
Abstract
Numerous permanent and temporary wetlands occur throughout the world’s drylands. Although characterised by diverse water inundation frequencies, durations and depths, these wetlands in drylands are typically hotspots of biological activity and productivity. The healthy functioning and even existence of many wetlands in drylands, however, is threatened by desiccation resulting from a combination of climate change and human disturbance. Near Alcañiz in arid northeast Spain, three adjacent saladas (playas) with contrasting hydroperiods provide an opportunity to investigate how moisture availability affects their soil carbon (C) stocks, CO2 efflux, and microbial communities. Predicting the impacts of changes in moisture availability on the C cycle in wetlands in drylands is challenging but important because many contain large C stocks and may be significant sources and sinks of greenhouse gases. Frequent inundation and/or near-permanent soil water saturation supports the generation of organic C from a range of different sources. Soil inorganic C was greatest on the driest salada (3.8%) compared to the wetter saladas (3.0% and 2.1%) owing to evaporative concentration and the reaction of CO2 with available Ca2+, Mg2+ and Na+ ions. CO2 efflux was greatest at intermediate moisture levels (142 mg CO2 m−2 h−1), but the spatial and temporal variability in CO2 efflux on salada surfaces is large, demonstrating the need for intensive sampling regimes to provide realistic estimates of their contribution to atmospheric CO2 exchanges. Different microbial community structures also characterise each salada. The saladas near Alcañiz, and many other similar features in northeast Spain, are renowned for their rare and threatened flora and fauna, yet their soil C cycle characteristics and soil microbial communities provide additional reasons to monitor the impacts of climate change and protect these vulnerable environments from further anthropogenic disturbances.
Similar content being viewed by others

Introduction
Throughout the world’s drylands, there are numerous permanent and temporary wetlands, such as floodplains, playas and oases (Tooth and McCarthy 2007). These wetlands in drylands are characterised by diverse hydroperiods (water inundation frequency, duration and depth) and are typically hotspots of biological activity and productivity compared to the surrounding landscape (Millennium Ecosystem Assessment 2005; Ezcurra 2006; Tooth et al. 2015). Nonetheless, the healthy functioning and possibly even existence of many wetlands in drylands is threatened by a combination of climate change and human disturbance that is projected to lead to changes in moisture availability (IPCC 2014a, b; Lin et al. 2015; Tooth 2018; Ramsar Convention 2018; Sandi et al. 2020; Xi et al. 2021). Changes in surface water and groundwater availability will be expressed through altered hydroperiods, which will impact on biological and chemical processes and characteristics, including those related to the carbon (C) cycle (Petrescu et al. 2015). Predicting the impacts of changes in moisture availability on the C cycle in wetlands in drylands is challenging but important because many contain large C stocks and may be significant sources and sinks of greenhouse gases (Nazaries et al. 2013).
Several distinctive characteristics of the C cycle in wetlands in drylands confound simple attempts to transfer data and concepts from better studied humid region wetlands (e.g. Mitsch and Gosselink 2015). First, in some wetlands in drylands, the majority of organic carbon (OC) may be generated by bacteria, algae, and aquatic organisms rather than by vascular plants (Domínguez‐Beisiegel et al. 2013a). Cyanobacterial photosynthesis has been shown, for example, to contribute OC to playa soils in the Makgadikgadi Basin in Botswana (Thomas et al. 2014). Although relatively little is known about controls on cyanobacterial photosynthesis in the soils of saline inland wetlands, or in other types of wetlands in drylands, periods of net CO2 uptake are likely to be short-lived and episodic (Ladrón de Guevara et al. 2014; Thomas et al. 2014) or seasonally controlled (Williams et al. 2014; Thomas et al. 2022). Second, wetlands in drylands are also sites of inorganic carbon (IC) production and storage (Xie et al. 2009; Yates et al. 2013). Globally, terrestrial IC formation occurs largely in dryland soils, and may be as much as 0.152 Pg C year−1, an amount similar to that buried in ocean sediments (Li et al. 2017). The generalised reaction that leads to IC formation involves CO32− ions, derived from CO2 dissolved in soil water, which combine with divalent metallic ions to form carbonates (Xie et al. 2009; Yates et al. 2013). Soil carbonate formation is of particular significance to the C cycle because carbonate is recalcitrant and less likely to be returned to the atmosphere as CO2 than is OC. Third, the extreme variations in moisture availability typical of many wetlands in drylands will have a profound effect on microbial communities, greenhouse gas emissions, and ultimately C stocks. Soil microbial activity, and consequently CO2 efflux, is typically reduced at high and low moisture extremes, and greatest at intermediate levels where water and oxygen availability are optimal (Orchard and Cook 1983; Schimel et al. 1999; Moyano et al. 2013).
Despite these previous findings regarding the distinctive aspects of the C cycle in wetlands in drylands, large gaps in knowledge remain, particularly for saline inland wetlands (e.g., playas, pans, salt lakes) where there are very few data on soil CO2 emissions, carbon stocks or microbial communities. Yet saline inland wetlands could be a significant source of CO2 and play a significant role in regional and global C cycles (Duarte et al. 2008). Whilst the IPCC Task Force on National Greenhouse Gas Inventories (IPCC 2014b) noted that saline inland wetlands are important features in dryland landscapes worldwide, little information was available to assess their role as sources or sinks of greenhouse gases, and consequently the Task Force was unable to provide guidance for these wetland types.
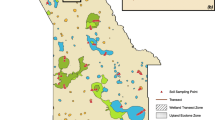
As a contribution towards this critical knowledge gap, we explore the contrasting hydroperiods of three adjacent playas, known locally as saladas, located near Alcañiz in northeast Spain (Fig. 1). The saladas are situated along a 3.5 km long hydrological gradient from wet to relatively dry and provide an opportunity to test hypotheses on how different moisture conditions affect critical aspects of the C cycle. For each salada, we characterised their hydroperiods, determined sediment chemistry (including stocks of OC and IC), calculated CO2 efflux, and investigated soil microbial communities. Our first hypothesis (H1) is that relatively wet soil moisture conditions will generate greater OC inputs than in drier soils, due to the controlling influence of moisture on biological productivity (Schimel et al. 1999). The net effect will be to elevate OC stocks in comparison to drier saladas. The effect of moisture on soil IC is less certain and our second hypothesis (H2) is that IC stocks will be dependent on the dominant carbonate formation processes (Xie et al. 2009; Yates et al. 2013). If evaporative concentration of solutes is the dominant process, then the drier saladas will have greater soil IC stocks than wetter saladas. If, however, IC formation is largely due to the supply of cations in inflowing waters and associated biomineralisation, then the wetter saladas will have higher soil IC stocks than drier saladas. Our third hypothesis (H3) is that very high soil moisture that is close to, or at saturated levels, will suppress microbial activity and CO2 efflux through a combination of O2 depletion and the impediment of gas transport (Moyano et al. 2013). Our final hypothesis (H4) is that each of the three saladas will contain different soil microbial communities due to their different inundation periods and soil moisture content. This is a phenomenon that has been observed in agricultural soils (e.g. Banerjee et al. 2016) but not previously in salada soils. The empirical data collected to test these hypotheses provide the basis for an improved conceptual understanding of the impact of climate or human-induced hydrological changes (e.g. transition from wetter to drier or, in rarer cases, drier to wetter) on the C cycle in the saladas. Ultimately, such knowledge may contribute to more effective management and protection of these important but threatened wetlands in drylands (c.f. Castañeda et al. 2013; Zhang et al. 2020).

a Outline of the three saladas near Alcañiz in northeast Spain. The black lines delineate the salada margins, and the red markers indicate the soil CO2 efflux sampling locations. b Panoramic photographs of the surfaces of Salada Jabonera, Salada Grande and Salada Pequeña
Methods
Study region
The town of Alcañiz is located in the southern part of the central Ebro basin in the Teruel Province, northeast Spain. The Ebro River, located ~ 22 km to the north of Alcañiz, bisects the basin as it flows from the northwest towards the Mediterranean Sea in the southeast. The river receives much of its discharge from snowmelt and rain-fed tributaries originating in the Pyrenees to the north and maintains perennial flow but the central basin has one of the highest annual water deficits in Europe (Herrero and Snyder 1997), with a mean air temperature of ~ 15 °C, an annual precipitation of 300–350 mm, and a mean annual potential evapotranspiration exceeding 1000 mm (e.g. Valero-Garcés et al. 2000a; Gutiérrez et al. 2005). Despite this widespread aridity, areas both south (Bajo Aragón region, including Alcañiz) and north (Monegros region) of the Ebro River host numerous saladas ranging in sizes up to ~ 200 ha (2 km2) (e.g. Sánchez et al. 1998; Gutiérrez et al. 2005; Castañeda et al. 2005; Domínguez-Beisiegel et al. 2013b). Many previous investigations have focused on the saladas in the Monegros region, particularly with respect to their geomorphological development and Quaternary history, and their present-day hydrology, geochemistry and ecology (e.g. Vizcayno et al. 1995; Samper-Calvete and García-Vera 1998; Castañeda et al. 2005, 2013; Castañeda and García-Vera 2008; González-Sampériz et al. 2008; Conesa et al. 2011; Mees et al. 2011; Domínguez-Beisiegel et al. 2013a; Gutiérrez et al. 2013; Casamayor et al. 2013). By contrast, and with a few exceptions (e.g. Ibáñez 1973, 1976; Macklin et al. 1994; Gutiérrez et al. 2005), the saladas near Alcañiz have been subject to fewer investigations. To the best of our knowledge, our study is the first to quantify soil C stocks and determine CO2 efflux for any of the saladas in the central Ebro basin, and the first to describe in detail the microbial communities of the saladas near Alcañiz.
