
Abstract
The endoplasmic reticulum (ER) stress response constitutes cellular reactions triggered by a wide variety of stimuli that disturb folding of proteins, often leading to apoptosis. ER stress-induced apoptotic cell death is thought to be an important contributor to many human pathological conditions. The molecular mechanism of this apoptosis process has been highly controversial with both the receptor and the mitochondrial pathways being implicated. Using knockout mouse models and RNAi-mediated gene silencing in cell lines, our group and others had demonstrated the importance of the mitochondrial apoptotic pathway in ER stress-induced cell death, particularly the role of the pro-apoptotic BH3-only BCL-2 family members, BIM and PUMA. However, a recent report suggested a central role for the death receptor, DR5, activated in a ligand-independent manner, and the initiator caspase, caspase-8, in ER stress-induced cell death. This prompted us to re-visit our previous observations and attempt to reproduce the newly published findings. Here we report that the mitochondrial apoptotic pathway, activated by BH3-only proteins, is essential for ER stress-induced cell death and that, in contrast to the previous report, DR5 as well as caspase-8 are not required for this process.
Similar content being viewed by others


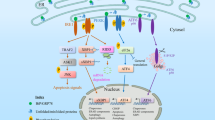
Main
The endoplasmic reticulum (ER) is the site for quality control for accurate folding of secretory and membrane-associated proteins in eukaryotic cells. This is achieved through the coordinated function of several proteins, including chaperones, protein peptide isomerases and glycosyl transferases.1 Several factors impact protein folding in the ER, including ATP, Ca2+, an oxidizing environment and gene mutations. As a consequence, the ER is highly sensitive to stress factors that alter cellular energy levels, calcium homeostasis or the redox status. Such stresses reduce the protein folding capacity of the ER, and this can result in the accumulation and aggregation of unfolded proteins, leading to a condition referred to as ER stress. In most cases, cellular responses ensure that ER stress does not overwhelm the protein-folding capacity. This is achieved by activating the so-called unfolded protein response (UPR). The UPR promotes cellular repair and survival by reducing the load of unfolded proteins through attenuating de novo protein synthesis and/or upregulation of chaperones, enzymes and structural components of the ER.2 However, unmitigated ER stress can lead to apoptotic cell death and this is thought to contribute to the development of several diseases, including type 2 diabetes and neurodegenerative disorders.3 For this reason, understanding the molecular mechanisms regulating ER stress-induced apoptosis has been the subject of intense research. This has led to the identification of the critical role of the pro-apoptotic BH3-only proteins, BIM and PUMA, in this process.4, 5, 6
However, a recent report suggested that the death receptor 5 (DR5), upregulated by the transcription factor CHOP, was a crucial mediator of ER‐stress‐induced apoptosis in cancer cells, with BH3-only proteins, such as BIM and PUMA, having only a minor role.7 It was suggested that ligand-independent DR5-mediated activation of caspase-8 could lead to the cleavage and activation of the BH3-only protein, BID, thereby initiating BAX/BAK-mediated apoptosis.7 This contradicted our previous findings and many other published results.4, 5, 6 This led us to re-visit the role of BH3-only proteins in ER stress-induced apoptosis. Testing the same HCT116 cell line as in Lu et al.7 (plus additional cell lines, such as MCF7 and primary MEFs), using CRISPR or conventional targeting to delete genes of interest and identical ER stress conditions as in Lu et al.,7 we sought to clarify the role of DR5, caspase-8 and BID in ER stress-induced apoptosis.
Results
The mitochondrial apoptotic pathway is upstream of caspase-8 activation during ER stress
Similar to what was reported previously by Lu et al.,7 we treated wild-type HCT116 and MCF7 cells with various ER stress-inducing agents. Efficient ER stress induction was confirmed by an increase in the expression of spliced XBP1 (XBP1s) and BiP (Figures 1a and b and Supplementary Figures S1a–c). In both cell lines, the pro-apoptotic BH3-only protein BIM was upregulated between 4 and 8 h of treatment, suggesting that the upregulation of BH3-only proteins during ER stress is an early event. It also confirmed that caspase-8 was activated in response to ER stress (Supplementary Figures S1a and b).7 Other ER stress-inducing agents, such as Subtilase cytotoxin (SubAB)8 or brefeldin A, were also used in HCT116 cells. Caspase-8 cleavage was seen after treatment with thapsigargin or SubAB and, albeit to a lesser extent, also after exposure to brefeldin A (Supplementary Figure S1c).

