
Abstract
Mass releases of sterilized male insects, in the frame of sterile insect technique programs, have helped suppress insect pest populations since the 1950s. In the major horticultural pests Bactrocera dorsalis, Ceratitis capitata, and Zeugodacus cucurbitae, a key phenotype white pupae (wp) has been used for decades to selectively remove females before releases, yet the gene responsible remained unknown. Here, we use classical and modern genetic approaches to identify and functionally characterize causal wp− mutations in these distantly related fruit fly species. We find that the wp phenotype is produced by parallel mutations in a single, conserved gene. CRISPR/Cas9-mediated knockout of the wp gene leads to the rapid generation of white pupae strains in C. capitata and B. tryoni. The conserved phenotype and independent nature of wp− mutations suggest this technique can provide a generic approach to produce sexing strains in other major medical and agricultural insect pests.
Similar content being viewed by others

Introduction
Tephritid species, including the Mediterranean fruit fly (medfly) Ceratitis capitata, the oriental fruit fly Bactrocera dorsalis, the melon fly Zeugodacus cucurbitae, and the Queensland fruit fly Bactrocera tryoni, are major agricultural pests worldwide1. The sterile insect technique (SIT) is a species-specific and environment-friendly approach to control their populations, which has been successfully applied as a component of area-wide integrated pest management programs2,3,4. The efficacy and cost-effectiveness of these large-scale operational SIT applications has been significantly enhanced by the development and use of genetic sexing strains (GSS) for medfly, B. dorsalis and Z. cucurbitae5,6.
A GSS requires two principal components: a selectable marker, which could be phenotypic or conditionally lethal, and the linkage of the wild-type allele of this marker to the male sex, ideally as close as possible to the male determining region. In a GSS, males are heterozygous and phenotypically wild type, whilst females are homozygous for the mutant allele thus facilitating sex separation6,7,8. Puparium color was one of the first phenotypic traits exploited as a selectable marker for the construction of GSS. In all three species, brown is the typical puparium color. However, naturally occurring color mutants such as white pupae (wp)9 and dark pupae (dp)10 have occurred in the field or laboratory stocks. The wp locus was successfully used as a selectable marker to develop GSS for C. capitata, B. dorsalis, and Z. cucurbitae6,11,12; however, its genetic basis has never been resolved.
Biochemical studies provided evidence that the white pupae phenotype in medfly is due to a defect in the mechanism responsible for the transfer of catecholamines from the hemolymph to the pupal cuticle13. In addition, classical genetic studies showed that the wp phenotype is due to a recessive mutation in an autosomal gene located on chromosome 5 of the medfly genome9,14. The development of translocation lines combined with deletion and transposition mapping and advanced cytogenetic studies allowed the localization of the gene responsible for the wp phenotype on the right arm of chromosome 5, at position 59B of the trichogen polytene chromosome map15. In the same series of experiments, the wp locus was shown to be tightly linked to a temperature-sensitive lethal (tsl) gene (position 59B–61C), which is the second selectable marker of the VIENNA 7 and VIENNA 8 GSS currently used in all medfly SIT operational programs worldwide7,15.
The genetic stability of a GSS is a major challenge, mainly due to recombination phenomena taking place between the selectable marker and the translocation breakpoint. To address this risk, a chromosomal inversion called D53 was induced and integrated into the medfly VIENNA 8 GSS (VIENNA 8D53+)6,8. Cytogenetic analysis indicated that the D53 inversion spans a large region of chromosome 5 (50B–59C on trichogen polytene chromosome map) with the wp locus being inside the inversion, close to its right breakpoint6.
Extensive genetic and cytogenetic studies facilitated the development of a physical map of the medfly genome8,16. The annotated gene set provided opportunities for the identification of genes or loci-associated mutant phenotypes, such as the wp and tsl, used for the construction of GSS16,17. Salivary gland polytene chromosome maps developed for C. capitata, B. dorsalis, Z. cucurbitae, and B. tryoni show that their homologous chromosomes exhibit similar banding patterns. In addition, in situ hybridization analysis of several genes confirmed that there is extensive shared synteny, including the right arm of chromosome 5 where the C. capitata wp gene is localized8. Interestingly, two recent studies identified SNPs associated with the wp phenotype in C. capitata and Z. cucurbitae that were also on chromosome 518,19.
In this work, we employ different strategies involving genetics, cytogenetics, genomics, transcriptomics, gene editing, and bioinformatics to identify independent natural mutations in a gene responsible for puparium coloration in three tephritid species of major agricultural importance, C. capitata, B. dorsalis, and Z. cucurbitae. We then functionally characterize causal mutations within this gene in C. capitata and B. tryoni resulting in development of new white pupae strains. Due to its conserved nature20 and widespread occurrence in many insect species of agricultural and medical importance, we also discuss the potential use of this gene as a generic selectable marker for the construction of GSS for SIT applications.
Results
Resolving the B. dorsalis wp locus by introgression experiments
The B. dorsalis white pupae phenotype was introgressed into B. tryoni to generate a strain referred to as the Bactrocera introgressed line (BIL, Supplementary Fig. 1). To determine the proportion of B. dorsalis genome introgressed into BIL, whole-genome sequence data from male and female B. dorsalis, B. tryoni, and BIL individuals were analyzed. Paired-end Illumina short read data from single B. oleae males (SRR826808) and females (SRR826807) were used as an outgroup. Single copy orthologs across the genome (n = 1,846) were used to reconstruct the species topology revealing a species-specific monophyly (Fig. 1a) consistent with published phylogenies21,22. Reconstruction also showed monophyly between B. tryoni and BIL across 99.2% of gene trees suggesting the majority of loci originally introgressed from B. dorsalis have been removed during backcrosses.

a Species tree constructed from 1846 single copy ortholog gene trees for four haplotypes of B. oleae, B. dorsalis, B. tryoni, and BIL. Branches corresponding to BIL individuals are shown in blue. All nodes were well supported with posterior probabilities >0.97. b Nei’s absolute genetic distance (dXY) calculated for tiled 100 kb windows across the genome between B. tryoni vs BIL (Bt vs BIL); B. tryoni vs B. dorsalis (Bt vs Bd); B. tryoni vs B. oleae (Bt vs Bo); and BIL vs B. oleae (BIL vs Bo). Box and whisker graphs (including outliers) represent a summary of 2294 genomic windows. Boxes show the first and third inter quartile range (IQR) while whiskers extend to a maximum of 1.5 ∗ IQR. All values outside 1.5 ∗ IQR are shown as plus signs. c The introgression estimator (ƒd) calculated across tiled 100 kb windows to identify regions of disproportionately shared alleles between BIL and B. dorsalis, ƒd (Bt, BIL, Bd; Bo). d The three evolutionary hypothesis/topologies of interest to identify introgressed regions and their representation across the genome: species (purple, 98.82%), introgression (orange, 1.04%) and a negative control tree (green, 0.14%). e Nei’s absolute genetic distance (dXY) calculated for tiled 10 kb windows across the candidate wp locus for B. tryoni vs BIL (purple), B. dorsalis vs BIL (orange), B. oleae vs BIL (green). f Topology weighting for each topology shown in d, calculated for 1 kb tiled local trees across the candidate wp locus. g The introgression estimator (ƒd) calculated across tiled 10 kb windows for the comparison ƒd (Bt, BIL, Bd; Bo) to identify the start and end of the introgressed locus. Source data are provided in a Source Data file.
Genomes were partitioned into 100 kb windows and pairwise absolute genetic distance (dXY) calculated between each species and BIL to estimate admixture. Bactrocera dorsalis was found to be highly similar to a small proportion of the BIL genome (Fig. 1b; purple), as indicated by dXY values approaching the median value of B. dorsalis vs B. tryoni (Fig. 1b; yellow).
Two formal tests for introgression were also carried out, the ƒ estimator ƒd (Fig. 1c) and topology weighting (Fig. 1d). Three distinct local evolutionary histories (Fig. 1d) were tested using dXY and topology weighting across the B. dorsalis wp Quantitative Trait Locus (QTL) i) BIL is closest to B. tryoni (Fig. 1d; purple, expected across most of the genome), ii) BIL is closest to B. dorsalis (Fig. 1d; orange, expected at the wp- locus), and iii) BIL is closest to B. oleae (Fig. 1d; green, a negative control). Across the nuclear genome the species topology was supported in 98.82% of windows. Both ƒd and topology weighting confirmed a lack of widespread introgression from B. dorsalis into BIL with few (n = 42) discordant outlier windows. Genomic windows discordant across all three tests were considered candidate regions for the wp mutation. Four scaffolds accounting for 1.18% of the B. dorsalis genome met these criteria and only two, NW_011876372.1 and NW_011876398.1, showed homozygous introgression consistent with a recessive white pupae phenotype (Supplementary Fig. 2).
To resolve breakpoints within the B. dorsalis wp- QTL, a windowed analysis across NW_011876398.1 and NW_011876372.1 was performed using dXY (Fig. 1e), topology weighting (Fig. 1f) and ƒd (Fig. 1g). The maximum range of the introgressed locus was 4.49 Mb (NW_011876398.1 was 2.9–5.94 Mb and NW_011876372.1 was 0–1.55 Mb) (Fig. 1e–g). The wp- QTL was further reduced to a 2.71 Mb region containing 113 annotated protein coding genes through analyzing nucleotide diversity (π) among eight pooled BIL genomes (3.8 Mb on NW_011876398.1 to 0.73 Mb on scaffold NW_011876372.1, Supplementary Fig. 2).
