
Abstract
Biological control against malaria and its transmission is currently a considerable challenge. Plant-associated bacteria of the genus Asaia are frequently found in nectarivorous arthropods, they thought to have a natural indirect action on the development of plasmodium in mosquitoes. However, virtually nothing is known about its natural cycle. Here, we show the role of nectar-producing plants in the hosting and dissemination of Asaia. We isolated Asaia strains from wild mosquitoes and flowers in Senegal and demonstrated the transmission of the bacteria from infected mosquitoes to sterile flowers and then to 26.6% of noninfected mosquitoes through nectar feeding. Thus, nectar-producing plants may naturally acquire Asaia and then colonize Anopheles mosquitoes through food-borne contamination. Finally, Asaia may play an indirect role in the reduction in the vectorial capacity of Anopheles mosquitoes in a natural environment (due to Plasmodium-antagonistic capacities of Asaia) and be used in the development of tools for Asaia-based paratransgenetic malaria control.
Similar content being viewed by others

Introduction
In the context of malaria pre-elimination, the emergence of resistance in both vectors and parasites threatens the progress made in recent decades. The search for alternative methods is becoming a priority for the WHO and the scientific community. The vector environment has very often been neglected during the implementation of control strategies.
The present study focuses on the role of nectar-producing plants in reducing the vectorial capacity of malaria-transmitting mosquitoes. We found that flower nectar is home to viable strains of Asaia, known for their natural antagonism to Plasmodium falciparum, and we isolated Asaia strains from Ipomoea pes-caprae. Moreover, we found that bacterial exchanges between plants and mosquitoes are common in the natural environment and thus discovered the natural Asaia/Anopheles/flower nectar cycle. Through these exchanges, plants, as the habitat for Plasmodium-suppressing Asaia bacteria, may contribute to the reduction in the vectorial capacity of Anopheles (which has already been empirically reported). It was recently reported that an Asaia-based paratransgenetic malaria control strategy may also rely on the Asaia natural cycle as a delivery tool.
Particular attention should be paid to the environment in which mosquitoes are founding the development of malaria control strategies.
Malaria continues to be one of the most common causes of death on earth. In addition, its transmission is only partially controlled due to the risk of the emergence of insecticide-resistant mosquitoes. This information justifies the search for alternative and nonpolluting solutions to control mosquito-borne infections, especially since, with the risk of global warming, the area affected by malaria may increase. In this context, the search for biological control agents against malaria and its transmission is a considerable challenge. The simultaneous presence of Asaia and Plasmodium falciparum has not yet been reported in mosquitoes suggesting antagonism1,2. Thus, identifying the source of Asaia contamination in Anopheles may help to determine how to apply biological control to limit Anopheles infestation by P. falciparum. Recently, a paratransgenetic strategy was proposed based on Asaia bacteria that conditionally expressed an anti-plasmodial protein only when a blood meal was present3.
Nectarivory is extremely common in insects. Nectar is a sugar-rich liquid specially produced by plant glands (nectaries) to attract pollinating animals. The majority of nectarivorous insect species belong to the families Diptera, Coleoptera, Lepidoptera, Hymenoptera and Hemiptera. Nectar consumption has been established for many lower dipteran taxa, including not only the Tipulidae (crane flies), Cecidomyiidae (gall midges), Sciaridae (root gnats) and Mycetophilidae but also blood-feeding species, such as Ceratopogonidae (biting midges) and Culicidae (mosquitoes)4. The Diptera possess an elongated tube, which has characteristic morphological and functional adaptations to feed on either floral nectar (e.g., Bombyliidae, Syrphidae, and Nemestrinidae), vertebrate blood (e.g., Tabanidae and Glossinidae) or both (Culicidae, Simuliidae, and Ceratopogonidae)5.
Male and female mosquitoes require sugar to obtain the energy they need to survive in their natural environment. Their sugar requirements are generally fulfilled from flower nectar near larval breeding sites or around adult resting sites. Plant nectar is a nutritional supplement that females often use between blood meals and when hosts are absent or rare. It is also used as an energy source and thus allows both male and female mosquitoes to survive in their environment6. Thus, plants play a very important role in mosquito survival in their natural environment, providing them with an easily accessible meal, which increases their life span and reduces contact with humans6. Plants also provide male mosquitoes with enough energy to successfully fertilize females, thus ensuring species perennity7. Mosquitoes are attracted by the odors emitted by plants and use those odors to locate plants that can provide the sugar and/or chemicals they need to survive in their natural environment6.
Several studies have discussed the importance of arthropod vector microbiota for the prevention of certain vector-borne diseases8,9,10,11. The composition of these microbiota is not only influenced by the bacterial flora in breeding sites but also by the sources of adult mosquitoes’ diets12,13. Some bacteria from larval breeding sites or plants can be absorbed by mosquitoes when ingesting food12,14.
Although highly suspected, very little data are available regarding the potential of flower nectar to facilitate the food-borne transmission of microorganisms between blood-feeding dipterans. Indeed, until a flower opens, the nectar inside of it remains sterile; however, once a flower opens, the nectar can be contaminated and serve as a source of contamination for all the insects that feed on it. We therefore hypothesized that plants are the key link between mosquitoes and the mosquito-borne transmission of Asaia spp. Asaia bacteria were isolated for the first time from the flowers of the tropical plants Bauhinia purpurea and Plumbago auriculata15,16,17,18. In such cases, plants may indirectly play a negative role in the transmission of vector-borne diseases, such as malaria, by being reservoirs and fomites of Plasmodium-antagonistic bacteria such as Asaia2.