Caspase-8 activation during ER stress occurs downstream of BAX/BAK activation. (a) HCT116 cells were treated with thapsigargin (Tg) and harvested at the indicated time points and analysed by western blotting. (b) WT and Bax−/−/Bak−/− HCT116 cells were treated with thapsisgargin and analysed by western blotting. (c) Activated caspases were pulled down from the lysates with Biotin-X-zVAD-fmk12 and were identified by western blotting. Treatment with FAS ligand was used as a positive control for caspase-8 activation (upper panel). The experiment was repeated and blots were probed for BiP (marker of ER stress) and caspase-3 (bottom panel)
To test the possibility that caspase-8 may have been activated as a feedback by downstream effector caspases (caspase-3, -7) or caspase-9, the initiator caspase in the mitochondrial apoptotic pathway, HCT116 cells were treated with thapsigargin and samples were harvested at 2 h intervals. Increased levels of BiP and pro-apoptotic BIM were seen by 8 h posttreatment (Figure 1a). However, significant levels of cleaved caspase-8, -9 and -3 were only observed by 10–12 h posttreatment (Figure 1a). This suggested that BIM upregulation is an early step in ER stress-induced apoptosis that clearly precedes the cleavage of the initiator caspase-8 and -9 as well as the effector caspase-3. BIM induction can therefore not be a consequence of caspase-8 activation as suggested by Lu et al.7 However, early induction of Bim does not necessarily mean that the mitochondrial pathway is the cause of apoptosis and detection of proteins by western blots could also be influenced by the quality of the antibodies used. To clarify the chronology of events in cell death induction, cell lysates from WT and BAX−/−;BAK−/− HCT116 cells (lacking the essential effectors of the mitochondrial apoptotic machinery9) at various time points of treatment with ER stress-inducing agents were subjected to western blotting analysis (Figure 1b). Though ER stress was induced equally in both these cell lines as seen by BiP induction and Xbp1 splicing, caspase-9 and caspase-3 processing and PARP cleavage were impaired in BAX−/−;BAK−/− HCT116 cells. These lysates were further subjected to pull-down with the pan-caspase inhibitor, biotin-X-VAD-fmk, followed by detection of the activated caspases by immunoblotting.10 Although activated caspase-8, -9 and -3 could be pulled down from the WT HCT116 cells, none of these caspases could be detected in the BAX−/−/BAK−/− HCT116 cells (Figure 1c). This suggests that caspase-8 activation seen in the WT cells must occur downstream of the activation of the mitochondrial apoptotic pathway, that is, downstream of the activation of BAX/BAK and the initiator caspase-9.
Caspase-8 is dispensable for ER stress-induced apoptosis
To verify the role of the mitochondrial apoptotic pathway in ER stress-induced cell killing, we compared the apoptotic response of CASP8−/− cells with BAX−/−;BAK−/− cells (both MEFs and HCT116 cells). When treated with various agents, ER stress was induced in these cells within 24–48 h, as demonstrated by upregulation of BiP (Figures 2b and d). Similar to our observation in HCT116 cells, BAX−/−;BAK−/− MEFs did not activate caspase-3 in response to ER stress (Figure 2b). However, caspase-8 deficiency did not have any impact on caspase-3 activation either in MEFs or in HCT116 cells (Figures 2b and d). Consistent with these findings, the BAX−/−;BAK−/− cells were highly resistant to ER stress-induced apoptosis, whereas CASP8−/− cells were as sensitive as their wild-type counterparts both in short-term apoptosis assays and in long-term clonogenic survival assays (Figures 2e–g; Supplementary Figures S2a, b and e).As expected,11 the CASP8−/− HCT116 clones were protected against FASL-induced apoptosis and had normal levels of RIPK1, RIPK3 and MLKL (the major regulators of the necroptotic pathway), they had impaired Bid cleavage in response to ER stress (Supplementary Figures S3a–c). Taken together, these results suggest that caspase-8 activation and Bid cleavage are dispensable for ER-stress induced apoptosis in MEFs and HCT116 cells.

