
Abstract
Context
Animal movement through the landscape is essential to several ecological processes, assuring genetic flow, reproductive success and population dynamics. In fragmented landscapes, species change their movement patterns according to their sensitivity to landscape cover and configuration. Increased landscape habitat loss and fragmentation affects resource distribution and habitat availability, compelling species to move more frequently through anthropized matrices.
Objectives
Our aim was to provide a new prospect for what is known about animal movement through the landscape anthropogenic matrix in the Atlantic Forest biodiversity hotspot, by analyzing the potential movement distances of several mammal, bird, reptile, and amphibian species.
Methods
We used recorded roadkills within the Brazilian Atlantic Forest as a source of information of species’ occurrence. We assumed the euclidean distance from the roadkill spot to the nearest forest fragment as the potential movement distance by different species through the anthropogenic matrix. Experts’ classification of species’ forest dependence degree, and animal movement information from the literature survey was used to analyze the variation in potential animal movement distance in these matrices.
Results
We analyzed 90 species, the majority of them being non-forest dependent (n = 64; 71%). The potential movement distances ranged from 0 to > 1500 m, and forest-dependent species presented lower mean distances (between 772 and 978 m) than non-forest dependent species (between 673 and 1015 m). The literature survey resulted in 44 articles regarding animal movement in the matrix, referred only to 22 species. No articles were found for 20 species, mainly reptiles and amphibians.
Conclusions
By comparing these values to the average distance between forest remnants in the Atlantic Forest, we expect that the distance to be traveled through the matrix to reach forest fragments may be restrictive, mainly for species with higher degree of forest dependence. Despite being descriptive, our results provide important insights on animal movement through the matrix, especially for less studied species, such as amphibians and reptiles.
Similar content being viewed by others

Introduction
The landscape, composed of elements of intrinsic dynamics, is prone to continuous changes resulting from interactions between natural and anthropic processes (Antrop 1998). Among these processes, fragmentation and habitat loss are two relevant landscape modifiers that can be even more influential in places with intense human activities (MacDonald et al. 2018). Fragmentation per se comprises the increase in the number of habitat fragments, which causes disruption in spatial continuity and changes in configuration (Fahrig 2003). Landscape configuration influences the parameters used by animals in their movement decisions, since the navigation capacity depends on the organism’s response to the dynamics and spatiotemporal structure of the environment (Nathan et al. 2008). Facing changes in resource distribution and landscape configuration, species can alter their movement patterns in order to adapt their ecological needs in an anthropized environment (Fahrig 2007). Changes in movement patterns in the modified landscape often occur in a non-optimal way, resulting in a use of the landscape that does not meet the needs of the species (Fahrig 2007). Although it affects the optimal movement, the anthropogenic matrix areas (i.e. pasture, agriculture, forestry, and urban areas; hereafter matrix) can be an important refugee for mammals and hold a significant richness of these animals in modified landscapes (Magioli et al. 2018), also acting as an additional resource area for some species (Magioli et al. 2019). In addition to the composition of the landscape being an influencing factor in the movement of species between habitat fragments (Prevedello et al. 2010), the degree of habitat dependence also influences animal movement through the matrix (e.g. non-habitat area). Thus, forest-dependent animals in highly fragmented landscapes will be confined in small fragments, at risk of reducing their population and, possibly, their persistence in the landscape (Fahrig 2003). On the other hand, more generalist species (species less dependent on forest) will take more chances crossing and exploring the matrix (Fahrig 2003). This ability, however, increases their exposure to risks, possibly resulting in higher mortality rates (Fahrig 2002).
Linear infrastructure (e.g. roads, powerlines, gas lines, railroads) constitutes a large part of anthropic changes in the landscape and remains in continuous expansion, with road length expected to increase more than 3 million km by 2050, mainly in developing countries (Forman et al. 2002; Meijer et al 2018). Roads play an important role in fragmentation, as they facilitate human access, promoting occupation and exploitation, also dividing animal populations and reducing habitat quality (Ewers et al. 2006; Di Marco et al. 2019). In the context of fragmented landscapes, road infrastructure is very relevant, as it can modify the movement pattern of organisms and pose risk of roadkills, thus causing population declines and even local and regional species extinctions (Freitas et al. 2010). Thus, for forest-dependent species that are sensitive to habitat loss, roads act as barriers to movement, preventing access to resources, while for habitat generalist species roads can enhance foraging activity and facilitate movement, increasing their area of use in the landscape (Trombulak and Frissell 2000; Chen and Koprowski 2016). Although roads provide benefits for species that are not affected by traffic and that manage to avoid collisions (Rytwinski and Fahrig 2013), organisms with low mobility capacity or that remain immobile when sighting a vehicle (‘freezing behavior’) can suffer significant drops in their populations, as the probability of being killed by vehicle collision is higher (Rytwinski and Fahrig 2013).
In Brazil, most roads were built in the industrialized portion of the territory along the east coast—the Atlantic Forest Biome—concentrated in the southeast and south regions (IBGE 2014). Despite the high density of roads in the Atlantic Forest, few studies assess their effects on this biome (Freitas et al. 2010). Previous mapping indicated the existence of about 245,000 fragments making up the Atlantic Forest (Ribeiro et al. 2009), but recent studies revealed that the biome is covered by 28% of native vegetation, including both forest and non-forest formations, which comprises more than the double of the former estimates (Rezende et al. 2018). Still, the Atlantic Forest remains highly fragmented, turning the movement of fauna through the matrix to reach other forest remnants critical for the maintenance of population dynamics (Fahrig and Paloheimo 1988). Animal locomotion occurs according to functional connectivity, determined by the behavioral responses of organisms to the landscape structure and its movement patterns (Goodwin 2003). The matrix composition characterizes different degrees of permeability, and the more similar it is to the habitat area, the greater its quality, which increases the connectivity between fragments (Prevedello and Vieira 2010a). Hence, the same landscape can be interpreted in different ways by each species moving through it, and cannot be simply classified as connected or fragmented based on a single criterion (Forero-Medina and Vieira 2007).