Indeed, the absorption of nectar from plants has already been reported to have negative effects on the transmission of certain diseases14. According to Hien et al., 2016, three mutually nonexclusive mechanisms could explain the effects of plant sugar on the ability of a vector to transmit P. falciparum19. First, the ingestion of secondary metabolites, such as alkaloids, terpenes and glycosides, has a negative effect on Plasmodium development. Indeed, these chemicals, present in all plant tissues, including flower nectar and fleshy fruits, are toxic to pests and reduce their life span and reproductive capacity and, therefore, play a key role in plants’ defense system against pests20,21. Second, feeding on poorly energetic plants could induce changes in the metabolism of mosquitoes and result in an energy deficiency that would limit the development of the parasite in the vector22. Third, plants can indirectly influence the development of parasites in mosquitoes through their effects on the immune system or gut microbiota. A deficit of energy would cause a dysfunction of the immune system and a reduction in the defenses against the parasite23. However, until now, the role of plants in the transmission of bacteria such as Asaia spp. and/or other microorganisms that have an antagonistic effect on Plasmodium sp. has not been described.
This study therefore aims to identify the role of plant nectar in the epidemiological cycle of Asaia bacteria and to test the hypothesis that plants act as a reservoir from which wild mosquitoes are constantly infected.
Results
Antibiogram and selective medium preparation
The resistance status of the previously isolated A. aff. bogorensis GD01 strain2 to different antibiotics was assessed (Table 1). The data obtained were then used to prepare an antibiotic-containing Columbia-based selective solid medium for the isolation of Asaia strains from naturally infected flowers and to obtain an Asaia-free mosquito line from a laboratory-reared colony of An. coluzzii.
In summary, A. aff. bogorensis GD01 was resistant to most families of the antibiotics tested (Table 1), except piperacillin-tazobactam, doxycycline, and carbapenems (doripenem and imipenem). Among them, gentamycin was used for the Columbia-based selective solid medium, and doxycycline was chosen to obtain our Asaia-free mosquito line.
Natural Asaia spp. infection in wild flowering plants
Flowers were collected from 11 endemic and widely distributed plant species (Table 2) across the study area, including those mainly encountered inside and around the villages of Dielmo and Simong. Among the collected flowers, only those of Datura metel 25% (2/8) and I. pes-caprae 25% (28/114) were found to be contaminated with Asaia spp. I. pes-caprae were collected at three different locations (Table 2); this was the most targeted plant due to its wide distribution in the study area and its proximity to mosquito breeding sites.
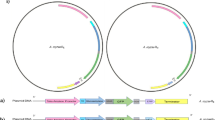
The colonies cultivated from all the infected specimens of I. pes-caprae that were morphologically similar to those of A. aff. bogorensis GD01, previously isolated by our team from An. gambiae s.l. collected from Dielmo (Senegal), were confirmed by qPCR2. The qPCR molecular screening showed that two colonies out of a total of 20 morphologically similar colonies isolated on Asaia-selective antibiotic-containing solid medium tested positive for A. aff. bogorensis. The subsequent phylogenetic analysis based on the sequences of almost the entire 16 S rRNA gene confirmed that the isolated strains were A. bogorensis, with slight differences from A. aff. bogorensis GD01, which was previously isolated from An. gambiae s.l. in the same village (Fig. 1), forming a clade within the A. bogorensis cluster.

Phylogeny of the A. aff. bogorensis strains isolated from I. pes-caprae. Figure 1 displays the molecular phylogenetic evolutionary reconstruction of the Senegalese strains of Asaia isolated from I. pes-caprae (Asaia IP-C1 and Asaia IP-C2) and those from An. gambiae s.l. from Dielmo (Asaia GD01) (in bold). The branch lengths are proportional to the genetic distance between the strains, considering the scale placed below the tree. The numbers in front of the nodes are the bootstrap values. The names of the reference strains retrieved from Genbank start with their accession number followed by the species name and the strain reference when available.
Natural prevalence of Asaia spp. in a laboratory mosquito strain
A subsample of 158 An. coluzzii (130 females and 28 males) of the laboratory strain were randomly selected and screened to determine the prevalence of natural infection by A. aff. bogorensis in our experimental study population. The results showed a global carriage rate of A. aff. bogorensis up to 61% (96/158), which was significantly higher among the female group (74%,96/130) than among the male group (46.4%,13/28)(χ2 = 6.864, df = 1, p = 0.008). The naturally infected An. coluzzii laboratory strain was used as the Asaia-positive line in the following experiments.
Experimental infection model
qPCR screening of newly opened Pseuderanthemum reticulatum flowers used in the experimental infection revealed the absence of natural Asaia infection. The plants were then isolated and protected from contact with arthropods. A batch of 100 An. coluzzii individuals, randomly selected from a naturally Asaia-infected laboratory colony, were exposed to P. reticulatum to confirm the effectiveness of transmission. During the entire experimental period, the only source of food for mosquitoes was the plant nectar, to which they were exposed for 3 consecutive days. Screening for Asaia infection on 36 randomly selected open flowers revealed the presence of Asaia spp. in 31/36 (86.1%) of the flowers. This shows that mosquito nectar feeding could be a significant route of bacterial contamination of plant flowers.
PCR screening of a randomly selected subsample of 56 anopheles (30 females and 26 males) out of the 200 An. coluzzii treated with antibiotics for 3 days to establish an Asaia-free lineage revealed no infection, thus confirming the Asaia-free status of the batch. The remaining females were reared to obtain the first generation (F1) of the lineage used for the following experiments. Samples of the Asaia-free lineage, selected randomly and in a timely manner, were regularly tested to verify whether reinfection with Asaia occurred. The lineage remained pure and uncontaminated throughout the experiments.
The subsequent exposure of two replicates of 265 An. coluzzii specimens belonging to the Asaia-free laboratory colony to P. reticulatum plants previously infected by the Asaia-positive lineage revealed a contamination rate of 27% (71/265) (Table 3). The infection rate was significantly lower in the female group (16.0%, 24/153) than in the male group (42.0%, 47/112) (χ2 = 21.45, df = 1, p < 0.05). These results have enabled us to propose a transmission cycle of Asaia spp. among mosquitoes in the natural environment (Fig. 2).