Resolving the C. capitata wp by genome sequencing and in situ hybridization
Cytogenetic studies have determined the gene responsible for the white pupae phenotype to be localized on the right arm of chromosome 5, at position 59B of the trichogen polytene chromosome map15. The equivalent of position 59B is position 76B of the salivary gland polytene chromosome map, inside but close to the right breakpoint of the D53 inversion (69C–76B on the salivary gland polytene chromosome map). Long read sequencing data were generated of the wild-type strain Egypt II (EgII, WT), the inversion line D53 and the genetic sexing strain VIENNA 8 (without the inversion; VIENNA 8D53−|−) (Supplementary Table 1) to enable a comparison of the genomes and locate the breakpoints of the D53 inversion, to subsequently narrow down the target region, and to identify wp candidate genes.
Chromosome 5-specific markers16 were used to identify the EgII_Ccap3.2.1 scaffold_5 as complete chromosome 5. Candidate D53 breakpoints in EgII scaffold_5 were identified using the alignment of three genome datasets EgII, VIENNA 8D53−|−, and D53 (see material and methods). The position of the D53 inversion breakpoints was located between 25,455,334 and 25,455,433 within a scaffold gap (left breakpoint), and at 61,880,224 bp in a scaffolded contig (right breakpoint) on EgII chromosome 5 (Ccap3.2.1; accession GCA_905071925) (Fig. 2a). The region containing the causal wp gene was known to be just next to the right breakpoint of the D53 inversion. Cytogenetic analysis and in situ hybridization using the WT EgII strain and the D53 inversion line confirmed the overall structure of the inversion, covering the area of 69C–76B on the salivary gland polytene chromosomes (Fig. 2), as well as the relative position of markers residing inside and outside the breakpoints (Fig. 2 and Supplementary Fig. 3). PCRs using two primer pairs flanking the predicted breakpoints (Supplementary Fig. 4) and subsequent sequencing confirmed the exact sequence of the breakpoints. Thereby, the wild-type status was confirmed for EgII flies and VIENNA 7D53+|− GSS males, which are heterozygous for the inversion. Correspondingly, these amplicons were not present in D53 males and females or in VIENNA 7D53+|+ GSS females (all homozygous for the inversion) (Supplementary Fig. 4). Positive signals for the inversion were detected in D53 and VIENNA 7D53+ GSS males and females, but not in WT flies using an inversion-specific primer pair (Supplementary Fig. 4).

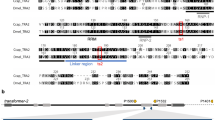
a Chromosome scale assembly of C. capitata EgII chromosome 5. Shown are the positions of in situ mapped genes white (w), 6-phosphogluconate dehydrogenase (Pgd), glucose-6-phosphate 1-dehydrogenase (Zw), and sex lethal (Sxl), the position of the D53 inversion breakpoints (blue; LB = left breakpoint, RB = right breakpoint), and the relative position of white pupae (wp) on the polytene chromosome map of chromosome 571 (left (L) and right (R) chromosome arm, linked at the centromeric region (C)) and the PacBio-Hi-C EgII scaffold_5 (bp = base pairs), representing the complete chromosome 5 (Ccap3.2.1, accession GCA_905071925). The position of the yellow gene (y, LOC101455502) was confirmed on chromosome 5 70A by in situ hybridization, despite its sequence not been found in the scaffold assembly. b Schematic illustration of chromosome 5 without (EgII, WT) and with (D53) D53 inversion, with additional marker genes Curly (Cy), integrin-aPS2 (PS2a), white (w), chorion S36/38 (Ccs36/38), vitellogenin-1/2-like (Vg1 + 2). The inverted part of chromosome 5 is shown in light blue, the centromere in yellow. Two probes, one inside (y, 70A) and one outside (Pgd, 68B) of the left inversion breakpoint were used to verify the D53 inversion breakpoints by in situ hybridization. WT EgII is shown in c and e, D53 in d and f. Chromosomal segments are numbered, arrows in micrographs indicate in situ hybridization signal. In situ hybridizations were done at least in duplicates and at least ten nuclei were analyzed per sample, scale bar = 10 µm. All replicates led to similar results. The source data underlying Fig. 2c–f are provided as a Source Data file.
Genome and transcriptome sequencing reveal a single candidate wp gene
Orthologs within the QTL of B. dorsalis, C. capitata, and scaffolds known to segregate with the wp phenotype in Z. cucurbitae (NW_011863770.1 and NW_011863674.1)18 were investigated for null mutations under the assumption that errors within a conserved gene result in white pupae. A single ortholog containing fixed indels absent from wild-type strains was identified in each species. White pupae B. dorsalis and BIL strains showed a 37 bp frame-shift deletion in the first coding exon of LOC105232189 introducing a premature stop codon 210 bp from the transcription start site (Fig. 3a). Presence of the deletion was confirmed in silico using whole genome resequencing from the wp and wildtype mapped to the reference, and by de novo assembly of Illumina RNAseq data transcripts (Fig. 3a).

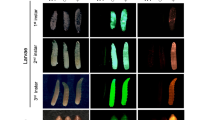
The gray graphs show expression profiles from the candidate wp loci in WT (wp+) and mutant (wp−) flies at the immobile pupae stages of a B. dorsalis, b C. capitata, and c Z. cucurbitae. The gene structure (not drawn to scale) is indicated below as exons (arrows labeled E1–E4) and introns (dashed lines), the Major Facilitator Superfamily (MFS) domain is shown in blue. The positions of independent wp mutations (Bd: 37 bp deletion, Cc: approximate 8150 bp insertion, Zc: 13 bp deletion) are marked with black dashed boxes in the expression profiles and are shown in detail below the gene models based on de novo assembly of RNAseq data from WT and white pupae phenotype individuals (nucleotide and amino acid sequences). Deletions are shown as dashes, alterations on protein level leading to premature stop codons are depicted as asterisks highlighted in black. In situ hybridization on polytene chromosomes for d B. dorsalis, e C. capitata, and f Z. cucurbitae confirmed the presence of the wp locus on the right arm of chromosome 5 in all three species (arrows in micrographs). In situ hybridizations were done at least in duplicates and at least ten nuclei were analyzed per sample, scale bar = 10 µm. The source data underlying Fig. 3d–f are provided as a Source Data file.
In C. capitata, a D53 Nanopore read alignment on EgII showed an independent approximate 8150 bp insertion into the third exon of LOC101451947 disrupting proper gene transcription 822 bp from the transcription start site (Fig. 3b). The insertion sequence is flanked by identical repeats, suggesting that it may originate from a transposable element insertion. The C. capitata mutation was confirmed in silico, as in B. dorsalis, using whole genome sequencing and RNAseq data (Fig. 3b).
Transcriptome data from the white pupae-based genetic sexing strain of Z. cucurbitae revealed a 13 bp deletion in the third exon of LOC105216239 on scaffold NW_011863770.1 introducing a premature stop codon (Fig. 3c).
The candidate white pupae gene in all three species had a reciprocal best BLAST hit to the putative metabolite transport protein CG14439 in Drosophila melanogaster and contains a Major Facilitator-like superfamily domain (MFS_1, pfam07690), suggesting a general function as a metabolite transport protein. In situ hybridization on polytene chromosomes of B. dorsalis, C. capitata and Z. cucurbitae was used to confirm the presence of the wp locus in the same syntenic position on the right arm of chromosome 5 (Fig. 3d–f). Therefore, all three species show a mutation in the same positional orthologous gene likely to be responsible for the phenotype in all three genera.
Knockout of the MFS gene causes white pupae phenotypes
An analogous B. dorsalis wp- mutation was developed in B. tryoni by functional knockouts of the putative Bt_wp using the CRISPR/Cas9 system. A total of 591 embryos from the Ourimbah laboratory strain were injected using two guides with recognition sites in the first coding exon of this gene (Fig. 4a). Injected embryos surviving to adulthood (n = 19, 3.2%) developed with either wild-type brown (n = 12) or somatically mosaic white-brown puparia (n = 7, Supplementary Fig. 5). Surviving G0 adults were individually backcrossed into the Ourimbah strain, resulting in potentially wp+|−(CRISPR) heterozygous brown pupae (Fig. 4c). Five independent G0 crosses were fertile (three mosaic white-brown and two brown pupae phenotypes). G1 offspring were sibling mated and visual inspection of G2 progeny revealed that three families contained white pupae individuals. Four distinct frameshift mutations were observed in screened G2 progeny (Fig. 4a) suggesting functional KO of putative Bt_wp is sufficient to produce the white pupae phenotype in B. tryoni. Capillary sequencing of cloned Bt_MFS amplicons revealed deletions ranging from a total of 4–155 bp, summed across the two guide recognition sites, introducing premature stop codons.

