
Abstract
LGR4/5 receptors and their cognate RSPO ligands potentiate Wnt/β-catenin signalling and promote proliferation and tissue homeostasis in epithelial stem cell compartments. In the liver, metabolic zonation requires a Wnt/β-catenin signalling gradient, but the instructive mechanism controlling its spatiotemporal regulation is not known. We have now identified the RSPO–LGR4/5–ZNRF3/RNF43 module as a master regulator of Wnt/β-catenin-mediated metabolic liver zonation. Liver-specific LGR4/5 loss of function (LOF) or RSPO blockade disrupted hepatic Wnt/β-catenin signalling and zonation. Conversely, pathway activation in ZNRF3/RNF43 LOF mice or with recombinant RSPO1 protein expanded the hepatic Wnt/β-catenin signalling gradient in a reversible and LGR4/5-dependent manner. Recombinant RSPO1 protein increased liver size and improved liver regeneration, whereas LGR4/5 LOF caused the opposite effects, resulting in hypoplastic livers. Furthermore, we show that LGR4+ hepatocytes throughout the lobule contribute to liver homeostasis without zonal dominance. Taken together, our results indicate that the RSPO–LGR4/5–ZNRF3/RNF43 module controls metabolic liver zonation and is a hepatic growth/size rheostat during development, homeostasis and regeneration.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Accession codes
Change history
27 September 2016
In the version of this Article originally published, the authors inadvertently omitted a key reference. The following reference has been added: '48. Rocha, A. S. et al. The angiocrine factor Rspondin3 is a key determinant of liver zonation. Cell Rep. 13, 1757–1764 (2015).' References 48–56 have been renumbered accordingly. Furthermore, a new sentence citing this reference has been added at the end of the first paragraph in the Discussion: 'A recent report that was published during the final revision of our manuscript suggests a role for pericentrally confined RSPO3 in metabolic liver zonation48.' These changes have been made in the online versions of the Article.
References
Gebhardt, R. & Hovhannisyan, A. Organ patterning in the adult stage: the role of Wnt/β-catenin signaling in liver zonation and beyond. Dev. Dyn. 239, 45–55 (2010).
Jungermann, K. & Katz, N. Functional specialization of different hepatocyte populations. Phys. Rev. 69, 708–764 (1989).
Jungermann, K. & Kietzmann, T. Zonation of parenchymal and nonparenchymal metabolism in liver. Annu. Rev. Nutr. 16, 179–203 (1996).
Monga, S. P. Role and regulation of β-catenin signaling during physiological liver growth. Gene Expr. 16, 51–62 (2014).
Torre, C., Perret, C. & Colnot, S. Transcription dynamics in a physiological process: β-catenin signaling directs liver metabolic zonation. Int. J. Biochem. Cell Biol. 43, 271–278 (2011).
Michalopoulos, G. K. & DeFrances, M. C. Liver regeneration. Science 276, 60–66 (1997).
Zaret, K. S. Regulatory phases of early liver development: paradigms of organogenesis. Nat. Rev. Genet. 3, 499–512 (2002).
Si-Tayeb, K., Lemaigre, F. P. & Duncan, S. A. Organogenesis and development of the liver. Dev. Cell 18, 175–189 (2010).
Apte, U. et al. β-catenin activation promotes liver regeneration after acetaminophen-induced injury. Am. J. Pathol. 175, 1056–1065 (2009).
Benhamouche, S. et al. Apc tumor suppressor gene is the ‘zonation-keeper’ of mouse liver. Dev. Cell 10, 759–770 (2006).
Boulter, L. et al. Macrophage-derived Wnt opposes Notch signaling to specify hepatic progenitor cell fate in chronic liver disease. Nat. Med. 18, 572–579 (2012).
Burke, Z. D. et al. Liver zonation occurs through a β-catenin-dependent, c-Myc-independent mechanism. Gastroenterology 136, 2316–2324 (2009).