Combined BAX/BAK deficiency protects MEFs and HCT116 cells against ER stress-induced apoptosis, whereas caspase-8 deficiency does not. (a and c) WT, BAX−/−;BAK−/− and CASPASE-8−/− MEFs and HCT116 cells were analysed by western blotting to confirm the knockout status of these proteins. (b) WT, BAX−/−;BAK−/− and CASPASE-8−/− MEFs were treated with thapsisgargin (Tg; 100 nM) or tunicamycin (Tm; 1 μg/ml) for 24 h and cell lysates were analysed for BiP induction (ER stress marker), caspase-9 cleavage (disappearance of the full-length protein) and caspase-3 cleavage. (d) WT and CASPASE-8−/−HCT116 clones were subjected to ER stress and analysed by western blotting. (e) WT, BAX−/−;BAK−/− and CASPASE-8−/− MEFs were treated with the ER stress inducing agents Tg, Tm or SubAB (200 ng/ml) and analysed for cell death by flow cytometry after annexin V plus PI staining. (f) WT and BAX−/−;BAK−/− HCT116 cells were treated with the ER stress-inducing agents and analysed for apoptosis induction by annexin V and PI staining followed by flow cytometry. (g) WT and CASPASE-8−/−HCT116 cells were treated with the ER stress-inducing agents and analysed for apoptosis induction by annexin V and PI staining followed by flow cytometry. Error bars±S.E.M.; N=3; unpaired, two-tailed Student’s t-test. Numerical values on histograms indicate lack of statistical significance
The death receptor DR5 is dispensable for ER stress-induced apoptosis
To further investigate a possible role of the death receptor (extrinsic) apoptotic pathway in ER stress-induced cell killing, we examined BID-deficient MEFs and HCT116 cells. Although the loss of BID had no impact on ER stress-induced apoptosis in MEFs (both in short-term apoptosis assays and in clonogenic assays), it significantly reduced ER stress-induced caspase-3 activation and apoptosis in HCT116 cells, which had normal levels of BAX and BAK (Figures 3a–e; Supplementary Figures S2c and S3e), consistent with the findings of Lu et al.7 Lu et al. also had suggested an essential role for the death receptor, DR5, stimulated in a ligand-independent manner, in the activation of caspase-8 and BID, leading to ER stress-induced apoptosis. To verify the role of DR5 in ER stress-induced cell death, we generated DR5-deficient HCT116 cells using CRISPR/Cas9 system and treated them with thapsigargin. These cells displayed markers of ER stress, cleaved (activated) caspase-8 and -3 and underwent apoptosis at a rate comparable to the WT HCT116 cells (Figures 4a–d and Supplementary Figure S2d). The DR5 knockout was not accompanied by a significant increase in DR4 mRNA levels (Supplementary Figure S4). These results reveal that DR5 is not essential for ER stress-induced apoptosis in HCT116 cells.