A schematic structure of the wp CDS exons (E1, E2, E3, E4) including the MFS domain in B. tryoni (a) and C. capitata (b) are shown. Positions of gRNAs targeting the first and third exon in B. tryoni and C. capitata, respectively, are indicated by green arrows. Nucleotide and amino acid sequences of mutant wp alleles identified in G1 individuals are compared to the WT reference sequence in B. tryoni (a) and C. capitata (b). Deletions are shown as dashes, alterations on protein level leading to premature stop codons are depicted as asterisks highlighted in black. Numbers on the right side represent InDel sizes (bp = base pairs). Crossing schemes to generate homozygous wp−(CRISPR) lines in B. tryoni (c) and C. capitata (d) show different strategies to generate wp strains. Bright-field images of empty puparia are depicted for both species. Genotype schematics and corresponding PCR analysis (for C. capitata) validating the presence of CRISPR-induced (orange) and natural (blue, for C. capitata) wp mutations are shown next to the images of the puparia. c Injected G0 B. tryoni were backcrossed to the Ourimbah laboratory strain resulting in uniformly brown G1 offspring (depicted as illustration because no images were acquired during G1). G1 inbreeding led to G2 individuals homozygous for the white pupae phenotype. d Injected WT G0 flies were crossed to flies homozygous for the naturally occurring wp− allele (wp−(nat)). wp−(nat) (457 bp amplicon) and wp−(CRISPR) or WT (724 bp amplicon) alleles were identified by multiplex PCR (left lane; L = NEB 2 log ladder). White pupae phenotypes in G1 indicated positive CRISPR events. G2 flies with a white pupae phenotype that were homozygous for the wp−(CRISPR) allele were used to establish lines. PCR was done once for each individual, wp−(CRISPR) alleles were verified and further analyzed via sequencing. The source data underlying Fig. 4d are provided as a Source Data file.
In C. capitata, CRISPR/Cas9 gene editing was used to knockout the orthologous gene and putative Cc_wp, LOC101451947, to confirm that it causes a white puparium phenotype. A mix of recombinant Cas9 protein and the gRNA_MFS, targeting the third exon and thereby the MFS domain of the presumed Cc_wp CDS (Fig. 4b), was injected into 588 EgII WT embryos of which 96 developed to larvae and 67 pupated. All injected G0 pupae showed brown pupal color. In total, 29 G0 males and 34 females survived to adulthood (9.3%) and were backcrossed individually or in groups (see material and methods) to a strain carrying the naturally occurring white pupae mutation (wp−(nat); strain #1402_22m1B)23 (Fig. 4d). As white pupae is known to be monogenic and recessive in C. capitata, this complementation assay was used to reveal whether the targeted gene is responsible for the naturally occurring white pupae phenotype or if the mutation is located in a different gene. G1 offspring would only show white pupae phenotypes if Cc_wp was indeed the white pupae gene, knocked-out by the CRISPR approach, and complemented by the natural mutation through the backcross (wp−(nat)|−(CRISPR)). In the case that the Cc_wp is not the gene carrying the natural wp− mutation, a brown phenotype would be observed for all offspring. Here, five out of 13 crosses, namely M1, M3, F2, F3, and F4, produced white pupae phenotype offspring. The crosses generated 221, 159, 70, 40, and 52 G1 pupae, of which 10, 30, 16, 1, and 1 pupa respectively, were white. Fifty-seven flies emerged from white puparia were analyzed via non-lethal genotyping, and all of them showed mutation events within the target region. Overall, eight different mutation events were seen, including deletions ranging from 1 to 9 bp and a 46 bp deletion combined with a 2 bp insertion (Fig. 4b). Five mutation events (B, D, E, G, H) caused frameshifts and premature stop codons. The remaining three (A, C, F), however, produced deletions of only one to three amino acids. Mutants were either inbred (mutation C) (Fig. 4d) or outcrossed to WT EgII (mutation A–H), both in groups according to their genotype. This demonstrated that Cc_wp is the gene carrying the wp−(nat), and that even the loss of a single amino acid without a frameshift at this position can cause the white pupae phenotype. Offspring from outcrosses of mutation A, D, and H, as well as offspring of the inbreeding (mutation C), were genotyped via PCR, and wp+|−(CRISPR) and wp−(CRISPR)|−(CRISPR) positive flies were inbred to establish homozygous wp-(CRISPR) lines.
Discussion
White pupae (wp) was first identified in C. capitata as a spontaneous mutation and was subsequently adopted as a phenotypic marker of fundamental importance for the construction of GSS for SIT6,9. Full penetrance expressivity and recessive inheritance rendered wp the marker of choice for GSS construction in two additional tephritid species, B. dorsalis and Z. cucurbitae11,12, allowing automated sex sorting based on pupal color. This was only possible because spontaneous wp mutations occur at relatively high rates either in the field or in mass rearing facilities and can easily be detected6,9. Despite the easy detection and establishment of wp mutants in these three species, similar mutations have not been detected in other closely or distantly related species such as B. tryoni, B. oleae, or Anastrepha ludens, despite large screens being conducted. In addition to being a visible GSS marker used to separate males and females, the wp phenotype is also important for detecting and removing recombinants in cases where sex separation is based on a conditional lethal gene such as the tsl gene in the medfly VIENNA 7 or VIENNA 8 GSS6,7. However, it took more than 20 years from the discovery and establishment of the wp mutants to the large-scale operational use of the medfly VIENNA 8 GSS for SIT applications6,9 and the genetic nature of the wp mutation remained unknown. The discovery of the underlying wp mutations and the availability of CRISPR/Cas genome editing would allow the fast recreation of such phenotypes and sexing strains in other insect pests. Isolation of the wp gene would also facilitate future efforts towards the identification of the closely linked tsl gene.
Using an integrated approach consisting of genetics, cytogenetics, genomics, transcriptomics, and bioinformatics, we identified the white pupae genetic locus in three major tephritid agricultural pest species, B. dorsalis, C. capitata, and Z. cucurbitae. Our study clearly shows the power of employing different strategies for gene discovery, one of which was species hybridization. In Drosophila, hybridization of different species has played a catalytic role in the deep understanding of species boundaries and the speciation processes, including the evolution of mating behavior and gene regulation24,25,26,27,28. In our study, we took advantage of two closely related species, B. dorsalis and B. tryoni, which can produce fertile hybrids and be backcrossed for consecutive generations. This allowed the introgression of the wp mutant locus of B. dorsalis into B. tryoni, resulting in the identification of the introgressed region, including the causal wp mutation via whole-genome resequencing and advanced bioinformatic analysis.
In C. capitata, we exploited two essential pieces of evidence originating from previous genetic and cytogenetic studies: the localization of wp to region 59B and 76B on chromosome 5 in the trichogen cells and salivary gland polytene chromosome map, respectively15,29, and its position close to the right breakpoint of the large inversion D536. This data prompted us to undertake a comparative genomic approach to identify the exact position of the right breakpoint of the D53 inversion, which would bring us in the vicinity of the wp gene. Coupled with comparative transcriptomic analysis, this strategy ensured that the analysis indeed tracked the specific wp locus on the right arm of chromosome 5, instead of any mutation in another, random locus which may participate in the pigmentation pathway and therefore result in the same phenotype. Functional characterization via CRISPR/Cas9-mediated knockout resulted in the establishment of new white pupae strains in C. capitata and B. tryoni and confirmed that this gene is responsible for the puparium’s coloration in these tephritid species. Interestingly, the wp phenotype is based on three independent and very different natural mutations of this gene, a rather large and transposon-like insertion in C. capitata, but only small deletions in the two other tephritids, B. dorsalis and Z. cucurbitae. In medfly, however, CRISPR-induced in-frame deletions of one or three amino acids in the MFS domain were sufficient to induce the wp phenotype, underlining the importance of this domain for correct coloration of the puparium.
It is worth noting that in the first stages of this study, we employed two additional approaches, which did not allow us to successfully narrow down the wp genomic region to the desired level. The first was based on Illumina sequencing of libraries produced from laser micro-dissected (Y;5) mitotic chromosomes that carry the wild-type allele of the wp gene through a translocation from the fifth chromosome to the Y. This dataset from the medfly VIENNA 7 GSS was comparatively analyzed to wild-type (Egypt II) Y and X chromosomes, and the complete genomes of Egypt II, VIENNA 7D53- GSS, and a D53 inversion line in an attempt to identify the chromosomal breakpoints of the translocation and/or inversion, which are close to the wp locus (Supplementary Table 2). However, this effort was not successful due to the short Illumina reads and the lack of a high-quality reference genome. The second approach was based on individual scale whole-genome resequencing/genotyping, and identifying fixed loci associated with pupal color phenotypes, which complemented the QTL analysis19. Seven loci associated with SNPs and larger deletions linked to the white pupae phenotype were analyzed based on their respective mutations and literature searches for their potential involvement in pigmentation pathways. However, we could not identify a clear link to the pupal coloration as shown by in silico, molecular, and in situ hybridization analysis (Supplementary Figs. 6 and 7, Supplementary Table 3).
The wp gene is a member of a Major Facilitator Superfamily (MFS). Orthologs of white pupae are present in 146 of 148 insect species aggregated in OrthoDB20 v9 and are single copy in 133 species. Furthermore, wp is included in the benchmarking universal single copy ortholog (BUSCO) gene set for Insecta and according to OrthoDB30 v10 has a below average evolutionary rate (0.87, OrthoDB group 42284at50557) suggesting an important and evolutionarily conserved function (Supplementary Fig. 8). Its ortholog in Bombyx mori, mucK, was shown to participate in the pigmentation at the larval stage31 whereas in D. melanogaster peak expression is during the prepupal stage after the larva has committed to pupation32, which is the stage where pupal cuticle sclerotization and melanization occurs. It is known that the insect cuticle consists of chitin, proteins, lipids, and catecholamines, which act as cross-linking agents thus contributing to polymerization and the formation of the integument33. Interestingly, the sclerotization and melanization pathways are connected and this explains the different mechanical properties observed in different medfly pupal color strains with the dark color cuticles being harder than the brown ones and the latter harder than the white color ones34. The fact that the white pupae mutants are unable to transfer catecholamines from the hemolymph to the cuticle is perhaps an explanation for the lack of the brown pigmentation13.
