

Abstract
Long-term human disturbance of tropical forests may favor generalist plant species leading to biotic homogenization. We aimed to a) assess if generalist species dominate across different successional ages and topographical positions in a tropical dry forest with a long history of human disturbance, b) to characterize functional traits associated with generalist and specialist species, and c) to assess if a predominance of generalists leads to a homogeneous functional structure across the landscape. We used a multinomial model of relative abundances to classify 118 woody species according to their successional/topographic habitat. Three species were classified as secondary-forest specialists, five as mature-forest specialists, 35 as generalists, and 75 as too rare to classify. According to topography, six species were hill specialists, eight flat-site specialists, 35 generalists, and 70 too rare. Generalists dominated across the landscape. Analysis of 14 functional traits from 65 dominant species indicated that generalists varied from acquisitive strategies of light and water early in succession to conservative strategies in older forests and on hills. Long-term human disturbance may have favored generalist species, but this did not result in functional homogenization. Further analyses considering other functional traits, and temporal and fine-scale microenvironmental variation are needed to better understand community assembly.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence.
Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Human-induced environmental change imposes new filters and dispersal constraints on species. These constraints may favor the spread of broadly tolerant, generalist species, capable of surviving in a wide range of environmental conditions (McKinney and Lockwood 1999, Olden et al 2004). Moreover, forest fragmentation may favor the replacement of species distinctive of old forests by young forest species, which may proliferate across human-modified landscapes (Laurance et al 2006, Santos et al 2008, Lôbo et al 2011). If so, this would lead to greater genetic, taxonomic and functional similarity among communities (i.e. biotic homogenization) (McKinney and Lockwood 1999, Olden et al 2004). The importance of defining the characteristics that differentiate generalist from specialist species increases as human influences on ecosystems become more widespread, frequent and long-lasting (Chazdon et al 2011).
Tropical dry forests (TDF) are among the most threatened ecosystems in the world in part because the severe dry season facilitates the use of fire and slows soil degradation characteristics that are highly valuable for agriculture and other land uses (Janzen 1988). In México, by 2009 more than 70% of TDF potential cover was lost, generating a mosaic of patches of different successional ages (Portillo-Quintero and Sánchez-Azofeifa 2010). In this mosaic, early successional stages have higher temperature and water vapor pressure deficit, and lower surface soil water content compared to older areas (Markesteijn et al 2010, Lebrija-Trejos et al 2011, Pineda-García et al 2013, Buzzard et al 2016). Similarly, soil water content is lower on hills than on flat sites and valleys (Markesteijn et al 2010, Méndez-Alonzo et al 2013).
Functional studies on TDF succession have found that species dominating early in succession, where radiation loads are higher (particularly during the dry season); i.e. young secondary forest specialists, show a greater water use efficiency, with traits associated with heat dissipation through leaf or leaflet movement, and avoidance of hydraulic failure through leaf shedding, high wood density, and/or deeper root foraging capacity (Lebrija-Trejos et al 2010, Pineda-García et al 2011, Méndez-Alonzo et al 2013, Paz et al 2015). Conversely, at later successional stages, the increase in density of individuals and foliage gives rise to a more shaded and humid microenvironment (Lebrija-Trejos et al 2011, Pineda-García et al 2013, Buzzard et al 2016). These changes favor species with an acquisitive strategy, i.e. old growth forest specialists, with traits such as high specific leaf area and low leaf dry matter content (Buzzard et al 2016) associated with higher water availability, or low wood density that increases hydraulic conductivity and water storage capacity (Reich and Borchert 1984, Markesteijn et al 2011).
Concerning topographic position, deciduous tree species with high wood density and water-storing trees with low wood density are predominantly distributed in upland dry forests (Borchert 1994, Méndez-Alonzo et al 2013), whereas evergreen species with either high or low wood density tend to dominate in the more humid lowland sites (Borchert 1994, Méndez-Alonzo et al 2013). However, studies addressing the functional differentiation between young- and old-growth forest specialists (Lebrija-Trejos et al 2010, Alvarez-Añorve et al 2012) or between upland and lowland specialists (Borchert 1994, Méndez-Alonzo et al 2013), have not addressed the functional traits that may facilitate the establishment and survival of generalist species along the complete successional and topographic gradients.