Across the central Ebro basin, the saladas are commonly found in depressions formed by physical weathering, dissolution, and aeolian deflation of fine-grained sedimentary rocks, principally siltstones, mudstones, gypsiferous units, and limestones (Sánchez et al. 1998; Castañeda et al. 2005; Gutiérrez et al. 2005). Consequently, many saladas are partially bordered by a bedrock escarpment or elevated ground and contain a central flat area that is periodically inundated for varying durations (Domínguez‐Beisiegel et al. 2013a). Approximately 50% of the water input into the saladas derives directly from rainfall, with 40% from groundwater and the remainder from surface runoff (Castañeda and García-Vera 2008; Castañeda et al. 2013). Local geological structure and lithology can exert strong controls on water supply and quality; for example, in the Monegros region, numerous northwest-southeast trending lineaments and faults (Arlegui and Soriano 1998) contribute to a complex hydrology and hydrochemistry that can vary markedly between adjacent saladas (Castañeda et al. 2013). Nevertheless, during the northern hemisphere autumn and winter (October through late February), the salada surfaces are typically saturated and/or inundated. The water in the saladas becomes increasingly saline as it evaporates during the spring and summer. Water tables are shallow and saline (> 100 g L−1), with the high evaporation rates leading to a concentration of evaporites within the soils (Vizcayno et al. 1995; Samper-Calvete and García-Vera 1998). In the Monegros region, for example, the evaporites take several forms, and include gypsum and halitic-mirabillitic efflorescences on soil surfaces, mirabillite crystallisation in small depressions, and saline crusts formed by precipitation of bloedite and halite (Vizcayno et al. 1995).
Across the central Ebro basin, salada soils are fine-grained (silt and clay dominated), and classified as Typic Haplogypsids, Typic Haplocalcids, and Gypsic Aquisalids (Soil Survey Staff 2010, cited in Castañeda et al. 2013). Soils are typically saline, gypsiferous and enriched in organic matter compared to the surrounding soils, owing to the remains of plants, insects, algae, extremophile microbes (Domínguez‐Beisiegel et al. 2013a) and crustaceans, including in some saladas the brine shrimp Artemia salina (Valero-Garcés et al. 2000a, b). Most salada surfaces are devoid of vascular plants, but halophytes can be found around the margins where moisture and salinity conditions are more favourable (Castañeda et al. 2013). In the Monegros region, many saladas have been degraded or lost due to ploughing or levelling, with numbers declining from 136 in 1927 to 96 in 2006 (Domínguez‐Beisiegel et al. 2013b). Since the 1960s, large parts of the central Ebro basin have been irrigated, leading to water seepage, leaching, salt redistribution, and local water table uplift within and around some saladas (Vizcayno et al. 1995). These changes have led to the disappearance of some protected halophyte species such as Chenopodioideae (Conesa et al. 2011; Domínguez‐Beisiegel et al. 2013b).
Study saladas
Data were collected in May 2018 at Salada Grande, Salada Pequeña and Salada de la Jabonera de las Torrazas (hereafter shortened to Salada Jabonera), located ~ 5 km west of Alcañiz (Fig. 1a). The area covered by Salada Grande, Pequeña and Jabonera is ~ 1.24, 0.19 and 0.19 km2, respectively. The saladas have developed in an area of sub-horizontal Miocene siltstones, mudstones and ribbon or sheet sandstones and microconglomerates (sensu Friend et al. 1979). Preferential weathering of the less resistant argillaceous strata has tended to give rise to the depressions, leaving some of the more resistant calcareous sandstone and microconglomerate bodies in positive relief in the form of sinuous, inverted palaeochannels on the salada margins (Sánchez et al. 1998; Gutiérrez et al. 2005). All three saladas form part of a 7 km2 Natura 2000 site (Saladas de Alcañiz, site code ES2420114), which was designated because it is one of the most important endorheic areas on the Iberian Peninsula, with plant communities adapted to saline conditions and a rich fauna linked to seasonal inundation (European Commission, no date). There is a topographic gradient from the surface of Salada Pequeña at ~ 359 m above sea level (masl), through Salada Grande at ~ 351 masl, and to Salada Jabonera at ~ 340 masl. Although topographically the highest, Pequeña is the wettest of the three saladas (Fig. 1a, b). Shallow (< 0.5 m deep) but frequently prolonged standing water occurs over much of the salada surface, partly because of rainfall and runoff but also because of contributions from groundwater and a small stream that periodically inflows from the southwest (Fig. 1a). Salada Grande is drier and Salada Jabonera drier still, but both typically experience shallow standing water over at least part of their surfaces for some of the year as a result of rainfall and runoff, and possibly some limited groundwater contributions. In all three saladas, the ephemeral nature of the standing water, combined with the widespread dissolution of halite and gypsum and its subsequent evaporative concentration, results in alkaline soils (Gutiérrez et al. 2005). Analysis of water extracts from saturated pastes of Salada Grande sediments indicates that soils are dispersive when wet, but prone to forming pellets when desiccating, which promotes aeolian deflation during dry periods (Gutiérrez et al. 2005). The surfaces of Pequeña and Grande are mostly unvegetated, except at the fringes where soils are elevated above the salada surface by a few metres (Fig. 1a, b). Salada Jabonera, the driest of the saladas, has a patchy cover of vascular halophytic shrubs and grasses across the surface (Fig. 1a, b), including Halopeplis amplexicaulis (Vahl) Ung.-Sternb., Arthrocnemum macrostachyum (Moric.) K. Koch, Microcnemum coralloides (Loscos & Pardo) and Atriplex halimus L. (Amaranthaceae). Around all three saladas, there are also localised zones where overland flow has transported fine-grained soil and associated organic matter from the surrounding slopes short distances onto the salada surface, and wind erosion may also contribute soil and organic matter. The lower topographic setting of Salada Jabonera with its steeper northern and eastern margins (Fig. 1b) may result in an enhanced contribution of soil and organic matter from overland flow.
Surface water coverage
To quantify the areal extent and frequency/duration of water coverage on each salada over six years, data were extracted from a time series of 86 cloud-free Landsat 8 OLI (2013–2016) and Sentinel-2 L1C (2016, 2017 and 2018) satellite images. Scenes were downloaded via EarthExplorer (USGS) and converted to Top of Atmosphere Reflectance using the Semi-Automatic Classification Plugin (SCP) in QGIS 3.4. Red, green (\(\rho Green\)) and blue bands were stacked to generate true-colour composite images, with near infrared and shortwave infrared bands (\(\rho SWIR\)) also used to aid water detection. A Modified Normalised Difference Water Index (MNDWI) was used to enhance the identification of surface water (Eq. 1; Xu 2007; Du et al. 2016; Li et al. 2018, 2021).
For each image, surface water area was manually digitised using ArcGIS 10.5. Mapping is deemed accurate to ± 1 pixel (20 m for Sentinel and 30 m for Landsat MNDWI). Applying a 20 m/30 m buffer to our digitised surface water areas, we calculated a maximum areal uncertainty of ± 12%, although uncertainty is considered much lower owing to the distinct spectral difference between the water surface and adjacent bare soil surface in the true-colour composites.
Soil moisture and temperature
The moisture content of salada soils from the surface to 0.05 m (probe length) was recorded in a grid pattern every 10 m across the surface of each salada using a DeltaT SM150 probe (n = 100, 120 and 180 on Saladas Pequeña, Grande and Jabonera, respectively). The grids corresponded approximately to the area of the sampling sites for CO2 efflux determination (Fig. 1a), although additional soil moisture readings were taken with the DeltaT probe adjacent to the chambers used to determine CO2 efflux (“Soil CO2 efflux” section).
