





Abstract
The benefits of colour change are expected to vary seasonally because of changes in reproductive activity, temperature and, potentially, predation risk; yet temporal variation in colour change has seldom been examined. We measured colour change in spring and autumn using captive individuals from two differently coloured populations of the central bearded dragon lizard, Pogona vitticeps. We predicted that colour change should be greater in spring than autumn because of the added requirements of reproductive and territorial activity. To elicit colour change in a standardized way, we placed lizards inside temperature-controlled chambers and measured colour at 15, 25, 35 and 40 °C, repeating experiments in spring and autumn. Lizards from both populations changed from dark grey to light yellowish or orange-brown (increasing luminance and saturation) with increasing temperature in both seasons, and both populations changed colour to a similar extent. As predicted, the maximal extent of temperature-induced colour change (in particular, luminance change) was greater in spring than autumn. Our results confirm that temperature-induced colour change is greater in the peak activity season, probably an adaptation to the greater thermal and/or signalling needs of that time of year.
INTRODUCTION
Colour change is a widespread form of phenotypic plasticity used to accommodate changes in the biotic and abiotic environment in space and time (Stuart-Fox & Moussalli, 2009; Duarte, Flores & Stevens, 2017). The time scale of change will depend on both the nature and the predictability of environmental variation. For example, ontogenetic and seasonal changes in integument colour may take up to months to achieve and are the result of an increase or decrease in pigment concentration and/or the quantity of chromatophores (Green, 1964). These morphological changes are common in species that experience major, predictable shifts in background colour or predator types at different life-history stages or in different seasons (e.g. blue tits, Örnborg et al., 2002; Arctic fox, Stoner, Bininda-Emonds & Caro, 2003; and some arthropods, Umbers et al., 2014). In contrast, rapid colour changes may take only seconds to minutes to occur and are effected through rapid dispersion or aggregation of pigments within the dermal chromatophores (Bagnara & Matsumoto, 2007; Teyssier et al., 2015). These rapid, physiological colour changes enable animals to respond to fine-scale spatial or temporal variation in background colour, changes in thermal requirements throughout the day, and to signal conspecifics or predators (Caro, Sherratt & Stevens, 2016). For example, chameleons, cephalopods and many other species use colour change to display conspicuous colours to mates or rivals during social interactions, but remain camouflaged at other times (Stuart-Fox, Whiting & Moussalli, 2006; Stuart-Fox & Moussalli, 2008). Likewise, bearded dragon lizards change colour for camouflage and signalling (Smith et al., 2016a, 2016b), as well as to reach active body temperatures more rapidly (Smith et al., 2016b).
Although rapid physiological colour change allows some animals to accommodate the multiple functions of colour, the benefits and potential costs of colour change are expected to vary seasonally because of changes in reproductive or social activity, temperature and, potentially, predation risk. For example, the benefits of colour change for social signalling might be lower outside the breeding season, and thermoregulatory benefits might be lower or higher during periods of reduced activity. Metabolic costs associated with short-term movement of pigments within pigment cells, pigment deposition and pigment cell synthesis have not been quantified but are likely to be dependent on the frequency of these changes. Occasional events are likely to incur minimal costs, but a high frequency of these events might impose higher associated costs. Metabolic costs of longer-term changes in pigment deposition and pigment cell synthesis can be significant, as recently shown for melanin synthesis (Talloen, Van Dyck & Lens, 2004; Ethier et al., 2015). Furthermore, seasonal variation in gonadal hormones, such as testosterone, can influence hormones that trigger colour change, notably α-melanocyte stimulating hormone (α-MSH; Castrucci et al., 1997; Nery & Castrucci, 1997). Examining temporal variation in phenotypic plasticity, including rapid colour change, is important for understanding its constraints and evolution; yet studies are seldom repeated at more than one point in time.
