
Abstract
This study demonstrates, for the first time, that loss of a single forkhead box class O (FoxO) transcription factor, can promote lymphomagenesis. Using two different mouse models, we show that FoxO3 has a significant tumour-suppressor function in the context of Myc-driven lymphomagenesis. Loss of FoxO3 significantly accelerated myeloid tumorigenesis in vavP-MYC10 transgenic mice and B lymphomagenesis in Eμ-myc transgenic mice. Tumour analysis indicated that the selective pressure for mutation of the p53 pathway during Eμ-myc lymphomagenesis was not altered. Frank tumours were preceded by elevated macrophage numbers in FoxO3−/− vavP-MYC10 mice but, surprisingly, pre-B-cell numbers were relatively normal in healthy young FoxO3−/−Eμ-myc mice. In vitro assays revealed enhanced survival capacity of Myc-driven cells lacking FoxO3, but no change in cell cycling was detected. The loss of FoxO3 may also be affecting other tumour-suppressive functions for which FoxO1/4 cannot fully compensate.
Similar content being viewed by others


Main
The evolutionarily conserved forkhead box class O (FoxO) transcription factors have important roles in cellular metabolism, stress tolerance and probably lifespan.1, 2 They are key regulators of cell proliferation and survival, a large number of their transcriptional targets being involved in apoptosis,3, 4, 5 cell cycle arrest,6, 7 DNA repair, oxidative stress resistance and metabolic processes.8, 9
Regulation of FoxOs is complex, involving acetylation and ubiquitination as well as phosphorylation (reviewed in Eijkelenboom and Burgering10). Under normal conditions of growth factor signalling, FoxO transcription factors are inactivated through phosphorylation by activated AKT. For FoxO3 this occurs at three conserved residues (Thr-32, Ser-253 and Ser-315), resulting in the export of FoxO3 from the nucleus into the cytoplasm11 and proteasomal degradation.12 During conditions of oxidative stress, nuclear translocation and activation of FoxOs occurs through Jun N-terminal kinase phosphorylation, which overrides growth factor signalling.13
The FoxOs are highly related and to some degree can act redundantly as they bind to the same consensus sequence.14 Of the four mammalian FoxO family members, FoxO1, FoxO3 and FoxO4 are widely expressed,14 whereas FoxO6 is mainly expressed in adult brain tissue.15 In haemopoietic tissues, FoxO1 and FoxO3 are the major FoxOs, with FoxO1 levels highest in lymphoid cells and FoxO3 levels highest in myeloid cells.16
Although FoxO4−/− mice do not have an obvious phenotype,17 FoxO6−/− mice show impaired memory consolidation18 and deletion of FoxO1 results in defects in embryonic vascular development, resulting in death at embryonic day 10.5.17, 19 FoxO3-deficient mice display early onset female infertility because of depletion of ovarian follicles caused by widespread follicular activation.17, 20 Aged FoxO3−/− mice were found to have reduced numbers of haemopoietic stem cells (HSCs),21 implying that FoxO3 contributes to the maintenance of the HSC pool during aging by reducing oxidative stress.22 FoxO3 primes HSCs for metabolic stress (starvation)-induced autophagy, which is necessary for HSC survival.23
Activation of AKT, and thus inactivation of FoxOs, is frequently observed in a range of cancers,24 raising the possibility that FoxO proteins may serve as tumour suppressors. An early study found that loss of individual FoxOs did not cause cancer predisposition, except for loss of FoxO1.17, 20 When FoxO1 was deleted using Mx-Cre (at 4 weeks of age), mice developed systemic hemangiomas, but with long latency and low frequency.25 However, compound somatic deletion of three FoxOs, FoxO1/3/4, resulted in thymic lymphomagenesis.25 Furthermore, a dominant-negative FoxO construct, which expresses the conserved DNA-binding domain of FoxO4 but lacks the transactivation domain and thus inhibits the activity of all FoxOs, greatly accelerated the onset of Eμ-myc lymphomas.26 This effect is due, at least in part, to FoxO upregulation of p19ARF expression, which promotes p53-dependent apoptosis. In fact, the dominant-negative FoxO Eμ-myc tumours arose as rapidly as those on a p53+/− background.26 FoxO3 and MYC compete for binding to some promoters and appear to antagonise each other’s activity.27, 28, 29 FoxO3 directly regulates expression of the MYC inhibitor MXI1, whereas other Max-interacting protein (MXD) family members are regulated indirectly.30
To determine whether loss of a single FoxO can cooperate in Myc-driven tumorigenesis, we crossed FoxO3−/− mice21 with vavP-MYC10 mice, which express the transgene in all haemopoietic lineages and primarily develop myeloid tumours with some T lymphomas,31, 32 and with Eμ-myc mice, which constitutively express Myc in B lymphoid cells and are susceptible to pre-B and B lymphomas.33, 34
Results
Loss of FoxO3 accelerates Myc-driven tumorigenesis
vavP-MYC10 (hereafter MYC10) transgenic mice develop macrophage tumours with a median onset of approximately 300 days. Lymphomagenesis was significantly accelerated by the loss of FoxO3, all FoxO3−/−MYC10 mice having succumbed to disease by 300 days compared with 400 days for the MYC10 mice (Figure 1a) (median survival of FoxO3−/−MYC10 256 days versus 295 days for MYC10; P<0.0001). Comparison of spleen weights and white blood cell (WBC) counts of tumour-bearing mice showed that the tumour burden was higher in the FoxO3−/−MYC10 mice (Figures 1b and c). The tumour phenotype (Figure 1d) was comparable between the cohorts, being predominantly monocyte/macrophage tumours (Mac1+F4/80+) with a small proportion of T-cell tumours (Thy1+CD3+ and variable CD4 and CD8 expression), as previously reported for MYC10 mice.32 Tumours of all genotypes were transplantable in C57BL/6 recipients (n=9–12 independent tumours).