Putative Asaia spp. natural transmission cycle involving endemic plants in the field. (A) Emerging adult mosquitoes (infected or not) feed on plants to obtain the nectar needed for their survival and ensure reproduction. During the diet or oral (potentially co-feeding) transmission phase (B), mosquitoes are contaminated or contaminate the plants they feed on. Then, during the copulation or sexual transmission phase (C), male or female mosquitoes can contaminate their partners. The fertilized females will search for a blood meal to ensure the maturation of their eggs (D), and the males will continue to feed on nectar from plants, such as I. pes-caprae, to satisfy their sugar needs and maintain the oral transmission cycle (E). After a blood meal, the females will lay eggs (F), some of which will be contaminated by Asaia spp. as well as larvae (known as the transstadial transmission phase).The infected emerging adults will initiate and maintain the transmission cycle.
Discussion
Arthropods maintain a complex relationship with the elements present in their natural environment. The impact of these elements on the transmission of vector-borne diseases has long been under-researched. However, nectar-producing plants play a very important role in the transmission of pathogens hosted by nectarivorous arthropods. Nectar is one of the main sources of food for insects and helps to increase their survival in the natural environment. As in larval breeding sites, where some bacteria belonging to the microbial flora can be ingested by mosquito larvae13,24, bacteria present in plant nectar can also pass into adult mosquitoes during nectar feeding. Bacterial species, such as those belonging to the Asaia genus, represent great potential for use in biological malaria control, especially in the current context of the emerging resistance of P. falciparum to artemisinin25,26,27 and of the main vectorial species of malaria to insecticides28,29,30,31. Asaia bacteria belong to the Acetobacteraceae family and are often isolated from fermented foods or plants32,33. They have also been described as symbionts of insects that often feed on natural sources of sugar34,35.
Asaia spp. could be modified by trangenesis to obtain a strong antagonistic effect against P. falciparum9,36 and may be able to protect mosquitoes against plasmodial infection. This antagonistic effect was first highlighted by Favia et al.9,37. Furthermore, the Asaia bacterium has been found to be stably associated with the microbiota of larvae and adults of An. stephensi, An. maculipennis and An. gambiae37,38. This bacterium has also been found to be stably associated with the microbiota of the laboratory strain of An. coluzzii, and naturally infect An. funestus2. Online searches in the metagenomic 16 S databases have also shown the presence of Asaia in different nectarivorous insects, such as butterflies and bees, in which the most likely propagation pathways identified were the venereal route, paternal transmission39,40 and food ingestion41.
Asaia, a genus of acetic acid bacteria was first discovered and isolated from plants, namely, Bauhinia purpurea and Plumbago auriculata flowers42. Currently, only 8 species of Asaia have been described: Asaia astilbis43, Asaia bogorensis42, Asaia krungthepensis18, Asaia lannensis44, Asaia platycodi43, Asaia prunellae43, Asaia siamensis16, and Asaia spathodeae45. To our knowledge, no studies have been able to highlight the role of plants in the contamination of arthropods by Asaia spp. In the study of tularemia, scientists have shown through an experimental model that nectar may play a key role in the spread of the disease. Indeed, Francisella tularensis has the ability to survive in nectar, thus facilitating its ingestion by vectors, which are then infected through the nectar feeding route, thus ensuring the transmission of the bacteria14. Asaia strains were found in the nectar of some plants in this study, suggesting that infected plants could be the missing link to the maintenance of mosquito infection in nature. Our hypothesis is reinforced by previous reports on the ability of bacteria to survive in plant nectar14. Using both molecular and microbiological methods, we demonstrated natural infection of flowers of I. pes-caprae and D. metel by Asaia bacteria. The existence of viable Asaia strains in I. pes-caprae flowers was demonstrated using bacterial isolation techniques with homemade selective medium. Moreover, we established an in vivo transmission model demonstrating the key role played by plants in the natural transmission of Asaia to Anopheles via contaminated nectar.
Together, the high prevalence of Asaia spp. in plants in the direct environment of mosquitoes (I. pes-caprae growing close to mosquito breeding sites and D. metel found in the vicinity of human dwellings, which also serving as adult mosquito resting sites) and the transmission of Asaia from infected to noninfected An. coluzzii mosquitoes via flower nectar provide enough evidence to support our hypothesis. Based on this evidence, we suggest the transmission cycle for Asaia spp. in the natural environment (Fig. 2). In addition to mosquitoes, the carriage of Asaia spp. is widespread in nectarivorous insects in nature. Indeed, Asaia spp. have already been isolated or identified in Anopheles mosquitoes46, monarch butterflies (Danaus plexippus, Genbank accession number KF414321), honey bees47, Ae. aegypti, Apis mellifera, Drosophila melanogaster, Saccharicoccus sacchari39 and many other insects35,48.
In the context of the spread of insecticide resistance and the increased need for innovative approaches to control vectors, the development of genetically modified mosquitoes (GMMs) that are resistant to pests has emerged as the most promising alternative to insecticide interventions49. However, the GMM approach has one main limitation: it modifies the fitness and competitiveness of mosquitoes in relation to natural mosquito populations9. Given the recent evidence of the stable natural infection of malaria vectors by several bacteria that may impede Plasmodium development, Asaia spp. have become one of the most promising potential paratransgenic weapons against malaria9. Asaia spp. are easily cultivable and can be genetically manipulated to deliver anti-pathogen effects on molecules that are then reinserted into their insect host36. A prominent model of the paratransgenetic malaria control strategy based on Asaia bacteria has been recently proposed3. Asaia bacteria have been genetically modified to release scorpine into the intestinal lumen of An. stephensi, leading to the elimination of P. berghei.