The central bearded dragon, Pogona vitticeps (Ahl 1926), is well known for its rapid colour change, which functions in thermoregulation, signalling and camouflage (de Velasco & Tattersall, 2008; Smith et al., 2016a, 2016b; Cadena et al., 2017b). The species is a large (ranging in size across its geographical range; 15–25 cm snout–vent length; 150–600 g) diurnal, semi-arboreal lizard that is widely distributed across semi-arid habitats in central, southern and eastern Australia. It is an omnivorous sit-and-wait forager, and males defend territories during the breeding season from conspicuous vantage points (e.g. fence posts, dead tree trunks). As in all seasonally reproducing lizards, signalling associated with territorial defence and courtship is most pronounced in the breeding season (Brattstrom, 1971). Both sexes change colour; during social interactions, the beard and chest changes from grey or cream to black and the colour of the rest of the body changes from grey to cream, yellow or rich orange, depending on the population (Smith et al., 2016a, 2016b). Apart from changes during social interactions, experimental studies have demonstrated colour change in response to temperature (de Velasco & Tattersall, 2008; Smith et al., 2016a, 2016b), circadian rhythms (Fan, Stuart-Fox & Cadena, 2014), social interactions (Smith et al., 2016b) and different background colours (Cadena et al., 2017b). Populations vary in colour, being locally adapted to different coloured substrates (e.g. orange-red sands of central Australia vs. cream sands and more vegetated environments in southern and eastern populations), but do not appear to differ in their capacity for colour change (Cadena et al., 2017b).
In this study, we tested whether the extent and nature of temperature-induced colour change differs between the spring reproductive season and autumn, after reproductive activity has ceased. To assess potential population variation in seasonal colour change, we measured colour change using captive individuals from two populations (Alice Springs and Mildura; Fig. 1) which differ in coloration, habitat and thermal environment. Lizards from Alice Springs, where orange-red sands predominate, tend to be more orange-red, whereas lizards from Mildura, where cream-yellow sands predominate, better match those backgrounds (Cadena et al., 2017b). Dark grey backgrounds (tree trunks, dead vegetation) are common in both populations, and lizards show a similar extent of background-dependent colour change (Cadena et al., 2017b). To compare the nature and extent of individual colour change at different time points, it was important to elicit colour change in a standardized, repeatable way. Varying temperature reliably elicits colour change in this species (Smith et al., 2016b); therefore, we placed lizards in temperature-controlled chambers and measured colour at 15, 25, 35 and 40 °C, repeating experiments in spring and autumn. We predicted that temperature-induced colour change would be greater in the period of peak reproductive and territorial activity (spring), when the thermal and signalling advantages of colour change are also likely to be greater.
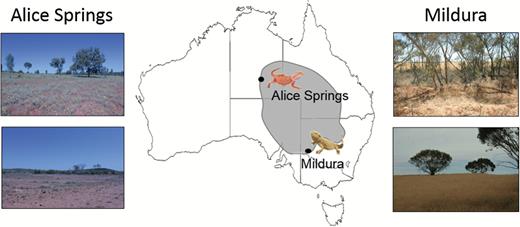
Geographical distribution of the two bearded dragon populations used in this study. Overall distribution of Pogona vitticeps is indicated by the grey area.
MATERIALS AND METHODS
Animals
Adult male bearded dragons, P. vitticeps, were hand captured in late September and early October 2012 from the vicinities of Mildura (34.18°S, 142.15°E; N = 11) and Alice Springs (23°42′S, 133°52′E; N = 11), Australia, and transported to indoor animal housing facilities at the School of BioSciences at The University of Melbourne. Bearded dragons were individually housed in sand-filled terraria fitted with an ultraviolet (UV) lamp and an incandescent basking lamp for heating, set to a 12 h–12 h light–dark photoperiod (lights on at 07.00 h), approximating the natural light conditions experienced by bearded dragons during breeding and post-breeding periods (spring and autumn). During the light phase, temperature inside the terrarium ranged between 25° (under the hiding place) and 50 °C (directly under the incandescent light), which allowed for behavioural thermoregulation; these temperatures are within the natural range experienced by bearded dragons in the wild (Smith et al., 2016a) and include the range of voluntary active body temperatures in the field (32–40 °C; Smith et al., 2016b). Each terrarium was also enriched with a hiding place and a natural tree branch for perching. Lizards were provided with water ad libitum and fed a diet of crickets and commercial bearded dragon food (10.04 URS Lizard Food; Ultimate Reptile Supplies, Australia) mixed with chopped green leafy vegetables, carrots and pumpkin, three times a week.