The discovery of the long-sought wp gene in this study and the recent discovery of the Maleness-on-the-Y (MoY) gene, which determines the male sex in several tephritids35, opens the way for the development of a generic approach for the construction of GSS for other species. Using CRISPR/Cas-based genome editing approaches, we can: (a) induce mutations in the wp orthologues of SIT target species and establish lines with wp phenotype and (b) link the rescue alleles as closely as possible to the MoY region. Given that the wp gene is present in diverse insect species including agricultural insect pests and mosquito disease vectors, this approach would allow more rapid development of GSS in SIT target species, members of diverse families, such as the agricultural pest species A. ludens, A. fraterculus, B. dorsalis, B. correcta, B. oleae, Drosophila suzukii, Cydia pomonella, Pectinophora gossypiella, Lobesia botrana; the livestock pests Glossina morsitans, G. pallidipes, G. palpalis gambiensis, G. austeni; and the mosquito disease vectors Aedes aegypti, Aedes albopictus, and Anopheles arabiensis. However, the biological quality of any new strain which is considered for SIT application should be first thoroughly tested in respect to their fitness and male mating competitiveness. In principle, these GSS will have higher fertility compared to the semi-sterile translocation lines6. In addition, these new generation GSS will be more stable since the rescue allele will be tightly linked to the male determining region thus eliminating recombination which can jeopardize the genetic integrity of any GSS. The concept of the generic approach can also be applied in species which lack a typical Y chromosome such as Ae. aegypti and Ae. albopictus. In these species, the rescue allele should be transferred close to the male determining gene (Nix) and the M locus36,37. It is hence important for this generic approach to identify regions close enough to the male determining loci to ensure the genetic stability of the GSS and to allow the proper expression of the rescue alleles. In the present study, we have already shown that CRISPR/Cas9-induced mutations resulting in the white pupae phenotype can be developed in SIT target species and the resulting strains provide already new opportunities for GSS based on visible markers.
Methods
Insect rearing
Ceratitis capitata, B. dorsalis, and Z. cucurbitae fly strains were maintained at 25 ± 1 °C, 48% RH and 14/10 h light/dark cycle, and fed with a mixture of sugar and yeast extract (3 v:1 v) and water. Larvae were reared on a gel diet, containing carrot powder (120 g/L), agar (3 g/L), yeast extract (42 g/L), benzoic acid (4 g/L), HCl (25%, 5.75 mL/L), and ethyl-4-hydroxybenzoate (2.86 g/L). Flies were anesthetized with N2 or CO2 for screening, sexing, and the setup of crosses. To slow down the development during the non-lethal genotyping process (C. capitata), adult flies were kept at 19 °C, 60% RH, and 24 h light for this period (1–4 days).
Bactrocera tryoni flies were obtained from New South Wales Department of Primary Industries (NSW DPI), Ourimbah, Australia and reared at 25 ± 2 °C, 65 ± 10% RH and 14/10 h light/dark cycle. Flies were fed with sugar, Brewer’s yeast and water and larvae were reared on a gel diet, containing Brewer’s yeast (204 g/L), sugar (121 g/L), methyl p-hydroxy benzoate (2 g/L), citric acid (23 g/L), wheat germ oil (2 g/L), sodium benzoate (2 g/L), and agar (10 g/L).
Introgression and identification of wp in B. dorsalis
Interspecific crosses between male B. tryoni (wp+|+) and female B. dorsalis (wp−|−) were carried out. The F1 wp+|− hybrids developed with brown puparia and were mass crossed. F2 wp−|− females were backcrossed into B. tryoni wp+|+ males. Backcrossing was then repeated five additional times to produce the white pupae Bactrocera introgressed line (BIL, Supplementary Fig. 1).
Genome sequencing using Illumina NovaSeq (2 ×150 bp, Deakin University) was performed on a single male and female from the B. dorsalis wp strain, B. tryoni, and the BIL (~ 26X) and two pools of five BIL individuals (~ 32X). Quality control of each sequenced library was carried out using FastQC v0.11.6 (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/) and aggregated using ngsReports38 v1.3. Adapter trimming was carried out using Trimmomatic39 v0.38 and paired reads were mapped to the B. dorsalis reference genome (GCF_000789215.1) using NextGenMap40 v0.5.5 under default settings. Mapped data were sorted and indexed using SAMtools, and deduplication was carried out using Picard MarkDuplicates v2.2.4 (https://github.com/broadinstitute/picard). Genotypes were called on single and pooled libraries separately with ploidy set to two and ten, respectively, using Freebayes41 v1.0.2. Each strain was set as a different population in Freebayes. Genotypes with less than five genotype depth were set to missing and sites with greater than 20% missing genotypes or indels filtered out using BCFtools42 v1.9. Conversion to the genomic data structure (GDS) format was carried out using SeqArray43 v1.26.2 and imported into the R package geaR44 v0.1 for population genetic analysis.
Single copy orthologs were identified in the B. dorsalis reference annotated proteins (NCBI Bactrocera dorsalis Annotation Release 100) with BUSCO45,46 v3 using the dipteran gene set45. Nucleotide alignments of each complete single copy ortholog were extracted from the called genotype set using geaR v0.1 and gene trees built using RAxML47 v8.2.10 with a GTR + G model. Gene trees were then imported into Astral III48 v5.1.1 for species tree estimation. Genome scans of absolute genetic divergence (dXY), nucleotide diversity (π), and the ƒ estimator ƒd were carried out using geaR v0.1. Two levels of analysis were carried out: i) genome wide scans of non-overlapping 100 kb windows and ii) locus scans of 10 kb tiled windows. Local phylogenies were built for nucleotide alignments of non-overlapping 1 kb windows using RAxML v8.2.10 with a GTR + G model and topology weighting was calculated using TWISST49.
Introgressed regions (i.e., candidate wp loci) were identified by extracting windows in the genome wide scan with topology weighting and ƒd greater than 0.75 and visually inspecting the ‘locus scan’ data set for dXY, ƒd, and topology weighting patterns indicative of introgression. Nucleotide alignments of all genes within candidate B. dorsalis introgressed regions were extracted from the GDS using geaR and visually inspected for fixed mutations in B. dorsalis wp, BIL individuals, and the two BIL pools. Candidate genes were then searched by tBLASTn against the D. melanogaster annotated protein set to identify putative functions and functional domains were annotated using HMMer50. Mapped read depth was calculated around candidate regions using SAMtools51 depth v1.9 and each sample’s read depth was normalized to the sample maximum to inspect putative deletions. Called genotypes were confirmed by de novo genome assembly of the B. dorsalis wp genome using MaSuRCA52 v3.3 under default settings. The de novo scaffold containing LOC105232189 was identified using the BLASTn algorithm. In silico exon–intron boundaries were then manually annotated in Geneious53 v11.
Identification of the D53 inversion and wp in C. capitata
Multiple C. capitata strains were used for this study. Egypt II (EgII) is a wild-type laboratory strain. D53 is a homozygous strain with an irradiation-induced inversion covering the area 69C–76B on the salivary gland polytene chromosome map (50B–59C on the trichogen cells polytene chromosome map). VIENNA 7 and VIENNA 8 are two GSS in which two (Y;5) translocations, in the region 58B and 52B of the trichogen cells polytene chromosome map, respectively, have resulted in the linkage of the wild-type allele of the wp and tsl genes to the male determining region of the Y chromosome. Thus, VIENNA 7 and VIENNA 8 males are heterozygous in the wp and tsl loci but phenotypically wild type while VIENNA 7 and VIENNA 8 females are homozygous for the mutant alleles and phenotypically white pupae, and they die when exposed to elevated temperatures. The VIENNA 7 and VIENNA 8 GSS can be constructed with and without the D53 inversion (VIENNA 7/8D53+ or D53−). When the GSS have the inversion, females are homozygous (D53+|+) for D53 while males are heterozygous (D53+|−)6,8,16.