We have noticed that in laboratory colonies of An. coluzzii, male mosquitoes are more likely than females to be infected by Asaia. We therefore speculate that the females (probably fertilized) were in search of a blood meal for maturing their eggs and, therefore, consume less Asaia-infected nectar. However, this hypothesis was not tested in the present study.
In fine, this study is an important steppingstone toward the successful use of any Asaia-based vector control approach. Given the strong antimalarial capacities of these bacteria, the involvement of plants as a key link between nectarivorous insects, including several hematophagous arthropods that transmit vector-borne diseases, in the natural transmission cycle of Asaia bacteria offers an unprecedented opportunity for the direct or indirect use of Asaia in an alternative biological and/or transgenic approach to malaria control.
Material and Methods
Description of the study area and plant collection
Entire flowers were manually collected from endemic plants growing on the banks of the Nema River, next to Dielmo (13°43′23.43′′N, 16°24′46.27′′W) and Simong (13°37′55.63′′N, 16°23′08.24′′W) (Fig. 3). Dielmo is located in the Fatick region of Senegal, in an area of Sudanese savannah approximately 174 miles from Dakar and 9.32 miles north of the Gambian Republic. Since 1990, the area near Dielmo has hosted one of the oldest cohort studies investigating the epidemiological relationships between the malaria parasite and its vectors and human hosts50. Previous studies have already laid out the geographical and epidemiological characteristics as well as the dynamics of malaria in Dielmo50,51.

Map of the study sites. The positions of the two collection sites are indicated by red (Simong) and green (Dielmo) dots. Both villages are close to the Saloum delta, one of the most irrigated areas in Senegal.
Members of the An. gambiae s.l. and An. funestus group are the main vectors of malaria in this area52,53.
The flowers were collected from 27 to 30 October 2016. They were cut at the stalk level, placed in bags (one bag per plant), stored at 4 °C in the Dielmo laboratory and transported to Dakar at the same temperature. In Dakar, the samples were immediately frozen at −80 °C until they were processed to assess their molecular biology and for bacterial cultures.
Molecular analyses
DNA extraction
The petals and sepals of each flower were removed, and then the peduncle was cut off. The remaining flower parts, including nectar and pistil, were immersed in an Eppendorf tube containing 600 μl of 1X phosphate buffered saline (PBS) and then crushed. After centrifugation, 200 μl of the supernatant was collected for DNA extraction, which was performed according to the 2% CTAB method54 supplemented with β-mercaptoethanol at a concentration of 0.2%. The mixture was left to digest overnight. The same DNA extraction procedure was performed for the mosquitoes. Before DNA extraction, individual specimens were sacrificed at −80 °C, washed with 70% ethanol to remove superficial environmental bacteria and then crushed. Then, genomic DNA was immediately extracted from the whole body of each individual specimen as previously described54.
Asaia bogorensis molecular detection
Taqman® qPCR methods with custom designed primers and probe sets were used to screen samples for the presence of Asaia spp. by targeting the rpoB gene (Fwd: 5′-GACGCCAAGGACCTGATCTA-3′; Rev: 5′-ATAGGCCAGGATTTCGTCCT-3′; Probe: 6-FAM-GGTCACGACCCTGCTCTATG-TAMRA)2. The targeted region was amplified in a total reaction volume of 20 μl, containing 10 μl of ROCHE® master mix (Roche Diagnostics, Indianapolis, IN, USA), 0.5 μl of each primer, 0.5 μl of the probe, 3 μl of distilled water, 0.5 µl of UDG, and 5.0 μl of test DNA. The conditions of the amplification with the CFX96 Touch detection system (Bio-Rad, Marnes-la-Coquette, France) were as follows: 2 minutes at 50 °C for UDG action followed by an initial denaturation of 5 minutes at 95 °C, then 39 cycles of 5 seconds at 95 °C and 30 seconds at 60 °C. A sample was considered positive when the Ct value was less than 35 cycles. The Anopheles negative control was prepared from a crush of certified Asaia-free An. coluzzii, and the positive control was prepared from An. coluzzii to which we added a suspension of A. aff. bogorensis GD01.
16 S rRNA gene sequencing and phylogenetic tree construction
The molecular phylogenetic evolutionary history was inferred from a nearly complete 16 S rDNA sequence (1249 bp), which was sequenced using universal 16 S rDNA primers fD1 and rp255, as described by Drancourt et al.56.
Initially, we used a Peltier PTC-200 model thermal cycler (MJ Research Inc., Watertown, MA, USA) to perform the PCR amplifications. The reactions were carried out using HotStar Taq polymerase (Qiagen, Hilden, Germany) following the manufacturer’s instructions. For each assay, we included negative and positive controls for the validation of the run. The success of amplification was confirmed by electrophoresis on a 1.5% agarose gel.
The purification of the PCR products was performed using NucleoFast 96 PCR plates (Macherey-Nagel EURL, Hoerdt, France) according to the manufacturer’s instructions. The products of the amplification were then sequenced using a Big Dye Terminator Cycle Sequencing Kit (Perkin Elmer Applied Biosystems, Foster City, CA) with an ABI automated sequencer (Applied Biosystems).
The amplified sequences were assembled and corrected using ChromasPro software (ChromasPro 1.7, Technelysium Pty Ltd., Tewantin, Australia) and then compared with reference sequences available in Genbank using the BLASTN server (http://blast.ncbi.nlm.nih.gov/Blast.cgi) to identify closely related species and/or strains.