During the winter months, bearded dragons in the wild enter a state of brumation, a sleep-like state similar to hibernation in mammals (Mayhew, 1965). To simulate natural behaviour and physiology in our captive lizards, we promoted brumation by introducing changes in their enclosures that mimicked winter in their natural environment; over the course of 3 weeks we gradually decreased the temperature in the room to 13 °C, reduced the length of time the lights were on, and reduced the amount of food until completely withdrawn. After 1 week in these conditions, all lizards had entered brumation and remained in this state from the beginning of June until the end of July 2013, when lighting and temperature were gradually returned to original conditions. Lizards were healthy and in good body condition, and all resumed normal eating, and thermoregulatory and social behaviours. All males, for example, performed courtship displays (e.g. head bobbing, chasing, beard darkening) when presented with our captive female bearded dragons (not used in this study).
Lizards from Mildura were obtained under Department of Sustainability and Environment Victoria permit number 10006453 and lizards from Alice Springs under Parks and Wildlife Commission Northern Territory permit number 44582. The Animal Ethics Committee of The University of Melbourne approved all experimental procedures (AEC no. 1212547).
Experimental procedures
Experiments were performed during the Austral spring (from September to November 2013) and autumn (April and early May 2013) before brumation. During experiments, lizards were placed inside a small temperature-controlled chamber (32 cm × 36 cm × 45 cm; Exo-Terra Thermoelectric Egg Incubator; Rolf C. Hagen Corp., USA) for a minimum of 45 min at each of four temperatures: 15, 25, 35 and 40 °C, sequentially in that order. A period of 45 min is considered sufficient for thermal equilibrium to occur in bearded dragon lizards (Tattersall & Gerlach, 2005) and for skin colour to stabilize after initial abrupt changes attributable to handling stress (Cadena et al., 2017b). We took photographs at 30 s intervals using a remotely operated digital camera (Canon EOS 600D; ISO 200; shutter speed 1/15; aperture 3.5) mounted on the ceiling of the chamber. We measured colour from photographs taken remotely rather than using a spectrometer to ensure that lizards were not disturbed. Bearded dragons have minimal UV reflectance, and near-simultaneous photographs and spectral measurements for this species have confirmed close correspondence of colours in RGB colour space and in avian or lizard visual colour space (Smith et al., 2016a). Photographs were obtained at 30 s intervals to monitor colour change as lizards reached thermal equilibrium at each of the four ambient temperatures. Preliminary analysis showed, however, that use of only the last photograph at each temperature was sufficient to obtain an accurate measurement of lizard stable skin colour, at full thermal equilibrium. Therefore, we analysed only the final photograph in the series of each 45 min period at a given temperature.
We obtained average red (R), green (G) and blue (B) from the dorsal surface of the lizard with custom Matlab script (Mathworks Inc., Natick, MA, USA; written by John Endler). To ensure a linear relationship between camera responses (R, G and B) and reflectance, raw RGB values were calibrated using six different grey standards of known reflectance (ColorChecker Passport, X-Rite, Grand Rapids, MI, USA). To linearize camera responses, we applied a function in the form of y = a × expbx + c × expdx, where y is the linearized pixel value, and a, b, c and d are empirically derived constants specific to our camera (Garcia et al., 2013). We then calculated the standardized differences between the calibrated red (R) and green (G) channels as (R − G)/(R + G + B) and between the green and blue (B) channels as (G − B)/(R + G + B) to provide a two-dimensional representation of colour space, where the distance from the origin corresponds to chroma and the angle relative to the axis corresponds to hue (Endler, 1990; Grill & Rush, 2000). Physical chroma was calculated as r = (x2 + y2)½ and physical hue as Θ = tan−1(y/x), where x and y are the standardized differences between red-green (R-G) and green-blue (G-B) channels, respectively (Endler, 1990), using the Matlab function cart2pol (Cartesian to polar coordinates). Luminance was calculated as the sum of the calibrated R, G and B values. As the colour of lizards was relatively uniform, we calculated mean dorsal coloration for each colour component (hue, chroma and luminance).
To estimate the degree of temperature-induced change in hue, chroma or luminance for each individual, we took the maximum difference between values at 15, 25, 35 and 40 °C. In the great majority of instances, the maximal individual change in hue, chroma or luminance was between 15 and 40 °C.
Statistical analysis
We first examined differences in hue, chroma and luminance between temperatures, seasons and populations using generalized linear mixed models (GLMMs; Bates et al., 2013). We used each of the colour variables (hue, chroma and luminance) as the dependent variables, and temperature (15, 25, 35, 40 °C), season (spring or autumn), population (Alice Springs or Mildura) and their interactions as the independent variables (fixed factors). Lizard identity was included as a random factor to account for repeated measures on individuals. Although hue is an angle, and angles are usually tested with circular statistics, the range of values in our study was small and does not cross 0°, so standard statistics based upon hue are valid in this study. Differences between temperatures were explored using Tukey’s post hoc tests (adjusted P-values reported).