To understand how movement patterns of organisms are shaped, given their perceptions of changes in the landscape, there are studies that analysed the use of space by different species in the Atlantic Forest. These studies are extremely important for understanding not only the animal movement per se, but also how space and resources are used by them and how interactions with other organisms occur (Cagnacci et al. 2010). According to Doherty et al. (2019), larger species, when in fragmented environments, expand their movements and tend to cross anthropogenic matrices to access different fragments, as small areas of habitat are not able to maintain sufficient resources to sustain the populations of these animals. The same is considered for habitat generalist species or those less sensitive to anthropogenic changes (Lees and Peres 2009). This is exemplified by species such as the maned wolf (Chrysocyon brachyurus), the cougar (Puma concolor) and the ocelot (Leopardus pardalis), which, within the Atlantic Forest, use fragments of native vegetation and non-habitat areas for foraging and dispersal, covering the non-habitat area as long as resources are available, which enable the maintenance of their population dynamics (Penteado 2012; Vynne et al. 2014). For small mammals and some reptiles, research indicates that the height of the vegetation that makes up the anthropic matrix and the arrangement in rows of cultivation areas placed between fragments have a great influence on movement, as they reduce the time spent in the matrix by providing better orientation and protection for these animals (Prevedello and Vieira 2010b; Kay et al. 2016). Regarding birds, some species find it extremely easy to travel through the matrix, due to their locomotion capacity, which allows them to fly for kilometers in regions outside their habitat (Lees and Peres 2009), as occurs with the white-shouldered fire-eye (Pyriglena leucoptera), capable of traversing open areas between fragments, and the blue manakin (Chiroxiphia caudata), as demonstrated by Uezu et al. (2005).
Despite these studies, there are still gaps in the knowledge regarding animal movement in anthropogenic matrices, especially amphibians and reptiles, which prevent us from better understanding their population dynamics and how they are affected by landscape modification. In this scenario, recorded roadkills can provide important information about these animals, such as their occurrence and movement. Thus, the use of roadkill data to infer the movement of organisms can help to elucidate distances traveled by them, giving insights on whether a given species moves a lot or a little in a context of fragmented habitat. Analyzing the potential movement distances in the matrix traveled by those species present in forest fragments can indicate how they are using the landscape and how far they can go. Although there has been advances in understanding how some species move in the non-habitat area, by adapting their movement to be faster (Crone et al. 2019) or by increasing or decreasing their movement distances (Doherty et al. 2021), there are still knowledge gaps about how far organisms can travel through the matrix. According to Forero-Medina and Vieira (2007), the distance traveled by different species in the matrix, especially forest-dependent ones, is thus far unclear. Even when considering habitat generalist species, that are able to use resources both in forest remnants and in the matrix, little is known about their movement in the non-habitat area, which makes the investigation of species' movement capacity relevant to understand other ecological aspects. The goal of this study is to provide a new prospect for what is known about animal movement through the landscape matrix in the Brazilian Atlantic Forest, by analyzing the potential movement distances of several mammal, bird, reptile, and amphibian species. We estimated the potential movement distance by calculating the Euclidean distance between the location of a species’ roadkill record to the nearest forest fragment. We used expert knowledge to assess species’ degree of forest dependence, and presented an overview of existing movement information in the scientific literature to reflect on the distribution of the potential movement distances of each species.
Material and methods
In the following subtopics we describe the extent and characterization of the study area, followed by the data sources and estimation of species potential movement. Then we present how the expert consultation and literature survey were conducted. The evaluation of the potential movement distance values was performed according to the expert’s classification for the forest dependence degree of each species, and any literature information about what is known about each species’ movement.
Study area and occurrence of species
The Atlantic Forest, composed of two main types of vegetation, namely the semideciduous seasonal forest and the dense ombrophilous forest (Morellato and Haddad 2000), originally occupied about 150 million hectares along the Brazilian coast. Due to its range, the biome varies in altitude (from sea level to more than 2000 m; IBGE 2012) -, annual rainfall (from 1000 mm in inland forests to 4000 mm in coastal areas; IBGE 2012), and latitude. The Atlantic Forest extension also encompasses on different transitional zones with neighboring biomes, such as Cerrado, Pampa, and Caatinga (IBGE 2012). These characteristics combined result in a wide range of environmental variability, which has led to the establishment of a highly diverse fauna and flora (Silva and Casteleti 2003).
The Atlantic Forest has been reduced over time, and in the first decades of 2000’s it accounted for about 16–26% coverage of remaining native vegetation distributed in dispersed forest fragments (depending on the quantification method, see Ribeiro et al 2009 and Rezende et al. 2018). Between 2012 and 2017, the road network increased more than 29,000 km in Brazil, due to economic and social growth (CNT 2021), leading to more impacts on wildlife. Currently, more than 152 thousand kilometers of roads run through the biome, 49.2% of which are paved (IBGE 2010). However, the biome still has one of the highest rates of biodiversity and endemism in the world, which classifies it as one of the 25 existing biodiversity hotspots (Tabarelli et al. 2005).
In this research, we used the integrative limit (the wider) of the delimitation of the Atlantic Forest biome (Muylaert et al. 2018), which comprises an area of more than 1.6 million km2. We also used a raster file of distances to the nearest forest remnant produced by LEEC Lab. The layer was generated based on maps of the World Wildlife Fund (Olson et al. 2001), MMA-IBGE (2004), Atlantic Forest Law (Brasil 2006), and Ribeiro et. al (2009). This raster file has a spatial resolution of 30 m and each pixel of the geographic space has the value of the Euclidean distance to the nearest forest remnant. Forest pixels have zero values, and the farther a pixel is from the nearest forest, the higher its value in meters.