Monga, S. P., Pediaditakis, P., Mule, K., Stolz, D. B. & Michalopoulos, G. K. Changes in WNT/β-catenin pathway during regulated growth in rat liver regeneration. Hepatology 33, 1098–1109 (2001).
Nejak-Bowen, K. & Monga, S. P. Wnt/β-catenin signaling in hepatic organogenesis. Organogenesis 4, 92–99 (2008).
Nejak-Bowen, K. N. et al. Accelerated liver regeneration and hepatocarcinogenesis in mice overexpressing serine-45 mutant β-catenin. Hepatology 51, 1603–1613 (2010).
Sekine, S., Gutierrez, P. J., Lan, B. Y., Feng, S. & Hebrok, M. Liver-specific loss of β-catenin results in delayed hepatocyte proliferation after partial hepatectomy. Hepatology 45, 361–368 (2007).
Sekine, S., Lan, B. Y., Bedolli, M., Feng, S. & Hebrok, M. Liver-specific loss of β-catenin blocks glutamine synthesis pathway activity and cytochrome p450 expression in mice. Hepatology 43, 817–825 (2006).
Yang, J. et al. β-catenin signaling in murine liver zonation and regeneration: a Wnt-Wnt situation. Hepatology 60, 964–976 (2014).
Niehrs, C. The complex world of WNT receptor signalling. Nat. Rev. Mol. Cell Biol. 13, 767–779 (2012).
Carmon, K. S., Gong, X., Lin, Q., Thomas, A. & Liu, Q. R-spondins function as ligands of the orphan receptors LGR4 and LGR5 to regulate Wnt/β-catenin signaling. Proc. Natl Acad. Sci. USA 108, 11452–11457 (2011).
de Lau, W. et al. Lgr5 homologues associate with Wnt receptors and mediate R-spondin signalling. Nature 476, 293–297 (2011).
Glinka, A. et al. LGR4 and LGR5 are R-spondin receptors mediating Wnt/β-catenin and Wnt/PCP signalling. EMBO Rep. 12, 1055–1061 (2011).
Ruffner, H. et al. R-Spondin potentiates Wnt/β-catenin signaling through orphan receptors LGR4 and LGR5. PLoS ONE 7, e40976 (2012).
Hao, H. X. et al. ZNRF3 promotes Wnt receptor turnover in an R-spondin-sensitive manner. Nature 485, 195–200 (2012).
Koo, B. K. et al. Tumour suppressor RNF43 is a stem-cell E3 ligase that induces endocytosis of Wnt receptors. Nature 488, 665–669 (2012).
Kinzel, B. et al. Functional roles of Lgr4 and Lgr5 in embryonic gut, kidney and skin development in mice. Dev. Biol. 390, 181–190 (2014).
Koo, B. K. & Clevers, H. Stem cells marked by the R-spondin receptor LGR5. Gastroenterology 147, 289–302 (2014).
Bird, T. G. & Forbes, S. J. Two fresh streams to fill the liver’s hepatocyte pool. Cell Stem Cell 17, 377–378 (2015).
Font-Burgada, J. et al. Hybrid periportal hepatocytes regenerate the injured liver without giving rise to cancer. Cell 162, 766–779 (2015).
Lu, W. Y. et al. Hepatic progenitor cells of biliary origin with liver repopulation capacity. Nat. Cell Biol. 17, 971–983 (2015).
Wang, B., Zhao, L., Fish, M., Logan, C. Y. & Nusse, R. Self-renewing diploid Axin2(+) cells fuel homeostatic renewal of the liver. Nature 524, 180–185 (2015).
Zaret, K. S. Regenerative biology: maintaining liver mass. Nature 524, 165–166 (2015).
Huch, M. et al. In vitro expansion of single Lgr5+ liver stem cells induced by Wnt-driven regeneration. Nature 494, 247–250 (2013).
Karaca, G. et al. TWEAK/Fn14 signaling is required for liver regeneration after partial hepatectomy in mice. PLoS ONE 9, e83987 (2014).