To test whether the extent of temperature-induced colour change differed between seasons and populations, we again ran LMMs with maximal hue, chroma or luminance change over the four temperatures as the dependent variable and season, population and their interaction as fixed factors. Lizard identity was included as a random factor to account for repeated measures on individuals.
The final models included interaction terms only when significant at P < 0.05. Homoscedasticity of the data was verified through visual examination of residual and QQ plots (Zuur, Ieno & Elphick, 2010).
RESULTS
Lizards from both populations showed temperature-induced colour change in both seasons and seasonal differences in colour (Fig. 2). For both luminance and chroma, there was a significant effect of temperature and season but not for their interaction (Table 1). In spring, lizards from both populations had higher and much more variable luminance (Fig. 3A) and lower chroma (Fig. 3B) than in autumn. In both seasons, luminance increased with increasing temperature, as one would expect from a thermoregulatory function (Table 1; Fig. 3A). The greatest increase in luminance between consecutive temperatures occurred from 35 to 40 °C (t150 = −1.92, P = 0.22), and the increases over larger temperature ranges, namely from 15 or 25 to 40 °C, were statistically significant (t150 = −4.13, P = 0.003 and t150 = −3.68, P = 0.002, respectively). Chroma increased in a similar manner with increasing temperature in both seasons (Table 1; Fig. 3B). The greatest increase in chroma between consecutive temperatures occurred from 15 to 25 °C (t150 = −3.87, P = 0.0009); increases from 25 to 35 °C and from 35 to 40 °C were not statistically significant (t150 = −0.59, P = 0.93 and t150 = −2.0, P = 0.19, respectively).

Digital photographs of the same individual from Alice Springs. Photographs were taken in autumn after 45 min at each temperature treatment: A, 15 °C; B, 25 °C; C, 35 °C; and D, 40 °C.
Differences in chroma, luminance and hue in response to population of origin (Alice Springs or Mildura, N = 11 for each), season (spring or autumn) and temperature (15, 25, 35 and 40 °C)
| Dependent | Independent | Fd.f. | P-value |
|---|---|---|---|
| Chroma | Population | 1.071,150 | 0.30 |
| Season | 16.771,150 | < 0.0001* | |
| Temperature | 14.583,150 | < 0.0001* | |
| Luminance | Population | 0.0001,150 | 0.99 |
| Season | 131.371,150 | < 0.0001* | |
| Temperature | 7.093,150 | 0.0002* | |
| Hue | Population | 7.781,147 | 0.006* |
| Season | 34.891,147 | < 0.0001* | |
| Temperature | 0.763,147 | 0.52 | |
| Season × temperature | 2.853,147 | 0.04* |
| Dependent | Independent | Fd.f. | P-value |
|---|---|---|---|
| Chroma | Population | 1.071,150 | 0.30 |
| Season | 16.771,150 | < 0.0001* | |
| Temperature | 14.583,150 | < 0.0001* | |
| Luminance | Population | 0.0001,150 | 0.99 |
| Season | 131.371,150 | < 0.0001* | |
| Temperature | 7.093,150 | 0.0002* | |
| Hue | Population | 7.781,147 | 0.006* |
| Season | 34.891,147 | < 0.0001* | |
| Temperature | 0.763,147 | 0.52 | |
| Season × temperature | 2.853,147 | 0.04* |
Colour components were derived from digital images. * denotes statistical significance. Pairwise interactions were included in initial models but removed from final models if they did not improve model fit (see Material and methods).