The taxonomic relationships of the new strains were inferred against the existing isolates. The reference sequences retrieved from the Genbank database, together with the Senegalese strains, were aligned using the ClustalW multisequence alignment program57 in BioEdit software58. A maximum likelihood phylogenetic tree was reconstructed using TOPALi v2.5 based on the Hasegawa-Kishino-Yano (HKY85) substitution model59, which includes the proportion of invariable sites and the gamma distribution. The robustness of the individual branches was estimated by bootstrapping with 100 replicates60.
Establishment of an Asaia-free An. coluzzii strain
Sensitivity of A. aff. bogorensis GD01 to antibiotics
The sensitivity of the A. aff. Bogorensis GD01 strain, previously isolated from An. gambiae from Dielmo village2, to antibiotics was determined. A pure suspension of A. aff. bogorensis GD01 at a concentration equal to the 0.5 McFarland standard was inoculated onto Mueller-Hinton agar medium61,62. Antibiotic (amoxicillin, amoxicillin/clavulanic acid, piperacillin/tazobactam, doripenem, imipenem, aztreonam, ceftriaxone, ceftazidime, cefpirome, gentamicin, doxycycline, erythromycin, trimethoprim/sulfamethoxazole, ciprofloxacin, linezolid, metronidazole and rifampicin) discs were deposited onto dried agar plates and the plates were then incubated at 28 °C under 5% CO2 atmosphere. Finally, the inhibition diameters were measured and compared with the reading charts according to the manufacturer’s instructions (i2a - Siège Social, 401 Avenue du Walhalla, CS83406, 34060 Montpellier Cedex 2,France) and the recommendations of the Antibiogram Committee of the French Society of Microbiology63.
Isolation of strains of Asaia spp. from wild plants
Suspensions of the qPCR-positive I. pes-caprae were seeded on Columbia agar supplemented with 5% fresh sheep blood and 4 mg/l gentamicin (batch No. F140512, manufactured by Xin K. Pharm Co. Ltd). The seeded medium was then incubated at 28 °C under an atmosphere of 5% CO2. The colonies morphologically resembling Asaia were seeded on another Columbia agar plate supplemented with 5% fresh sheep blood but without antibiotics and incubated under the same conditions. Approximately 50 purified colonies were then collected and suspended in 200 μl of sterile 1X PBS and used for bacterial DNA extraction using the 2% CTAB method supplemented with 25 μl of proteinase K (Tritirachium album, 25 mg, ref. EUC0090-A, EUROMEDEX, 24, rue des Tuileries BP684 67460 Souffelweyersheim, France). Finally, the extracted DNA was screened for the presence of Asaia using A. bogorensis-specific qPCR.
Treatment of adult mosquitoes with antibiotics
Adult An. coluzzii colonies maintained at a temperature of 26 ± 2 °C and a relative humidity of 80 ± 10% in our laboratory, as described previously50,51,52,53, were fed a mixture of sterile water containing doxycycline (batch No. B113304 manufactured by SERB Laboratories) at a concentration of 8 mg/l and 10% sucrose solution for 4 days. qPCR was performed on the treated mosquitoes to confirm their Asaia-free status.
Insectary rearing of the Asaia-free mosquito strain
The Asaia-free mosquito line, fed sugar solution prepared with autoclaved water, was maintained at a temperature of 26 ± 2 °C and a relative humidity of 80 ± 10%. When feeding the mosquitoes, a volume of 50 ml of autoclaved water was filtered with a 0.2 µm filter and then mixed with sterile sucrose solution at a concentration of 10%. This mixture was offered daily to mosquitoes and changed daily to prevent mosquito reinfection. The physical conditions of breeding were the same as those of the other mosquitoes in the insectarium. The control of the absence of Asaia in this mosquito line was assessed weekly and before each experiment using our Asaia-specific custom-designed qPCR system.
Design of the experimental infection through transmission of A.aff. bogorensis
The two An. coluzzii colonies (Asaia-infected and Asaia-free), maintained separately in the insectarium of the VITROME laboratory of the Institut de Recherche pour le Développement (Dakar, Senegal), were used for the experimental infection of A. aff. bogorensis transmission. The common ornamental plant P. reticulatum (Acanthaceae) was selected as the source of nectar during the experimental infection study after the confirmation of its natural Asaia-free status and due to its easy procurement from urban horticulturists in Dakar, where our laboratory is based. The plants were grown in pots within a closed room with no access to flying arthropods to prevent the contamination of newly opened flowers with Asaia. Randomly selected flowers from each plant were screened for the presence of Asaia spp. and only plants that were negative were used for the subsequent experimental infection.
Overall, 310 mosquitoes of the naturally Asaia-infected laboratory colonies (123 males and 187 females) were brought into contact with an uninfected P. reticulatum bearing newly opened flowers. The plant in its pot was introduced into a closed 100 × 50 × 50 cm mosquito cages for 3 days. During the experimental infection, the average temperature of the standard thermo-hygronomic device was 26 ± 2 °C, and the relative humidity was 80 ± 10%. At the end of the third day, the mosquitoes were sacrificed at −80 °C and then screened with randomly selected flowers using our qPCR method to confirm that the mosquitoes and the flowers of the exposed plants were infected with Asaia. The following day, an unfed batch of mosquitoes from the Asaia-free lines was allowed to feed for 3 days on the Asaia-infected plants under the same conditions described above. After exposure, the mosquitoes were sacrificed, stored individually in Eppendorf tubes for DNA extraction and then examined for Asaia infection. This experiment was repeated twice with the same number of anopheles for both lineages and the same plant species under identical physical conditions.