Colnot, S. & Perret, C. Molecular Pathology of Liver Diseases Vol. Molecular Pathology Library 5. (ed. S. P. Monga) (Springer Science and Business Media, 2011).
Ferrer-Vaquer, A. et al. A sensitive and bright single-cell resolution live imaging reporter of Wnt/ß-catenin signaling in the mouse. BMC Dev. Biol. 10, 121 (2010).
Gerbal-Chaloin, S. et al. The WNT/β-catenin pathway is a transcriptional regulator of CYP2E1, CYP1A2, and aryl hydrocarbon receptor gene expression in primary human hepatocytes. Mol. Pharmacol. 86, 624–634 (2014).
Soriano, P. Generalized lacZ expression with the ROSA26 Cre reporter strain. Nat. Genet. 21, 70–71 (1999).
Barker, N. et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 449, 1003–1007 (2007).
Clevers, H., Loh, K. M. & Nusse, R. Stem cell signaling. An integral program for tissue renewal and regeneration: Wnt signaling and stem cell control. Science 346, 1248012 (2014).
Snippert, H. J. et al. Lgr6 marks stem cells in the hair follicle that generate all cell lineages of the skin. Science 327, 1385–1389 (2010).
Klochendler, A. et al. A transgenic mouse marking live replicating cells reveals in vivo transcriptional program of proliferation. Dev. Cell 23, 681–690 (2012).
Miyajima, A., Tanaka, M. & Itoh, T. Stem/progenitor cells in liver development, homeostasis, regeneration, and reprogramming. Cell Stem Cell 14, 561–574 (2014).
Gougelet, A. et al. T-cell factor 4 and β-catenin chromatin occupancies pattern zonal liver metabolism in mice. Hepatology 59, 2344–2357 (2014).
Clevers, H. Wnt/β-catenin signaling in development and disease. Cell 127, 469–480 (2006).
Fitamant, J. et al. YAP inhibition restores hepatocyte differentiation in advanced HCC, leading to tumor regression. Cell Rep. 10, 1692–1707 (2015).
Marti, P. et al. YAP promotes proliferation, chemoresistance, and angiogenesis in human cholangiocarcinoma through TEAD transcription factors. Hepatology 62, 1497–1510 (2015).
Rocha, A. S. et al. The angiocrine factor Rspondin3 is a key determinant of liver zonation. Cell Rep. 13, 1757–1764 (2015).
Bralet, M. P., Branchereau, S., Brechot, C. & Ferry, N. Cell lineage study in the liver using retroviral mediated gene transfer. Evidence against the streaming of hepatocytes in normal liver. Am. J. Pathol. 144, 896–905 (1994).
Ong, C. K. et al. Exome sequencing of liver fluke-associated cholangiocarcinoma. Nat. Genet. 44, 690–693 (2012).
Tchorz, J. S. et al. A modified RMCE-compatible Rosa26 locus for the expression of transgenes from exogenous promoters. PLoS ONE 7, e30011 (2012).
Postic, C. et al. Dual roles for glucokinase in glucose homeostasis as determined by liver and pancreatic beta cell-specific gene knock-outs using Cre recombinase. J. Biol. Chem. 274, 305–315 (1999).
Mitchell, C. & Willenbring, H. A reproducible and well-tolerated method for 2/3 partial hepatectomy in mice. Nat. Protoc. 3, 1167–1170 (2008).
Dill, M. T. et al. Constitutive Notch2 signaling induces hepatic tumors in mice. Hepatology 57, 1607–1619 (2013).
Wieland, S. et al. Simultaneous detection of hepatitis C virus and interferon stimulated gene expression in infected human liver. Hepatology 59, 2121–2130 (2014).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Wu, D. & Smyth, G. K. Camera: a competitive gene set test accounting for inter-gene correlation. Nucleic Acids Res. 40, e133 (2012).