Previous studies in TDF in the Yucatan Peninsula show a high similarity among plant communities, and suggest that pioneer and generalist species may predominate in the mostly early successional habitats due to a prolonged and recurrent use of vegetation over 2000 years, from pre-Columbian Mayas to current inhabitants (Mizrahi et al 1997, González-Iturbe et al 2002, Schultz 2003). Other studies in the Peninsula found that plant species composition and structure were mainly determined by topography, soil properties, and successional age (White and Hood 2004, Dupuy et al 2012a, 2012b, López-Martínez et al 2013).
The aim of this study is to assess if secondary succession and topographic position differentiate species into generalists and specialists in a TDF in Yucatan, and to answer the following questions: Given the millennial land use in our study site, (1) Are generalist species more abundant than specialists over the whole successional and topographical gradients; (2) Do generalist species show primarily acquisitive or conservative strategies; (3) Does a large pool of generalist species lead to a homogeneous functional structure of vegetation across the landscape? To address these questions, we used a multinomial model of relative abundances of woody species across plots, and classified species according to their successional/ topographic habitat into specialists, generalists, or too rare to classify with confidence. We then analyzed functional traits of the dominant species to determine how strategies varied between groups growing in different habitats. We expected that: (1) Generalist species would be more abundant than specialists along the successional and topographical gradients; (2) If disturbance generates habitats that limit growth, generalists will tend to have conservative strategies; conversely, if disturbance releases resources, generalists will tend to be acquisitive. (3) In line with previous studies, conservative species would dominate in sites with the most limiting conditions: early successional sites and hills; whereas acquisitive species would dominate in later-successional and flat sites.
2. Materials and methods
2.1. Study site
The study area covers a 352 km2 landscape of semi-deciduous tropical forest in the Yucatan Peninsula, Mexico, with warm sub-humid AW1 climate (García 1981), summer rains, a dry season from November to April, mean annual temperature of 26 °C and mean annual precipitation between 900 and 1100 mm. The landscape consists of flat areas with relatively deep (40–100 cm) clayey Luvisols and Cambisols and small limestone hills (elevations between 60 and 190 m asl) with shallow (5–20 cm) rocky Lithosols and Rendzines (Flores and Espejel 1994, Bautista-Zúñiga et al 2003). Soil fertility tends to increase with forest stand age and is higher on hills than on flat areas (Dupuy et al 2012b, López-Martínez et al 2013). The predominant land use has been traditional slash and burn agriculture, practiced by the Mayan people for over 2000 years (Rico-Gray and García-Franco 1991). For further details (see Dupuy et al 2012b).
2.2. Sampling design
A Spot 5 satellite image (year 2005) was used to conduct a supervised classification using the following vegetation classes (1) 3 to 8 years of secondary succession, (2) 9 to 15 years, (3) > 15 years on flat areas, and (4) > 15 years on hills (as flat sites are preferred by local farmers) (Dupuy et al 2012a). In the summers of 2008 and 2009, twenty-three 1 km2 landscape units were selected across the landscape, and 12 sampling plots (three per vegetation class) were established in each unit (276 plots in total). Stand age of each plot was determined from interviews with local residents who owned or had used the land. Each plot consisted of a circular 200 m2 area where all woody plants with a diameter at breast height (DBH, measured at 1.3 m height) > 5 cm were identified and measured (1–99 individuals per plot, mean = 33.3, 9129 individuals in total) (see Hernández-Stefanoni et al 2011 for further details).
2.3. Species selection
Relative importance value (RIV) for all species in each vegetation class was calculated, as the sum of their relative abundance, frequency and basal area. For each vegetation class and separately for trees, lianas and shrubs, species that constituted at least 90% of the cumulative RIV were selected, yielding a total of 72 species. However, only 65 species were included for the analyses (those for which we could collect material from at least five individuals), representing 90% ± 10% (mean ± standard deviation) of the total richness per plot (Pakeman and Quested 2007). Of these species, 45 were trees, 11 shrubs, and 9 lianas.