To perform whole genome sequencing of C. capitata strains, high-molecular-weight (HMW) DNA was extracted from C. capitata lines (males and females of the WT EgII strain, the VIENNA 7D53−|− and VIENNA 8D53−|− GSS and the inversion line D53) and sequenced. Freshly emerged, virgin and unfed males and females were collected from all strains. For 10X Genomics linked read and Nanopore sequencing, the HMW was prepared as follows: twenty individuals of each sex and strain were pooled, ground in liquid nitrogen, and HMW DNA was extracted using the QIAGEN Genomic tip 100/G kit (Qiagen, Germany). For PacBio Sequel an EgII line was created with single pair crossing and subsequent sibling-mating for six generations. In all generations adult and larval diet contained 100 μg/mL tetracycline. HMW DNA from G6 individuals was prepared as follows: five males from this EgII line were pooled and ground in liquid nitrogen, and HMW DNA was extracted using the phenol/chloroform Phase Lock Gel tubes (QuantaBio)54. DNA for Illumina applications was extracted from individual flies (Supplementary Table 1). PacBio de novo sequencing: samples were purified with AMPure beads (Beckman Coulter, UK) (0.6 volumes) and QC checked for concentration, size, integrity, and purity using Qubit (Qiagen, UK), Fragment Analyser (Agilent Technologies) and Nanodrop (Thermo Fisher) machines. The samples were then processed without shearing using the PacBio Express kit 1 for library construction and an input of 4 µg DNA following the manufacturer’s protocol. The final library was size-selected using the Sage Blue Pippin (Sage Sciences) 0.75% cassette U1 marker in the range of 25–80 kb. The final library size and concentrations were obtained on the Fragment Analyser before being sequenced using the Sequel 1 2.1 chemistry with V4 primers at a loading on plate concentration of 6 pM and 10 h movie times. For Nanopore sequencing, the ligation sequencing kits SQK-LSK109 or SQK-RAD004 were used as recommended by the manufacturer (Oxford Nanopore Technologies, Oxford, United Kingdom). Starting material for the ligation library preparation were 1–1.5 µg HMW gDNA for the ligation libraries and 400 ng for the rapid libraries. The prepared libraries were loaded onto FLO-PRO002 (R9.4) flow cells. Data collection was carried out using a PromethION Beta with live high accuracy base calling for up to 72 h and with mux scan intervals of 1.5 h. Each sample was sequenced at least twice. Data generated were 7.7 Gb for EgII male, 31.09 Gb for D53 male, 26.72 Gb for VIENNA 7D53−|− male, and 24.83 Gb for VIENNA 8D53−|− male. Run metrics are shown in Supplementary Table 4. The PacBio data were assembled using CANU55 v1.8 with two parameter settings: the first to avoid haplotype collapsing (genomeSize = 500 m corOutCoverage=200 ‘batOptions = -dg 3 -db 3 -dr 1 -ca 500 -cp 50’) and the second to merge haplotypes together (genomeSize = 500 m corOutCoverage=200 correctedErrorRate=0.15). The genome completeness was assessed with BUSCO45,46 v3 using the dipteran gene set45. The two assemblies were found to be duplicated due to alternative haplotypes. To improve the contiguity and reduce duplication, haploMerger2 v20161205 was used56 and the assembly was assessed with BUSCO v3. Phase Genomics Hi-C libraries were made by Phase genomics from males (n = 2) of the same family used for PacBio sequencing. Initial scaffolding was completed by Phase Genomics, but edited using the Salsa57 v2.2 and 3D-DNA (3D de novo assembly pipeline v180419; https://github.com/theaidenlab/3d-dna) software. The resulting scaffolds were allocated a chromosome number using chromosome specific markers16. Specific attention was made to the assembly and scaffolding of chromosome 5. Two contig misassemblies were detected by the Hi-C data and fitted manually. The new assembly (EgII_Ccap3.2.1) was then validated using the Hi-C data. Genes were called using the Funannotate v1.6.0-24f34f6 software making use of the Illumina RNAseq data generated by this project; mRNA mapping to the genome is described below.
To identify possible breakpoint positions, the Nanopore D53 fly assembly contig_531 was mapped onto the EgII_scaffold_5 (from the EgII_CCAP3.2_CANU_Hi-C_scaffolds.fasta assembly) using MashMap v2.0 (https://github.com/marbl/MashMap). This helped to visualize the local alignment boundaries (Supplementary Fig. 10). MashMap parameters were set to kmer size = 16; window size = 100; segment length = 500; alphabet = DNA; percentage identity threshold = 95%; filter mode = one-to-one. Subsequent to this, and to help confirm the exact location of the identified breakpoints, minimap2 (v2.17, https://github.com/lh3/minimap2) was used to align D53 as well as VIENNA 8D53−|− and VIENNA 7D53−|− Nanopore reads onto the EgII scaffold_5 reference (Supplementary Fig. 10). Minimap2 parameters for Nanopore reads were: minimap2 -x map-ont -A 1 -a --MD -L -t 40. Samtools (v1.9, https://github.com/samtools/samtools) was used to convert the alignment.sam to.bam and prepare the alignment file to be viewed in the Integrative Genomics Viewer (IGV, http://software.broadinstitute.org/software/igv/). The expectation was to see a leftmost breakpoint in D53 read set alignments but not in VIENNA 8D53−|− and VIENNA 7D53−|− when compared to the EgII reference (Supplementary Fig. 9). Due to an assembly gap in the EgII scaffold_5 sequence, the exact location of the leftmost inversion breakpoint was not conclusive using this approach. A complementary approach was then used to facilitate detection of the leftmost inversion breakpoint in the D53 inversion line. Minimap2 was again used, but here D53 contig_531 was used as reference for the mapping of EgII male PacBio reads as well as VIENNA 8D53−|− male and VIENNA 7D53−|− male Nanopore reads (Supplementary Fig. 10). Minimap2 parameters for PacBio reads were: minimap2 -x map-pb -A 1 -a --MD -L -t 40. Minimap2 parameters for Nanopore reads were: minimap2 -x map-ont -A 1 -a --MD -L -t 40. Samtools (v1.9, https://github.com/samtools/samtools) was used to convert the alignment.sam to.bam and prepare the alignment file to be viewed in the Integrative Genomics Viewer (IGV, http://software.broadinstitute.org/software/igv/). The expectation was to see a common breakpoint for all three of the above read set alignments when compared to the D53 genome in the area of the inversion. Position ~3,055,294 was identified in the D53 contig_531 as the most likely leftmost breakpoint. To determine the rightmost breakpoint, D53, VIENNA 8D53−|− and VIENNA 7D53−|− male nanopore reads were aligned on the EgII_scaffold_5 sequence. The expectation was to see a breakpoint in D53 read set alignments but not in VIENNA 7D53−|− and VIENNA 8D53−|−. This is the case here, since read alignments coming from both sides of the inversion are truncated at one position (Supplementary Fig. 9). Findings from genome version EgII_Ccap3.2 were extrapolated to the manually revised genome version EgII_Ccap3.2.1.
Predicted D53 inversion breakpoints were verified via PCRs on EgII, D53, and VIENNA 7D53+ GSS male and female genomic DNA, using PhusionFlash Polymerase in a 10 µL reaction volume [98 °C, 10 s; 30 cycles of (98 °C, 1 s; 56 °C, 5 s; 72 °C, 35 s); 72 °C, 1 min] (Supplementary Fig. 4). Sequences of all oligonucleotides used in this study are listed in Supplementary Table 5. The primer pair for the right breakpoint was designed based on EgII sequence information, primers for the left breakpoint were designed based on D53 sequence information. The wild-type status of chromosome 5 (EgII male and female, VIENNA 7D53+|− male) was amplified using primer pairs P_1794 and P_1798 (1950 bp) and P_1795 and P_1777 (690 bp). Chromosome 5 with the inversion (D53 male and female, VIENNA 7D53+|− male and VIENNA 7D53+|+ female) was verified using primer pairs P_1777 and P_1798 (1188 bp) and P_1794 and P_1795 (1152 bp) and amplicon sequencing (Macrogen Europe, Amsterdam).
Transcriptomic analysis of C. capitata, B. dorsalis, and Z. cucurbitae species were then conducted for RNA samples from 3rd instar larval and pre-pupal stages (Supplementary Table 1). Total RNA was extracted by homogenizing three larvae of C. capitata and B. dorsalis and a single larvae of Z. cucurbitae in liquid nitrogen, and then using the RNeasy Mini kit (Qiagen). Three replicates per strain and time point were performed. mRNA was isolated using the NEBNext polyA selection and the Ultra II directional RNA library preparation protocols from NEB and sequenced on the Illumina NovaSeq 6000 using dual indexes as 150 bp paired end reads (library insert 500 bp). Individual libraries were sequenced to provide >1 million paired end reads per sample. Each replicate was then assembled separately using Trinity58 v2.8.5. The assembled transcripts from Trinity were mapped to the Ccap3.2 genome using minimap59 (parameters -ax splice:hq -uf). The Illumina reads were mapped with STAR60 v2.5.2.a. IGV61 v2.6 was used to view all data at a genomic and gene level. Given that the white pupae GSS12,62 was used to collect samples for RNA extraction from single larvae of Z. cucurbitae, larval sex was confirmed by a maleness-specific PCR on the MoY gene of Z. cucurbitae35 using cDNA synthesized with the OneStep RT-PCR Kit (Qiagen) and the primer pair ZcMoY1F and ZcMoY1R amplifying a 214 bp fragment. Conditions for a 25 µL PCR reaction using the 1× Taq PCR Master Mix kit (Qiagen) were: [95 °C, 5 min; 30 cycles of (95 °C, 1 min; 51 °C, 1 min; 72 °C, 1 min); 72 °C, 10 min]. Presence of a PCR product indicated a male sample. Each, male and female sample was a pool of three individuals. Three replicates per strain and time point were collected.
Cytogenetic verification of D53 inversion and wp genes
Polytene chromosomes for in situ hybridization were prepared from third-instar larvae salivary glands63. In brief, the glands were dissected in 45% acetic acid and placed on a coverslip in a drop of 3:2:1 solution (3 parts glacial acetic acid: 2 parts water: 1-part lactic acid) until been transparent (approximately 5 min). The coverslip was picked up with a clean slide. After squashing, the quality of the preparation was checked by phase contrast microscope. Satisfactory preparations were left to flatten overnight at −20 °C and dipped into liquid nitrogen until the bubbling stopped. The coverslip was immediately removed with razor blade and the slides were dehydrated in absolute ethanol, air dried, and kept at room temperature.
Probes were prepared by PCR. Single adult flies were used to extract DNA with the Extract me kit (Blirt SA), following the manufacturer’s protocol. NanoDrop spectrometer was used to assess the quantity and quality of the extracted DNA which was then stored at −20 °C until used. Primers (P_1790/P_1791, P_1821/P_1822, Pgd_probe_F/R, vg1_probe_F/R, Sxl_probe_F/R, y_probe_F/R, zw_probe_F/R, P_1633/P_1634, Zc_F/R, Bd_F/R, P_1395/P_1396, P_1415/P_1416) were designed for each targeted gene using the Geneious Prime software. PCR was performed in a 25 µL reaction volume using 12.5 μL PCR Master mix 2x Kit (Thermo Fisher Scientific), 60–80 ng DNA, and the following PCR settings [94 °C, 5 min; 35 cycles of (94 °C, 45 s; 56 °C, 30 s; 72 °C, 90 s); 72 °C, 1 min].