Soil physical and chemical properties
On each salada, four pits up to 0.5 m deep (to the water table) were excavated at locations evenly spaced across the centre of the soil moisture sampling grid. From each pit, up to five soil samples were collected from a range of depths (see Supplementary Information), then air dried and bagged prior to analyses. H+ ion activity was determined in a 1:2.5 soil–water mixture using a pH probe. Approximately 5 g of each sample was dried at 105 °C to a constant weight and heated at 430 °C for 16 h in a muffle furnace to determine organic matter by loss-on-ignition. Bulk density was determined after weighing the oven dry mass of a soil sample collected from the subsurface using a stainless steel tube with an internal volume of 99 cm3. Together with soil C data, bulk density was used to determine soil C stocks using Eq. 2:
where OC and IC are the mean concentrations of soil organic and inorganic carbon (%) respectively, and BD is the bulk density (in kg m−3).
An elemental analyser (vario PYRO cube, Elementar UK Ltd.) was used to determine the total C, N and S content of 10–30 mg sub-samples in tin capsules (the mass was dependent on the amount of loss-on-ignition). C stable isotope measurements were made on the same sub-samples using a coupled mass spectrometer (visION, Elementar UK Ltd.). Organic C content and isotopic compositions were determined from separate sub-samples, after removal of inorganic C with 50 μL of 10% HCl in silver capsules (Brodie et al. 2011) and then overnight drying at 60 °C. Stable isotope data are reported as δ13CVPDB values, the proportion per thousand (‰) variation from the ratio of 13C/12C in V-Pee Dee Belemnite (VPDB) (Thompson et al. 2006; Ibell et al. 2013) (Eq. 3):
where Rsample C indicates the ratio of 13C/12C in the sample, and RVPDB is the ratio of 13C/12C in the VPDB. Isotope ratio measurements were calibrated to the VPDB scale using commercially available standard reference materials (B2205 EMA P2, B2153 low organic content soil, B2151 high organic content sediment and B2159 sorghum flour, all from Elemental Microanalysis, UK). Further elemental analysis on each sediment sample was performed with a Niton XL3t 950 GOLDD+ portable X-ray fluorescence spectrometer (pXRF). Data in the results section of this article are reported as depth averages, with depth specific data reported in Supplementary Information (S1).
Soil CO2 efflux
To quantify soil CO2 efflux, respiration chambers were positioned at 12 locations in a grid across the water-free surface of each salada (Fig. 1a). Maximum distances between chambers were c. 50 m on Salada Pequeña, c. 120 m on Salada Grande and c. 70 m on Salada Jabonera. Comprehensive details of the chamber design and method can be found in Thomas (2012) and Thomas et al. (2018). In summary, the chambers are made from white uPVC and comprise two parts: (i) a lower chamber that when pushed 3 cm into the surface forms an air-tight seal; and (ii) a screw-on lid that enables soil gases to accumulate inside the chamber and to be sampled during measurement cycles. The chamber lids contain a sampling port covered with a Suba seal for gas extraction and a two-way valve to ensure any pressure differences between the chamber and atmosphere are minimal and rapidly equilibrated. Chamber surface area is 83 cm2 and in this study chamber, volume ranged from 0.48 to 0.52 L depending on insertion depth. Heat sinks mounted through the chamber walls ensured the internal air temperatures were not elevated above ambient. Three of the 12 chambers were equipped with a sensor (USB502, Adept Science, UK) to record the air temperature and humidity inside the chamber at 2 min intervals.
Measurements were taken three times each day to capture a range of temperature and light conditions, giving a total of 36 measurements at each of the three saladas. Following previous sampling protocols to determine soil CO2 efflux (cf. Thomas 2012; Thomas et al. 2018), the lid was placed on the chamber and 12 ml of gas was immediately extracted through the sample port using a syringe and hypodermic needle and secured with a Leur lock. After approximately five min, another syringe was gently pumped to mix the air within the chamber before a second sample was collected. Both sample CO2 concentrations were determined immediately after each of the three measurement cycles using an EGM-4 infrared gas analyser (PP Systems, Amesbury, USA). Mass CO2 flux in mg m−2 h−1 was determined from the changes in CO2 concentration normalised to mean temperature and pressure during measurement (Kutzbach et al. 2007). To correct for the effect of any diffusion suppression owing to the accumulation of CO2 inside the chamber, a diffusion correction factor was applied (for details see Thomas 2012). At the time of sampling, surface soil temperature and moisture adjacent to each respiration chamber were determined (n = 3) using an infra-red thermometer and a soil moisture probe (SM150, Delta-T Devices Ltd., Cambridge, UK).
DNA extraction and sequencing
At two locations on each salada, one gram of soil was collected from three depths (0–1 cm, 20–21 cm and 49–50 cm) for DNA extraction. Samples were stored in a 15 mL aseptic centrifuge tube containing 2 mL of Life Guard Soil Preservation solution (Qiagen, UK). The DNA was amplified by polymerase chain reaction (PCR) and sequenced for three primers, although sequencing failed for six of the 54 analyses (because of low DNA concentrations). Total DNA was extracted using a DNA extraction kit (Omega, USA), and DNA concentration and purity were determined using a NanoDrop 2000 spectrophotometer (Thermo Scientific, USA). The quality of the extracted DNA was verified using 1% agarose gel electrophoresis, before amplification using a GeneAmp 9700 PCR System (ABI, USA). Prokaryotic bacterial 16S rRNA genes were amplified using primers 338F and 806R (Lan et al. 2021). Archaeal 16S rRNA genes were amplified using primers 524F10extF and Arch958RmodR (Ye et al. 2021). Eukaryotic 18S rRNA genes were amplified using primers 3NDF and 4-euk-R2R (Zhang et al. 2018). The PCR reaction systems were done under conditions previously published (Lan et al. 2021), and the PCR amplification program comprised the following cycles: (i) initial denaturation at 95 °C for 3 min; (ii) 29 (bacteria), 37 (archaea) and 37 (eukaryotes) cycles at 95 °C for 30 s, annealing at 53 °C for 30 s, and elongation at 72 °C for 45 s; and (iii) the final extension at 72 °C for 10 min. The PCR products were extracted from a 2% agarose gel, and further purified using an AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, U.S.A.). The purified PCR products were then quantified using a QuantiFluor-ST quantitative system (Promega, USA) and sequenced on an Illumina MiSeq platform (Illumina, USA).
The raw sequencing data were quality filtered (< Q20) using a Trimmomatic trimmer, and merged using FLASH (Magoč and Salzberg 2011). The chimera were eliminated using UCHIME, and the high-quality sequences were classified into different operational taxonomic units (OTUs) at a 97% similarity cut-off using UPARSE (Yuan et al. 2018). The taxonomic information from all the sequences was annotated by an RDP classifier using the Silva database (SSU132), with a confidence threshold set at 70%. Shannon index was analysed by Mothur (V1.30.1), with the details for this index calculation described at https://mothur.org/wiki/shannon/.
Statistical analyses
Statistical analyses of the soil chemical and CO2 efflux data were performed using SPSS (IBM v. 25). One-way analysis of variance (ANOVA) was used to test whether mean values of the dependent factors (moisture, temperature, total C, total N, S, pH, δ13C) were significantly different between saladas. For CO2 efflux, where multiple readings were taken at each site, repeated measure ANOVA was used. The Levene’s F statistic was used to test equality of variance. Although ANOVA can tolerate inhomogeneous variance, where these conditions were not met, the more robust Welch and Brown Forsythe tests of significance were used. Tukey’s HSD post-hoc test was undertaken to determine whether saladas were significantly different with a probability of p < 0.05.
Results
Salada hydroperiods
Over the six year monitoring period, the saladas had contrasting surface water inundation regimes (Table 1, Fig. 2). Salada Pequeña had water covering up to 50% of the surface for 86% of the time, with > 50% water coverage for 47% of the time, and > 80% water coverage for 10% of the time. Saladas Jabonera and Grande were both drier. On Salada Grande, water covered up to 50% of the surface for only 26% of the time, with > 50% water coverage for only 16% of the time (Fig. 2). Salada Jabonera was drier still, with water covering up to 50% of the surface for only 19% of the time, and water coverage never exceeding > 50%. The soil moisture conditions at the time of field sampling (May 2018) reflected the longer term differences in surface water inundation regimes (Table 1), with significant differences evident in the mean soil moisture content for the three saladas (F = 87.4, df = 2, p < 0.001) (Fig. 3a). Nevertheless, on all three saladas, large within-site variations were evident (Fig. 3a), with moisture typically increasing with distance from the fringes towards the salada centres. Mean soil temperatures were similar at Saladas Grande and Jabonera but significantly warmer at Salada Pequeña (df 2, f 7.0, p ≤ 0.001) but variations in soil temperature within each salada were also large (Fig. 3b).