We used the Brazil RoadKill Dataset (Grilo et al. 2018) with the location records of roadkilled fauna species. The database consists of a compilation of roadkill points present in published research, thesis, dissertations, and unpublished records of wildlife-vehicle collisions in Brazil. It is intended to assist the analysis of spatial and temporal patterns of wildlife roadkill, the influence of the landscape on these occurrences and the identification of species vulnerable to roads (Grilo et al. 2018). The database has 21,512 roadkill records, ranging from 1988 to 2017, with 83% of the roadkilled animals being identified to the species level. Each point contained in the database has details about the geographic coordinate of the roadkill, the date of registration, the classification in the IUCN red list and the source of the information. Among the records, there are 31 species of amphibians, 229 species of birds, 90 species of reptiles, and 99 species of mammals. These numbers refer to 2,87% (31 out of 1080), 11,93% (229 out of 1919), 11,64% (90 out of 773), and 14,12% (99 out of 701), respectively, when compared to the number of species found in Brazil (ICMBIO 2018).
First, we selected the records within the integrating limit of the Atlantic Forest, totalizing 14,465 records, 6966 of which are mammals, 3141 reptiles, 2601 birds, and 1757 amphibians. Then, we filtered the roadkill points identified to the level of species and/or genus that occurred from the year 2000–2017 (when the database was published). Based on this selection, we kept the species or genus with 15 or more roadkill occurrences.
Evaluation of potential movement distances
The potential distance traveled by each animal through the matrix (hereafter potential movement distance) was estimated as a straight line (i.e. Euclidean distance) from its roadkill point to the nearest forest fragment. From these values, the mean distance traveled for every species was calculated. These values were extracted from the distance raster, after the cartographic reprojection of the vector layer (shapefile) containing the roadkill records to the same geographic coordinate system as the distance raster (SAD 69). Then, for each animal class, the distribution of the potential movement distances of each species was represented in a dotplot along with the mean and median, using the ggplot2 package in software R version 4.1.0. All analyzes were performed using Quantum Gis Desktop software version 3.4.5 and R × 64 version 3.6.1.
Literature survey
For comparative purposes, we made a review on the existing scientific literature about the movement in the anthropic matrix for each of the studied species. The survey was carried out on the Web of Science (WoS) platform, using the keywords: “xx” movement OR “xx” dispersion OR “xx” “step lenght” AND “xx” distance NOT “seed dispersion”, covering the years from 2005 to 2020, with “xx” replaced by the scientific name of each focal species. The starting year of the search interval was chosen because it was in the last 2 decades that animal tracking studies started to rely on methods such as GPS (Global Positioning System), with the advance of telemetry technologies (Cagnacci et al 2010). All abstracts from selected articles were scanned, and the ones that measured species’ movement distances were fully read. Articles that made available animal movement distance values were selected, with no restriction for the location where they were carried out nor to the type of distance measure (e.g., average movement distance, full movement distance, daily movement distance). We compiled information such as the study coordinates, country, ecosystem or biome, the movement distance values (including the characterization of the movement type, e.g., dispersion, daily movement), and the research method (e.g., GPS colar, radiotelemetry). The full list of selected articles is available in the Supplementary Information.
Expert knowledge
When information is unavailable or scarce, expert knowledge portrays an essential role as a source of information (McBride and Burgman 2012). According to their training, experience or technical practices, experts can be qualified to correctly apply their knowledge to new contexts, corroborate existing information or act as a benchmark to unknown parameters (McBride and Burgmann 2012). Since the degree of forest dependence for many species is not available, a consultation with experts was carried out to define the most appropriate classification of the studied species according to their degree of habitat specialization. We contacted full professors and PhD candidates, with peer recognition and more than 5 years of work experience, to classify the selected species. The number of experts consulted varied between animal classes, with consultation of only one expert for mammals, amphibians, and reptiles, and two experts for birds. For this classification, the experts defined a value from 0 to 5 to categorize the degree of forest dependence of each species, where 0 referred to no forest dependence (habitat generalist species) and 5 to a high forest dependence (habitat specialist species). Species assigned with values between 5 and 4 were considered as forest-dependent, being the value 5 referred to strictly forest-dependent species. The species given the values between 0 and 3 were considered non-forest dependent (habitat generalist), and able to use the matrix area. Although being considered non-forest dependent, species assigned with values 2 and 3 were related to a higher degree of association with forest remnants, when compared to the other ones. Species with unknown forest dependence degree (no value assigned by expert), were assigned with N/A. Species for which two experts were consulted had their degree of forest dependence determined by the average of the values, resulting in a single value.
Results
Study area and occurrence of species
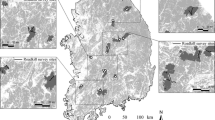
After the roadkill data screening, 14,462 records were obtained within the integrative limit of the Atlantic Forest, 1757 (12.2%) of them referring to amphibians, 2601 (18%) to birds, 3141 (21.7%) to reptiles, and 6963 (48.1%) to mammals. The selection of the species with more than 15 roadkill records led to a total of 90 species for analysis (summing up to 12,237 records in total; Fig. 1). The group with the highest number of species was mammals (n = 34), followed by birds (n = 26), reptiles (n = 22), and amphibians (n = 8) (Table 1).

The integrative limit of the Atlantic Forest and the locations of the roadkill records referring to the 90 species analyzed, filtered from the Brazil RoadKill Dataset (Grilo et al. 2018). Dark green represents forest remnants
Evaluation of potential movement distances
The potential movement distances ranged from 0 to 1500 m, in many cases with values distributed along the whole range of values for species in every animal group. For some species, mainly the ones with higher roadkill records, an aggregation of values is observed, especially in the extremes of the distance range (lower than 500 m and higher than 1000 m). Despite the amount of roadkill records, a great variability of potential movement distances is seen in the four animal groups, even for amphibians, whose species had the lowest number of records and still had their potential movement distance values distributed along all the distance range. The dotplots generated for each animal group contain the distribution of the potential movement distance values for all species, which enable the examination of every potential movement distance obtained despite the mean and median values for species of mammals (Fig. 2), birds (Fig. 3), reptiles (Fig. 4), and amphibians (Fig. 5).