Loss of FoxO3 accelerates Myc-induced myeloid tumorigenesis. (a) Kaplan–Meier survival curves for MYC10 (n=32, median survival 295 days), FoxO3+/−MYC10 (n=35, median survival 270 days), FoxO3−/−MYC10 (n=33, median survival 256 days) and FoxO3−/− (n=24). Non-tumour-related deaths have been censored. Tumorigenesis is significantly accelerated in FoxO3−/−MYC10 versus MYC10 mice, P<0.0001, log-rank test. (b) Spleen weights and (c) WBC counts of sick mice. Leukocytes were elevated in FoxO3−/−MYC10 mice, **P<0.01, Student’s t-test. (d) Proportions of myeloid and T-cell tumours in sick mice. Tumour cell suspensions were stained for cell surface markers and analysed by FACS. n=22–32
When we assessed the impact of FoxO3 loss on lymphomagenesis in Eμ-myc mice, again we found a significant acceleration in morbidity. The median survival of FoxO3−/−Eμ-myc mice was 93 versus 125 days for Eμ-myc mice (P<0.0001), with no FoxO3−/−Eμ-myc mice surviving beyond 150 days (Figure 2a). Tumour-bearing mice typically presented with enlarged spleen, lymph nodes and/or thymus, with no significant difference in lymphoma burden between genotypes (Figures 2b and c). Although Eμ-myc mice generally succumb to sIgM−B220+ pre-B lymphomas or sIgM+B220+ B-cell lymphomas,33, 34 certain crosses have led to changes in tumour phenotype, either predominantly B-cell lymphomas (e.g., in p53+/−/Eμ-myc and vavP-Mcl-1/Eμ-myc mice),35, 36 or a complete change in immunophenotype (e.g., in Eμ-bcl-2/ Eμ-myc and Eμ-v-abl/ Eμ-myc mice).37, 38 However, immunophenotyping of the FoxO3−/−Eμ-myc lymphomas revealed no major difference in the proportion of pre-B versus B-cell tumours compared with Eμ-myc lymphomas (Figure 2d).

Loss of FoxO3 accelerates Myc-induced lymphomagenesis. (a) Kaplan–Meier survival curves for Eμ-myc (n=33, median survival 125 days), FoxO3+/− Eμ-myc (n=59, median survival 101 days) and FoxO3−/− Eμ-myc (n=23, median survival 93 days). Non-tumour-related deaths have been censored. Lymphomagenesis is significantly accelerated by loss of FoxO3. FoxO3−/− Eμ-myc versus Eμ-myc mice, P<0.0001; FoxO3+/− Eμ-myc versus Eμ-myc mice, P=0.0502; FoxO3−/− Eμ-myc versus FoxO3+/− Eμ-myc, P=0.0076; log-rank test. (b) Spleen weights and (c) WBC counts of sick mice. (d) Proportions of pre-B, mixed (pre-B/B), B and other lymphoid (T or primitive) tumours in sick mice; determined by staining for cell surface markers and FACS analysis. n=20–42
Haemopoietic homeostasis is perturbed by loss of FoxO3
To determine whether loss of FoxO3 alters the preneoplastic phenotype of MYC10 mice, which could contribute to the enhanced tumorigenesis, we compared the composition of blood and haemopoietic tissues of healthy mice at 8 weeks of age (Figure 3, Supplementary Figure 1 and Supplementary Table 1).

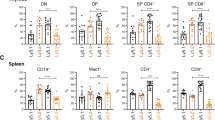
Loss of FoxO3 increases myeloid and T-cell populations in MYC10 mice. Enumeration of total leukocytes and indicated populations in the spleen of preneoplastic 8-week-old male mice (n=8–9 per genotype). Bars represent mean±S.E.M.; statistical significance is shown only for WT versus FoxO3−/−, MYC10 versus FoxO3−/−MYC10 and FoxO3−/− versus FoxO3−/−MYC10 (*P<0.05, **P<0.01, ***P<0.001, Student’s t-test). See also Supplementary Table S1 and Supplementary Figure 1
Perhaps surprisingly, loss of FoxO3 alone had a greater impact on haemopoietic homeostasis than expression of the MYC10 transgene, which provokes only a mild preneoplastic phenotype.32 The FoxO3−/− mice had significantly increased T cells in the spleen and blood, but not in lymph nodes. Herold et al.39 have also reported an increase in splenic T cells using the same FoxO3−/− mice, although increased T-cell numbers were not observed in an earlier study using a different line.40 Consistent with previous reports, the FoxO3−/− mice also had increased myeloid cells in the spleen and bone marrow,21, 41, 42 reduced recirculating B cells in the bone marrow43 and elevated immature erythroid cells (Ter119+), presumably due to impaired maturation.44, 45
In general, the phenotype of FoxO3−/−MYC10 mice resembled that of FoxO3−/− mice. However, in the spleen, there was an additional increase in the number of T cells in FoxO3−/−MYC10 mice compared with FoxO3−/− mice (Figure 3). Significant increases in myeloid populations were also observed in the blood, spleen and bone marrow, in particular Mac1+Gr1− cells, which were elevated approximately 2- to 2.5-fold in each organ (Figure 3, Supplementary Figure 1). As the majority of tumours arising in the MYC10 mice are Mac1+Gr1−, it is likely that the increase in this population in FoxO3−/−MYC10 mice is a factor in the acceleration of tumour onset.