Frequency (columns) and cumulative frequency (dashed lines) distributions of the area covered by surface water in each of the saladas over the 6 year monitoring period

Data from the three saladas, illustrating within- and between-site variations in: a moisture; b temperature; and c CO2 efflux. Individual values shown with mean (adjacent larger marker) and standard deviation (line). Horizontal displacement of the data points for each salada are partly for clarity but partly schematic as they correspond to the shorter lines of sampling locations in each of the sampling grids (see Fig. 1a). p values indicate significance of differences
Soil Properties and C Stocks
All salada soils were alkaline (pH 8.8–9.1) but differences in sediment chemistry between the saladas are evident (Table 2). Concentrations of K+, Ca+ and organic matter were lowest in the soils of the wetter Salada Pequeña, intermediate in Salada Grande and significantly larger in the drier Salada Jabonera (Table 2). Concentrations of S were lowest in the soils of the drier Salada Jabonera and highest in the soils of the wetter Salada Pequeña. Fe concentrations were very similar to each other in the soils of the drier Saladas Jabonera and Grande but significantly lower in the soils of the wetter Salada Pequeña (Table 2).
Total C stocks were greatest on the drier Salada Jabonera and least on the wetter Salada Pequeña (Fig. 4). Total C stocks on all saladas were dominated by the inorganic fraction (Table 3, Fig. 4), comprising 75% of the total C stock on the wetter Salada Pequeña and 90% on the drier Saladas Grande and Jabonera. Organic C concentrations were highest on the wetter Salada Pequeña and lower on the drier Saladas Grande and Jabonera (Table 3, Fig. 4). For all three saladas, the δ13CVPDB of total C was less negative than the organic C fraction (Table 3). There were also significant differences in the δ13CVPDB of the soil, with the wetter Salada Pequeña having the most negative δ13totalC and the drier Saladas Grande and Jabonera having less negative δ13OrgC (Table 3).

Inorganic (dark shade) and organic (lighter shade) carbon stocks to 0.5 m depth in Saladas Jabonera (Jab), Grande (Gra) and Pequeña (Peq)
Soil CO2 efflux
At the time of sampling, soils on all three saladas was a net source of CO2, although there was a limited number of individual negative efflux readings on Saladas Pequeña and Jabonera, indicating intermittent net CO2 uptake (Table 4 and Fig. 3c). Soil CO2 efflux varied significantly between saladas (F = 18.3, df = 2, p ≤ 0.001), with the mean value from Salada Grande significantly larger than the mean values from Saladas Pequeña and Jabonera. Nevertheless, there was also significant variation in soil CO2 efflux within each salada in both space and time (i.e. through the measurement day) (Fig. 3c), and this is quantified in Table 4 as the coefficient of variation. Spatial variations in CO2 efflux were smaller on Salada Pequeña (58.0%) and Salada Grande (30.5%) and larger on Salada Jabonera (95.6%) (Table 4). Temporal variations in CO2 efflux were smallest on Salada Pequeña (38.0%) and Salada Grande (58.4%) and larger on Salada Jabonera (71.1%) (Table 4).
The combined CO2 efflux data from all three saladas are not well predicted by soil temperature (Fig. 5a). This can be explained by the contrasting CO2 efflux-temperature relationships on each salada (Fig. 5b–d). Soil temperature alone explains very little of the variance in CO2 efflux on the wetter Salada Pequeña (Fig. 5d) or on the drier Salada Jabonera (Fig. 5b). On Salada Grande, however, 40% of the variance in soil CO2 efflux is explained by temperature (Fig. 5c). Soil moisture content is a slightly better single variable predictor of CO2 efflux from the three saladas than temperature, with a tendency for CO2 efflux to be higher at intermediate moisture levels of between 40 and 60% vol/vol (hereafter v/v) (Fig. 6a). At higher moisture contents (> 75% v/v) where the soil is saturated, and where soil moisture is low (< 25% v/v), CO2 efflux tends to be lower and less variable (Fig. 6a–d). On Salada Pequeña, the initially high moisture content means that efflux tends to decline with increasing moisture (Fig. 6d) but on the drier Saladas Grande and Jabonera, the initially lower moisture contents mean that efflux tends to increase with moisture (Fig. 6c, d).

Soil CO2 efflux and soil temperature at: a all sites; b Jabonera; c Grande and d Pequeña

Soil CO2 efflux and soil moisture at: a all sites; b Jabonera; c Grande and d Pequeña
Microbial communities
For soils from the three saladas, Shannon index values ranged from 0.3 to 4.4 for bacteria, 1.6 to 3.7 for eukaryota, and 0.7 to 4.0 for archaea (Fig. 7). The bacterial and eukaryotic diversities represented by the Shannon index were not significantly different between saladas (p > 0.05; Fig. 7), but the archaeal diversity in Salada Grande was significantly higher than in Saladas Pequeña and Jabonera (p < 0.05; Fig. 7). At phylum level, the composition of bacteria, eukaryota and archaea were similar in the saladas, but their relative abundances varied between the saladas (Fig. 8). Bacterial communities were dominated by Proteobacteria and Firmicutes, and together these phyla accounted for 83.4% of bacterial sequences (Fig. 8a). The Proteobacteria were more abundant in Salada Pequeña, while Firmicutes were more abundant in Saladas Jabonera and Grande. The most abundant eukaryotic sequences belonged to Chloroplastida (30.5%), Amoebozoa (8.5%) and Fungi (8.0%) (Fig. 8b). The Chloroplastida and Fungi were more abundant in Saladas Jabonera and Pequeña, while Amoebozoa were more abundant in Salada Grande. The archaea were dominated by Euryarchaeota (53.6%) and Thaumarchaeota (41.2%) (Fig. 8c). The Euryarchaeota were more abundant in Salada Grande, while Thaumarchaeota were more abundant in Saladas Jabonera and Pequeña.

Soil microbial diversities at Saladas Jabonera (Jab), Grande (Gra) and Pequeña (Peq). The box lines show the 25th, 50th and 75th percentiles and error bars show the 5th and 95th percentiles. * indicates the Shannon index of archaea is significantly higher in Salada Grande

Soil microbial composition at Saladas Jabonera (Jab), Grande (Gra) and Pequeña (Peq): a bacteria; b eukaryota; c archaea
Interpretation and discussion
The three saladas near Alcañiz form a hydrological continuum. Our findings show how their contrasting hydroperiods are associated with different soil properties, C stocks, CO2 efflux and microbial communities. Changes in these soil properties and functions in response to increasing or decreasing wetness are not necessarily linear, but many statistically significant differences between the wetter and drier extremes are nonetheless evident. By integrating our interpretations of the hydrological controls on these soil properties, functions and microbial characteristics, we can use an ergodic approach (space-for-time substitution) to assess how these saladas, and potentially other similar saladas in the central Ebro basin and farther afield, may respond to twenty-first century climate or human-induced hydrological changes. The ergodic approach is widely used across geomorphology, soil science and ecology to infer sequences of changes that cannot be monitored in real time (e.g. Walker and Coventry 1976; Birkeland 1992; Huggett 1998; Fryirs et al. 2012). For instance, with caution, the ergodic approach can be used to reconstruct sequences of changes to landforms, processes and ecological community structures that have occurred in the past, or that may occur in future. Previous studies in South Africa and Australia have successfully used this approach to assess potential changes to floodplain wetlands that occur across wetter-to-drier hydroclimatic gradients (Grenfell et al. 2014; Larkin et al. 2017, 2020).
Soil organic carbon (OC)
We hypothesised (H1) that relatively wet soil moisture conditions would stimulate biological productivity and elevate OC stocks in comparison to drier saladas. On each of the three saladas, soil OC is likely derived from various sources, including microbes, aquatic organisms, birds, insects, plants, and soil washed or blown in from the surrounding landscape. The proportion of soil OC derived from these sources will change with water availability. The highest concentrations of soil OC were found in the wetter Salada Pequeña (Table 3, Fig. 4). The wetter (frequently inundated and/or near-permanently saturated) condition of Salada Pequeña (Fig. 1b, Table 1) allows aquatic life to flourish, particularly algae. At the time of sampling, for instance, an ~ 0.2–0.5 cm thick black sapropelic layer (ooze) rich in decaying organic matter was present beneath the surficial saline crusts. Similar observations of microbial mats and associated sapropelic layers have been made on other saladas in the central Ebro basin (e.g. Pueyo 1980; Valero-Garcés et al. 2000a, b; Gutiérrez et al. 2005) and in saline inland wetlands farther afield (McKay et al. 2016), and indicate the potential for these deposits to form significant OC inputs. Aquatic life also includes various crustaceans, other micro-invertebrates, and amphibians. These provide potential food sources and attract aquatic and wading birds (at the time of sampling there were protected Avocet (Recurvirostra avosetta) nesting sites near the inflowing stream on the southwestern margin—Fig. 1a), which will also add organic material to the water column and sediment. Plants are entirely absent from the salada surface, presumably because the saturated, saline conditions inhibit their establishment, and are only present on the higher surfaces elevated above the level of inundation (Fig. 1a). In contrast, at the time of sampling on the drier Salada Jabonera (Fig. 1c, Table 1), there was little evidence of aquatic or bird life, but there are patches of halophytic shrubs across much of the salada surface and a greater density and diversity of shrubs and grasses around the margins. The plants may contribute spatially localised organic matter inputs and alter the microclimate. Salada Grande, with its intermediate surface water and soil moisture conditions (Fig. 1b, Table 1), has characteristics both of Salada Pequeña and Salada Jabonera. At the time of sampling, it was largely dry with little evidence of aquatic or bird life, and while the salada surface is largely devoid of plants, patches of halophytic shrubs and grasses grow in a narrow band at the margins (Fig. 1b). These different combinations of OC sources may explain why the δ13CVPDB of the soil OC fraction from the drier Salada Jabonera is significantly less negative than in soils from Saladas Pequeña and Grande (Table 3). Typical δ13C values for C3 plants (i.e., generally shrubs and trees) range from − 22 to − 38‰ and for C4 plants (generally grasses) range from − 8 to −15‰ (Yeh and Wang 2001), which suggests that a greater proportion of soil OC on Salada Jabonera may be derived from plant inputs, namely the shrubs and grasses around the salada margins, as well as plant litter and soil inputs associated with overland flow from the steeper northern and eastern margins.