Statistical analyses
This study was carried out over two consecutive years. Two groups of mosquioes were used each time. This first group was the An. coluzzii strain naturally infected by A. aff. bogorensis and the second was as An. coluzzii Asaia-free group obtained after antibiotic treatment and then exposed to infected P. reticulatum. Within each group, we analyzed the differences in infection rates between the two sexes. Statistical analyses were performed with Epi Info software version 7.0.8.8.0 (Centers for Disease Control and Prevention, Atlanta, GA, USA). The differences were analyzed using the Yates corrected χ2 test with one degree of freedom and a 95% confidence interval. The difference was considered significant when the bilateral p value < 0.05
References
Capone, A. et al. Interactions between Asaia, Plasmodium and Anopheles: new insights into mosquito symbiosis and implications in Malaria Symbiotic Control. Parasites & vectors 6, 182, https://doi.org/10.1186/1756-3305-6-182. (2013).
Bassene, H. et al. A pilot study on isolation of Asaia and detecting its co-presence with Plasmodium falciparum in two major malaria vectors in Senegal. J. Parasitol. Vector Biol. 11, 1–9, https://doi.org/10.5897/JPVB2018.0341 (2019).
Shane, J. L., Grogan, C. L., Cwalina, C. & Lampe, D. J. Blood meal-induced inhibition of vector-borne disease by transgenic microbiota. Nature communications 9, 4127, https://doi.org/10.1038/s41467-018-06580-9 (2018).
Yeates, D. K. & Wiegmann, B. M. The Evolutionary Biology of Flies (Columbia University Press, 2005).
Krenn, H. W., Plant, J. D. & Szucsich, N. U. Mouthparts of flower-visiting insects. Arthropod Structure & Development 34, 1–40 (2005).
Nikbakhtzadeh, M. R., Terbot, J. W. 2nd, Otienoburu, P. E. & Foster, W. A. Olfactory basis of floral preference of the malaria vector Anopheles gambiae (Diptera: Culicidae) among common African plants. Journal of vector ecology: journal of the Society for Vector Ecology 39, 372–383, https://doi.org/10.1111/jvec.12113 (2014).
Stone, C. M., Taylor, R. M., Roitberg, B. D. & Foster, W. A. Sugar deprivation reduces insemination of Anopheles gambiae (Diptera: Culicidae), despite daily recruitment of adults, and predicts decline in model populations. Journal of medical entomology 46, 1327–1337, https://doi.org/10.1603/033.046.0611 (2009).
Aksoy, S. Control of tsetse flies and trypanosomes using molecular genetics. Veterinary parasitology 115, 125–145, https://doi.org/10.1016/S0304-4017(03)00203-6 (2003).
Favia, G. et al. Bacteria of the genus Asaia: a potential paratransgenic weapon against malaria. Advances in experimental medicine and biology 627, 49–59, https://doi.org/10.1007/978-0-387-78225-6_4 (2008).
Jayakrishnan, L., Sudhikumar, A. V. & Aneesh, E. M. Role of gut inhabitants on vectorial capacity of mosquitoes. Journal of vector borne diseases 55, 69–78, https://doi.org/10.4103/0972-9062.242567 (2018).
Telleria, E. L., Martins-da-Silva, A., Tempone, A. J. & Traub-Cseko, Y. M. Leishmania, microbiota and sand fly immunity. Parasitology 145, 1336–1353, https://doi.org/10.1017/S0031182018001014 (2018).
Gimonneau, G. et al. Composition of Anopheles coluzzii and Anopheles gambiae microbiota from larval to adult stages. Infection, Genetics and Evolution 28, 715–724, https://doi.org/10.1016/j.meegid.2014.09.029 (2014).
Akorli, J. et al. Seasonality and Locality Affect the Diversity of Anopheles gambiae and Anopheles coluzzii Midgut Microbiota from Ghana. PloS one 11, 1–18, https://doi.org/10.1371/journal.pone.0157529 (2016).
Kenney, A. et al. The potential for flower nectar to allow mosquito to mosquito transmission of Francisella tularensis. PloS one 12, e0175157, https://doi.org/10.1371/journal.pone.0175157 (2017).
Yamada, Y. et al. Asaia bogorensis gen. nov., sp nov., an unusual acetic acid bacterium in the α-Proteobacteria. Int. J. Syst. Evol. Microbiol. 50, 823–829, https://doi.org/10.1099/00207713-50-2-823 (2000).
Katsura, K. et al. Asaia siamensis sp. nov., an acetic acid bacterium in the alpha-proteobacteria. International journal of systematic and evolutionary microbiology 51, 559–563, https://doi.org/10.1099/00207713-51-2-559 (2001).
Moore, J., Mccalmont, M., Xu, J., Millar, B. & Heaney, N. Asaia sp., an unusual spoilage organism of fruit-flavored bottled water. Appl Environ Microbiol 68, 4130–4131, https://doi.org/10.1128/aem.68.8.4130-4131.2002 (2002).
Yukphan, P., Potacharoen, W., Tanasupawat, S., Tanticharoen, M. & Yamada, Y. Asaia krungthepensis sp. nov., an acetic acid bacterium in the alpha-Proteobacteria. International journal of systematic and evolutionary microbiology 54, 313–316 (2004).
Gu, W., Muller, G., Schlein, Y., Novak, R. J. & Beier, J. C. Natural plant sugar sources of Anopheles mosquitoes strongly impact malaria transmission potential. PloS one 6, e15996, https://doi.org/10.1371/journal.pone.0015996 (2011).
Stam, J. M. et al. Plant interactions with multiple insect herbivores: from community to genes. Annual review of plant biology 65, 689–713, https://doi.org/10.1146/annurev-arplant-050213-035937 (2014).
Ode, P. J. Plant chemistry and natural enemy fitness: effects on herbivore and natural enemy interactions. Annu Rev Entomol 51, 163–185, https://doi.org/10.1146/annurev.ento.51.110104.151110 (2006).
Hall, S. R. et al. Quality matters: resource quality for hosts and the timing of epidemics. Ecology letters 12, 118–128, https://doi.org/10.1111/j.1461-0248.2008.01264.x (2009).