Haemopoietic analysis of pre-leukaemic FoxO3−/−Eμ-myc mice and littermates was performed at 4 weeks of age (Figure 4, Supplementary Figure 2 and Supplementary Table 2). Loss of FoxO3 resulted in increased myeloid cells in the blood, spleen and bone marrow of Eμ-myc mice, as in MYC10 mice, but no significant differences were seen in T-cell numbers. As Eμ-myc and FoxO3−/−Eμ-myc mice develop pre-B or B-cell lymphomas, the impact of FoxO3 loss on B lymphopoiesis was of particular interest. As documented previously,46 young preneoplastic Eμ-myc mice had a greatly expanded pre-B-cell (B220+IgM−IgD−) population, with 15- to 20-fold increases in the spleen, blood and lymph nodes, and there was a modest further increase in the blood of FoxO3−/−Eμ-myc mice.

Loss of FoxO3 increases myeloid and B-cell populations in the blood of Eμ-myc mice. Total WBC count and quantification of the indicated populations in the blood of preneoplastic 4-week-old male mice (n=8–9 per genotype). Bars represent mean±S.E.M.; statistical significance is shown only for WT versus FoxO3−/− and Eμ-myc versus FoxO3−/− Eμ-myc (*P<0.05, **P<0.01, ***P<0.001, Student’s t-test). See also Supplementary Table S2 and Supplementary Figure 2
Cell cycle analysis
Deregulation of c-myc promotes tumorigenesis through altered expression of genes regulating cell cycling and proliferation.47 Loss of FoxO may be expected to amplify the cell cycle impact of myc, as FoxO3 regulates genes that promote cell cycle arrest.6, 48 We tested this possibility in preneoplastic populations, to avoid the complication of additional oncogenic mutations.
As the MYC10 and FoxO3−/−MYC10 mice are predisposed to myeloid tumours, we analysed the cell cycle profiles of bone marrow macrophages and granulocytes from healthy young mice. No differences in the cell cycle distribution were detectable in either Mac1+Gr1− or Mac1+Gr1+ cells (Figure 5).

Loss of FoxO3 has no impact on the cycling of preneoplastic MYC10 myeloid cells. Bone marrow from MYC10 and FoxO3−/−MYC10 mice was surface stained with Mac1 and Gr1 before fixation. Cell cycle analysis of macrophages (Mac1+Gr1−) and granulocytes (Mac1+Gr1+) was then performed following incubation with PI/RNase staining solution. Data represent mean±S.E.M.; n= 6–7 per genotype
For the Eμ-myc and FoxO3−/− Eμ-myc mice, preneoplastic pre-B cells were analysed immediately after isolation from the bone marrow and also during culture in simple medium lacking exogenous cytokines. However, there was no difference between Eμ-myc and FoxO3−/−Eμ-myc pre-B-cell cycle profiles either before culture (first panel, Figure 6a) or at any of the time points analysed (4 h, 8 h, 24 h; Figure 6a). Furthermore, tumour cells isolated from Eμ-myc and FoxO3−/−Eμ-myc mice had similar cell cycle profiles (Figure 6b).

Loss of FoxO3 has no impact on the cycling of preneoplastic and neoplastic Eμ-myc pre-B cells. (a) Pre-B cells (B220+sIg−) were purified from the bone marrow of 4-week-old Eμ-myc and FoxO3−/− Eμ-myc mice by flow cytometry and cultured without cytokines. Cell cycle analysis was performed using Nicoletti staining at the indicated time points and flow cytometry. Cell cycle phases were determined using the Watson pragmatic model within the flow cytometry data analysis software FlowJo. Bars represent mean±S.E.M.; n= 4–5 per genotype. (b) Pre-B/B cells were purified from tumours of Eμ-myc and FoxO3−/− Eμ-myc mice using CD19 beads and MACS columns. Cells were fixed then stained with PI/RNase staining solution. Cell cycle populations were analysed by FACS. Bars represent mean±S.E.M.; n= 5 per genotype
Impact of loss of FoxO3 on apoptosis of Myc-overexpressing cells
The proliferative effect of Myc overexpression is partially counteracted by Myc-driven apoptosis.49, 50 We therefore investigated whether loss of FoxO3 inhibited apoptosis of cells overexpressing Myc. We reasoned that the impact of loss of FoxO3 may become more apparent during culture, as the deficiency in cytokine signalling would normally inactivate AKT and activate FoxO3.
As expression of the vavP-MYC transgene is highest in thymocytes,32 we isolated the four major thymocyte populations by flow cytometry from wild-type (WT), FoxO3−/−, MYC10 and FoxO3−/− MYC10 mice and cultured them in simple medium (i.e., in the absence of cytokines) either immediately (Supplementary Figure 3A) or following exposure to 1.25 Gy γ-irradiation (Supplementary Figure 3B). For double-positive thymocytes, the most sensitive population, there was a trend toward an increased viability of FoxO3−/− compared with WT cells under these conditions, but this was not apparent on the MYC10 background. Loss of FoxO3 also enhanced the survival of peripheral T-cell blasts following removal of interleukin 2 (IL-2), as noted previously,39 and this was evident even in the face of expression of the MYC10 transgene (Supplementary Figure 3C).
Apoptosis was also assessed for B lymphoid cells from Eμ-myc and FoxO3−/−Eμ-myc mice. Frank tumours had comparable numbers of apoptotic cells (Figure 7a and Supplementary Figure 4). When pre-leukaemic pre-B cells sorted from bone marrow were cultured in the absence of cytokines, no significant survival advantage could be detected for those lacking FoxO3 (Figure 7b). However, when IL-7 was removed from IL-7-supported cultures of pre-B cells, loss of FoxO3 significantly enhanced the survival of Eμ-myc (but not WT) pre-B cells (Figure 7c). As IL-7 has a critical role in early B lymphopoiesis, this protective effect may account for the modest increase in pre-B cells in FoxO3−/−Eμ-myc compared with Eμ-myc mice (Figure 4, Supplementary Table 2).