Soil inorganic C (IC)
Compared to the soil OC concentrations, the higher concentrations of soil IC in all three saladas suggests that they may be active sites of carbonate formation due to a combination of evaporative concentration and the reaction of CO2 with available ions such as Ca2+, Mg2+ and Na+ (Thomas et al. 2014; Zhao et al. 2016). For all three saladas, the δ13CVPDB of total C was less negative than the OC fraction (Table 3), suggesting that the isotopic signal from plant and animal material is diluted by the isotopic signal from carbonates. However, because many parts of the central Ebro basin are underlain by carbonate-bearing lithologies (e.g. limestone, calcareous sandstone) and other evaporites we cannot discern from our data whether the IC is from geological sources (i.e. derived from bedrock at the base of the soil profile, or supplied by overland flow or aeolian inputs) or whether it is being formed in situ through near-surface pedogenic processes. Nevertheless, although the exact source of soil IC remains unclear, Xie et al. (2009) reported rates of non-biological CO2 adsorption by alkaline soils of 0.3–3.0 μmol m−2 s−1 and concluded that this mechanism is likely to be associated with a hugely underestimated global terrestrial C sink. In Xie et al.’s (2009) study, the intensity of non-biological CO2 absorption increased with soil salinity and soil alkalinity, and it is reasonable to assume that the saladas are also sites of active and significant carbonate formation. Our hypothesis (H2) was that drier saladas would have greater soil IC than wetter saladas if the evaporative concentration of solutes was the dominant process of carbonate formation. In our study, the soil IC in the wetter Salada Pequeña was lower than IC concentrations in the drier Saladas Grande and Jabonera, and also comprised a lower proportion of total C (Table 3, Fig. 4) suggesting that evaporative concentration could be a dominant process of IC formation. This pattern is to be expected, for carbonate-forming conditions tend to be more favourable in drier soils where longer periods of water loss by evaporation lead to an increase in salinity and alkalinity (Xie et al. 2009).
CO2 efflux
At the time of the measurements, the salada soils were a net source of CO2 to the atmosphere (Table 4) with mean CO2 efflux similar to the ~ 74 mg CO2 m−2 h−1 reported in Thomas et al. (2014) from the hyper-saline Ntwetwe pan in Botswana. There were occasional instances of net CO2 uptake on Saladas Jabonera and Pequeña (Fig. 3c) which could be related to photosynthesis in biological crusts and algal mats (c.f. Thomas et al. (2014) on Ntwetwe pan) or by abiotic (physicochemical) processes (c.f. Yates et al. (2013) for alkaline playa sediment in Nevada, USA).
The highest CO2 efflux was found at intermediate soil moisture conditions of c. 40–60% v/v with lower values when soil moisture was > 75% v/v or < 25% v/v (Fig. 6). These data support our initial hypothesis (H3) regarding the nature of the relationship between CO2 efflux and soil moisture (Orchard and Cook 1983; Schimel et al. 1999; Moyano et al. 2013), where high soil moisture contents greatly reduce gas diffusion, and the metabolic activity of aerobic microorganisms is impeded by oxygen deficit (Moyano et al. 2013). Nevertheless, soil CO2 efflux is highly spatially and temporally variable both within and between the saladas (Table 4, Fig. 3c). In saturated or near-saturated soils, like those in Salada Pequeña, temperature has very little effect on CO2 emissions (Fig. 5d). In drier soils, like those in Salada Jabonera (Fig. 5b), CO2 efflux also can become insensitive to temperature (c.f. Thomas 2012).
The complex spatial and temporal patterns of soil CO2 efflux from the three saladas illustrate the pitfalls of making single variable predictions using either soil moisture or temperature, something Rey et al. (2017) conclude could lead to errors and inappropriate interpretations if using these variables to predict the response of soil CO2 efflux to climate change in drylands. The very high spatial and temporal variability in CO2 efflux demonstrates the need for extensive sampling, for single point measurements of efflux in time and space are unlikely to provide a reliable estimate of CO2 emissions from the saladas.
Microbial diversity and composition
Our soil microbial data (Figs. 7 and 8) reflect the broad microbial genetic diversity observed in water samples from 11 shallow, saline lakes in the Monegros region (Casamayor et al. 2013). In their study of bacteria, eukaryota and archaea, Casamayor et al. (2013) found that water salinity levels had little or no impact on microbial ecological diversity (Shannon–Weaver index) or genetic diversity (novelty level). In our study, we hypothesised (H4) that there would be different soil microbial communities in each of the three saladas as a direct or indirect consequence of their contrasting inundation regimes and soil moisture content. From our data, salada surface water regimes and soil moisture conditions appear to have no effect on bacterial and eukaryotal diversities, although archaeal diversity was significantly higher in Salada Grande with its intermediate moisture conditions (Fig. 7). There are, however, different microbial community structures in each salada. These results suggest that different microbial species may be involved in the C metabolisms and cycles depending on the water availability in the salada. Consequently, these distinct community structures will likely lead to differences in C storage between different saladas (Vikram et al. 2015; Maier et al 2018). Prokaryotic Proteobacteria and Firmicutes (Fig. 8a), together with eukaryotic Fungi (Ascomycota) and Amoebozoa (Protosporangiida) (Fig. 8b) are likely responsible for the majority of soil OC respiration and CO2 emissions (Zhang et al. 2018; Swenson et al. 2018). Cyanobacterial abundance was very low in the salada soils, especially when compared to biocrusts and soils in other playas worldwide (e.g. Elliott et al. 2014; Thomas et al. 2014). This low abundance may be related to the high salinity or C content in the saladas, and help explain the infrequent negative CO2 efflux detected on the salada soils. Nevertheless, Chloroplastida (Chlorophyta), a common dryland eukaryotic photosynthetic microbe (Bates and Garcia-Pichel 2009; Zhang et al. 2018) is also widespread in the salada soils (Fig. 8b), which suggests that it may generate microbial OC in the salada soils. However, the prevalence and persistence of microbial C in salada soils remains an area of uncertainty as other studies have shown that periods of microbial photosynthesis and net CO2 uptake are intermittent, unpredictable, and likely to be only short-lived (Thomas et al. 2014, 2022; Williams et al. 2014). Euryarchaeota were detected in all the salada soils and they are known to include methanogens (Watanabe et al. 2011). Given the saturated condition of some of the salada soils, at least for part of the year, CH4 may also be generated and released, but we do not have the data to test this notion.
Potential changes to the C cycle in saladas with twenty-first century warming
Many of the world’s drylands are predicted to warm by 3 to 4 °C and experience a reduction in precipitation of 5 to 30% by the end of this century (Lin et al. 2015), including those in Spain (Drobinski et al. 2020). These changes will have a profound effect on multiple ecosystem characteristics and functions within many wetlands in drylands (Huang et al. 2016). For example, for many riverine wetlands, increasing aridification, coupled with greater hydroclimatic variability, may induce major changes to channel-floodplain wetland structure and thus patterns of water and sediment dispersal (Tooth 2018; Larkin et al. 2017, 2020). To date, such studies have not considered the implications of such physical and associated ecological changes for the C cycle, although many aspects of the C cycle are likely to be affected. For saline inland wetlands (including saladas) in particular, the impacts of climate change on C cycles are essentially unknown (IPCC 2014b).