Differences in chroma, luminance and hue in response to population of origin (Alice Springs or Mildura, N = 11 for each), season (spring or autumn) and temperature (15, 25, 35 and 40 °C)
| Dependent | Independent | Fd.f. | P-value |
|---|---|---|---|
| Chroma | Population | 1.071,150 | 0.30 |
| Season | 16.771,150 | < 0.0001* | |
| Temperature | 14.583,150 | < 0.0001* | |
| Luminance | Population | 0.0001,150 | 0.99 |
| Season | 131.371,150 | < 0.0001* | |
| Temperature | 7.093,150 | 0.0002* | |
| Hue | Population | 7.781,147 | 0.006* |
| Season | 34.891,147 | < 0.0001* | |
| Temperature | 0.763,147 | 0.52 | |
| Season × temperature | 2.853,147 | 0.04* |
| Dependent | Independent | Fd.f. | P-value |
|---|---|---|---|
| Chroma | Population | 1.071,150 | 0.30 |
| Season | 16.771,150 | < 0.0001* | |
| Temperature | 14.583,150 | < 0.0001* | |
| Luminance | Population | 0.0001,150 | 0.99 |
| Season | 131.371,150 | < 0.0001* | |
| Temperature | 7.093,150 | 0.0002* | |
| Hue | Population | 7.781,147 | 0.006* |
| Season | 34.891,147 | < 0.0001* | |
| Temperature | 0.763,147 | 0.52 | |
| Season × temperature | 2.853,147 | 0.04* |
Colour components were derived from digital images. * denotes statistical significance. Pairwise interactions were included in initial models but removed from final models if they did not improve model fit (see Material and methods).
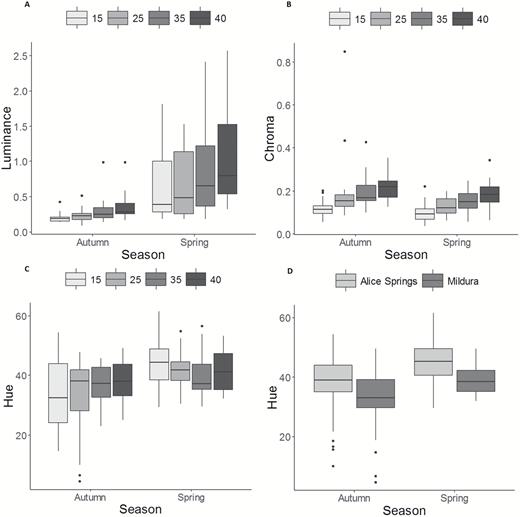
Colour properties of bearded dragon lizards in autumn and spring during exposure to different temperature treatments (15, 25, 35 and 40 °C). A (luminance), B (chroma) and C (hue) show lizards for both populations combined, whereas D (hue) shows each population in autumn and spring; note the higher hue of lizards from Alice Springs (more orange) than from lizards from Mildura (yellower). Boxplots show the median, and first and third quartiles of the range of the data. Whiskers extend from the upper and lower hinges to the largest and smallest values, respectively, no further than 1.5 times the interquartile range. Data points beyond the end of boxes hinges are considered outlier values and are plotted as individual dots.
For hue, there was a significant interaction between temperature and season (Table 1; Fig. 3C). Lizards were more orange (larger hue angle) in spring than autumn at 15 and 25 °C (t150 = −4.89, P < 0.0001 and t150 = −3.84, P = 0.004, respectively), but there was no difference in hue between seasons at 35 and 40 °C (t150 = −1.53; P = 0.79 and t150 = −1.55; P = 0.78, respectively). Within seasons, there were no significant differences in hue between any of the temperature treatments (P > 0.1 in all instances; Fig. 3C). Additionally, hue showed a significant population effect (Table 2; Fig. 3D), indicating that lizards from the two populations (Alice Springs and Mildura) show different coloration independent of season and temperature; lizards from Alice Springs tend to be more orange, whereas those from Mildura are more yellow.
Differences in the maximal extent of temperature-induced colour (chroma, luminance and hue) change between populations (Alice Springs and Mildura, N = 11 for both) and seasons (spring and autumn)
| Dependent | Independent | Fd.f. | P-value |
|---|---|---|---|
| Chroma | Population | 0.881,21 | 0.36 |
| Season | 3.391,21 | 0.08 | |
| Luminance | Population | 0.851,21 | 0.37 |
| Season | 20.471,21 | 0.0002* | |
| Hue | Population | 1.081,21 | 0.31 |
| Season | 11.301,21 | 0.003* |
| Dependent | Independent | Fd.f. | P-value |
|---|---|---|---|
| Chroma | Population | 0.881,21 | 0.36 |
| Season | 3.391,21 | 0.08 | |
| Luminance | Population | 0.851,21 | 0.37 |
| Season | 20.471,21 | 0.0002* | |
| Hue | Population | 1.081,21 | 0.31 |
| Season | 11.301,21 | 0.003* |
Colour components were derived from digital images. * denotes statistical significance. Pairwise interactions were included in initial models but removed from final models if they did not improve model fit (see Material and methods).