Loss of BID protects HCT116 cells from ER stress-induced apoptosis. (a) WT and BID−/− MEFs and HCT clones were analysed by western blotting to confirm the knockout status of these genes. (b) WT and BID−/−MEFs were treated with ER stress-inducing agents and analysed by western blotting for BiP induction and caspase-3 activation. (c) WT and BID−/−HCT116 clones were treated with ER stress-inducing agents and analysed by western blotting for BiP induction and caspase-3 activation. (d) WT and BID−/− MEFs were treated with ER stress-inducing agents and analysed for induction of apoptosis by Annexin V and PI staining followed by flow cytometry. (e) WT and BID−/− HCT116 clones were treated with ER stress-inducing agents and analysed for induction of apoptosis by Annexin V and PI staining followed by flow cytometry. Error bars±S.E.M.; N=3; unpaired, two-tailed Student’s t-test. Numerical values on histograms indicate lack of statistical significance

DR5 deficiency does not protect HCT116 cells from ER stress-induced apoptosis. (a) DR5−/− HCT116 clones were analysed by western blotting to confirm the knockout. The DR5-specific protein band is indicated by an arrowhead. (b) HCT116 cells were treated with thapsigargin for various time points and analysed by western blotting for BiP, XBP1 splicing, DR5 and BIM levels and CASPASE-3 cleavage. (c) WT and DR5−/− HCT116 cells were treated for 24 h with Tg and analysed by western blotting for BiP induction and caspase-3 activation. (d) WT and DR5−/− HCT116 clones were treated for 24 or 48 h with Tg, SubAB or Tm and analysed for cell death by flow cytometry. Error bars±S.E.M.; N=3; unpaired, two-tailed Student’s t-test. Numerical values on histograms indicate lack of statistical significance
Several BH3-only proteins are critical for ER stress-induced apoptosis in HCT116 cells
Our results so far suggest the following: (1) Caspase-8 cleavage can occur in response to ER stress; (2) Caspase-8 is not essential for ER stress-induced killing of MEFs and HCT116 cells; (3) Caspase-8 cleavage (activation) is independent of DR5; (4) the BAX/BAK-dependent mitochondrial pathway is essential for ER stress-induced cell killing in both MEFs and HCT116 cells; (5) the BH3-only protein BID appears to be critical for ER stress-induced apoptosis in HCT116 cells but not in MEFs; and (6) BID cleavage does not appear to be necessary for ER stress-induced apoptosis in HCT116 cells. Using RNAi knockdown experiments, we have previously shown that BIM is essential for ER stress-induced apoptosis in a broad range of cell types, including MCF7, Vero cells and lymphocytes.4 To confirm our findings in MCF7 and extend them to the HCT116 cell line, we deleted the BIM gene using the CRISPR/Cas9 system. Loss of BIM provided MCF7 cells a significant protection from thapsigargin-induced apoptosis and greatly increased their clonogenic survival (Figures 5a–d). However, BIM deletion did not offer any protection in HCT116 cells (Figures 5e and f), in contrast to the complete or partial protection afforded by combined loss of BAX and BAK or loss of BID, respectively, in these cells. Absolute quantitation of transcript during ER stress in MEFs and HCT116 cells show that CHOP mRNA induction is accompanied by both BIM and PUMA mRNA induction (Supplementary Figure S5). These results suggests that BAX and BAK are essential for ER stress-induced apoptosis in all cells tested and BID is dispensable in most cells but, curiously, has a critical contributory (along with other BH3-only proteins) role in HCT116 cells.

Loss of BIM protects MCF7 cells but not HCT-116 cells from ER stress-induced apoptosis. (a) BIM−/− MCF7 clones generated by CRISPR/Cas9 were analysed by western blotting for the expression of the indicated proteins. (b) WT and BIM−/− MCF7 cells were treated with thapsigargin (100 nM) and visualized by phase-contrast microscopy. (c and d) WT and BIM−/− MCF7 cells were treated as in panel (b) and clonogenic survival was measured. (e) BIM−/− HCT116 clones generated by CRISPR/Cas9 were analysed by western blotting for the expression of the indicated proteins. (f) WT and BIM−/− HCT-116 cells were treated with Tg (100 nM), SubAB (200 ng/ml) and Tm (1 μg/ml) for the periods of 24 and 48 h and analysed for apoptosis by annexin V and PI staining followed by flow cytometry. Error bars±S.E.M.; N=3; unpaired, two-tailed Student’s t- test. Numerical values on histograms indicate lack of statistical significance