2.4. Functional traits
Leaf samples were collected from at least five individuals per species (14 individuals per species on average, 934 in total) and wood samples from four individuals per species from August to November, 2011 and from September to November, 2012. We calculated total leaf area (LA), minimal photosynthetic unit (MPU, leaf area for entire leaves or leaflet area for compound leaves), leaf dry matter content (LDMC), specific leaf area (SLA) and wood specific gravity (WSG) (for details see online supplementary material 1). The following binary traits were determined from field specimens, parataxonomists, and bibliography: leaf pulvination (LPulv); leaf pubescence (LPb); plant exudates (Ex); plant spininess (Sp; with 0 = absent, 1 = present in all cases); leaf compoundness (LC; 0 = simple, 1 = compound); dispersal syndrome (Dis; 0 = abiotic, 1 = biotic) and deciduousness (LD; 0 = evergreen, 1 = deciduous). Finally, seed volume (SV) was obtained for 55 species as a proxy for seed mass by measuring at least three (and in most cases ten) seeds per individual from samples collected in the field, obtained from the Herbarium at the Centro de Investigación Científica de Yucatán, or from botanical descriptions. Seed volume (SV) was calculated using the formula of an ellipsoid.
2.5. Statistical analyses
We used a multinomial model based on estimated species relative abundances (Chazdon et al 2011) to identify generalist and specialist species, choosing a conservative threshold of k = 2/3, 40 points to define boundaries among groups and a p value of 0.005, as recommended by Chazdon et al (2011). Since the multinomial model classifies species only between two habitats, we performed two analyses, one considering the successional gradient, and another considering the topographical gradient. In the first analysis, abundances of 118 species were compared in 158 young (3–29 y-old) forest plots, versus 53 old-growth forest (30–70 y-old) plots, both groups on flat areas. The successional age limit for old-growth forest was based on a previous finding that abandoned agricultural fields in the Yucatan Peninsula recover a tree species community indistinguishable from extant mature forest in 25–30 years (Turner II et al 2001). The second analysis compared abundances of 119 species in forests on flat areas (106 plots) versus hills (63 plots). We excluded young forest plots from this second analysis (<13 years), because in our study site agriculture is established mainly on flat areas, and we could not find enough replicates of young forests on hills. The multinomial models were performed with the vegan package (Oksanen et al 2016) in R software (R Development Core Team 2015).
To characterize multivariate functional strategies of the 65 most abundant species (for which functional traits were determined), we performed a Principal Component Analysis (PCA) on a species x traits matrix (LA, MPU and PL were log10 transformed). We performed an ANOVA among specialists and generalists PCA scores and post hoc Tukey test to determine if they showed different functional strategies, and compared each trait among groups with Kruskall-Wallis tests for continuous traits and X2 on absolute frequencies for binary traits (Zar 1999). We also calculated the relative abundance of specialists and generalists per plot and performed a Kruskall-Wallis analysis to determine if generalists dominated across forest stand ages and topographic positions.
To analyze the turnover of dominating functional strategies over the successional and topographical gradients, we calculated community-weighted means (hereafter CWM) for each trait, defined as the mean of values present in the community (i.e. plot) weighed by the relative abundance of taxa bearing each value (Lavorel et al 2008), using the LD package (Laliberté et al 2014) in R software. We then performed a PCA on a plot x traits CWM matrix. Since we had seed volume data for only 55 species, we excluded this trait from both PCA analyses. To determine if there were significant differences in trait dominance (CWM) among successional age and topographic position classes, we performed non-parametric Kruskal-Wallis tests and paired comparisons among three vegetation classes (1) 3 to 15 years of secondary succession, (2) 16 to 24 years and (3) ≥ 25 years) and two topographic positions (flat areas and hills) in InfoStat software (Di Rienzo et al 2013). To evaluate how functional dominance of plant strategies varied along secondary succession, we used linear regression of the first two axes of the CWM PCA on successional age. Finally, we analyzed continuous variation of traits (CWM) with successional age by using heteroskedasticity-consistent standard error (HCSE) regressions (HC3 version) separately for plots on flat sites and on hills (online supplementary figure S1, available at stacks.iop.org/ERL/12/055004/mmedia), using the SPSS macro of Hayes and Cai (2007) in SPSS v. 16.0.
3. Results
In the successional gradient, the multinomial model classified three species as secondary forest specialists, five as old-growth forest specialists, 35 as generalists and 75 as too rare to classify with confidence. According to topography, the model identified six hill specialists, eight flat site specialists, 35 generalists and 70 were too rare (table S1). Generalist species dominated all successional ages and topographical positions (p < 0.001, online supplementary figure S2).