Sheldon, B. & Verhulst, S. Ecological immunity: costly parasite defences and trade-offs in evolutionary ecology. Trends Ecol Evol. 11, 317–321, https://doi.org/10.1016/0169-5347(96)10039-2 (1996).
Gimonneau, G. et al. Behavioural responses of Anopheles gambiae sensu stricto M and S molecular form larvae to an aquatic predator in Burkina Faso. Parasites & vectors 5, 65, https://doi.org/10.1186/1756-3305-5-65 (2012).
Dondorp, A. M. et al. Artemisinin resistance in Plasmodium falciparum malaria. The New England journal of medicine 361, 455–467, https://doi.org/10.1056/NEJMoa0808859 (2009).
WHO. Global Report on Antimalarial Drug Efficacy and Drug Resistance: 2000–2010. (World Health Organization, Geneve, 2010).
Ariey, F. et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 505, 50–55, https://doi.org/10.1038/nature12876 (2014).
Chandre, F. et al. Pyrethroid cross resistance spectrum among populations of Anopheles gambiae s.s. from Cote d’Ivoire. Journal of the American Mosquito Control Association 15, 53–59 (1999).
Martinez-Torres, D. et al. Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae s.s. Insect molecular biology 7, 179–184 (1998).
Ranson, H. et al. Identification of a point mutation in the voltage-gated sodium channel gene of Kenyan Anopheles gambiae associated with resistance to DDT and pyrethroids. Insect molecular biology 9, 491–497, https://doi.org/10.1046/j.1365-2583.2000.00209.x (2000).
Cuamba, N., Morgan, J. C., Irving, H., Steven, A. & Wondji, C. S. High level of pyrethroid resistance in an Anopheles funestus population of the Chokwe District in Mozambique. PloS one 5, e11010, https://doi.org/10.1371/journal.pone.0011010 (2010).
Kommanee, J. et al. Identification of Acetobacter strains isolated in Thailand based on 16S-23S rRNA gene ITS restriction and 16S rRNA gene sequence analyses. Ann Microbiol 58, 319–324, https://doi.org/10.1007/Bf03175337 (2008a).
Kommanee, J. et al. Identification of Gluconobacter strains isolated in Thailand based on 16S-23S rRNA gene ITS restriction and 16S rRNA gene sequence analyses. Ann Microbiol 58, 741–747, https://doi.org/10.1007/Bf03175584 (2008b).
Crotti, E. et al. Acetic acid bacteria, newly emerging symbionts of insects. Appl Environ Microbiol 76, 6963–6970, https://doi.org/10.1128/AEM.01336-10 (2010).
Kounatidis, I. et al. Acetobacter tropicalis Is a Major Symbiont of the Olive Fruit Fly (Bactrocera oleae). Appl Environ Microb 75, 3281–3288, https://doi.org/10.1128/Aem.02933-08 (2009).
Mancini, M. V. et al. Paratransgenesis to control malaria vectors: a semi-field pilot study. Parasites & vectors 9, 140, https://doi.org/10.1186/s13071-016-1427-3 (2016).
Favia, G. et al. Bacteria of the genus Asaia stably associate with Anopheles stephensi, an Asian malarial mosquito vector. PNAS 104, 9047–9051, https://doi.org/10.1073/pnas.0610451104 (2007).
Damiani, C. et al. Mosquito-bacteria symbiosis: the case of Anopheles gambiae and Asaia. Microbial ecology 60, 644–654, https://doi.org/10.1007/s00248-010-9704-8 (2010).
Crotti, E. et al. Asaia, a versatile acetic acid bacterial symbiont, capable of cross-colonizing insects of phylogenetically distant genera and orders. Environm Microbiol. 11, 3252–3264, https://doi.org/10.1111/j.1462-2920.2009.02048.x (2009).
Damiani, C. et al. Paternal transmission of symbiotic bacteria in malaria vectors. Current biology: CB 18, R1087–1088, https://doi.org/10.1016/j.cub.2008.10.040 (2008).
Gonella, E. et al. Horizontal transmission of the symbiotic bacterium Asaia sp. in the leafhopper Scaphoideus titanus Ball (Hemiptera: Cicadellidae). BMC Microbiol 12(Suppl 1), S4, https://doi.org/10.1186/1471-2180-12-S1-S4 (2012).
Yamada, Y. et al. Asaia bogorensis gen. nov., sp. nov., an unusual acetic acid bacterium in the alpha-Proteobacteria. Int J Syst Evol Microbiol 50, 823–829, https://doi.org/10.1099/00207713-50-2-823 (2000).
Suzuki, R. et al. Asaia astilbes sp. nov., Asaia platycodi sp. nov., and Asaia prunellae sp. nov., novel acetic acid bacteria isolated from flowers in Japan. The Journal of general and applied microbiology 56, 339–346, https://doi.org/10.2323/jgam.56.339 (2010).
Malimas, T. et al. Asaia lannaensis sp. nov., a new acetic acid bacterium in the Alphaproteobacteria. Bioscience, biotechnology, and biochemistry 72, 666–671, https://doi.org/10.1271/bbb.70233 (2008).
Kommanee, J. et al. Asaia spathodeae sp. nov., an acetic acid bacterium in the alpha-Proteobacteria. The Journal of general and applied microbiology 56, 81–87, https://doi.org/10.2323/jgam.56.81 (2010).
Samaddar, N. et al. Nitrogen fixation in Asaia sp. (family Acetobacteraceae). Current microbiology 63, 226–231, https://doi.org/10.1007/s00284-011-9968-3 (2011).
Good, A. P., Gauthier, M. P. L., Vannette, R. L. & Fukami, T. Honey Bees Avoid Nectar Colonized by Three Bacterial Species, But Not by a Yeast Species, Isolated from the Bee Gut. PloS one 9, doi:ARTN e8649410.1371/journal.pone.0086494 (2014).