Dot plot with potential movement distances generated for mammal species within the Brazilian Atlantic Forest biodiversity hotspot, representing species mean (red) and median (black) potential movements

Dot plot with potential movement distances generated for bird species within the Brazilian Atlantic Forest biodiversity hotspot, representing species mean (red) and median (black) potential movements

Dot plot with potential movement distances generated for reptile species within the Brazilian Atlantic Forest biodiversity hotspot, representing species mean (red) and median (black) potential movements

Dot plot with potential movement distances generated for amphibian species, representing species’ mean (red) and median (black) potential movements
Regarding the mean and median of the potential movement distance values, reptiles consisted of the group with the highest mean values, which varied between 800 and 1000 m, whereas the median values for 21 of its 22 species are above 1000 m, showing that there is a concentration of values of potential movement distance higher than 1000 m for the group (Fig. 4). This pattern is also seen for mammals, whose mean potential movement distance for the majority of the species remained between 800 and 920 m, with median values above 1000 m for 32 of the 34 species of the group (Fig. 2), and birds, whose mean potential movement distance values varied between 631 and 1058 m, and the median values for 22 of 26 species were higher than 1000 m (Fig. 3). Amphibians were the group with the lowest mean potential movement distances, ranging from 738 to 886 m, with three of the species’ median values lower than the mean value, and the remaining five species with median values higher than the mean values.
In general, the mean and median potential movement distance values didn’t have a great difference among the species studied, having a consistent pattern between the animal groups. Only four of the 90 species analyzed obtained mean potential movement distances superior to 1000 m: Turdus rufiventris (1058 m), Nyctidromus albicollis (1044 m), Chrysocyon brachyurus (1015 m), and Boa constrictor (1026 m), the first one being the holder of the highest mean value among all of the species considered.
Literature survey
The literature review resulted in 477 articles, from which only 46 contained the required information of species’ movement, referring to 22 species. The number of studies for each species were discrepant, given that 37.5% of them referred to Puma concolor (n = 11) and Athene cunicularia (n = 7), whereas for the amphibian and reptile species there was only one study. For 35 out of the total 90 species analyzed in this study no specific study regarding movement was found (mostly reptiles). In addition, there were studies found for 33 species, but none of them contained the necessary information regarding movement distances. The analyzed literature presented a great variability of methods, which provided different measures of movement, such as daily movement, movement speed, dispersion in a long period of time, minimum and maximum movement distance, and short movement distances. Besides, the studies considered different parameters (e.g. different periods of the day, reproductive season, movement within and between forest fragments, and in urban areas, etc.) which made it difficult to compile the results (Table S1).
Expert knowledge
Mammals
Of the 34 mammal species, only three had no assigned values for their degree of forest specialization (Table 2)—A. lituratus, H. brasiliensis, and L. geoffroyi. The species with highest forest dependence were A. guariba, C. insidiosus, C. spinosus, D. aurita, L. guttulus, L. wiedii, and T. tetradactyla, all of them designated with value 5, being, thus, considered forest specialists. The species given value 4 of forest dependence were D. novemcinctus, D. albiventris, E. barbara, G. cuja, L. pardalis, L. tigrinus, N. nasua, and P. cancrivorus.
Birds
None of the bird species were classified as forest-dependent, the majority of them being assigned with value 1 of dependence (Table 3). T. rufiventris was the species with the higher degree of habitat dependence (2.5), followed by A. clamator, C. ruficapillus, C. picui, N. albicollis, P. coronata, T. sayaca,and T. aedon, whose degree was 2. The remaining species’ forest dependence degree varied between 0, 0.5, 1, and 1.5. Even though some species have some preference for arboreal vegetation, none of them is considered strictly forest specialists, because they occur in different configurations of habitats, with many of them occurring in open and urbanized areas.
Reptiles
Of the 22 reptile species, six had no assigned values, due to the expert’s lack of acquaintance with those species. Four species–H. tectifera, P. olfersii, S. merianae and D. neuwiedi–were classified as value 3 for forest dependence, which was the highest value among the reptiles (Table 4). Two species were defined as habitat generalists, B. alternatus, and O. rhombifer. All of the other species had their values assigned between 2 and 1, which means little forest dependence.
Amphibians
According to the classification, the species with the highest forest dependence were R. icterica (3), followed by H. faber (2.5), and L. latrans (1). The other species were considered as not dependent on forest habitats (Table 5). The only species without data was B. guentheri.
Discussion
The obtained data, substantially of a descriptive character, provide an initial basis to inform the potential movement capacity of species from different groups through the anthropogenic matrix, within the Atlantic Forest biodiversity hotspot, which is unknown for many of them. Despite some subjectivity or bias that can be associated to the use of expert knowledge, it can still serve as a benchmark or reference values, since empirical values for the degree of forest dependence of the studied species are not available. Expert opinion has been widely used to quantify resistance to movement based on landscape features (Zeller et al 2012), and to support different types of decision making, such as development or evaluation of projects, study designs, planning, interpretation of results, and improvement of model parameters (Martin et al. 2012). In this study, it was used as a guideline for the identified patterns of potential movement distance. Therefore, the degree of forest dependence, even though it can not define the locomotion capacity of the species, is capable of indicating the resistance to the use of anthropized environments, which, in turn, can inhibit long-distance movements that cross non-forest areas. Consequently, in the context of the Atlantic Forest biome, which is highly fragmented, we expect that species with greater forest dependence move shorter distances outside the fragment, due to their sensitivity to modified areas of the landscape. As to species like C. brachyurus, T. rufiventris, A. lituratus, C. thous, and H. yagouaroundi, which are considered non-forest dependent species or species associated with open environments, there are studies that demonstrate continuous use of forest fragments or forest edges in different areas of the Atlantic Forest, besides being pointed out a higher occurrence of P. concolor in areas with greater forest cover in the biome (Michalski et al 2006; Trevelin et al. 2013; Da Silveira et al. 2016; Regolin et al. 2017; Eigenheer 2018). Thus, even with non-forest dependent species analyzed in this study, we can assume that such species maintain a certain degree of association with forest remnants, supporting the potential movement distances found here.