Impact of loss of FoxO3 on the apoptosis of preneoplastic and neoplastic Eμ-myc cells. (a) Quantification of cleaved caspase-3 positive cells in the spleen and lymph node of sick Eμ-myc and FoxO3−/− Eμ-myc mice. For each tumour section (blinded as to the genotype), cleaved caspase-3-positive cells were enumerated from images of three independent fields of view at x200 magnification. Bars represent mean±S.E.M.; n=4–6 per genotype. (b) Spontaneous death of FACS purified preneoplastic pre-B cells (B220+sIg-) from 4-week-old Eμ-myc and FoxO3−/− Eμ-myc mice cultured in simple medium (no cytokines) was assessed by staining with PI and annexin V followed by FACS analysis (n=5–6 per genotype). Values represent mean±S.E.M. (c) Enhanced survival upon IL-7 withdrawal of cultured FoxO3−/− Eμ-myc pre-B cells. Pre-B cells isolated from bone marrow of mice of the indicated genotypes were cultured in the presence of 2% IL-7 supernatant for 5 days. Survival following removal of IL-7 was determined at the indicated time points by flow cytometry, with data normalised to the viability of each culture when IL-7 was withdrawn, day 0. Bars represent mean±S.E.M.; n=3–4 mice for each genotype. FoxO3−/− Eμ-myc pre-B cells were significantly more viable than Eμ-myc pre-B cells on day 1 (P=0.0028) and day 2 (P=0.0250). *P<0.05, **P<0.01, ***P<0.001, Student’s t-test
Loss of FoxO3 does not select against inactivation of the p53 pathway
Lymphomas arising in Eμ-myc mice commonly bear mutations that inactivate the p19Arf/Mdm2/p53 pathway, either by p53 mutation or deletion, p19Arf deletion, or overexpression of Mdm2.51 Although expression of dominant-negative FoxO (dnFoxO) reduced selective pressure to inactivate the p19Arf/p53 pathway during lymphomagenesis in Eμ-myc mice,26 this was not the case for FoxO3−/−Eμ-myc lymphomas (Figure 8). One FoxO3−/−Eμ-myc tumour (# 81) had high levels of p53 and p19Arf, suggestive of a p53 inactivating mutation (as carried by the positive control, Eμ-myc lymphoma #22). Certain other FoxO3−/−Eμ-myc lymphomas (#322 and #430) had high levels of p19Arf but expressed no p53, apparently because of upregulation of Mdm2. Two, perhaps three, other FoxO3−/−Eμ-myc lymphomas lacking p53 expression had elevated Mdm2 but did not express p19Arf (#124, #240 and perhaps #94), suggestive of p19Arf deletion, as observed previously.51 Thus, overall, 5 or 6 of 13 (38–46%) FoxO3−/−Eμ-myc lymphomas analysed showed evidence of loss of p53 or Arf function, comparable to the 13/25 (52%) reported by Eischen et al.51 Thus, the absence of FoxO3 alone appears insufficient to select against inactivation of the p53 pathway.

Loss of FoxO3 does not relieve the pressure for mutation of the p53 pathway in Eμ-myc lymphomagenesis. Western blot analysis of p53, p19ARF and Mdm2 in the indicated Eμ-myc and FoxO3−/− Eμ-myc lymphomas; Eμ-myc #22 (right-hand lane on each gel), a known p53 mutant cell line, was included as a positive control. MW markers are indicated (kDa). High expression of p19ARF is indicative of p53 mutation (e.g., FoxO3−/− Eμ-myc #81) or loss (e.g., FoxO3−/− Eμ-myc #322, #430 and Eμ-myc #97). High expression of Mdm2 promotes ubiquitinylation and degradation of p53, thereby abrogating p53-mediated apoptosis
Discussion
We have demonstrated, for the first time, that loss of a single FoxO transcription factor, can promote lymphomagenesis. Whereas Paik et al.25 found that complete loss of FoxO activity (both alleles of FoxO1, FoxO3 and FoxO4) is required for lymphomagenesis, we have shown using two different mouse models that loss of FoxO3 alone suffices to accelerate Myc-driven lymphomagenesis. In the absence of FoxO3, myeloid tumours arose more rapidly in vavP-MYC10 mice and pre-B and B lymphomas arose more rapidly in Eμ-myc transgenic mice (Figures 1 and 2). Unsurprisingly, the reduction in tumour latency was not as great as that observed when a dominant-negative FoxO construct was used to inhibit all FoxOs in the Eμ-myc model.25 Deficiency in a single FoxO does not have an observable effect in all tumour models, however, as loss of FoxO3 or FoxO4 does not accelerate or increase the incidence of DMBA-induced tumorigenesis.25
The larger population of myeloid cells in FoxO3-deficient MYC10 mice is likely to be a significant factor contributing to accelerated myeloid tumour formation in this model (Figure 3,Supplementary Figure 1 and Supplementary Table 1). In the Eμ-myc model, expansion of the pre-B-cell pool, although modest, may contribute to accelerated lymphoma development in the absence of FoxO3.
Several factors may contribute to increasing the size of the susceptible populations. The absence of FoxO3 had no discernible impact on the cycling of pre-leukaemic myeloid cells isolated from MYC10 mice (Figure 5) or of pre-leukaemic or neoplastic B lymphoid cells taken from Eμ-myc mice (Figure 6). Presumably other FoxOs can compensate in this role – FoxO1, in particular, was readily detected in lymphoid cells from Eμ-myc and vavP-MYC mice (Supplementary Figure 5), as reported previously for WT lymphoid cells.16 Previous studies have reported that FoxO3-deficiency increased the viability of mast cells and thymocytes cultured in the absence of cytokines39, 52 and we were able to demonstrate enhanced viability of certain T-cell populations from vavP-MYC10 mice, particularly after cytokine removal (Supplementary Figure 3). Similarly, lack of FoxO3 enhanced the viability of cultured Eμ-myc pre-B cells following IL-7 removal (Figure 7c). Reduced apoptosis has also been documented in dnFoxO-expressing Eμ-myc lymphoma cells.26
Activation of p53 is a principal route for Myc-induced apoptosis. Myc upregulates p19Arf expression, probably indirectly, and p19Arf binds to and neutralises Mdm2, thereby preventing inhibition and degradation of p53 and facilitating expression of pro-apoptotic p53 targets, BH3-only proteins Puma and Noxa and, in a negative feedback loop, Mdm2.53 Although expression of dnFoxO reduced the selective pressure to inactivate the p19Arf/p53 pathway during lymphomagenesis in Eμ-myc mice,26 this was not the case for FoxO3−/−Eμ-myc lymphomas (Figure 8).