Species strategies in the first two axes of PCA (figure 1) showed the functional continuum of species from acquisition to conservation of resources. The first axis (which explained 28.1% of variation) was largely related to the leaf and stem economics spectrum, as it tended to separate acquisitive species with big compound leaves (LA and LC), long petioles (PL), high specific leaf area (SLA), low wood specific gravity (WSG), low leaf dry matter content (LDMC) and abiotic dispersal, from conservative, drought-resistant species with opposite traits (table 1). The second axis (which explained 23% of variation) was associated with traits related to leaf temperature dissipation and plant defense, as it separated conservative deciduous species, mainly with compound leaves, high LDMC, high WSG, very small MPU, pulvini and spines, from more acquisitive species with mostly simple, perennial leaves with high MPU and exudates.
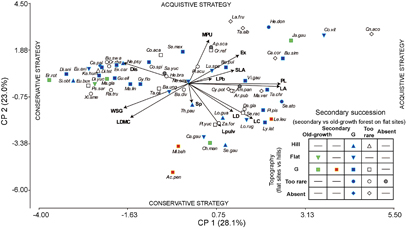
Figure 1 PCA of species according to their functional traits. Symbols represent species classification by the multinomial model: G = generalist; Old-growth = old-growth forest specialist; Secondary = secondary forest specialist; Hill = hill specialist; Flat = flat site specialist; Too rare = too rare to classify with confidence, and Absent = species present in one gradient but not in the other. Functional traits are the same as in table 1. Species (in italics and abbreviated) are in online supplementary table S1.
Download figure:
Standard image High-resolution imageTable 1. Correlations of species traits and CWM with the first two principal components. Italics indicate non-significant correlations (p ≥ 0.05). See eigenvector scores in online supplementary table S4.
| Species | CWM | ||||
|---|---|---|---|---|---|
| Functional traits | PC1 (28.1%) | PC2 (22.7%) | PC1 (28.7%) | PC2 (22.2%) | |
| WSG | Wood specific gravity | −0.61 | −0.44 | −0.72 | 0.58 |
| LC | Leaf compoundness (0 = simple; 1 = compound) | 0.60 | −0.63 | 0.63 | 0.48 |
| Ex | Plant exudates (0 = absent; 1 = present) | 0.47 | 0.51 | 0.55 | −0.54 |
| Sp | Plant spininess (0 = absent; 1 = present) | 0.03 | −0.34 | 0.09 | 0.62 |
| Dis | Dispersal syndrome (0 = abiotic; 1 = biotic) | −0.61 | 0.37 | −0.20 | −0.77 |
| LD | Leaf deciduousness (0 = evergreen; 1 = deciduous) | 0.42 | −0.50 | 0.75 | 0.30 |
| LPb | Leaf pubescence (0 = absent; 1 = present) | 0.22 | 0.09 | 0.04 | 0.14 |
| LPulv | Leaf pulvination (0 = absent; 1 = present) | 0.30 | −0.77 | 0.44 | 0.80 |
| PL | Petiole length | 0.86 | −0.01 | 0.88 | −0.30 |
| MPU | Minimal photosynthetic unit | 0.19 | 0.78 | 0.24 | −0.36 |
| LA | Leaf area | 0.86 | −0.05 | 0.88 | 0.01 |
| LDMC | Leaf dry matter content | −0.55 | −0.61 | −0.60 | 0.13 |
| SLA | Specific leaf area | 0.43 | 0.24 | 0.14 | 0.29 |
Secondary forest specialists (which were all legumes) differed from the other groups –although only in the scores of the second PCA axis (F2,35 = 6.63, p = 0.0036, first axis F2,35 = 0.64, p = 0.53), by traits that favor an efficient use of water, such as smaller MPU (compared to generalists, p = 0.021) and the presence of pulvini (compared to primary forest specialists, p = 0.028). Secondary forest specialist also differed from generalists by the presence of spines (p = 0.002) (see online supplementary tables S2 and S3). Regarding topography, we found no differences among groups either in PCA scores (first axis F2,39 = 0.42, p = 0.66 and second axis F2,39 = 0.24, p = 0.79 respectively) or in traits (tables S2 y S3).