Deutscher, A. T. et al. Near full-length 16S rRNA gene next-generation sequencing revealed Asaia as a common midgut bacterium of wild and domesticated Queensland fruit fly larvae. Microbiome 6, doi:ARTN 8510.1186/s40168-018-0463-y (2018).
Grossman, G. L. et al. Germline transformation of the malaria vector, Anopheles gambiae, with the piggyBac transposable element. Insect molecular biology 10, 597–604 (2001).
Trape, J. F. et al. The Dielmo project: a longitudinal study of natural malaria infection and the mechanisms of protective immunity in a community living in a holoendemic area of Senegal. Am J Trop Med Hyg 51, 123–137, https://doi.org/10.4269/ajtmh.1994.51.123 (1994).
Rogier, C. & Trape, J. F. Study of premunition development in holo- and meso-endemic malaria areas in Dielmo and Ndiop (Senegal): preliminary results, 1990–1994. Medecine tropicale: revue du Corps de sante colonial 55, 71–76 (1995).
Sougoufara, S., Harry, M., Doucoure, S., Sembene, P. M. & Sokhna, C. Shift in species composition in the Anopheles gambiae complex after implementation of long-lasting insecticidal nets in Dielmo, Senegal. Medical and veterinary entomology 30, 365–368, https://doi.org/10.1111/mve.12171 (2016).
Trape, J. F. et al. The rise and fall of malaria in a West African rural community, Dielmo, Senegal, from 1990 to 2012: a 22 year longitudinal study. The Lancet. Infectious diseases 14, 476–488, https://doi.org/10.1016/S1473-3099(14)70712-1 (2014).
Sambou, M. et al. Identification of rickettsial pathogens in ixodid ticks in northern Senegal. Ticks and tick-borne diseases 5, 552–556, https://doi.org/10.1016/j.ttbdis.2014.04.002 (2014).
Weisburg, W. G., Barns, S. M., Pelletier, D. A. & Lane, D. J. 16S ribosomal DNA amplification for phylogenetic study. Journal of bacteriology 173, 697–703, https://doi.org/10.1128/jb.173.2.697-703.1991 (1991).
Drancourt, M., Bollet, C. & Raoult, D. Stenotrophomonas africana sp. nov., an opportunistic human pathogen in Africa. International journal of systematic bacteriology 47, 160–163, https://doi.org/10.1099/00207713-47-1-160 (1997).
Thompson, J. D., Higgins, D. G. & Gibson, T. J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22, 4673–4680 (1994).
Hall, T. A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 41, 95–98 (1999).
Hasegawa, M., Kishino, H. & Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. Journal of molecular evolution 22, 160–174 (1985).
Felsenstein, J. Confidence-Limits on Phylogenies - an Approach Using the Bootstrap. Evolution; international journal of organic evolution 39, 783–791, https://doi.org/10.2307/2408678 (1985).
Matuschek, E., Ahman, J., Webster, C. & Kahlmeter, G. Antimicrobial susceptibility testing of colistin - evaluation of seven commercial MIC products against standard broth microdilution for Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Acinetobacter spp. Clinical microbiology and infection: the official publication of the European Society of Clinical Microbiology and Infectious Diseases 24, 865–870, https://doi.org/10.1016/j.cmi.2017.11.020 (2018).
Safari Sinegani, A. A. & Younessi, N. Antibiotic resistance of bacteria isolated from heavy metal-polluted soils with different land uses. Journal of global antimicrobial resistance 10, 247–255, https://doi.org/10.1016/j.jgar.2017.05.012 (2017).
Comité de l’antibiogramme. Recommandations 2012. Janvier 2012 edn, (Société Française de Microbiologie, 2012).
Acknowledgements
The authors are grateful to Dr. Ibrahima Dia of the Pasteur Institute for their support in providing mosquito breeding materials and valuable suggestions. We also thank Sacy Nadarajane and Vanessa Makanta for their technical assistance and Omar Thiaw, third year PhD student at the Vecteurs - Infections TROpicales et MEditerranéennes - (VITROME) of the French Institute of Recherche for sustainable Development, for his advice on breeding anopheles and his availability. This study was supported by the Institut Hospitalo-Universitaire (IHU) Méditerranée Infection, the National Research Agency under the program “Investissements d’avenir”, reference ANR-10-IAHU-03, the Région Provence Alpes Côte d’Azur and European funding FEDER PRIMI.
Author information
Authors and Affiliations
Contributions
Conceptualization: O.M., H.B., D.R. Data curation: H.B., O.M., D.R. Formal analysis: H.B. Funding acquisition: F.F., D.R. Investigation: H.B., O.M., E.H.A.N., F.F., S.D., O.F., C.S., D.R. Methodology: O.M., H.B., E.H.A.N., D.R. Project administration: O.M., F.F., D.R. Writing–original draft: H.B., O.M., F.F., D.R. Writing–review & editing: H.B., O.M., E.H.A.N., F.F., S.D., O.F., C.S., D.R.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bassene, H., Niang, E.H.A., Fenollar, F. et al. Role of plants in the transmission of Asaia sp., which potentially inhibit the Plasmodium sporogenic cycle in Anopheles mosquitoes. Sci Rep 10, 7144 (2020). https://doi.org/10.1038/s41598-020-64163-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-64163-5
This article is cited by
-
Two promising candidates for paratransgenesis, Elizabethkingia and Asaia, increase in both sexes of Anopheles gambiae mosquitoes after feeding
Malaria Journal (2024)
-
Characterization of the reproductive tract bacterial microbiota of virgin, mated, and blood-fed Aedes aegypti and Aedes albopictus females
Parasites & Vectors (2021)
-
Molecular detection and maternal transmission of a bacterial symbiont Asaia species in field-caught Anopheles mosquitoes from Cameroon
Parasites & Vectors (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