Acknowledgements
We thank J. Wirsching, C. Manneville, M. Lemaistre, M. Haffner, B. Leonhard, S. Ley, N. Guo, T. Lewis, M. Li, M. van de Velde, D. Breustedt, H. Lei, R. Boomer, S. Zurbruegg, L. Perrot, F. Cordoba, S. Schuierer, A. Arif and J. Gfeller for technical assistance. For helpful discussion and critical reading of the manuscript, we thank E. Wiellette, G. Hintzen, B. Dietrich, D. Liu, F. Bassilana, C. Parker, K. Seuwen and J. Porter. This work was supported by the Leverhulme Trust ECF-2012-262 to L.B., SNF grant 310030B_147089 to M.H.H., MRC Centre Grant and UKRMP Niche hub grant MR/K026666/1 to S.J.F. and the Novartis Institutes for BioMedical Research Postdoctoral Program.
Author information
Authors and Affiliations
Contributions
Experimental work and data analysis: L.P.-P., V.O., L.B., D.C., M.P., Y.X., A.D., P.M., N.B., M.T.D., W.C., R.Z. and P.C.; data analysis: F.N., G.R., S.B., R.V., L.M.T. and J.S.T.; mouse model generation: A.I., M.M., B.K., Y.Y., X.M. and T.B.N.; RSPO1 production: D.R. and A.L.; concept and design, and manuscript writing: L.P.-P., L.B., C.U., T.B., F.C., M.H.H., S.J.F., H.R. and J.S.T.
Corresponding author
Ethics declarations
Competing interests
All authors except S.J.F., L.B., L.M.T., M.H.H., D.C. and M.T.D. are employed by and/or shareholders of Novartis Pharma AG.
Integrated supplementary information
Supplementary Figure 1 LGR4 and LGR5 expression and mouse model generation.
(a) Lgr4 and Lgr5 ISH in liver parenchyma of control mice. The images are representative for 4 mice each. (b) Percent of hepatocytes expressing Lgr4 and Lgr5 in the indicated liver zones. n = 4 mice. These data involved assessment of 3346 cells from the indicated mice. (c) Scheme depicting the generation of Rnf43-LacZ mice. (d) Scheme depicting the generation of liver-specific Lgr4/5dLKO mice by crossing with AlbCre mice. Data represent mean ± s.d. Scale bars, (a) 20 μm and (magnification in a) 10 μm.
Supplementary Figure 2 LGR4 and LGR5 lineage tracing.
(a) Scheme depicting lineage tracing in Lgr4ki/R26-LacZ and Lgr5ki/R26-LacZ mice. (b) LacZ staining in Lgr4ki/R26-LacZ mice showing LGR4 + hepatocytes in the liver (magnified area: parenchyma) after 10 days or 10 months of tracing. (c) LacZ staining in Lgr5ki/R26-LacZ mice showing no LGR5 + hepatocytes in the liver (magnified area: parenchyma) after 10 days or 18 months of tracing. (d) Distribution of Lgr5ki/R26-LacZ + hepatocytes after 10 days or 18 months of tracing. n = 3 mice per group. These data involved assessment of 210 cells (10 days) and 330 cells (18 months) from the indicated mice. (e) Axin2 ISH and β-Gal staining in Lgr5ki/R26-LacZ mice after 10 days of tracing showing Axin2 + /LacZ + hepatocytes. (f) LacZ staining, Axin2 and Lgr5 co-ISH in consecutive liver sections of Lgr5ki/R26-LacZ mice after 10 days of tracing. (g) Percent of hepatocytes expressing Axin2 and Lgr5 in Lgr5ki/R26-LacZ + and Lgr5ki/R26-LacZ- hepatocytes. n = 4 mice. (h) Scheme depicting lineage tracing in Lgr5ki/tdTOM mice 2 days post-PH. (i) tdTOM and Ki67 staining in consecutive liver sections of Lgr5ki/R26-tdTOM mice 2 days post-PH. Arrowheads point at tdTOM + /Ki67 + hepatocytes. (j) tdTOM + /Ki67 + hepatocytes quantified in Lgr5ki/R26-tdTOM mice 2 days post-PH. n = 4 mice. (k) Scheme depicting EdU injections in WT mice. (l) GS and EdU co-staining in control mice. Arrowheads point at EdU + hepatocytes. (m) EdU + hepatocytes