In summary, we have demonstrated using two different mouse models that FoxO3 has a significant tumour-suppressor function in the context of Myc-driven lymphomagenesis. Although our data suggest a role for FoxO3-mediated inhibition of apoptosis in restraining Myc-driven tumour development, we suspect that loss of FoxO3 may also be affecting other tumour-suppressive functions for which FoxO1/4 cannot fully compensate. Of note, the tumour-suppressive activity of p53 does not involve its capacity to promote apoptosis or cell cycle arrest.54, 55
Materials and Methods
Mice
Experimental protocols involving the use of mice were approved by the Walter and Eliza Hall Institute’s Animal Ethics Committee. All mice were on a C57BL/6 background and were bred at the Walter and Eliza Hall Institute (WEHI). To generate FoxO3−/−MYC10 mice, FoxO3+/− (ref. 21) females were crossed with MYC10hom32 males (i.e., homozygous for the MYC10 transgene) then offspring were interbred. FoxO3−/− Eμ-myc mice were produced by mating FoxO3+/− females with Eμ-myc33 males followed by interbreeding of offspring. Analysis was performed on healthy young mice (4 or 8 weeks of age) and cohorts of mice were aged to ethical end point.
Haemopoietic analysis
Blood counts and composition were determined using an ADVIA 2120 haematology analyser (Siemens, Erlangen, Germany). In addition, for preneoplastic analysis, the remaining blood was depleted of red blood cells using 0.168 M ammonium chloride and cell composition determined by flow cytometry, as below. Single-cell suspensions were prepared from spleen, thymus, lymph nodes (axillary, brachial, inguinal) and bone marrow and leukocytes enumerated with a CASY Cell Counter (Scharfe System GmbH, Reutlingen, Germany). Cell composition was determined by staining with fluorochrome-labelled surface marker-specific monoclonal antibodies followed by fluorescence-activated cell sorting (FACS) analysis using an LSRI (BD Biosciences, Franklin Lakes, NJ, USA). Data were processed using FlowJo Version 9.3.2 (TreeStar, Ashland, OR, USA). The monoclonal antibodies, produced and labelled with fluorescein isothiocyanate (FITC), R-phycoerythrin or allophycocyanin at WEHI, were: RB6-8C5, anti-Gr1; MI/70, anti-Mac1; H129.19, anti-CD4; YTS169, anti-CD8; Ter119, anti-erythroid marker; RA3-6B2, anti-CD45R-B220; 5.1, anti-IgM; 11-26C, anti-IgD; T24-31, anti-Thy1.
Cell cycle analysis
Pre-B cells (B220+IgM−IgD−) were purified from the bone marrow of 4-week-old mice by flow cytometry, or by using CD19 MicroBeads (Miltenyi Biotech, Bergisch Gladbach, Germany) and MS columns (Miltenyi Biotech) according to the manufacturer’s protocols. The Nicoletti assay56 was used for cell cycle analysis of the purified pre-B cells – cells were resuspended in 0.1% sodium citrate, 0.1% Triton X and 50 μg/ml propidium iodide (PI) and incubated for 30 min on ice before FACS analysis. Alternatively, mixed cell populations, such as bone marrow, were stained with surface markers for the population of interest before fixation and permeabilisation with the Transcription Factor Staining Buffer Set (eBioscience, San Diego, CA, USA). Cells were then stained with PI/RNase staining solution (Cell Signaling Technology, Danvers, MA, USA) for 30 min before analysis on an LSR II flow cytometer (BD Biosciences). Cell cycle profiles were characterised using FlowJo using the Watson pragmatic model (see above).
Survival assays
Thymocyte populations or pre-B cells isolated by flow cytometry were cultured at 0.2–0.5x106 cells/ml in high-glucose Dulbecco’s Modified Eagle’s medium supplemented with 10% foetal calf serum (JRH Biosciences, Brooklyn, VIC, Australia), 50 μM 2-mercaptoethanol (2-ME; Sigma-Aldrich, St Louis, MO, USA) and 100 μM asparagine (Sigma-Aldrich) without additional cytokines. In addition, thymocytes were treated before culture with 1.25 Gy γ-irradiation. Cell viability was determined by staining with FITC-conjugated annexin V and PI followed by flow cytometry. Specific viability was calculated at each time point as (viability of treated cells/viability of untreated cells) x 100%.
Cytokine withdrawal
Pre-B cells (B220+IgM−IgD−) purified from bone marrow by flow cytometry were cultured at a starting density of 1x106/ml in Iscove’s modified Dulbecco’s medium supplemented with 1x GlutaMAX, 1 mM sodium pyruvate, 0.1 mM nonessential amino acids, 10 mM HEPES pH 7.4 (all media supplements from Life Technologies, Carlsbad, CA, USA), 50 μM 2-ME, 10% foetal calf serum and 2% mIL-7 supernatant (produced by transfected x63/0 hybridoma cells).57 Cultures were split if required. After 5 days, cells were washed three times to remove IL-7, then replated at 0.3x106/ml with or without IL-7 for 2 days. Viability was determined by flow cytometry as above.