CWM showed that traits associated with light capture and efficient use of water (leaf area, compoundness, deciduousness, pulvination, petiole length) had their highest values in young plots, and decreased with successional age. Conversely, animal seed dispersal, seed volume and LDMC increased with successional age (figures 2, 3 and S1). Considering only old-growth plots (figure 2), hills had more species with pulvini, compound and perennial leaves and spines, higher LDMC, WSG, and lower leaf area, MPU, petiole length and leaf pubescence than flat sites. When considering forest stand age as a continuous variable (figure S1), SLA increased with age on hills while exudates decreased with age on flat sites and hills. Besides, PCA of CWM (figure 3) and regressions of this PCA scores (first two axes) on successional age (figure 4) showed that acquisitive species are dominant at early stages of succession, while more conservative drought resistant species become dominant at latter successional ages. While leaf pulvination and spines are mainly present in the youngest (3–15 year-old) forest plots, exudates and animal seed dispersal predominate at older plots on flat sites and represent the main traits differentiating plots along the second PCA axis (figures 3 and 4).

Figure 2 CWM (mean ± standard error) per successional age category and topographical position. Filled circles correspond to flat sites, while open inverted triangles correspond to hills. Successional age categories are: 1 = plots 3 to 15 years old; 2 = 16 to 24 years old and 3 = > 25 years old. Functional traits as in table 1.
Download figure:
Standard image High-resolution image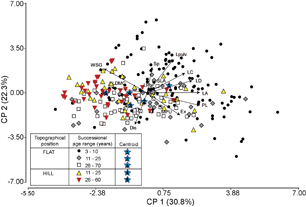
Figure 3 PCA of trait's CWM. Functional traits as in figure 1.
Download figure:
Standard image High-resolution image
{kind=link}
{kind=link}
{kind=link}
Figure 4 Changes in the dominant plant strategies with succession and topographical position gradients. Regression lines were obtained with first and second axes of traits' CWM PCA (figure 3).
Download figure:
Standard image High-resolution image{kind=link}
4. Discussion
4.1. Functional variation across the landscape gradients
Our results show that patterns of plant functional variation in the landscape are driven mostly by generalist species as they dominate (in species number and relative abundance) across successional ages and topographic positions. This agrees with our first prediction and may be due to the long history of disturbance in this region (Rico-Gray and García-Franco 1992, Mizrahi et al 1997, Dupuy et al 2012b), and partly also due to the prevalence of earlier stages of forest succession –158 out of 211 plots (75%) were in young forests.
As described in previous studies for several plant communities (Wright et al 2004, Chave et al 2009, Reich 2014), and particularly in dry forests (Méndez-Alonzo et al 2012, Lohbeck et al 2013, Buzzard et al 2016), species lifestyles are distributed along axes of functional variation between acquisitive and conservative strategies of resource use. Specialists in this forest appear to be highly variable functionally, with the exception of secondary forest specialists, which were all bipinnate legumes. Reyes-García et al (2012) found that large legumes in a TDF in Yucatan had lower whole-tree water use than non-legume species due to higher allocation to non-conductive heartwood in legumes. Powers and Tiffin (2010) found in TDF of Costa Rica that, compared to leaf-habit functional types, legume tree species had a different suite of traits, including high wood density, leaf nitrogen and leaf carbon. In our study, legumes had higher WSG (mean ± SE: 0.784 ± 0.16) than non-legumes (0.694 ± 0.204), but the difference was not significant. Contrary to our second prediction, generalist species were not functionally closer to acquisitive or conservative strategies; instead, they displayed the entire range of variation between both extremes of the resource use continuum (figure 1). This is intriguing and its generality should be explored including forests with higher levels of water limitation, where generalists are expected to be conservative and drought resistant species.
Our results did not support our third prediction: that young forest stands and hills would share conservative functional strategies since they represent the driest habitats in the landscape. Patterns of trait CWM showed that, unlike other dry forest (Buzzard et al 2016), acquisitive species dominate in the youngest plots (figures 2–4). Although dominant species of young forests showed functional traits associated with an efficient use of water (such as large divided leaves with small leaflets, leaf pulvini or leaf deciduousness), they also showed traits (long petioles and large leaf area) associated with an increased efficiency and area for light harvest (Takenaka 1994, Niinemets et al 2004), as found previously in another Mexican TDF (Lohbeck et al 2013). We concur with Lohbeck et al (2013) who suggested that the number of months without leaves and root depth may explain the survival of species with cheap and large leaves in tropical dry forests (Pineda-García et al 2013).