Discussion
In the present study, we used HCT116 cells (one of the two cell lines used by Lu et al.7) as well as MCF7 cells and MEFs, which we had both used in our previous study.4 Although Lu et al.7 suggested that BIM induction was a late event during ER stress-induced apoptosis, this was certainly not the case in our previous experiments,4 as well as in the present study. In fact, a marked increase in BIM protein was observed at the same time or just after the appearance of XBP1s and the induction of BiP (markers of ER stress), that is, well before cleaved caspase-8 could be detected. However, this does not necessarily mean that BIM is critical for apoptosis in these cells and this assertion was later found to be true in these cells (Figure 5). Although it is true that caspase-8 cleavage can be seen during ER stress (Figure 1a), this process occurs downstream of the activation of BAX/BAK and the initiator caspase-9 activation and is dispensable for ER stress-induced apoptosis. This is corroborated by our studies using caspase-8 deficient MEFs and HCT116 cells (Figure 2). Moreover, while in HCT116 cells, BID along with other BH3-only proteins appear to be necessary for apoptosis (Figures 4 and 5 and Supplementary Figure S5), the loss of the death receptor, DR5, had no impact on ER stress-induced UPR pathway activation, cleavage of caspase-8 and -3 and apoptosis (Figure 4). Finally, upon exposure to ER stress-inducing agents, BAX−/−BAK−/− MEFs and BAX−/−BAK−/− HCT116 cells did not display cleaved caspase-8, -9 or -3 and were fully protected from apoptosis. This underscores the importance of the mitochondrial apoptotic pathway in ER stress-induced cell killing and the absence of a role of death receptor-induced apoptosis in this process.
It is our (and many others’) long held view that the BH3-only proteins are critical initiators of ER stress-induced apoptosis.4, 5, 6 It appears that different BH3-only proteins are essential for ER stress-induced apoptosis in different cell types, that is, BIM is essential in MCF7 cells, whereas BID and other BH3-only proteins appear to have significant roles in HCT116 cells. As we did not detect any BID cleavage in CASP8−/− HCT116 cells and as these cells were normally susceptible to ER stress-induced apoptosis, BID must be activated through a caspase-8-independent process in response to ER stress. Cleavage of BID by caspases to its active form tBID was reported to be essential for its mitochondrial localization, activation of BAX/BAK and consequent induction of apoptosis downstream of death receptor stimulation.12, 13 It is, however, noteworthy that it has also been reported that BID in uncleaved form can induce apoptosis as potently as tBID.14
In conclusion, our results underscore the previously published results that several BH3-only proteins induce apoptosis in a cell-type-specific manner in response to ER stress;4, 5, 6 that is, BIM predominates in lymphoid cells, PUMA in neuronal cells and multiple BH3-only proteins in HCT116 cells. It will be of great interest to understand how BID is activated in response to ER stress in the HCT116 cells, but our studies reveal that this does not require DR5 or any death receptor-mediated activation of caspase-8.
Materials and methods
Culture conditions for mammalian cell lines
All cultures were grown in complete DMEM containing 10% FCS. HCT-116 cells were cultured in high-glucose DMEM (11995-065, Gibco, Thermo Fisher, Brendale, QLD, Australia) at 37 °C and 5% CO2. MCF-7, 293 T and MEF cells were cultured in normal glucose containing DMEM (11885-084, Gibco, Thermo Fisher) at 37 °C and 10% CO2.
Western blotting and immunoprecipitation
Western blotting and z-VAD-fmk pull-down analysis methods were described previously.10, 12, 15 A list of the antibodies that were used is given in Supplementary Table S2.