With twenty-first century warming, major structural changes to the saladas near Alcañiz, or more generally across the central Ebro basin, are unlikely. The saladas are likely to remain as topographic depressions in an increasingly arid landscape. Nonetheless, these climatic changes will impact on regional hydrology (e.g., lower and more variable rainfall, greater evaporation) and will likely result in altered salada hydroperiods (e.g., less frequent, shorter inundation phases, and more frequent, longer drying phases). Hence, for many saladas there may be a temporal shift along the hydrological continuum from wetter to drier conditions (i.e., a shift from a Pequeña-type situation through a Grande-type situation to a Jabonera-type situation). These hydrological changes may be exacerbated or compounded by human impacts (e.g., river diversion or water abstraction that results in groundwater table lowering). More rarely, some saladas may experience a temporal shift from drier to wetter conditions; for example, where excess irrigation runoff or deliberate damming leads to raising of local groundwater tables and establishment of permanent lakes, as has happened at La Estanca, a former salada located ~ 5 km to the west-northwest of Alcañiz (Gutiérrez et al. 2005), and also at La Laguna de Sariñena, a former salada in the northern Monegros region (Rodrigo and Alastrué, no date).
In both increasing drying or increasing wetting scenarios, C cycling is likely to be affected. However, few studies have investigated the C cycle in wetlands in drylands, despite increasing studies of soil C and CO2 efflux in wetlands generally (e.g. Mitsch et al. 2010; Nahlik and Fennessy 2016; Rogers et al. 2019; Were et al. 2019), Consequently, many knowledge gaps remain, particularly regarding how climate changes will affect the mechanisms and rates of IC formation, the residence times of OC, and the interaction between OC and IC. These gaps contribute to uncertainty surrounding the size of the C stock in wetlands in drylands as well as the processes that lead to C uptake and release, and to CO2 emissions. However, it is evident from our work and other studies in the region that the saladas near Alcañiz, and possibly similar features across the wider central Ebro basin, are currently sites of C storage, contain active microbial communities and typically release CO2 to the atmosphere. Although the impacts of aridification will depend on the initial hydration status of the saladas, our findings suggest that we can perhaps expect a reduction in OC stocks, as the OC sources most vulnerable to declines in water availability are those associated with stream inputs and aquatic fauna (e.g., crustaceans, amphibians, bird life). Aridification of the saladas may increase soil IC formation through evaporative concentration, at least in the short term. Over longer time periods, desiccation will reduce the supply of cations and bicarbonate to the saladas, thus limiting the capacity to form carbonates. In addition, a reduction in sediment moisture content in initially very wet saladas is likely to lead to an increase in CO2 efflux as conditions become more favourable for aerobic microbial activity in the soil. The increase in microbial activity may reduce soil OC residence times as it is more likely to be respired. Further drying of saladas where soil moisture content is already typically < 50% v/v may inhibit microbial activity and lead to a reduction in CO2 efflux. Any future reduction in the extent and duration of inundation and/or soil saturation on the saladas may shift the composition of C stocks to less organic-rich, more carbonate-based stocks, at least in the short term, with an overall decline in the total C stock.
Caution needs to be exercised in extrapolating the findings from the saladas near Alcañiz and those in the wider central Ebro basin to other saladas in Spain or to saline inland wetlands more generally (e.g., Zhang et al. 2020). Combined with the aforementioned process uncertainties, different playas may respond to changes in hydroperiods in different ways depending on the dominant ions in solution. Evaporation of water from saladas with appreciable concentrations of calcium bicarbonate or calcium sulphate is likely to result in the precipitation of calcrete (and/or gypsum) and thus increase IC stocks (Eugster and Jones 1979; Tooth and McCarthy 2007). Where saladas are dominated by sodium bicarbonate, sodium chloride, or sodium sulphate, evaporation is more likely to result in the development of increasingly saline surface water that is toxic to many biota and lead to a reduction or change in OC inputs (Eugster and Jones 1979; Tooth and McCarthy 2007).
The potential influence of changing hydroperiods on the soil microbial community in saladas and the implications for OC and IC formation, storage, and release also remains poorly understood. Our data (Fig. 8) suggest that prevailing moisture conditions affect the soil microbial community structure through changes to relative abundances. A reduction in salada soil moisture may lead to lower relative abundances of Proteobacteria, Amoebozoa and Euryarchaeota, ultimately influencing C assimilation, metabolism and storage. Nevertheless, drying of saladas may also cause greater aerobic microbial activity (at least initially), which will probably reduce soil OC residence times and further affect the microbial community structure (Swenson et al. 2018). These uncertainties notwithstanding, our data can serve as a framework to guide future investigations.
Conclusions
Three adjacent saladas near Alcañiz with contrasting surface water regimes and soil moisture conditions provided an opportunity to investigate, for the first time, how water availability affects their C stocks, CO2 emissions, and microbiology. Frequent inundation and/or near-permanent soil saturation promotes higher soil OC stocks owing to input from a greater range of OC sources, especially biological. Drier salada soils, however, have higher soil IC stocks owing to evaporative concentration and formation of carbon-bearing mineral compounds. CO2 efflux is highest at intermediate moisture conditions, but the spatial and temporal variability in CO2 efflux on salada surfaces was very high, demonstrating the need for intensive sampling regimes to provide realistic estimates of their contribution to atmospheric CO2 exchanges. There were also different microbial community structures in each salada, suggesting a possible link to moisture availability. These three saladas, and numerous other similar features across the central Ebro basin, are important areas of C storage and likely play an important role in local and regional C cycles. They also host many rare and threatened plants and animals (Conesa et al. 2011; European Commission, no date), and their C cycle and microbial community characteristics provide an additional reason to monitor changes resulting from twenty-first century climate changes and protect these vulnerable environments from anthropogenic disturbances.
References
Arlegui LE, Soriano MA (1998) Characterizing lineaments from satellite images and field studies in the central Ebro Basin (NE Spain). Int J Remote Sens 19:3169–3185
Banerjee S, Helgason B, Wang L, Winsley T, Ferrari BC, Siciliano SD (2016) Legacy effects of soil moisture on microbial community structure and N2O emissions. Soil Biol Biochem 95:40–50
Bates ST, Garcia-Pichel F (2009) A culture-independent study of freeliving fungi in biological soil crusts of the Colorado Plateau: their diversity and relative contribution to microbial biomass. Environ Microbiol 11:56–67
Birkeland PW (1992) Quaternary soil chronosequences in various environments—extremely arid to humid tropical. Editor(s): I.P. Martini, W Chesworth. Dev Earth Surf Process 2:261–281
Brodie CR, Leng MJ, Casford JSL, Kendrick CP, Lloyd JM, Yongqiang Z, Bird MI (2011) Evidence for bias in C and N concentrations and δ13C composition of terrestrial and aquatic organic materials due to pre-analysis acid preparation methods. Chem Geol 282:67–83
Casamayor EO, Triadó-Margarit X, Castañeda C (2013) Microbial biodiversity in saline shallow lakes of the Monegros Desert, Spain. FEMS Microbiol Ecol 85:503–518
Castañeda C, García-Vera MA (2008) Water balance in the playa-lakes of an arid environment, Monegros, NE Spain. Hydrogeol J 16:87–102
Castañeda C, Herrero J, Casterad MA (2005) Facies identification within the playa-lakes of the Monegros desert, Spain, from field and satellite data. Catena 63:39–63
Castañeda C, Herrero J, Conesa JA (2013) Distribution, morphology and habitats of saline wetlands: a case study from Monegros, Spain. Geol Acta 11(4):371–388