quantified in liver zones of control mice. n = 5 mice. (n) Distribution of EdU + hepatocytes in the indicated liver zones. n = 5 mice. These data involved assessment of (d) 210 cells (10 days) and 330 cells (18 months); (g) 1970 LacZ- and 87 LacZ + cells; (j) 14681 tdTOM- and 49 tdTOM + cells; (m) 2650 cells; and (n) 577 cells from the indicated mice. CV, central vein; PV, portal vein. The images in (b,c) and (e,f,i,l) are representative for 3 and 4 mice each, respectively. Data represent mean ± s.d. ns, not significant; two-tailed unpaired t-test (j) and one-way ANOVA with Tukey’s test (m) were used. Scale bars, (b,c,f) 100 μm, (magnifications in b,c) 50 μm, (e,l, magnification in f) 20 μm and (i) 50 μm.
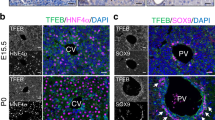
Supplementary Figure 3 Role of LGR4 and LGR5 during hepatocyte maturation and differentiation.
(a) GS and CK19 co-staining in P2, P10 and P30 control and Lgr4/5dLKO mice. (b) GS + staining quantified in the indicated mice. n = 4 mice (P10 control and P30 control) and 5 mice (all other groups). (c) HNF4α and CK19 co-staining in P2, P10 and P30 control and Lgr4/5dLKO mice. (d) SOX9 staining in P2, P10 and P30 control and Lgr4/5dLKO mice. CV, central vein; PV, portal vein. The images in (a,c,d) are representative for 29 mice, and the stainings were repeated 2 times. Data represent mean ± s.d. ∗∗, P < 0.01; ∗∗∗, P < 0.001; ∗∗∗∗, P < 0.0001; two-tailed unpaired t-test (b) was used. Scale bars, 20 μm.
Supplementary Figure 4 Lgr4 and Lgr5 control liver zonation.
(a) Volcano plot showing pericentral genes (in green), periportal genes (in blue) and genes involved in Wnt signalling (in purple) that are differentially expressed in livers of the indicated mice. n = 5 mice per group. (b) Gene sets downregulated in livers of Lgr4/5dLKO mice compared to control mice. n = 5 mice per group. (c,d) ARG1 (c) and PCK1 (d) staining in the indicated mice. CV, central vein; PV, portal vein. The images in (c,d) are representative for 10 mice, and the stainings were repeated 3 times. Empirical Bayes with Benjamini–Hochberg test (a), and weighted Kolmogorov–Smirnov and Mann–Whitney U-test with Benjamini-Hochberg test (b) were used. Scale bars, 100 μm.
Supplementary Figure 5 Role of LGR4 and LGR5 in the control of liver growth.
(a,b,c) GS, CK19 and Ki67 co-staining in P2 (a), P10 (b) and P30 (c) control and Lgr4/5dLKO mice, showing proliferating hepatocytes (arrowheads). Bottom: Hepatocyte proliferation quantified in liver zones of the indicated mice. n = 4 mice (P30 controls), 5 mice (P2 and P10), and 6 mice (P30 Lgr4/5dLKO). These data involved assessment of 2582 cells (P2 control), 2705 cells (P2 Lgr4/5dLKO), 4217 cells (P10 control), 3911 cells (P10 Lgr4/5dLKO), 2882 cells (P30 control) and 3171 cells (P30 Lgr4/5dLKO) from the indicated mice. (d) Relative liver weight of the indicated mice. n = 5 mice (P2 control) and 6 mice (all other groups). (e) Ki67 staining of the indicated mice at embryonic day (E)16.5. (f) Liver cell proliferation quantified in the indicated mice at E16.5. n = 5 mice (control, Lgr5LKO and Lgr4/5dLKO) and 6 mice (Lgr4LKO). CV, central vein; PV, portal vein. The images in (a–c) and (e) are representative for 30 and 21 mice, respectively, and the stainings were repeated 2 times. Data represent mean ± s.d. ∗, P < 0.05; ∗∗, P < 0.01; ∗∗∗, P < 0.001; ns, not significant; two-tailed unpaired t-test (a–d, f) was used. Scale bars, 50 μm.