Probe labeling was carried out according to the Dig DNA Labelling Kit manual (Roche). Prior to in situ hybridization64, stored chromosome preparations were hydrated by placing them for 2 min at each of the following ethanol solutions: 70%, 50%, and 30%. Then they were placed in 2× SSC at room temperature for 2 min. The stabilization of the chromosomes was done by placing them in 2× SSC at 65 °C for 30 min, denaturing in 0.07 M NaOH 2 min, washing in 2× SSC for 30 s, dehydrating (2 min in 30%, 50%, 70%, and 95% ethanol), and air drying. Hybridization was performed on the same day by adding 15 μL of denatured probe (boiled for 10 min and ice-chilled). Slides were covered with a siliconized coverslip, sealed with rubber cement, and incubated at 45 °C overnight in a humid box. At the end of incubation, the coverslip was floated off in 2× SSC and the slide washed in 2× SSC for 3 × 20 min at 53 °C.
After 5 min wash in Buffer 1 (100 mM tris-HCl pH 7.5/ 1.5 M NaCl), the preparations were in Blocking solution (Blocking reagent 0.5% in Buffer 1) for 30 min, and then washed for 1 min in Buffer 1. The antibody mix was added to each slide and a coverslip was added. Then the slides were incubated in a humid box for 45 min at room temperature, following 2× 15 min washes in Buffer 1, and a 2 min wash in detection buffer (100 mM Tris-HCl pH 9.5/ 100 mM NaCl). The color was developed with 1 mL of NBT/BCIP solution during a 40 min incubation in the dark at room temperature. The removal of the NBT/BCIP solution was done by rinsing in water twice. Hybridization sites were identified using 40× or 100× oil objectives (phase or bright field) and a Leica DM 2000 LED microscope, with reference to the salivary gland chromosome maps65. Well-spread nuclei or isolated chromosomes were photographed using a digital camera (Leica DMC 5400) and the LAS X software 3.7.0. All in situ hybridizations were performed at least in duplicates and at least ten nuclei were analyzed per sample.
Gene editing and generation of homozygous wp - strains
For CRISPR/Cas9 gene editing in B. tryoni, purified Cas9 protein (Alt-R S.p. Cas9 Nuclease V3, #1081058, 10 µg/µL) and guide RNAs (customized Alt-R CRISPR/Cas9 crRNA, 2 nmol and Alt-R CRISPR/Cas9 tracrRNA, #1072532, 5 nmol) were obtained from Integrated DNA Technologies (IDT). The guide RNAs were individually resuspended to a 100 μM stock solution with nuclease-free duplex buffer before use. The two customized 20 bp crRNA sequences (Bt_MFS-1 and Bt_MFS-2) were designed using CRISPOR66. Injection mixes for microinjection of B. tryoni embryos comprise of 300 ng/µL Cas9 protein, 59 ng/µL of each individual crRNA, 222 ng/µL tracrRNA, and 1x injection buffer (0.1 mM sodium phosphate buffer pH 6.8, 5 mM KCI) in a final volume of 10 µL. The guide RNA complex containing the two crRNAs and tracrRNA was prepared by heating at 95 °C for 5 min before cooling to room temperature. The Cas9 enzyme along with the other injection mix components were then added to the guide RNA complex and incubated at room temperature for 5 min to assemble the ribonucleoprotein (RNP) complexes. Microinjections were performed in B. tryoni Ourimbah laboratory strain embryos that were collected over a 1 h time period. Injections were performed under paraffin oil using borosilicate capillary needles (#30-0038, Harvard Apparatus) drawn out on a Sutter P-87 flaming/brown micropipette puller and connected to an air-filled 20 mL syringe, a manual MM-3 micromanipulator (Narishige) and a CKX31-inverted microscope (Olympus). Microscope slides with the injected embryos were placed on agar in a Petri dish inside a vented container containing moist paper towels at 25 °C ( ± 2 °C). Hatched first instar larvae were removed from the oil and transferred to larval food. Individual G0 flies were crossed to six virgin flies from the Ourimbah laboratory strain and eggs were collected overnight for two consecutive weeks. G1 flies were then allowed to mate inter se and eggs were collected in the same manner. G2 pupae were then analyzed phenotypically and separated according to color of pupae (brown, mosaic, or white).
For C. capitata CRISPR/Cas9 gene editing, a guide RNA (gRNA_MFS), targeting the third CDS exon of CcMFS was designed and tested for potential off target effects using Geneious Prime53 and the C. capitata genome annotation Ccap2.116. In silico target site analysis predicted an on-target activity score of 0.615 (scores are between 0 and 1; higher score corresponds to higher expected activity67) and zero off-targets sites in the medfly genome. gRNA_MFS was synthesized by in vitro transcription of linear double-stranded DNA template. Therefore, a linear DNA template was amplified in a 100 µL PCR reaction using primers P_1753 and P_369 and Q5 HF polymerase (NEB) according to the manufacturers protocol (Bio-Rad C1000 Touch thermal cycler [98 °C, 30 s; 35 cycles of (98 °C, 10 s; 58 °C, 20 s; 72 °C, 20 s); 72 °C, 2 min]). The PCR reaction was purified using the Clean and Concentrator-25 kit. Subsequently, 500 ng were transcribed using the HiScribe T7 High Yield RNA Synthesis kit (NEB), followed by an DNase treatment (Invitrogen) and a final purification of the RNA using the Mega Clear Kit (Invitrogen). Injection mix for microinjection of embryos contained 360 ng/µL Cas9 protein (1 µg/µL, dissolved in its formulation buffer (PNA Bio Inc, CP01)), 200 ng/µL gRNA_MFS, and an end-concentration of 300 mM KCl68,69. The mix was freshly prepared on ice followed by an incubation step for 10 min at 37 °C to allow pre-assembly of gRNA-Cas9 RNP complexes and stored on ice until use. Microinjections were conducted in WT EgII C. capitata embryos, collected over a 30–40 min period, chemically dechorionized (sodium hypochlorite, 3 min), fixed on double-sided sticky tape (Scotch 3 M), and covered with halocarbon oil 700 (Sigma-Aldrich). For injections, siliconized quartz glass needles (Q100-70-7.5; LOT171381; Science Products, Germany), drawn out on a laser-based micropipette puller (Sutter P-2000), were used with a manual micromanipulator (MN-151, Narishige), an Eppendorf FemtoJet 4i microinjector, and an Olympus SZX16 microscope (SDF PLAPO 1xPF objective). Injected embryos were placed in an oxygen chamber (max. 2 psi), first instar larvae were transferred from the oil to larval food.
As complementation assay, reciprocal crosses between surviving G0 adults and virgin adults of the white pupae strain #1402_22m1B (pBac_fa_attP-TREhs43-Cctra-I-hidAla5-SV40_a_PUb-nls-EGFP-SV40) (wp−(nat))23 were set up either single paired (six cages) or in groups of seven to ten flies (seven cages). Eggs were collected three times every 1–2 days. Progeny (G1) exhibiting the white pupae phenotype (wp−(nat)|−(CRISPR)) were assayed via non-lethal genotyping and sorted according to mutation genotype (see Fig. 4). Genotypes ‘A-H’ were group-backcrossed to WT EgII (wp+|+), genotype ‘C’ siblings mass-crossed. Eggs were collected four times every 1–2 days. Generation G2 flies were analyzed via multiplex PCR using three primers, specific for wp+ and wp−(CRISPR) or wp−(nat) allele size, respectively (see molecular analyses of wp mutants and mosaics, C. capitata non-lethal genotyping). Offspring of outcross cages showed brown pupae phenotype and either wp+|−(nat) or wp+|−(CRISPR) genotype. In order to make mutations A, D, and H homozygous, 40 flies (25 females, 15 males) were genotyped each, and wp+|−(CRISPR) positive flies were inbred (mutation A: 15 females, 7 males, mutation D: 12 females, 7 males, mutation H: 11 females, 8 males). G3 showing white pupae phenotype was homozygous for wp−(CRISPR) mutations A, D, or H, respectively, and was used to establish lines. Inbreeding of mutation C wp−(nat)|−(CRISPR) flies produced only white pupae offspring, based on either the wp−(nat)|−(nat), wp−(nat)|−(CRISPR), or wp−(CRISPR)|−(CRISPR) genotype. 94 flies (46 females, 48 males) were genotyped, homozygous wp−(CRISPR) were inbred to establish a line (13 females, 8 males).
Molecular analyses of wp mutants and mosaics
In B. tryoni, genomic DNA was isolated for genotyping from G2 pupae using the DNeasy Blood and Tissue Kit (Qiagen). PCR amplicons spanning both BtMFS guide recognition sites were generated using Q5 polymerase (NEB) with primers BtMFS_5primeF and BtMFS_exon2R. Products were purified using MinElute PCR Purification Kit (Qiagen), ligated into pGEM-t-easy vector (Promega) and transformed into DH5α cells. Plasmids were purified with Wizard Plus SV Minipreps (Promega) and sequenced.