Conesa JA, Castañeda C, Pedrol J (2011) Las saladas de Monegros y su entorno. Hábitats y paisaje vegetal. Zaragoza (Spain). Consejo de Protección de la Naturaleza de Aragón pp 539
Drobinski P, Da Silva N, Bastin S, Mailler S, Muller C, Ahrens B, Christensen OB, Lionello P (2020) How warmer and drier will the Mediterranean region be at the end of the twenty-first century? Reg Environ Change 20:78
Domínguez-Beisiegel M, Castañeda C, Herrero J (2013a) Two microenvironments at the soil surface of saline wetlands in Monegros, Spain. Soil Sci Soc Am J 77(2):653–663
Domínguez-Beisiegel M, Herrero J, Castañeda C (2013b) Saline wetlands’ fate in inland deserts: an example of 80 years’ decline in Monegros, Spain. Land Degrad Dev 24:250–265
Du Y, Zhang Y, Ling F, Wang Q, Li W, Li X (2016) Water bodies’ mapping from Sentinel-2 imagery with Modified Normalized Difference Water Index at 10-m spatial resolution produced by sharpening the SWIR band. Remote Sens 8(4):354
Duarte CM, Prairie YT, Montes C, Cole JJ, Striegl R, Melack J, Downing JA (2008) CO2 emissions from saline lakes: a global estimate of a surprisingly large flux. J Geophys Res 113:G04041
Elliott DR, Thomas AD, Hoon SR, Sen R (2014) Niche partitioning of bacterial communities in biological crusts and soils under grasses, shrubs and trees in the Kalahari. Biodivers Conserv 23:1709–1733
Eugster HP, Jones BF (1979) Behavior of major solutes—closed-basin brine evolution. Am J Sci 279:609–631
European Commission (no date) Natura 2000. https://ec.europa.eu/environment/nature/natura2000/index_en.htm. Accessed 1 June 2021
Ezcurra E (ed) (2006) Global Deserts Outlook. United Nations Environment Programme, Nairobi
Friend PF, Slater MJ, Williams RC (1979) Vertical and lateral building of river sandstone bodies, Ebro Basin, Spain. J Geol Soc Lond 136:39–46
Fryirs K, Brierley GJ, Erskine WD (2012) Use of ergodic reasoning to reconstruct the historical range of variability and evolutionary trajectory of rivers. Earth Surf Proc Land 37:763–773
González-Sampériz P, Valero-Garcés BL, Moreno A, Morellón M, Navas A, Machín J, Delgado-Huertas A (2008) Vegetation changes and hydrological fluctuations in the Central Ebro Basin (NE Spain) since the Late Glacial period: saline lake records. Palaeogeogr Palaeoclimatol Palaeoecol 259:157–181
Grenfell S, Grenfell M, Rowntree K, Ellery W (2014) Fluvial connectivity and climate: a comparison of channel pattern and process in two climatically contrasting fluvial sedimentary systems in South Africa. Geomorphology 205:142–154
Gutiérrez M, Desir G, Gutiérrez F, Sánchez JA, Castañeda C, Lucha P (2005) Aeolian Landforms and Saline Lakes (Central Ebro Basin). In: Sixth international conference on geomorphology, field trip guide B-7
Gutiérrez F, Valero-Garcés BL, Desir G, González-Sampériz P, Gutiérrez M, Linares R, Zarroca M, Moreno A, Guerrero J, Roqué C, Arnold LJ, Demuro M (2013) Late Holocene evolution of playa lakes in the central Ebro depression based on geophysical surveys and morpho-stratigraphic analysis of lacustrine terraces. Geomorphology 196:177–197
Herrero J, Snyder RL (1997) Aridity and irrigation in Aragon, Spain. J Arid Environ 35(3):535–547
Huang J, Yu H, Guan X, Wang G, Guo R (2016) Accelerated dryland expansion under climate change. Nat Clim Change 6(2):166–171
Huggett RJ (1998) Soil chronosequences, soil development, and soil evolution: a critical review. Catena 32:155–172
Ibáñez MJ (1973) Contribución al estudio del endorreísmo de la Depresión del Ebro: el foco endorreico al W y SW de Alcañiz (Teruel). Geográfica 1:21–32
Ibáñez MJ (1976) El Piedemonte Ibérico bajoaragonés. Estudio Geomorfológico. Instituto de Geografía Aplicada, C.S.I.C., 523
Ibell PT, Xu Z, Blumfield TJ (2013) The influence of weed control on foliar δ15N, δ13C and tree growth in an 8 year-old exotic pine plantation of subtropical Australia. Plant Soil 369(1):199–217
IPCC (2014a) Summary for Policymakers. In: Edenhofer et al. (eds) Climate change 2014, Mitigation of climate change. Contribution of working group III to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge
IPCC (2014b) 2013 supplement to the 2006 IPCC guidelines for national greenhouse gas inventories: wetlands. In: Hiraishi T, Krug T, Tanabe K, Srivastava N, Baasansuren J, Fukuda M, Troxler TG (eds). Published: IPCC
Kutzbach L, Schneider J, Sachs T, Giebels M, Nykänen H, Shurpali NJ, Martikainen PJ, Alm J, Wilmking M (2007) CO2 flux determination by closed-chamber methods can be seriously biased by inappropriate application of linear regression. Biogeosciences 4:2279–2328
Ladrón de Guevara M, Lázaro R, Quero JL, Ochoa V, Gozalo B, Berdugo M, Uclés O, Escolar C, Maestre FT (2014) Simulated climate change reduced the capacity of lichen-dominated biocrusts to act as carbon sinks in two semi-arid Mediterranean ecosystems. Biodivers Conserv 23:1787–1807
Lan S, Thomas AD, Rakes JB, Garcia-Pichel F, Wu L, Hu C (2021) Cyanobacterial community composition and their functional shifts associated with biocrust succession in the Gurbantunggut Desert. Environ Microbiol Rep 13(6):884–898
Larkin ZT, Ralph TJ, Tooth S, McCarthy TS (2017) The interplay between extrinsic and intrinsic controls in determining floodplain wetland characteristics in the South African drylands. Earth Surf Proc Land 42:1092–1109
Larkin ZT, Ralph TJ, Tooth S, Fryirs KA, Carthey AJR (2020) Identifying threshold responses of Australian dryland rivers to future global hydroclimatic change. Scientific Reports, 10, article no. 6653
Li J, Yang X, Maffei C, Tooth S, Yao G (2018) Applying independent component analysis on Sentinel-2 imagery to characterize geomorphological responses to an extreme flood event near the non-vegetated Río Colorado terminus, Salar de Uyuni, Bolivia. Remote Sensing, 10(5), article no. 725
Li J, Tooth S, Zhang K, Zhao Y (2021) Visualisation of flooding along an unvegetated, ephemeral river using Google Earth Engine: implications for assessment of channel-floodplain dynamics in a time of rapid environmental change. J Environ Manag 278, article no. 111559
Li Y, Zhang C, Wang N, Han Q, Zhang X, Liu Y, Xu L, Ye W (2017) Substantial inorganic carbon sink in closed drainage basins globally. Nat Geosci 10:501–506
Lin L, Gettelman A, Feng S, Fu Q (2015) Simulated climatology and evolution of aridity in the 21st century. J Geophys Res Atmos 120(12):5795–5815
Macklin MG, Passmore DG, Stevenson AC, Davis BA, Benavente JA (1994) Responses of rivers and lakes to Holocene environmental change in the Alcañiz region, Teruel, north-east Spain. In: Millington AC, Pye K (eds) Environmental change in drylands: biogeographical and geomorphological perspectives. Wiley, Chichester, pp 113–130
Magoč T, Salzberg SL (2011) FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics 27(21):2957–2963
Maier S, Tamm A, Wu D, Caesar J, Grube M, Weber B (2018) Photoautotrophic organisms control microbial abundance, diversity, and physiology in different types of biological soil crusts. ISME J 12:1032–1046
McKay CP, Rask JC, Detweiler AM, Bebout BM, Everroad RC, Lee JZ, Chanton JP, Mayer MH, Caraballo AAL, Kapili B, Al-Awar M, Al-Farraj A (2016) An unusual inverted saline microbial mat community in an interdune sabkha in the Rub’ al Khali (the Empty Quarter) United Arab Emirates. PLoS ONE 11(3):e0150342
Mees F, Castañeda C, Van Ranst E (2011) Sedimentary and diagenetic features in saline lake deposits of the Monegros region, northern Spain. Catena 85:245–252
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: wetlands and water synthesis. World Resources Institute, Washington, DC
Mitsch WJ, Gosselink JG (2015) Wetlands, 5th edn. Wiley, New York
Mitsch WJ, Nahlik A, Wolski P, Bernal B, Zhang L, Ramberg L (2010) Tropical wetlands: seasonal hydrologic pulsing, carbon sequestration, and methane emissions. Wetlands Ecol Manage 18:573–586
Moyano FE, Manzoni S, Chenu C (2013) Responses of soil heterotrophic respiration to moisture availability: an exploration of processes and models. Soil Biol Biochem 59:72–85
Nahlik A, Fennessy M (2016) Carbon storage in US wetlands. Nat Commun 7:13835
Nazaries L, Murrell JC, Milard P, Baggs L, Singh BK (2013) Methane, microbes and models: fundamental understanding of the soil methane cycle for future predictions. Environ Microbiol 15(9):2395–2417
Orchard VA, Cook FJ (1983) Relationship between soil respiration and soil moisture. Soil Biol Biochem 15(4):447–453
Petrescu AM, Lohila A, Tuovinen JP, Baldocchi D, Desai A, Roulet N et al (2015) The uncertain climate footprint of wetlands under human pressure. Proc Natl Acad Sci USA 112:4594–4599