Differences in the maximal extent of temperature-induced colour (chroma, luminance and hue) change between populations (Alice Springs and Mildura, N = 11 for both) and seasons (spring and autumn)
| Dependent | Independent | Fd.f. | P-value |
|---|---|---|---|
| Chroma | Population | 0.881,21 | 0.36 |
| Season | 3.391,21 | 0.08 | |
| Luminance | Population | 0.851,21 | 0.37 |
| Season | 20.471,21 | 0.0002* | |
| Hue | Population | 1.081,21 | 0.31 |
| Season | 11.301,21 | 0.003* |
| Dependent | Independent | Fd.f. | P-value |
|---|---|---|---|
| Chroma | Population | 0.881,21 | 0.36 |
| Season | 3.391,21 | 0.08 | |
| Luminance | Population | 0.851,21 | 0.37 |
| Season | 20.471,21 | 0.0002* | |
| Hue | Population | 1.081,21 | 0.31 |
| Season | 11.301,21 | 0.003* |
Colour components were derived from digital images. * denotes statistical significance. Pairwise interactions were included in initial models but removed from final models if they did not improve model fit (see Material and methods).
There were significant differences in the maximal extent of temperature-induced luminance and hue change between seasons (Table 2; Figs 1, 4A, C). As predicted, lizards showed substantially greater luminance change in spring than autumn (Table 2). In contrast, the maximal extent of hue change was greater in autumn than spring, even though differences between temperature treatments were not statistically significant in either season (Fig. 3C). There was no difference in temperature-induced chroma change between seasons (Table 2; Fig. 4B). Seasonal differences in the extent of colour change were consistent in the two populations; there were no differences between populations in the maximal extent of temperature-induced hue, chroma or luminance change, and no interaction between population and season (Table 2).
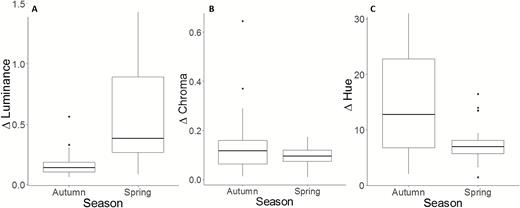
Maximal extent of temperature-induced colour change in bearded dragons in autumn and spring. Maximal change in luminance (A), chroma (B) and hue (C). Boxplots show the median, and first and third quartiles of the range of the data. Whiskers extend from the upper and lower hinges to the largest and smallest values, respectively, no further than 1.5 times the interquartile range. Data points beyond the end of boxes hinges are considered outlier values and are plotted as individual dots.
DISCUSSION
Rapid physiological colour change, a visually striking form of phenotypic plasticity, may vary seasonally in relationship to social and reproductive activity and environmental conditions. We assessed the nature and extent of temperature-induced colour change in two populations of bearded dragons in spring and autumn. Despite the different colour of lizards in the two populations, both populations showed consistent changes between temperatures and seasons. Lizards from both populations showed higher, more variable luminance, as well as greater changes in this parameter in spring than in autumn. As chroma is negatively correlated with luminance (a necessary property of reflectance spectra), lizard colour was also less saturated (lower chroma) in spring; however, they showed similar temperature-induced chroma change in the two seasons. In contrast, lizards showed greater change in hue in autumn than spring but had similar hue at 35 and 40 °C in both seasons. Overall, these results confirm seasonal differences in the extent and nature of colour change, consistent with the prediction that benefits of colour change should be greater in the reproductive season, when bearded dragons have a higher need for signalling conspecifics and experience higher variations in ambient temperature because of higher levels of activity.
As predicted, luminance was higher and changed more in spring than autumn. Luminance is the most important colour component affecting absorption of solar radiation and therefore heat transfer (Smith et al., 2016b). Bearded dragons use colour change for thermoregulation in the wild; free-ranging bearded dragons show changes in colour in relationship to both skin and core body temperature (Smith et al., 2016a). Biophysical models of heat transfer indicate that the thermal benefits of colour change for bearded dragons at Mildura are generally higher in spring, although they are also very variable in both seasons depending on whether conditions enable a basking lizard to reach its minimal activity threshold (Smith et al., 2016b). Colour change allows lizards to reach active temperatures more quickly, which might have greater fitness benefits in spring, when social and foraging activity is higher. The lower chroma of lizards in spring is consistent with higher luminance, because lighter colours tend to be less saturated. However, hue, which is less affected by changes in luminance, was similar in the two seasons for lizards at 35 and 40 °C, the two temperature treatments within the range of voluntary active body temperatures in the field (32–40 °C; Smith et al., 2016b). Overall, these results indicate a greater seasonal effect of temperature-induced colour change on luminance, consistent with expectations of seasonal variation in the thermal benefits of colour change. Luminance and chroma also show the greatest change for background matching (Smith et al., 2016a). If predation risk varies seasonally, this might alter potential trade-offs between crypsis and thermoregulation; although this possibility remains to be tested.