In all experiments, 5 × 106 wild-type HCT116 and Bax/Bak DKO cells were treated with Fc-FAS ligand (20 ng/ml) or thapsigargin (100 nM) and lysed in ONYX buffer (20 mM Tris/HCl pH 7.4, 135 mM NaCl, 1.5 mM MgCl2, 1 mM EGTA, 10% glycerol, 1% Triton X-100) supplemented with protease inhibitor cocktail tablets (Roche, Hawthorn, VIC, Australia, 04693132001) and phosphatase inhibitors β-glycerophosphate (5 mM), Na Molybdate (1 mM), Na pyrophosphate (2 mM) and NaF (10 mM). Active caspases from cell lysates were irreversibly bound to 2.5 mM of the pan-caspase inhibitor biotin-X-VAD-fmk (Calbiochem, Bayswater, VIC, Australia, 218742) for 30 min at 37 °C. Streptavidin-conjugated sepharose beads (ThermoFisher, 20347) were added as a 50% slurry in ONYX buffer and incubated for 45 min on a rotating wheel at 4 °C to capture biotin-X-VAD-fmk and bound active caspases. The sepharose beads were pelleted by centrifugation and the supernatant was recovered and mixed with reducing SDS-PAGE sample buffer for analysis of sample input. Beads were washed five times in ONYX buffer before boiling for 7 min in reducing SDS-PAGE sample buffer. Proteins in boiled samples were separated on SDS-PAGE gels, western blotted and blots were then probed for human caspase-3, -8 and -9 with the following antibodies: pAb anti-cleaved caspase-3 (Asp175) (Cell Signaling, Danvers, MA, USA, 9661), mAb anti-caspase-8 (MBL M032-3), and pAb caspase-9 (kindly provided by Dr. Lorraine O’Reilly). A complete list of antibodies used is given in Supplementary Table S1.
Generation of transgenic cell lines
CRISPR knockout cell lines were generated as described before.16 CRISPR guides were generated according to the MIT algorithm (http://crispr.mit.edu/). The guide sequences for each deletion are given in Supplementary Table S1. Lentivirally infected cells were sorted using a FACS Aria and gene deletions in cell populations were induced by adding 1 μg/ml doxycycline hyclate (D9891-5G, Sigma-Aldrich, Castle Hill, NSW, Australia). Clones with gene-specific deletions were confirmed by Next Gen Sequencing as described before.16 Next Gen Sequencing data on the knockout clones are shown in Supplementary Figure S3d. List of oligos used for gene knockouts are given in Supplementary Table S2.
Gene expression analysis
Absolute quantitation of transcript levels was determined by digital droplet PCR on a QX200 Droplet Digital System using EvaGreen ddPCR Supermix (Bio-Rad, Gladesville, NSW, Australia) as described before.16 The details of the primers used are given in Supplementary Table S3.
Drug treatment and apoptosis measurement
All cells were treated with identical concentrations of the ER stress-inducing agents thapsigargin (100 nM), brefeldin A (1 μg/ml), SubAB (200 ng/ml) and tunicamycin (1 μM) for all relevant experiments, replicating the conditions used by Lu et al.7 Treatment for death receptor apoptosis pathway activation was performed using TNFα (Fc-TNFα, 100 ng/ml) combined in solution with cycloheximide (2.5 μg/ml) or by Fc-FAS ligand (20 ng/ml). Apoptosis measurement was carried out by Annexin V/PI staining followed by flow cytometric analysis. In some cases, where there was interference by GFP/RFP fluorescence, we only stained with PI. Clonogenic survival assay was performed as described before.15
References
Weston RT, Puthalakath H . Endoplasmic reticulum stress and BCL-2 family members. Adv Exp Med Biol 2010; 687: 65–77.
Bravo R, Vicencio JM, Parra V, Troncoso R, Munoz JP, Bui M et al. Increased ER-mitochondrial coupling promotes mitochondrial respiration and bioenergetics during early phases of ER stress. J Cell Sci 2011; 124 (Pt 13): 2143–2152.
Kaufman RJ . Orchestrating the unfolded protein response in health and disease. J Clin Invest 2002; 110: 1389–1398.
Puthalakath H, O'Reilly LA, Gunn P, Lee L, Kelly PN, Huntington ND et al. ER stress triggers apoptosis by activating BH3-only protein Bim. Cell 2007; 129: 1337–1349.
Reimertz C, Kogel D, Rami A, Chittenden T, Prehn JH . Gene expression during ER stress-induced apoptosis in neurons: induction of the BH3-only protein Bbc3/PUMA and activation of the mitochondrial apoptosis pathway. J Cell Biol 2003; 162: 587–597.
Li JLB, Lee AS . Endoplasmic reticulum stress-induced apoptosis: multiple pathways and activation of p53-up-regulated modulator of apoptosis (PUMA) and NOXA by p53. J Biol Chem 2006; 281: 7260–7270.