The 15 forest-dependent species (with designated values from 4 to 5), all belonging to the mammal group, had mean potential movement distances between 772 m (A. guariba) and 978 m (L. triginus). Despite being close to 1 km, the potential movement distance for these species may not mean that they can move effectively to another fragment within the Atlantic Forest, since the average distance between the biome's forest remnants—including small fragments (< 50 ha)—is about 1400 m, which represents a large matrix area to be crossed (Ribeiro et al. 2009). Thus, strictly forest species (designated value 5), less tolerant to habitat modification, avoid the use of open areas and forest edges, and face the risk of becoming isolated in the fragments they inhabit, affecting their population dynamics and gene flow (Fahrig and Paloheimo 1988). On the other hand, the more tolerant forest species (designated value 4) may risk moving outside the fragments, but without the guarantee that they will reach new fragments. According to the maximum potential movement distance values, 9 of the 15 species have rates above 1400 m, which may indicate a certain degree of success for some individuals who try to disperse across the matrix, but that are not necessarily able to accomplish the demand of reaching another suitable fragment. In this context, small fragments are of great importance when acting as stepping stones (Ribeiro et al. 2009), but, according to the results found here, mitigation measures are necessary to expand the functional connectivity for the movement of such species.
Among the non-forest dependents (with assigned values between 0 and 3), represented by the four animal groups, there are species with different physical and ecological characteristics that maintain different relationships with the forest remnants of the Atlantic Forest. For instance, aquatic and semi-aquatic species depend on the presence of water bodies at different stages of their life, as is the case of mammals such as H. hydrochaeris, E. barbara, L. longicaudis, and M. coypus, reptiles such as A. spixii, H. tectifera, P. hilarii, T. dorbigni, and E. jaegeri, and most amphibian species, that depend on the presence of water to develop. The movement of these animals is often related to the presence of water bodies in the landscape, which, in degraded areas, are commonly related to forest remnants, being this proximity associated with the preservation of water provision (Molin et al. 2017). For the chelonian species considered here (T. dorbigni, P. hilarii, H. tectifera), for example, there are records of preference for less modified environments, which does not, however, prevent the use of water bodies in anthropic places (Horta 2008; Lopez et al. 2013). Thus, some aquatic and semi-aquatic generalist species in this study possibly manage to cross areas of non-habitat during locomotion, and some of them occur in close proximity to urban centers, as they are more tolerant to anthropization (e.g., capybara (H. hydrochaeris), argentine snake-necked turtle (H. tectifera)). In the present study, the mean potential movement distance values for these animals are relatively wide, reaching 1000 m for mammals and ranging from 673 (B. guentheri) to 972 m (H. tectifera), which indicates that movement may be occurring in broad extensions through the landscape, due to their lower habitat quality requirements, allowing them to use different conformations of the environment. Even so, locomotion in the matrix area still poses threats to such species, which can be subject to predation and roadkill, for example.
Similarly to non-forest dependent aquatic and semi-aquatic species, non-forest dependent species (assigned values between 0 and 3) are also associated, in the context of the Atlantic Forest biome, with forest remnants. The mammal species with the greatest mean potential movement distance value (1015 m) was the maned wolf (C. brachyurus) which, when in transitional environments between Cerrado and the Atlantic Forest or in the Atlantic Forest itself, has records of remaining close to forest remnants, demonstrating a certain degree of association with these environments (Eigenheer 2018). Being a species that inhabits open vegetation areas, mainly in the Cerrado, and having great locomotion capacity, the maned wolf is able to move through open areas of the Atlantic Forest with ease, crossing large areas and reaching different configurations of the landscape. The same is observed for the only bat species analyzed, A. lituratus, whose mean potential movement distance was greater than for other mammal species. Despite being present in restored areas, they are able to widely use the different features of the landscape (Trevelin et al. 2013), due to their size and adaptation to urban and anthropic environments (Bredt and Uieda 1996).
Non-forest dependent species of reptiles and birds (assigned values between 0 and 3) are also associated with forest edges, whether for predation, nesting or protection against predators. These associations were reported for snake species like P. olfersii, B. constrictor, and P. patagoniensis (Marques et al. 2017), rattlesnakes (Carfagno and Weatherhead 2006), and birds like T. rufiventris (Gasperin and Pizo 2009; Da Silveira et al. 2016), and T. alba (Foster and Johnson 1974). For these species, that are capable of using non-forest areas, the small forest fragments act as important stepping stones, which help them cross matrix areas (Fischer and Lindenmayer 2002), and significantly reduce the functional distance between the fragments, allowing them to reach larger remnants where better maintenance of population dynamics is possible (Ribeiro et al. 2009). The results found for the potential movement distance values of such animals, whose mean values vary between 631 m (C. campestris) and 1058 m (T. rufiventris)—56.25% of them being above 900 m –, and the maximum reaches 3774 m (Z. capensis), imply the ability of these species to travel through different landscape formations, demonstrating a wide ability to move through the matrix. When analyzing all the potential movement distances for the non-forest dependent species (amphibians, birds, mammals and reptiles with values between 0 and 3), there is an aggregation of values above 1000 m, which suggests large displacement in non-forest areas, enabling their presence in more altered regions of the landscape and in more distant forest fragments, contributing for the maintenance of their population dynamics.
When comparing our potential movement distance with the movement values found in the literature, we found great discrepancy, with much larger or much smaller distance values in the literature than those presented here. The greatest similarity between those values were from the articles that had the Atlantic Forest as their study area. For instance, C. chinga (Kasper et al. 2012), N. nasua (Hirsch et. al 2013), and T. dorbigni (Bager et al. 2012), presented similar minimum and maximum movement values. Complementary, one article for A. cunicularia, carried out in the USA (Catlin and Rosenberg 2014), also showed similar minimum and maximum movement values. Despite the divergent values found for most of the other species, this literature survey helped to define the state of knowledge regarding the movement in the matrix of the animals analyzed here, including the identification of meaningful knowledge gaps. The various measures used in the compiled movement data, along with the different parameters considered, makes it challenging to identify patterns in the available information. Having similar standardized approaches in research about the movement of animals is a crucial step to further understand how animals’ movement capacity is affected in anthropized areas.