T-cell blasts were generated by stimulation of splenocytes (2x106/ml) for 3 days with 1% mIL-2 supernatant57 and 2 μg/ml concanavalin A (Sigma-Aldrich) in cell culture medium as for survival assays above. Cells were then washed three times to remove concanavalin A and cultured with IL-2 alone for 24 h. To set up the death assay, cells were washed three times to remove IL-2 then cultured with or without IL-2 for 2 days. Viability was determined by flow cytometry as above.
Cleaved caspase-3 immunohistochemistry
Spleens and lymph nodes from lymphoma-bearing mice were fixed in 10% formalin then embedded in paraffin. Sections were stained for cleaved caspase-3 using the SignalStain Apoptosis (Cleaved Caspase-3) IHC Detection Kit (Cell Signaling Technology) according to the manufacturer’s protocol. Stained slides were imaged (blinded as to genotype) using an Olympus BX43 microscope (Olympus, Tokyo, Japan) and Olympus DP72 camera at x200 magnification.
Immunoblotting
Protein extracts were prepared by lysis in RIPA buffer (300 mM NaCl, 2% IGEPAL CA-630, 1% deoxycholic acid, 0.2% SDS, 100 mM Tris-HCl pH 8.0) containing complete ULTRA protease inhibitors (Roche, Basel, Switzerland). Western blots were carried out using 30 μg total protein per sample run on NuPAGE Bis-Tris gels (Life Technologies) and transferred to nitrocellulose membranes with an iBlot (Life Technologies) according to the manufacturer’s protocols. Blots were probed with the following antibodies: p53 (FL-393, Santa Cruz Biotechnology, Santa Cruz, CA, USA); p19ARF (p19ARF exon 2, Rockland, Gilbertsville, PA, USA); Mdm2 (C-18, Santa Cruz Biotechnology); FoxO3 (75D8, Cell Signaling Technology); FoxO1 (C29H4, Cell Signaling Technology); and β-actin (clone AC-74, Sigma-Aldrich).
Statistical analysis
GraphPad Prism (Version 6.0d, GraphPad Software, San Diego, CA, USA) was used to graph and statistically analyse data. Student’s t-test (an unpaired two-tailed t-test) or the log-rank (Mantel–Cox) test, for Kaplan–Meier mouse survival curves, were used to determine statistical significance.
Abbreviations
- dnFoxO:
-
dominant-negative FoxO
- FACS:
-
fluorescence-activated cell sorting
- FoxO:
-
Forkhead-box O class
- HSC:
-
haemopoietic stem cell
- IHC:
-
immunohistochemistry
- IL-2:
-
interleukin 2
- IL-7:
-
interleukin 7
- PI:
-
propidium iodide
- sIg:
-
surface immunoglobulin
- WBC:
-
white blood cell
- WT:
-
wild type.
References
van der Horst A, Burgering BM . Stressing the role of FoxO proteins in lifespan and disease. Nat Rev Mol Cell Biol 2007; 8: 440–450.
Lam EW, Brosens JJ, Gomes AR, Koo CY . Forkhead box proteins: tuning forks for transcriptional harmony. Nat Rev Cancer 2013; 13: 482–495.
Dijkers PF, Medema RH, Lammers JJ, Koenderman L, Coffer PJ . Expression of the pro-apoptotic Bcl-2 family member Bim is regulated by the forkhead transcription factor FKHR-L1. Curr Biol 2000; 10: 1201–1204.
Dijkers PF, Birkenkamp KU, Lam EW, Thomas NS, Lammers JW, Koenderman L et al. FKHR-L1 can act as a critical effector of cell death induced by cytokine withdrawal: protein kinase B-enhanced cell survival through maintenance of mitochondrial integrity. J Cell Biol 2002; 156: 531–542.
Stahl M, Dijkers PF, Kops GJ, Lens SM, Coffer PJ, Burgering BM et al. The forkhead transcription factor FoxO regulates transcription of p27Kip1 and Bim in response to IL-2. J Immunol 2002; 168: 5024–5031.
Medema RH, Kops GJ, Bos JL, Burgering BM . AFX-like Forkhead transcription factors mediate cell-cycle regulation by Ras and PKB through p27kip1. Nature 2000; 404: 782–787.
Alvarez B, Martinez AC, Burgering BM, Carrera AC . Forkhead transcription factors contribute to execution of the mitotic programme in mammals. Nature 2001; 413: 744–747.
Ferber EC, Peck B, Delpuech O, Bell GP, East P, Schulze A . FOXO3a regulates reactive oxygen metabolism by inhibiting mitochondrial gene expression. Cell Death Differ 2012; 19: 968–979.
van der Vos KE, Eliasson P, Proikas-Cezanne T, Vervoort SJ, van Boxtel R, Putker M et al. Modulation of glutamine metabolism by the PI(3)K-PKB-FOXO network regulates autophagy. Nat Cell Biol 2012; 14: 829–837.
Eijkelenboom A, Burgering BM . FOXOs: signalling integrators for homeostasis maintenance. Nat Rev Mol Cell Biol 2013; 14: 83–97.
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo P, Hu LS et al. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 1999; 96: 857–868.
Matsuzaki H, Daitoku H, Hatta M, Tanaka K, Fukamizu A . Insulin-induced phosphorylation of FKHR (Foxo1) targets to proteasomal degradation. Proc Natl Acad Sci USA 2003; 100: 11285–11290.
Essers MA, Weijzen S, de Vries-Smits AM, Saarloos I, de Ruiter ND, Bos JL et al. FOXO transcription factor activation by oxidative stress mediated by the small GTPase Ral and JNK. EMBO J 2004; 23: 4802–4812.
Furuyama T, Nakazawa T, Nakano I, Mori N . Identification of the differential distribution patterns of mRNAs and consensus binding sequences for mouse DAF-16 homologues. Biochem J 2000; 349: 629–634.