Lu M, Lawrence DA, Marsters S, Acosta-Alvear D, Kimmig P, Mendez AS et al. Cell death. Opposing unfolded-protein-response signals converge on death receptor 5 to control apoptosis. Science 2014; 345: 98–101.
Paton AW, Beddoe T, Thorpe CM, Whisstock JC, Wilce MC, Rossjohn J et al. AB5 subtilase cytotoxin inactivates the endoplasmic reticulum chaperone BiP. Nature 2006; 443: 548–552.
Lindsten T, Ross AJ, King A, Zong WX, Rathmell JC, Shiels HA et al. The combined functions of proapoptotic Bcl-2 family members bak and bax are essential for normal development of multiple tissues. Mol Cell 2000; 6: 1389–1399.
Jost PJ, Grabow S, Gray D, McKenzie MD, Nachbur U, Huang DC et al. XIAP discriminates between type I and type II FAS-induced apoptosis. Nature 2009; 460: 1035–1039.
Varfolomeev EE, Schuchmann M, Luria V, Chiannilkulchai N, Beckmann JS, Mett IL et al. Targeted disruption of the mouse Caspase 8 gene ablates cell death induction by the TNF receptors, Fas/Apo1, and DR3 and is lethal prenatally. Immunity 1998; 9: 267–276.
Kaufmann T, Jost PJ, Pellegrini M, Puthalakath H, Gugasyan R, Gerondakis S et al. Fatal hepatitis mediated by tumor necrosis factor TNFalpha requires caspase-8 and involves the BH3-only proteins Bid and Bim. Immunity 2009; 30: 56–66.
Puthalakath H, Strasser A . Keeping killers on a tight leash: transcriptional and post-translational control of the pro-apoptotic activity of BH3-only proteins. Cell Death Differ 2002; 9: 505–512.
Sarig R, Zaltsman Y, Marcellus RC, Flavell R, Mak TW, Gross A . BID-D59A is a potent inducer of apoptosis in primary embryonic fibroblasts. J Biol Chem 2003; 278: 10707–10715.
Gangoda L, Doerflinger M, Srivastava R, Narayan N, Edgington LE, Orian J et al. Loss of Prkar1a leads to Bcl-2 family protein induction and cachexia in mice. Cell Death Differ 2014; 21: 1815–1824.
Aubrey BJ, Kelly GL, Kueh AJ, Brennan MS, O'Connor L, Milla L et al. An inducible lentiviral guide RNA platform enables the identification of tumor-essential genes and tumor-promoting mutations in vivo. Cell Rep 2015; 10: 1422–1432.
Acknowledgements
JG, CN and GM are supported by LTU PhD scholarship and this project was funded by an Australian National Health and Medical Research Council Project grant APP1085328 (to HP).
Author contributions
JG, MD, CN, IJ, GM and GB conducted the experiments. AK, DS, MJ and DH generated the CRISPR knockout cell lines. MJ, DH, AP, JP and HP designed the experiments and MD, JG and HP wrote the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Edited by G Melino
Supplementary Information accompanies this paper on Cell Death and Differentiation website
Rights and permissions
About this article

Cite this article
Glab, J., Doerflinger, M., Nedeva, C. et al. DR5 and caspase-8 are dispensable in ER stress-induced apoptosis. Cell Death Differ 24, 944–950 (2017). https://doi.org/10.1038/cdd.2017.53
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cdd.2017.53
This article is cited by
-
IRE1 RNase controls CD95-mediated cell death
EMBO Reports (2024)
-
When cell death goes wrong: inflammatory outcomes of failed apoptosis and mitotic cell death
Cell Death & Differentiation (2023)
-
Negligible role of TRAIL death receptors in cell death upon endoplasmic reticulum stress in B-cell malignancies
Oncogenesis (2023)
-
Mast cells inhibit colorectal cancer development by inducing ER stress through secreting Cystatin C
Oncogene (2023)
-
Stress-induced inflammation evoked by immunogenic cell death is blunted by the IRE1α kinase inhibitor KIRA6 through HSP60 targeting
Cell Death & Differentiation (2022)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}