The lack of studies focusing on animal movement in the anthropogenic matrix—especially for amphibians and reptiles but also for the other groups–, shows that there is still room for research in this field, with a great potential of enlightening aspects of species movement capacity through non-habitat areas. This type of knowledge can guide strategies to enhance connectivity by considering species with limited movement capacity through the anthropogenic matrix; in this study, those with potential movement distance values smaller than the average distance between the forest remnants of the Atlantic Forest. Regarding the importance of the Atlantic Forest and the species that inhabit this biodiversity hotspot, it can be considered that, even being descriptive and having limitations, the results we presented have great potential to fill gaps in knowledge about animal mobility, also pointed out by the experts themselves. Although being a simple way to infer animal movement, the application of roadkill data along the assumptions considered in this study—such as that all species, in the context of the biome, have some kind of association with forest remnants, and that their movement occured in a straight line between the roadkill to the nearest fragment—were able to bring additional understanding of how much the animals are capable to move outside forest patches. The results provided by our study can guide future research, by giving a starting point for movement distances that different species are potentially able to move in non-habitat areas. Therefore, when researchers establish new agenda that will focus on species movement patterns, they can use the information summarized here to define their movement data sampling strategy, taking into account how data will be recorded in a spatio-temporal perspective. The experts’ classification on forest dependence degree can also be used by other researchers, especially if they aim to study species’ relation with forest remnants and other landscape features. Lastly, the literature compilation grants directly what is known for species’ movement distance values through the matrix, not only in the Atlantic Forest, but for different biomes worldwide. The presentation of the results generated insights about the movement of species in the modified landscapes of the Brazilian Atlantic Forest, by linking expert knowledge with roadkill data. The results provide a general overview of how much these species can move through the modified landscape, outside forest remnants, mainly for forest-dependent species, with a certain degree of forest dependence, and for species for which movement is poorly studied. Even for non-forest dependent species, the distance values from the roadkill spot to the nearest forest fragment can also contribute to a better understanding of how these species use the space in a biome with high levels of habitat loss and fragmentation.
Data availability
The dataset used in this study is publicly available at https://zenodo.org/record/1420508#.YnTlfdpByUk.
References
Antrop M (1998) Landscape change: plan or chaos? Landsc Urban Plan 41:155–161
Bager A, Krause L, De Freitas TR (2012) Fidelity to nesting sites and orientation of Trachemys dorbigni (Duméril & Bibron, 1835) (Testudines: Emydidae) female in southern Brazil. Trop Zool. https://doi.org/10.1080/03946975.2012.679393
Brasil (2006) Lei Federal nº 11.428. Definições, objetivos e príncipios do regime jurídico do Bioma Mata Atlântica. Brasília, DF
Bredt A, Uieda W (1996) Bats from urban and rural environments of the Distrito Federal, Mid-western Brazil. Chiroptera Neotropical 2:54–55
Cagnacci F, Boitani L, Powell RA, Boyce MS (2010) Animal ecology meets GPS-based radiotelemetry: a perfect storm of opportunities and challenges. Bioll Sci. https://doi.org/10.1098/rstb.2010.0107
Carfagno GLF, Weatherhead PJ (2006) Intraspecific and interspecific variation in use of forest-edge habitat by snakes. Can J Zool. https://doi.org/10.1139/Z06-124
Catlin DH, Rosenberg DK (2014) Association of sex, fledging date, and sibling relationships with post-fledging movements of burrowing owls in a nonmigratory population in the imperial valley California. J Raptor Res. https://doi.org/10.3356/JRR-12-24.1
Chen HL, Koprowski JL (2016) Differential effects of roads and traffic on space use and movements of native forest-dependent and introduced edge-tolerant species. PLoS ONE. https://doi.org/10.1371/journal.pone.0148121
CNT, Confederação Nacional do Transporte (2021) Anuário CNT do Transporte - Estatísticas consolidadas. Brasília, DF
Crone EE, Brown LM, Hodgson JA et al (2019) Faster movement in nonhabitat matrix promotes range shifts in heterogeneous landscapes. Ecology. https://doi.org/10.1002/ecy.2701
da Silveira NS, Niebuhr BBS, Muylaert RDL et al (2016) Effects of land cover on the movement of frugivorous birds in a heterogeneous landscape. PLoS ONE. https://doi.org/10.1371/journal.pone.0156688
di Marco M, Ferrier S, Harwood TD et al (2019) Wilderness areas halve the extinction risk of terrestrial biodiversity. Nature. https://doi.org/10.1038/s41586-019-1567-7
Doherty TS, Fist CN, Driscoll DA (2019) Animal movement varies with resource availability, landscape configuration and body size: a conceptual model and empirical example. Landsc Ecol. https://doi.org/10.1007/s10980-019-00795-x
Doherty TS, Hays GC, Driscoll DA (2021) Human disturbance causes widespread disruption of animal movement. Nat Ecol Evol. https://doi.org/10.1038/s41559-020-01380-1
Eigenheer MAA (2018) Animal movement, social interactions and mutualism in Brazilian ecosystems. Dissertation, Universidade Estadual Paulista.