Jacobs FM, van der Heide LP, Wijchers PJ, Burbach JP, Hoekman MF, Smidt MP . FoxO6, a novel member of the FoxO class of transcription factors with distinct shuttling dynamics. J Biol Chem 2003; 278: 35959–35967.
Dejean AS, Hedrick SM, Kerdiles YM . Highly specialized role of Forkhead box O transcription factors in the immune system. Antioxid Redox Signal 2011; 14: 663–674.
Hosaka T, Biggs WH 3rd, Tieu D, Boyer AD, Varki NM, Cavenee WK et al. Disruption of forkhead transcription factor (FOXO) family members in mice reveals their functional diversification. Proc Natl Acad Sci USA 2004; 101: 2975–2980.
Salih DA, Rashid AJ, Colas D, de la Torre-Ubieta L, Zhu RP, Morgan AA et al. FoxO6 regulates memory consolidation and synaptic function. Genes Dev 2012; 26: 2780–2801.
Nakae J, Biggs WH 3rd, Kitamura T, Cavenee WK, Wright CV, Arden KC et al. Regulation of insulin action and pancreatic beta-cell function by mutated alleles of the gene encoding forkhead transcription factor Foxo1. Nat Genet 2002; 32: 245–253.
Castrillon DH, Miao L, Kollipara R, Horner JW, DePinho RA . Suppression of ovarian follicle activation in mice by the transcription factor Foxo3a. Science 2003; 301: 215–218.
Miyamoto K, Araki KY, Naka K, Arai F, Takubo K, Yamazaki S et al. Foxo3a is essential for maintenance of the hematopoietic stem cell pool. Cell Stem Cell 2007; 1: 101–112.
Yalcin S, Zhang X, Luciano JP, Mungamuri SK, Marinkovic D, Vercherat C et al. Foxo3 is essential for the regulation of ataxia telangiectasia mutated and oxidative stress-mediated homeostasis of hematopoietic stem cells. J Biol Chem 2008; 283: 25692–25705.
Warr MR, Binnewies M, Flach J, Reynaud D, Garg T, Malhotra R et al. FOXO3A directs a protective autophagy program in haematopoietic stem cells. Nature 2013; 494: 323–327.
Altomare DA, Testa JR . Perturbations of the AKT signaling pathway in human cancer. Oncogene 2005; 24: 7455–7464.
Paik JH, Kollipara R, Chu G, Ji H, Xiao Y, Ding Z et al. FoxOs are lineage-restricted redundant tumor suppressors and regulate endothelial cell homeostasis. Cell 2007; 128: 309–323.
Bouchard C, Lee S, Paulus-Hock V, Loddenkemper C, Eilers M, Schmitt CA . FoxO transcription factors suppress Myc-driven lymphomagenesis via direct activation of Arf. Genes Dev 2007; 21: 2775–2787.
Bouchard C, Marquardt J, Bras A, Medema RH, Eilers M . Myc-induced proliferation and transformation require Akt-mediated phosphorylation of FoxO proteins. EMBO J 2004; 23: 2830–2840.
Amente S, Zhang J, Lavadera ML, Lania L, Avvedimento EV, Majello B . Myc and PI3K/AKT signaling cooperatively repress FOXO3a-dependent PUMA and GADD45a gene expression. Nucleic Acids Res 2011; 39: 9498–9507.
Eijkelenboom A, Mokry M, de Wit E, Smits LM, Polderman PE, van Triest MH et al. Genome-wide analysis of FOXO3 mediated transcription regulation through RNA polymerase II profiling. Mol Syst Biol 2013; 9: 638.
Delpuech O, Griffiths B, East P, Essafi A, Lam EW, Burgering B et al. Induction of Mxi1-SR alpha by FOXO3a contributes to repression of Myc-dependent gene expression. Mol Cell Biol 2007; 27: 4917–4930.
Smith DP, Bath ML, Harris AW, Cory S . T-cell lymphomas mask slower developing B-lymphoid and myeloid tumors in transgenic mice with broad hematopoietic expression of MYC. Oncogene 2005; 24: 3544–3553.
Smith DP, Bath ML, Metcalf D, Harris AW, Cory S . MYC levels govern hematopoietic tumor type and latency in transgenic mice. Blood 2006; 108: 653–661.
Adams JM, Harris AW, Pinkert CA, Corcoran LM, Alexander WS, Cory S et al. The c-myc oncogene driven by immunoglobulin enhancers induces lymphoid malignancy in transgenic mice. Nature 1985; 318: 533–538.
Harris AW, Pinkert CA, Crawford M, Langdon WY, Brinster RL, Adams JM . The Eμ-myc transgenic mouse: a model for high-incidence spontaneous lymphoma and leukemia of early B cells. J Exp Med 1988; 167: 353–371.
Michalak EM, Jansen ES, Happo L, Cragg MS, Tai L, Smyth GK et al. Puma and to a lesser extent Noxa are suppressors of Myc-induced lymphomagenesis. Cell Death Differ 2009; 16: 684–696.
Campbell KJ, Bath ML, Turner ML, Vandenberg CJ, Bouillet P, Metcalf D et al. Elevated Mcl-1 perturbs lymphopoiesis, promotes transformation of hematopoietic stem/progenitor cells, and enhances drug resistance. Blood 2010; 116: 3197–3207.
Strasser A, Harris AW, Bath ML, Cory S . Novel primitive lymphoid tumours induced in transgenic mice by cooperation between myc and bcl-2. Nature 1990; 348: 331–333.
Rosenbaum H, Harris AW, Bath ML, McNeall J, Webb E, Adams JM et al. An Em-v-abl transgene elicits plasmacytomas in concert with an activated myc gene. EMBO J 1990; 9: 897–905.