In C. capitata, non-lethal genotyping was performed to identify parental genotypes before setting up crosses. Therefore, genomic DNA was extracted from single legs of G1 and G2 flies following an adapted version of an established protocol70. Single legs of anesthetized flies were cut at the proximal femur, placed in vials containing ceramic beads and 50 µL buffer (10 mM Tris-Cl, pH 8.2, 1 mM EDTA, 25 mM NaCl), and homogenized for 15 s (6 m/s) using a FastPrep-24 5 G homogenizer. Then, 28.3 µL buffer and 1.7 µL proteinase-K (2.5 U/mg) were added. The reaction mix was incubated for 1 h at 37 °C, followed by 4 min at 98 °C, and subsequently cooled down on ice and used for PCR. For G1 flies, PCR on wp was performed in a 25 µL reaction volume using the DreamTaq polymerase, primers P_1643 and P_1644, and 3.75 µL reaction mix, whereby different amplicon sizes were expected for different alleles (wp+ and wp−(CRISPR): 724 bp, wp−(nat): 8872 bp). The wp−(nat) amplicon was excluded via PCR settings [95 °C, 3 min; 35 cycles of (95 °C, 30 s; 56 °C, 30 s; 72 °C, 1 min); 72 °C, 5 min]. The 724 bp PCR product was verified by gel electrophoresis and purified from the PCR reaction using the DNA Clean & Concentrator-5 kit. PCR products were sequenced (P_1644) and analyzed using Geneious Prime53. In generation G2, flies were analyzed using multiplex PCR with primers P_1657, P_1643, and P_1644, to distinguish between the wp-(nat) (457 bp; P_1643/P_1657), and wp−(CRISPR) alleles (724 bp; P_1643/P_1644) using the above-described PCR protocol.
Image acquisition
Images of B. tryoni pupae were taken with an Olympus SZXI6 microscope, Olympus DP74 camera, and Olympus LF-PS2 light source using the Olympus stream basic 2.3.3 software. Images of C. capitata pupae were taken with a Keyence digital microscope VHX-5000. Image processing was conducted with Adobe Photoshop CS5.1 software to apply moderate changes to image brightness and contrast. Changes were applied across the entire image.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Data supporting the findings of this work are available within the paper and its Supplementary Information files. A reporting summary for this article is available as a Supplementary Information file. The datasets and insect strains generated and analyzed during the current study are available from the corresponding authors upon request. All sequences generated in this study from B. dorsalis, B. tryoni, Bactrocera introgressed line (BIL), C. capitata, and Z. cucurbitae samples are publicly available on NCBI within the ENA BioProject PRJEB36344 (for Ccap genome assembly EgII-3.2.1, WGS, PacBio, chromosome dissections, Illumina MiSeq, Illumina HiSeq 4000, RNAseq, Illumina NovaSeq 6000, Hi-C, and Nanopore data; see Supplementary Table 1 for detailed sample designation), BioProject PRJNA629430 (for WGS and Illumina DNAseq 2 × 250 PE data; see Supplementary Fig. 6 for detailed sample designation), and BioProject PRJNA682907 (for WGS and Illumina NovaSeq 6000 data; see Supplementary Table 1 for detailed sample designation). The source data underlying Figs. 1, 2c–f, 3d–f, and 4d, as well as Supplementary Figs. 3a–b, 4a–b, 4d–e, and 7 are provided as a Source Data file. Source data are provided with this paper.
References
Robinson, A. S. & Hooper, G. Fruit Flies: Their Biology, Natural Enemies, and Control Vol. 1 (Elsevier, 1989).
Suckling, D. M. et al. Eradication of tephritid fruit fly pest populations: outcomes and prospects. Pest Manag. Sci. 72, 456–465 (2016).
Dyck, V. A. et al. Sterile Insect Technique – Principles and Practice in Area-Wide Integrated Pest Management (eds Dyck, V. A., Hendrichs, J. & Robinson, A. S.) (Springer, 2005).
Vreysen, M., Robinson, A. S. & Hendrichs, J. Area-Wide Control of Insect Pests: from Research to Field Implementation (Springer, 2007).
Rendon, P., McInnis, D., Lance, D. & Stewart, J. Medfly (Diptera: Tephritidae) genetic sexing: large-scale field comparison of males-only and bisexual sterile fly releases in Guatemala. J. Econ. Entomol. 97, 1547–1553 (2004).
Franz, G. Sterile Insect Technique – Principles and Practice in Area-Wide Integrated Pest Management (eds Dyck, V. A., Hendrichs, J. & Robinson, A. S.) (Springer, 2005).
Augustinos, A. A. et al. Ceratitis capitata genetic sexing strains: laboratory evaluation of strains from mass‐rearing facilities worldwide. Entomol. Exp. Appl. 164, 305–317 (2017).
Zacharopoulou, A. et al. A review of more than 30 years of cytogenetic studies of Tephritidae in support of sterile insect technique and global trade. Entomol. Exp. Appl. 164, 204–225 (2017).
Rössler, Y. The genetics of the Mediterranean fruit fly: a “white pupae” mutant. Ann. Entomol. Soc. Am. 72, 583–585 (1979).
Rössler, Y. & Koltin, Y. The genetics of the Mediterranean fruit fly, Ceratitis capitata: three morphological mutations. Ann. Entomol. Soc. Am. 69, 604–608 (1976).
McCombs, S. D. & Saul, S. H. Linkage analysis of five new genetic markers of the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). J. Hered. 83, 199–203 (1992).
McInnis, D. O. et al. Development of a pupal color-based genetic sexing strain of the melon fly, Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 97, 1026–1033 (2004).
Wappner, P. et al. White pupa: a Ceratitis capitata mutant lacking catecholamines for tanning the puparium. Insect Biochem. Molec. Biol. 25, 365–373 (1995).
Rössler, Y. & Rosenthal, H. Genetics of the mediterranean fruit fly (Diptera: Tephritidae): morphological mutants on chromosome five. Ann. Entomol. Soc. Am. 85, 525–531 (1992).
Kerremans, P. & Franz, G. Cytogenetic analysis of chromosome 5 from the Mediterranean fruit fly, Ceratitis capitata. Chromosoma 103, 142–146 (1994).
Papanicolaou, A. et al. The whole genome sequence of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann), reveals insights into the biology and adaptive evolution of a highly invasive pest species. Genome Biol. 17, 192 (2016).
Papathanos, P. A. et al. A perspective on the need and current status of efficient sex separation methods for mosquito genetic control. Parasites Vectors 11, 654 (2018).
Sim, S. B. & Geib, S. M. A chromosome-scale assembly of the Bactrocera cucurbitae genome provides insight to the genetic basis of white pupae. G3 7, 1927–1940 (2017).
Sim, S. B., Ruiz-Arce, R., Barr, N. B. & Geib, S. M. A new diagnostic resource for Ceratitis capitata strain identification based on QTL mapping. G3 7, 3637–3647 (2017).
Zdobnov, E. M. et al. OrthoDB v9.1: cataloging evolutionary and functional annotations for animal, fungal, plant, archaeal, bacterial and viral orthologs. Nucleic Acids Res. 45, D744–D749 (2017).
San Jose, M. et al. Incongruence between molecules and morphology: a seven-gene phylogeny of Dacini fruit flies paves the way for reclassification (Diptera: Tephritidae). Mol. Phylogen. Evol. 121, 139–149 (2018).
Choo, A., Crisp, P., Saint, R., O’Keefe, L. V. & Baxter, S. W. CRISPR/Cas9-mediated mutagenesis of the white gene in the tephritid pest Bactrocera tryoni. J. Appl. Entomol. 142, 52–58 (2018).
Ogaugwu, C. E., Schetelig, M. F. & Wimmer, E. A. Transgenic sexing system for Ceratitis capitata (Diptera: Tephritidae) based on female-specific embryonic lethality. Insect Biochem. Mol. Biol. 43, 1–8 (2013).
Davis, A. W. et al. Rescue of hybrid sterility in crosses between D. melanogaster and D. simulans. Nature 380, 157–159 (1996).
Araripe, L. O., Montenegro, H., Lemos, B. & Hartl, D. L. Fine-scale genetic mapping of a hybrid sterility factor between Drosophila simulans and D. mauritiana: the varied and elusive functions of “speciation genes”. BMC Evol. Biol. 10, 385 (2010).
Brideau, N. J. & Barbash, D. A. Functional conservation of the Drosophila hybrid incompatibility gene Lhr. BMC Evol. Biol. 11, 57 (2011).
Kotov, A. A. et al. piRNA silencing contributes to interspecies hybrid sterility and reproductive isolation in Drosophila melanogaster. Nucleic Acids Res. 47, 4255–4271 (2019).
Barbash, D. A. Ninety years of Drosophila melanogaster hybrids. Genetics 186, 1–8 (2010).
Bedo, D. G. & Zacharopoulou, A. Inter-tissue variability of polytene chromosome banding patterns. Trends Genet. 4, 90–91 (1988).
Kriventseva, E. V. et al. OrthoDB v10: sampling the diversity of animal, plant, fungal, protist, bacterial and viral genomes for evolutionary and functional annotations of orthologs. Nucleic Acids Res. 47, D807–d811 (2019).
Zhao, Y. et al. A major facilitator superfamily protein participates in the reddish brown pigmentation in Bombyx mori. J. Insect Physiol. 58, 1397–1405 (2012).
The modEncode Consortium et al. Identification of functional elements and regulatory circuits by Drosophila modENCODE. Science 330, 1787–1797 (2010).
Wright, T. R. The genetics of biogenic amine metabolism, sclerotization, and melanization in Drosophila melanogaster. Adv. Genet. 24, 127–222 (1987).