Ewers RM, Kliskey AD, Walker S et al (2006) Past and future trajectories of forest loss in New Zealand. Biol Cons. https://doi.org/10.1016/j.biocon.2006.06.018
Fahrig L (2002) Effect of habitat fragmentation on the extinction threshold: a synthesis. Ecol Appl 12(2):346–353
Fahrig L (2003) Effect of habitat fragmentation on biodiversity. Annu Rev Ecol Evol Syst 34:487–515
Fahrig L (2007) Non-optimal animal movement in human-altered landscapes. Funct Ecol 21:1003–1015
Fahrig L, Paloheimo J (1988) Effect of spatial arrangement of habitat patches on local population size. Ecology 69:468–475
Fischer J, Lindenmayer DB (2002) The conservation value of paddock trees for birds in a variegated landscape in southern New South Wales 1. Species composition and site occupancy patterns. Biodivers Conserv 11:807–832
Forero-Medina G, Vieira MV (2007) Conectividade funcional e a importância da interação organismo-paisagem. Oecologia Brasiliensis 11:493–502
Forman RTT et al (2002) Road Ecology: Science and Solutions. Island Press, Washington
Foster MS, Johnson NK (1974) Notes on birds of Costa Rica. Wilson Bulletin 86:59–63
Freitas SR, Hawbaker TJ, Metzger JP (2010) Effects of roads, topography, and land use on forest cover dynamics in the Brazilian Atlantic Forest. Ecol Manag. https://doi.org/10.1016/j.foreco.2009.10.036
Gasperin G, Pizo MA (2009) Frugivory and habitat use by thrushes (Turdus spp.) in a suburban area in south Brazil. Urban Ecosyst. https://doi.org/10.1007/s11252-009-0090-2
Goodwin BJ (2003) Is landscape connectivity a dependent or independent variable? Landsc Ecol 18:687–699
Grilo C, Coimbra MR, Cerqueira RC et al (2018) Brazil road-kill: a data set of wildlife terrestrial vertebrate road-kills. Ecology. https://doi.org/10.1002/ecy.2464/suppinfo
Hirsch BT, Tujague MP, Di Blanco YE, Di Bitetti MS, Janson CH (2013) Comparing capuchins and coatis: causes and consequences of differing movement ecology in two sympatric mammals. Anim Behav. https://doi.org/10.1016/j.anbehav.2013.05.023
Horta GDF (2008) Movimentação e uso do espaço por Acanthochelys spixii (Testudines, Chelidae) no Parque Nacional de Brasília, Distrito Federal. Dissertation, Universidade de Brasília
IBGE. Instituto Brasileiro de Geografia e Estatística (2010) Base vetorial contínua da carta internacional do mundo ao milionésimo (BCIM).
IBGE (2012) Manual Técnico da Vegetação Brasileira. Instituto Brasileiro de Geografa e Estatistica,
IBGE. Instituto Brasileiro de Geografia e Estatística (2014) Nota técnica da logística dos transportes no Brasil. Coordenação de Geografia.
ICMBIO. Instituto Chico Mendes de Conservação da Biodiversidade (2018) Livro Vermelho da Fauna Brasileira Ameaçada de Extinção. Volume I. 492p. Brasília, DF: ICMBIO/MMA.
Kasper CB, Soares JB, Freitas TR (2012) Differential patterns of home-range, net displacement and resting sites use of Conepatus chinga in southern Brazil. Mamm Biol. https://doi.org/10.1016/j.mambio.2012.03.006
Kay GM, Driscoll DA, Lindenmayer DB et al (2016) Pasture height and crop direction influence reptile movement in an agricultural matrix. Agr Ecosyst Environ. https://doi.org/10.1016/j.agee.2016.10.019
Lees AC, Peres CA (2009) Gap-crossing movements predict species occupancy in Amazonian forest fragments. Oikos. https://doi.org/10.1111/j.1600-0706.2008.16842.x
López MS, Sione W, Leynaud GC et al (2013) How far from water? Terrestrial dispersal and nesting sites of the freshwater turtle Phrynops hilarii in the floodplain of the Paraná river (Argentina). Zool Sci. https://doi.org/10.2108/zsj.30.1063
MacDonald ZG, Anderson ID, Acorn JH, Nielsen SE (2018) Decoupling habitat fragmentation from habitat loss: butterfly species mobility obscures fragmentation effects in a naturally fragmented landscape of lake islands. Oecologia. https://doi.org/10.1007/s00442-017-4005-2
Magioli M, de Bovo AAA, Percequillo AR et al (2018) Human-modified landscape acts as refuge for mammals in Atlantic Forest. Biota Neotrop. https://doi.org/10.1590/1676-0611-BN-2017-0395
Magioli M, Moreira MZ, Fonseca RCB et al (2019) Human-modified landscapes alter mammal resource and habitat use and trophic structure. Proc Natl Acad Sci. https://doi.org/10.1073/pnas.1904384116
Marques R, Rödder D, Solé M, Santos Tinôco M (2017) Diversity and habitat use of snakes from the coastal Atlantic rainforest in northeastern Bahia, Brazil. Salamandra 53:34–43
Martin TG, Burgman MA, Fidler F et al (2012) Eliciting expert knowledge in conservation science. Conserv Biol. https://doi.org/10.1111/j.1523-1739.2011.01806.x
McBride MF, Burgman MA (2012) What is expert knowledge, how is such knowledge gathered, and how do we use it to address questions in Landscape Ecology? Exp Knowl Its Appl Landsc Ecol. https://doi.org/10.1007/978-1-4614-1034-8_2
Meijer JR, Huijbregts MAJ, Schotten KCGJ, Schipper AM (2018) Global patterns of current and future road infrastructure. Environ Res Lett. https://doi.org/10.1088/1748-9326/aabd42
Michalski F, Crawshaw PG, de Oliveira TG, Fabián ME (2006) Notes on home range and habitat use of three small carnivore species in a disturbed vegetation mosaic of Southeastern Brazil. Mammalia. https://doi.org/10.1515/MAMM.2006.004
MMA-IBGE: IBGE. Instituto Brasileiro de Geografia e Estatística (2004) Mapa de biomas e de vegetação. Ministério do Meio Ambiente.