Herold MJ, Rohrbeck L, Lang MJ, Grumont R, Gerondakis S, Tai L et al. Foxo-mediated Bim transcription is dispensable for the apoptosis of hematopoietic cells that is mediated by this BH3-only protein. EMBO Rep 2013; 14: 992–998.
Dejean AS, Beisner DR, Ch'en IL, Kerdiles YM, Babour A, Arden KC et al. Transcription factor Foxo3 controls the magnitude of T cell immune responses by modulating the function of dendritic cells. Nat Immunol 2009; 10: 504–513.
Yalcin S, Marinkovic D, Mungamuri SK, Zhang X, Tong W, Sellers R et al. ROS-mediated amplification of AKT/mTOR signalling pathway leads to myeloproliferative syndrome in Foxo3(-/-) mice. EMBO J 2010; 29: 4118–4131.
Kang H, Corr M, Mansson R, Welinder E, Hedrick SM, Stone EL . Loss of murine FOXO3 in cells of the myeloid lineage enhances myelopoiesis but protects from K/BxN-serum transfer-induced arthritis. PLoS One 2015; 10: e0126728.
Hinman RM, Nichols WA, Diaz TM, Gallardo TD, Castrillon DH, Satterthwaite AB . Foxo3-/- mice demonstrate reduced numbers of pre-B and recirculating B cells but normal splenic B cell sub-population distribution. Int Immunol 2009; 21: 831–842.
Bakker WJ, van Dijk TB, Parren-van Amelsvoort M, Kolbus A, Yamamoto K, Steinlein P et al. Differential regulation of Foxo3a target genes in erythropoiesis. Mol Cell Biol 2007; 27: 3839–3854.
Marinkovic D, Zhang X, Yalcin S, Luciano JP, Brugnara C, Huber T et al. Foxo3 is required for the regulation of oxidative stress in erythropoiesis. J Clin Invest 2007; 117: 2133–2144.
Langdon WY, Harris AW, Cory S, Adams JM . The c-myc oncogene perturbs B lymphocyte development in Eμ-myc transgenic mice. Cell 1986; 47: 11–18.
Eilers M, Eisenman RN . Myc's broad reach. Genes Dev 2008; 22: 2755–2766.
Kops GJ, Medema RH, Glassford J, Essers MA, Dijkers PF, Coffer PJ et al. Control of cell cycle exit and entry by protein kinase B-regulated forkhead transcription factors. Mol Cell Biol 2002; 22: 2025–2036.
Evan GI, Wyllie AH, Gilbert CS, Littlewood TD, Land H, Brooks M et al. Induction of apoptosis in fibroblasts by c-myc protein. Cell 1992; 69: 119–128.
Evan GI, Vousden KH . Proliferation, cell cycle and apoptosis in cancer. Nature 2001; 411: 342–348.
Eischen CM, Weber JD, Roussel MF, Sherr CJ, Cleveland JL . Disruption of the ARF-Mdm2-p53 tumor suppressor pathway in Myc-induced lymphomagenesis. Genes Dev 1999; 13: 2658–2669.
Ekoff M, Kaufmann T, Engström M, Motoyama N, Villunger A, Jönsson JI et al. The BH3-only protein Puma plays an essential role in cytokine deprivation-induced apoptosis of mast cells. Blood 2007; 110: 3209–3217.
Sherr CJ . The INK4a/ARF network in tumour suppression. Nat Rev Mol Cell Biol 2001; 2: 731–737.
Brady CA, Jiang D, Mello SS, Johnson TM, Jarvis LA, Kozak MM et al. Distinct p53 transcriptional programs dictate acute DNA-damage responses and tumor suppression. Cell 2011; 145: 571–583.
Valente LJ, Gray DH, Michalak EM, Pinon-Hofbauer J, Egle A, Scott CL et al. p53 efficiently suppresses tumor development in the complete absence of its cell-cycle inhibitory and proapoptotic effectors p21, Puma, and Noxa. Cell Rep 2013; 3: 1339–1345.
Nicoletti I, Migliorati G, Pagliacci MC, Grignani F, Riccardi C . A rapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J Immunol Meth 1991; 139: 271–279.
Karasuyama H, Melchers F . Establishment of mouse cell lines which constitutively secrete large quantities of interleukin 2, 3, 4 or 5, using modified cDNA expression vectors. Eur J Immunol 1988; 18: 97–104.
Acknowledgements
We thank our institute colleagues A Strasser, MJ Herold and Steve Nutt for useful discussions; J Mansheim, K Hughes, S Allan, M Robati and J Corbin for excellent technical assistance; and the institute’s Flow Cytometry Facility for skilled support. This work was supported by the NHMRC (Australia) program grants 461221; 1016647; NCI grant CA43540; Leukemia and Lymphoma Society Specialized Center for Research Grant 7015-02; an NHMRC Career Development Award (CJV); and infrastructure support to WEHI from the NHMRC Independent Research Institute Infrastructure Support Scheme (IRISS) and the Victorian State Government Operational Infrastructure Support (OIS).
Author contributions
CJV and SC conceived the studies, planned experiments, analysed data and wrote the manuscript. CJV performed the experiments. NM provided mice and intellectual input.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Edited by G Melino
Supplementary Information accompanies this paper on Cell Death and Disease website
Supplementary information
Rights and permissions
Cell Death and Disease is an open-access journal published by Nature Publishing Group. This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Vandenberg, C., Motoyama, N. & Cory, S. FoxO3 suppresses Myc-driven lymphomagenesis. Cell Death Dis 7, e2046 (2016). https://doi.org/10.1038/cddis.2015.396
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cddis.2015.396
This article is cited by
-
The APC/C E3 ligase subunit ANAPC11 mediates FOXO3 protein degradation to promote cell proliferation and lymph node metastasis in urothelial bladder cancer
Cell Death & Disease (2023)
-
Mnt modulates Myc-driven lymphomagenesis
Cell Death & Differentiation (2017)