Bourtzis, K., Psachoulia, C. & Marmaras, V. J. Evidence that different integumental phosphatases exist during development in the Mediterranean fruit fly Ceratitis capitata: possible involvement in pupariation. Comp. Biochem. Physiol. Part B 98, 411–416 (1991).
Meccariello, A. et al. Maleness-on-the-Y (MoY) orchestrates male sex determination in major agricultural fruit fly pests. Science 365, 1457–1460 (2019).
Hall, A. B. et al. Sex determination. A male-determining factor in the mosquito Aedes aegypti. Science 348, 1268–1270 (2015).
Liu, P. et al. Nix is a male-determining factor in the Asian tiger mosquito Aedes albopictus. Insect Biochem. Mol. Biol. 118, 103311 (2019).
Ward, C. M. & Pederson, T. H. S. M. ngsReports: a Bioconductor package for managing FastQC reports and other NGS related log files. Bioinformatics 36, 2587–2588 (2020).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Sedlazeck, F. J., Rescheneder, P. & von Haeseler, A. NextGenMap: fast and accurate read mapping in highly polymorphic genomes. Bioinformatics 29, 2790–2791 (2013).
Garrison, E. & Marth, G. Haplotype-based variant detection from short-read sequencing. arXiv:1207.3907 (2012).
Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 27, 2987–2993 (2011).
Zheng, X. et al. SeqArray-a storage-efficient high-performance data format for WGS variant calls. Bioinformatics 33, 2251–2257 (2017).
Ward, C. M., Ludington, A. J., Breen, J. & Baxter, S. W. Genomic evolutionary analysis in R with geaR. https://doi.org/10.1101/2020.08.06.240754 (2020).
Simão, F. A., Waterhouse, R. M., Ioannidis, P., Kriventseva, E. V. & Zdobnov, E. M. BUSCO: assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 31, 3210–3212 (2015).
Waterhouse, R. M. et al. BUSCO applications from quality assessments to gene prediction and phylogenomics. Mol. Biol. Evol. 35, 543–548 (2018).
Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313 (2014).
Zhang, C., Rabiee, M., Sayyari, E. & Mirarab, S. ASTRAL-III: polynomial time species tree reconstruction from partially resolved gene trees. BMC Bioinforma. 19, 153 (2018).
Martin, S. H. & Van Belleghem, S. M. Exploring evolutionary relationships across the genome using topology weighting. Genetics 206, 429–438 (2017).
Finn, R. D., Clements, J. & Eddy, S. R. HMMER web server: interactive sequence similarity searching. Nucleic Acids Res. 39, W29–W37 (2011).
Li, H. et al. The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Zimin, A. V. et al. The MaSuRCA genome assembler. Bioinformatics 29, 2669–2677 (2013).
Kearse, M. et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649 (2012).
Green, M. R. & Sambrook, J. Isolation of High-Molecular-Weight DNA using organic solvents. Cold Spring Harb. Protoc. 2017, pdb.prot093450 (2017).
Koren, S. et al. Canu: scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 27, 722–736 (2017).
Huang, S. et al. HaploMerger: reconstructing allelic relationships for polymorphic diploid genome assemblies. Genome Res 22, 1581–1588 (2012).
Ghurye, J., Pop, M., Koren, S., Bickhart, D. & Chin, C. S. Scaffolding of long read assemblies using long range contact information. BMC Genomics 18, 527 (2017).
Haas, B. J. et al. De novo transcript sequence reconstruction from RNA-seq using the trinity platform for reference generation and analysis. Nat. Protoc. 8, 1494–1512 (2013).
Li, H. Minimap2: pairwise alignment for nucleotide sequences. Bioinformatics 34, 3094–3100 (2018).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Robinson, J. T. et al. Integrative genomics viewer. Nat. Biotechnol. 29, 24–26 (2011).
Zacharopoulou, A. & Franz, G. Genetic and cytogenetic characterization of genetic sexing strains of Bactrocera dorsalis and Bactrocera cucurbitae (Diptera: Tephritidae). J. Econ. Entomol. 106, 995–1003 (2013).
Zacharopoulou, A. et al. The genome of the Mediterranean fruit fly Ceratitis capitata: localization of molecular markers by in situ hybridization to salivary gland polytene chromosomes. Chromosoma 101, 448–455 (1992).
Mavragani-Tsipidou, P. et al. Protocols for Cytogenetic Mapping of Arthropod Genomes (ed Sakharov, I.) (CRC Press, Taylor and Francis Group, LLC, 2014).
Zacharopoulou, A. Polytene chromosome maps in the medfly Ceratitis capitata. Genome 33, 184–197 (1990).
Concordet, J. P. & Haeussler, M. CRISPOR: intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic Acids Res. 46, W242–W245 (2018).
Doench, J. G. et al. Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation. Nat. Biotechnol. 32, 1262–1267 (2014).
Aumann, R. A., Schetelig, M. F. & Häcker, I. Highly efficient genome editing by homology-directed repair using Cas9 protein in Ceratitis capitata. Insect Biochem. Mol. Biol. 101, 85–93 (2018).
Burger, A. et al. Maximizing mutagenesis with solubilized CRISPR-Cas9 ribonucleoprotein complexes. Development 143, 2025–2037 (2016).
Carvalho, G. B., Ja, W. W. & Benzer, S. Non-lethal PCR genotyping of single Drosophila. Biotechniques 46, 312–314 (2009).
Gariou-Papalexiou, A. et al. Polytene chromosomes as tools in the genetic analysis of the Mediterranean fruit fly, Ceratitis capitata. Genetica 116, 59–71 (2002).
Acknowledgements
This study was financially supported by the Joint FAO/IAEA Insect Pest Control Subprogramme of the Joint FAO/IAEA Programme of Nuclear Techniques in Food and Agriculture, the Emmy Noether program of the German Research Foundation (SCHE 1833/1-1; to MFS), the LOEWE Center for Insect Biotechnology and Bioresources of the Hessen State Ministry for Higher Education, Research and the Arts (HMWK; to MFS), and the SITplus collaborative fruit fly program funded by the Hort Frontiers Fruit Fly Fund, part of the Hort Frontiers strategic partnership initiative developed by Hort Innovation, with co-investment from Macquarie University, USDA, and JLU Gießen and contributions from the Australian Government (FF17000 to MFS). The project was further supported by Hort Innovation, using the Apple & Pear, Strawberry, Citrus, Cherry, Summerfruit, Table Grape and Vegetable research and development levies (MT13059 to PC, AC, EF), with co-investment from South Australian Research and Development Institute (SARDI) and Primary Industries and Regions South Australia (PIRSA) and contributions from the Australian Government. Hort Innovation is the grower-owned, not-for-profit research and development corporation for Australian horticulture. SWB was supported by the Australian Research Council (FT140101303) and the Hermon Slade Foundation grant HSF 18/6. Furthermore, this work was supported by the Canadian Foundation for Innovation (33408, to JR) and Genome Canada Genome Technology Platform awards (JR), as well as the International Atomic Energy Agency research contracts no. 23358 (JR) and no. 23379 (FM) as part of the Coordinated Research Project “Generic approach for the development of genetic sexing strains for SIT applications”. The project benefitted from discussions at this CRP. Furthermore, resources were provided by the SCINet project of the USDA-ARS, the Detection, Control, and Area-wide Management of Fruit Flies and Other Quarantine Pests of Tropical/Subtropical Crops, and a USDA-NIFA grant (0500-00093-001-00-D, 2040-22430-026-00-D, and 2017-67012-26087 to SBS and SMG). The USDA-ARS is an equal opportunity/affirmative action employer, and all agency services are available without discrimination. Mention of commercial products and organizations in this manuscript is solely to provide specific information. It does not constitute endorsement by USDA-ARS over other products and organizations not mentioned. The authors also wish to thank Tanja Rehling, Jakob Martin, and Johanna Rühl for technical assistance and Germano Sollazzo for input on injections and primers design (Justus-Liebig University Gießen and Insect Pest Control Laboratory); Elena Isabel Cancio Martinez, Thilakasiri Dammalage, Sohel Ahmad, and Gülizar Pillwax for insect rearing (Insect Pest Control Laboratory), Shu-Huang Chen (McGill University) for technical assistance with Nanopore library preparations; and Arjen van´t Hof (University of Liverpool) for constructing libraries from micro-dissected chromosomes.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
R.A.A., C.M.W., C.C., P.C., S.B.S., S.M.G., I.H., J.R., A.C.D., K.B., S.W.B., and M.F.S. designed the research; C.M.W., R.A.A., M.A.W., K.N., G.G., E.F., S.J.R., M.A.H., C.C., T.N.M.N., A.C., S.B.S., S.M.G., A.C.D., K.B., S.W.B., and M.F.S. performed the research; R.A.A., C.M.W., H.D., G.L., F.M., J.R., K.B., S.W.B., and M.F.S. contributed new reagents/analytic tools; C.M.W., R.A.A., M.A.W., K.N., G.L., G.G., H.D., S.W., T.N.M.N., A.C., S.B.S., S.M.G., I.H., J.R., A.C.D., K.B., S.W.B., and M.F.S. analyzed the data; R.A.A., C.M.W., K.N., G.L., G.G., S.J.R., S.W., A.C., S.B.S., S.M.G., I.H., J.R., A.C.D., K.B., S.W.B., and M.F.S. wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ward, C.M., Aumann, R.A., Whitehead, M.A. et al. White pupae phenotype of tephritids is caused by parallel mutations of a MFS transporter. Nat Commun 12, 491 (2021). https://doi.org/10.1038/s41467-020-20680-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-020-20680-5
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