Molin PG, Gergel SE, Soares-Filho BS, Ferraz SFB (2017) Spatial determinants of Atlantic Forest loss and recovery in Brazil. Landsc Ecol. https://doi.org/10.1007/s10980-017-0490-2
Morellato LPC, Haddad CFB (2000) Introduction: the Brazilian Atlantic Forest. Biotropica 32:786–792
Muylaert RL, Vancine MH, Bernardo R et al (2018) A note on the territorial limits of the Atlantic Forest. Oecologia Australis. https://doi.org/10.4257/oeco.2018.2203.09
Muylaert, R. L. et al (2018) Uma nota sobre os limites territoriais da Mata Atlântica. Repositório GitHub, https://github.com/LEEClab/ATLANTIC-limits.
Nathan R, Getz WM, Revilla E et al (2008) A movement ecology paradigm for unifying organismal movement research. Proc Natl Acad Sci. https://doi.org/10.1073/pnas.0800375105
Olson DM, Dinerstein E, Wikramanayake ED et al (2001) Terrestrial ecoregions of the world: a new map of life on Earth. Bioscience. https://doi.org/10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2
Penteado MJF (2012) Área de vida, padrões de deslocamento e seleção de habitat por: pumas (Puma concolor) e jaguatiricas (Leopardus pardalis), em paisagem fragmentada do Estado de São Paulo. Dissertation, Universidade Estadual de Campinas.
Prevedello JA, Vieira MV (2010a) Does the type of matrix matter? A quantitative review of the evidence. Biodivers Conserv 19:1205–1223
Prevedello JA, Vieira MV (2010b) Plantation rows as dispersal routes: a test with didelphid marsupials in the Atlantic Forest Brazil. Biol Conserv. https://doi.org/10.1016/j.biocon.2009.09.016
Prevedello JA, Forero-Medina G, Vieira MV (2010) Movement behaviour within and beyond perceptual ranges in three small mammals: effects of matrix type and body mass. J Anim Ecol. https://doi.org/10.1111/j.1365-2656.2010.01736.x
Regolin AL, Cherem JJ, Graipel ME et al (2017) Forest cover influences occurrence of mammalian carnivores within Brazilian Atlantic Forest. J Mammal. https://doi.org/10.1093/jmammal/gyx103
Rezende CL, Scarano FR, Assad ED et al (2018) From hotspot to hopespot: an opportunity for the Brazilian Atlantic Forest. Perspect Ecol Conserv. https://doi.org/10.1016/j.pecon.2018.10.002
Ribeiro MC, Metzger JP, Martensen AC et al (2009) The Brazilian Atlantic Forest: how much is left, and how is the remaining forest distributed? Implic Conserv Biol Conserv. https://doi.org/10.1016/j.biocon.2009.02.021
Rytwinski T, Fahrig L (2013) Why are some animal populations unaffected or positively affected by roads? Oecologia. https://doi.org/10.1007/s00442-013-2684-x
Silva JMC, Casteleti CH (2003) Status of the biodiversity of the Atlantic forest of Brazil. In: Galindo-Leal C, Câmara IG (eds) The Atlantic Forest of South America: biodiversity status, threats, and outlook. Island Press, Washington, DC, pp 43–59
Tabarelli M et al (2005) Desafios e oportunidades para a conservação da biodiversidade na Mata Atlântica brasileira. Megadiversidade 1:132–138
Trevelin LC, Silveira M, Port-Carvalho M et al (2013) Use of space by frugivorous bats (Chiroptera: Phyllostomidae) in a restored Atlantic forest fragment in Brazil. For Ecol Manag. https://doi.org/10.1016/j.foreco.2012.11.013
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic communities. Conserv Biol 14:18–30
Uezu A, Metzger JP, Vielliard JME (2005) Effects of structural and functional connectivity and patch size on the abundance of seven Atlantic Forest bird species. Biol Cons. https://doi.org/10.1016/j.biocon.2005.01.001
Vynne C, Booth RK, Wasser SK (2014) Physiological implications of landscape use by free-ranging maned wolves (Chrysocyon brachyurus) in Brazil. J Mammal. https://doi.org/10.1644/12-MAMM-A-247
Zeller KA, McGarigal K, Whiteley AR (2012) Estimating landscape resistance to movement: a review. Landsc Ecol. https://doi.org/10.1007/s10980-012-9737-0
Acknowledgements
We thank Marcelo Magioli, Célio F. B. Haddad, João C. Pena, Vinicius Tonetti, and Renato S. Bérnils for the fundamental knowledge shared in the expert consultation. We also thank the Spatial Ecology and Conservation lab (LEEC) team for helping with data handling and additional technical support provided, especially Milene Alves-Eigenheer and Maurício H. Vancine. ICMF was supported by a National Council for Scientific and Technological Development—CNPQ scholarship (PIBIC). MCR thanks to FAPESP (processes #2013/50421-2; #2020/01779-5; #2021/08534-0; #2021/10195-0; #2022/10760-1) and National Council for Scientific and Technological Development – CNPQ (processes #442147/2020-1; #402765/2021-4; #313016/2021-6; #440145/2022-8) for their financial support.
Funding
ICMF was supported by a National Council for Scientific and Technological Development—CNPQ scholarship (PIBIC). MCR thanks to FAPESP (processes #2013/50421–2; #2020/01779–5; #2021/08534–0; #2021/10195–0; #2022/10760–1) and National Council for Scientific and Technological Development – CNPQ (processes #442147/2020–1; #402765/2021–4; #313016/2021–6; #440145/2022–8) for their financial support.
Author information
Authors and Affiliations
Contributions
Conceptualization: IMFC, MCR, JCA; Methodology: IMFC, MCR, JCA; Formal analysis and investigation: IMFC; Writing – original draft preparation: IMFC, JCA; Writing – review and editing: MCR, JCA; Funding acquisition: IMFC, MCR; Supervision: MCR, JCA.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cassimiro, I.M.F., Ribeiro, M.C. & Assis, J.C. How did the animal come to cross the road? Drawing insights on animal movement from existing roadkill data and expert knowledge. Landsc Ecol 38, 2035–2051 (2023). https://doi.org/10.1007/s10980-023-01671-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-023-01671-5