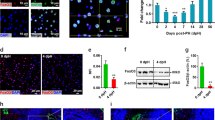
Supplementary Figure 6 Role of LGR4 and LGR5 during liver regeneration.
(a) Axin2 ISH in wild type mice +/− PH. (b) Axin2 ISH quantified in wild type mice +/− PH in the indicated liver zones. n = 5 mice (WT) and 3 mice (WT d2 post-PH). These data involved assessment of 4225 cells (WT) and 1905 cells (WT d2 post-PH) from the indicated mice. (c) Volcano plot showing genes differentially expressed in livers of the indicated mice. n = 5 mice per group. (d) Lgr4 and Lgr5 ISH in wild type mice +/− PH. (e,f) Lgr4 (e) and Lgr5 (f) ISH quantified in wild type mice +/− PH in the indicated liver zones. n = 5 mice (WT, Lgr5), 4 mice (WT, Lgr4) and 3 mice (WT d2 post-PH). (g) Volcano plot showing genes differentially expressed in livers of the indicated mice. n = 5 mice per group. (h) Scheme depicting lineage tracing in Lgr5ki/R26-LacZ mice 7 days post-PH. (i) LacZ staining showing that Lgr5 + hepatocytes (arrowheads) did not overtly proliferate during liver regeneration following PH. CV, central vein; PV, portal vein. The images in (a,d) and (i) are representative for 8 and 6 mice, respectively. Data represent mean ± s.d. ∗, P < 0.05; ∗∗∗∗, P < 0.0001; two-way ANOVA with Sidak’s test (b,e,f) and Empirical Bayes with Benjamini–Hochberg test (c,g) were used. Scale bars, (a,d) 10 μm and (i) 100 μm. Statistics source data for (b,e,f) can be found in Supplementary Table 4.
Supplementary information
Supplementary Information
Supplementary Information (PDF 1394 kb)
Supplementary Table 1
Supplementary Information (XLSX 47 kb)
Supplementary Table 2
Supplementary Information (XLSX 71 kb)
Supplementary Table 3
Supplementary Information (XLSX 11 kb)
Supplementary Table 4
Supplementary Information (XLSX 16 kb)
Rights and permissions
About this article

Cite this article
Planas-Paz, L., Orsini, V., Boulter, L. et al. The RSPO–LGR4/5–ZNRF3/RNF43 module controls liver zonation and size. Nat Cell Biol 18, 467–479 (2016). https://doi.org/10.1038/ncb3337
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ncb3337
This article is cited by
-
Liver organoid culture methods
Cell & Bioscience (2023)
-
Hepatocyte generation in liver homeostasis, repair, and regeneration
Cell Regeneration (2022)
-
Integrative network analysis of circular RNAs reveals regulatory mechanisms for hepatic specification of human iPSC-derived endoderm
Stem Cell Research & Therapy (2022)
-
Cell adhesion molecule KIRREL1 is a feedback regulator of Hippo signaling recruiting SAV1 to cell-cell contact sites
Nature Communications (2022)
-
RNF43/ZNRF3 loss predisposes to hepatocellular-carcinoma by impairing liver regeneration and altering the liver lipid metabolic ground-state
Nature Communications (2022)