Pueyo JJ (1980) Procesos diagenéticos observados en las lagunas tipo playa de la zona de Bujaraloz-Sástago (provincias de Zaragoza y Teruel). Revista Del Instituto De Investigaciones Geológicas 34:195–207
Ramsar Convention (2018) Global wetland outlook: state of the world's wetlands and their services to people 2018. Gland: Ramsar Convention Secretariat. https://www.global-wetland-outlook.ramsar.org
Rey A, Oyonarte C, Morán-López T, Raimundo J, Pegoraro E (2017) Changes in soil moisture predict soil carbon losses upon rewetting in a perennial semiarid steppe in SE Spain. Geoderma 287:135–146
Rodrigo RM, Alastrué IM (no date) Guía de La Laguna de Sariñena: Refugio de Fauna Silvestre y Zepa (Zóna de Especial Protecctión Para Las Aves), Ayuntamiento de Sariñena, Diputación Provincial de Huesca
Rogers K, Kelleway JJ, Saintilan N, Megonigal JP, Adams JB, Holmquist JR, Lu M, Schile-Beers L, Zawadzki A, Mazumder D, Woodroffe CD (2019) Wetland carbon storage controlled by millennial-scale variation in relative sea-level rise. Nature 567:91–95
Samper-Calvete FJ, García-Vera MA (1998) Inverse modeling of groundwater flow in the semiarid evaporitic closed basin of Los Monegros, Spain. Hydrogeol J 6:33–49
Sánchez JA, Pérez A, Martínez-Gil J (1998) Combined effects of groundwater and aeolian processes in the formation of the northernmost closed saline depressions of Europe: north-east Spain. Hydrol Process 12:813–820
Sandi SG, Rodriguez JF, Saintilan N, Wen L, Kuczera G, Riccardi G, Saco PM (2020) Resilience to drought of dryland wetlands threatened by climate change. Sci Rep 10:13232
Schimel JP, Gulledge JM, Clein-Curley JS, Lindstrom JE, Braddock JF (1999) Moisture effects on microbial activity and community structure in decomposing birch litter in the Alaskan taiga. Soil Biol Biochem 31:831–883
Soil Survey Staff (2010) Keys to Soil Taxonomy, 11th edn. USDA, National Resources Conservation Service, National Soil Survey Center, Lincoln
Swenson TL, Karaoz U, Swenson JM, Bowen BP, Northern TR (2018) Linking soil biology and chemistry in biological soil crust using isolate exometabolomics. Nat Commun 9:19
Tooth S (2018) The geomorphology of wetlands in drylands: resilience, nonresilience, or …? Geomorphology 305:33–48
Tooth S, McCarthy TS (2007) Wetlands in drylands: geomorphological and sedimentological characteristics, with emphasis on examples from southern Africa. Prog Phys Geogr 31(1):3–41
Tooth S, Grenfell M, Thomas AD, Ellery W (2015) Wetlands in drylands: ‘hotspots’ of ecosystem service provision in marginal environments. Science Brief for the United Nation’s Global Sustainable Development Report, 2015 Edition, https://sustainabledevelopment.un.org/content/documents/640493-Tooth-Wetlands%20in%20Drylands_Hotspots%20of%20Ecosystem%20Services%20in%20Marginal%20Environments.pdf
Thomas AD (2012) Impact of grazing intensity on seasonal variations of soil organic carbon and soil CO2 efflux in two semi-arid grasslands in southern Botswana. Philos Trans R Soc B 367:3076–3086
Thomas AD, Dougill AJ, Elliott D, Mairs H (2014) Seasonal differences in CO2 efflux and C storage in salt pan sediments on Ntwetwe Pan, Makgadikgadi, Botswana. Geoderma 219–220:72–81
Thomas AD, Elliott DR, Dougill AJ, Stringer LC, Hoon SR, Sen R (2018) The influence of trees, shrubs and grasses on micro-climate, soil C, N and CO2 efflux: potential implications of shrub encroachment for Kalahari rangelands. Land Degrad Dev 29:1–11
Thomas AD, Elliott DR, Hardcastle D, Strong CL, Bullard J, Webster R, Lan S (2022) Soil biocrusts affect metabolic response to hydration on dunes in west Queensland Australia. Geoderma 405:115464
Thompson LG, Mosley-Thompson E, Brecher H, Davis M, Leon B, Les D, Lin PN, Mashiotta T, Mountain K (2006) Abrupt tropical climate change: past and present. Proc Natl Acad Sci 103(28):10536–10543
Valero-Garcés BL, Navas A, Machín J, Stevenson T, Davis B (2000a) Responses of a saline lake ecosystem in a semiarid region to irrigation and climate variability: the history of Salada Chiprana, central Ebro Basin, Spain. Ambio 29(6):344–350
Valero-Garcés BL, Delgado-Huertas A, Navas A, Machín J, González-Samériz P, Kelts K (2000b) Quaternary palaeohydrological evolution of a playa lake: Salada Mediana, central Ebro Basin, Spain. Sedimentology 47:1135–1156
Vikram S, Guerrero LD, Makhalanyane TP, Le PT, Seely M, Cowan DA (2015) Metagenomic analysis provides insights into functional capacity in a hyperarid desert soil niche community. Environ Microbiol 18:1875–1888
Vizcayno C, Garcia-Gonzalez MT, Gutierrez M, Rodriguez R (1995) Mineralogical, chemical and morphological features of salt accumulations in the Flumen-Monegros district NE, Spain. Geoderma 68(3):193–210
Walker PH, Coventry RJ (1976) Soil profile development in some alluvial deposits of eastern New South Wales. Soil Res 14:305–317
Watanabe T, Wang G, Lee CG, Murase J, Asakawa S, Kimura M (2011) Assimilation of glucose-derived carbon into methanogenic archaea in soil under unflooded condition. Appl Soil Ecol 48(2):201–209
Were D, Kansiime F, Fetahi T, Cooper A, Jjuuko C (2019) Carbon sequestration by wetlands: a critical review of enhancement measures for climate change mitigation. Earth Syst Environ 3:327–340
Williams WJ, Büdel B, Reichenberger H, Rose N (2014) Cyanobacteria in the Australian northern savannah detect the difference between intermittent dry season and wet season rain. Biodivers Conserv 23(7):1827–1844
Xi Y, Peng S, Ciais P, Chen Y (2021) Future impacts of climate change on inland Ramsar wetlands. Nat Clim Chang 11:45–51
Xie J, Li Y, Zhai C, Li C, Lan Z (2009) CO2 absorption by alkaline soils and its implication to the global carbon cycle. Environ Geol 56:953–961
Xu HQ (2007) Modification of normalised difference water index (NDWI) to enhance open water features in remotely sensed imagery. Int J Remote Sens 27:3025–3033
Yates EL, Detweiler AM, Iraci LT, Bebout BM, McKay CP, Schiro K, Sheffner EJ, Kelley CA, Tadić TM, Loewenstein M (2013) Assessing the role of alkaline soils on the carbon cycle at a playa site. Environ Earth Sci 70:1047–1056
Ye F, Hu C, Wang Y, Wu J, Hong Y (2021) Biogeographic pattern of methanogenic community in surface water along the Yangtze River. Geomicrobiology 38(4):1–10
Yeh HW, Wang WM (2001) Factors affecting the isotopic composition of organic matter. (1) Carbon isotopic composition of terrestrial plant materials. Proc Natl Sci Counc Repub China B 25:137–147
Yuan X, Zhang X, Chen X, Kong D, Liu X, Shen S (2018) Synergistic degradation of crude oil by indigenous bacterial consortium and exogenous fungus Scedosporium boydii. Bioresour Technol 64:190–197
Zhang Y, Cong J, Lu H, Li G, Xue Y, Deng Y, Li H, Zhou J, Li D (2015) Soil bacterial diversity patterns and drivers along an elevational gradient on Shennongjia Mountain, China. Microb Biotechnol 8:739–746
Zhang Q, Wang Q, Ouyang H, Lan S, Hu C (2018) Pyrosequencing reveals significant changes in microbial communities along the ecological successions of biological soil crusts in Tengger Desert of China. Pedosphere 28:350–362
Zhang H, Tang Z, Bishop A, Drahota J, LaGrange T, Varner D (2020) Conservation significantly improves wetland conditions: evaluation of playa wetlands in different conservation status. Wetlands Ecol Manag 28:85–102
Zhao X, Zhao C, Wang J, Stahr K, Kuzyakov Y (2016) CaCO3 recrystallization in saline and alkaline soils. Geoderma 282(2016):1–8
Acknowledgements
The authors report no financial conflicts or conflicts of interests. The microbial data in the paper have been deposited to NCBI Sequence Read Archive (SRA) database under the accession numbers PRJNA683155, PRJNA683730, and PRJNA684402. The soil chemical and CO2 efflux data have been deposited at https://doi.org/10.6084/m9.figshare.13395959.
Funding
The research was funded through the European Union's Horizon 2020 Research and Innovation Programme under a Marie Skłodowska-Curie Grant (No. 663830). Tarren is grateful for the financial support provided by the Department of Geography and Earth Sciences’s Postgraduate Research Fund. The paper was prepared while Shubin Lan was a Sêr Cymru II Fellow at Aberystwyth University. The authors are grateful for input from two anonymous reviewers.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Thomas, A.D., Tooth, S., Lan, S. et al. Soil properties across a hydrological gradient in saladas from northeast Spain: what are the implications for soil carbon stocks, CO2 efflux and microbial communities in a warming world?. Wetlands Ecol Manage 30, 715–735 (2022). https://doi.org/10.1007/s11273-022-09856-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-022-09856-6