In contrast, species that dominated on hills showed traits associated with a more water-limiting environment, such as the smallest leaves (both in terms of LA and MPU) with high LDMC, and the maximum WSG. This latter trait has been directly associated with high resistance to cavitation, low water storage capacity, and low hydraulic conductivity (Santiago et al 2004, Ishida et al 2008, Markesteijn et al 2011, Pineda-García et al 2013). These distinct strategies of species dominating on hills vs young plots suggest that conditions and resources differ between these portions of the landscape; microenvironmental studies are needed to assess this.
Late successional plots on flat sites also showed more conservative species with small perennial leaves of high LDMC. However, WSG was remarkably similar along succession on flat sites (figures 2 and S1), as also reported for TDF in Costa Rica (Becknell and Powers 2014), but contrary to the lower WSG found at older stands in other Mexican TDF (Lebrija-Trejos et al 2010, Lohbeck et al 2015). It is possible that, in our study area, a fine scale heterogeneity of water availability in the porous and soil-filled cavities in limestone bedrock may allow local niche partitioning of water (Querejeta et al 2007), and consequently the coexistence of low and high wood density species with different rooting strategies across succession, although this also requires further investigation.
4.2. Generalist species dominance does not mean functional homogeneity in the landscape
Even though our results show that considering CWM some ecological strategies are more abundant in certain parts of the landscape, when considered individually most of the species abundances are not significantly different among successional ages or topographic positions. These results indicate that there are multiple ways to be a generalist, and suggest that environmental conditions across the successional and topographical gradient are not sufficiently different compared to the range of conditions in which generalist species can persist and become abundant. However, we cannot discard the possibility that the most influential factors affecting species filtering do not respond to spatial environmental gradients. Temporal variation of environmental resources and conditions could have a more important role determining plant reproduction and recruitment (i.e. the storage effect; Warner and Chesson 1985), which could result in the coexistence of distinctive functional strategies independently of successional age or topographical position.
Furthermore, according to local farmers, all 65 species included in our analyses are capable of sprouting after slash-and-burn agriculture. Sprouting is an important regeneration mechanism (Kammesheidt 1999), especially in a context of recurrent, but spatially patchy and low-intensity shifting cultivation, as practiced for millennia in our study area. It remains to be investigated if sprouting capacity is a key trait for being a generalist, since sprouts have a well-developed root system packed with reserves, which gives them a competitive advantage over seedlings (Miller and Kauffman 1998, Kennard et al 2002).
Finally, more sensitive statistical methods to determine habitat specialization for rare species are needed. It is possible that rare species could be highly sensitive to environmental heterogeneity, while generalist species may be largely tolerant and perhaps common for that reason (Pitman et al 2001). Although the multinomial method has been shown to be more sensitive than other methods in classifying specialist species, and also detects generalists statistically (Chazdon et al 2011), it was unable to classify rare species that represent over 50% of the species in our landscape.
5. Conclusions
Generalist species dominated across the successional and topographic gradient, possibly reflecting the millennial use of land in our studied landscape. However, this did not produce a functional homogenization, since there are many ways to be a generalist. Further studies are needed to evaluate the generality of these findings, especially considering the increasing prevalence of human alterations of all ecosystems, including tropical dry forests. In this study, species that dominated early in succession had acquisitive functional traits, while those that dominated at later successional ages and hills showed more conservative traits, even though almost all species can establish along the complete environmental gradient. Further analyses considering fine scale microenvironmental differences (particularly soil-rock water availability), other key traits such as rooting strategy or sprouting capacity, and the temporal variability of favorable conditions are required to understand community assembly in these tropical dry forests. Finally, new methods to determine habitat specialization for rare species are needed, particularly for tropical forests and other species-rich ecosystems, where most of the species are represented by few individuals.
Acknowledgments
We thank the communities of Xkobenhaltún, Xul and Yaxhachén for allowing us to work in their lands. James Callaghan and Kaxil Kiuic A C provided logistic support. We are grateful to Filogonio May-Pat, Roberth Armando Us Santamaría and Luis Simá Gómez for their help with fieldwork, and to Santos Uc Uc for his help with species identification and fieldwork. The comments of Jennifer Powers and an anonymous reviewer helped improve the clarity and reach of this paper. LSV acknowledges CONACYT, Mexico, for the grant awarded (169510).