Colour change also plays an important role in signalling in bearded dragons, particularly during the breeding season. During social interactions, the beard and chest change from grey or cream to jet black and the dorsal surface becomes much lighter and more intensely coloured. Thus, the greater change in luminance in spring might also reflect greater need for signalling, although we found no difference in the extent of chroma change between seasons. We examined only temperature-induced colour change, because it can be measured in a standardized and repeatable way; however, bearded dragons show substantially greater colour change during social interactions than in response to temperature. Bearded dragons from Mildura show a maximum of 15% change in overall dorsal reflectance (equivalent to luminance) from 15 to 40 °C, but up to 23% change during social interactions (Smith et al., 2016b).
Colour change for signalling may be influenced by short- and long-term changes in gonadal hormones (possibly induced by brumation) associated with social and reproductive activity (e.g. testosterone), because gonadal hormones can mediate effects of α-MSH, the primary melanotrophic hormone that induces skin darkening (Nery & Castrucci, 1997). For example, in tree lizards, Urosaurus ornatus, the sensitivity of skins to α-MSH in vitro decreased ninefold in the non-breeding season compared with the breeding season, presumably owing to effects of testosterone (Castrucci et al., 1997). Thus, seasonal variation in the extent of temperature-induced colour change may reflect physiological differences associated with reproductive activity, in addition to seasonal differences in the thermal or signalling benefits of colour change.
The marked change in luminance in response to temperature is consistent with underlying mechanisms. In reptiles, colour change is primarily caused by the dispersion or aggregation of melanin-containing organelles (melanosomes) within melanophores, but may also be influenced by changes to spacing of reflecting platelets within iridophores (Bagnara & Matsumoto, 2007; Teyssier et al., 2015). Changes in both melanosome dispersion and iridosome spacing primarily cause changes in luminance, but also influence hue and chroma. Furthermore, temperature influences the release of α-MSH, which triggers melanin dispersion (Fernandez & Bagnara, 1991; Nery & Castrucci, 1997; Höglund, Balm & Winberg, 2002). If seasonal variation in colour change involves melanin synthesis as well as short-term changes in dispersion, then the associated metabolic costs of darker coloration in autumn must be offset by greater signalling and thermal advantages in spring.
Overall, our study demonstrates seasonal differences in temperature-induced colour change, consistent with greater thermal and/or signalling benefits in spring than autumn. These effects were similar in two populations, each locally adapted to different background colours (Cadena et al., 2017b) and occupying thermally different habitats. An interesting avenue for further research is to examine seasonal variation in the metabolic or physiological costs of colour change. This is particularly relevant if colour change is mediated by testosterone or other hormones that are known to incur high physiological costs (Zera & Harshman, 2001; Perez-Rodriguez et al., 2006; Alonso-Alvarez et al., 2007).
SHARED DATA
Data for this study have been archived in Dryad (Cadena, Rankin, Smith, Endler & Stuart-Fox, 2017a).
ACKNOWLEDGEMENTS
We are grateful to Adam Elliott for his help with lizard collection, to Stefania Milano and Ashton Dickerson for their assistance with data acquisition and to Christopher Gatto, Jacob Gardiner, Vivian Truong, Georgia Goodchild, Anna Lewis, Silvia Swan and Jess Rowland for their help with photographic processing and animal care. We also extend our thanks to three anonymous reviewers for their insightful comments on a previous version of the manuscript. This study was supported by an Australian Research Council grant (DP120100105) to D.S.-F. and J.A.E. V.C., J.A.E. and D.S.-F. contributed to the research conception and design; V.C. and K.R.S. performed the experiments; J.A.E. provided photographic analysis methods and Matlab scripts; V.C. and K.R. processed raw data and conducted statistical analysis; V.C., K.R. and D.S.-F. wrote the manuscript; all authors revised and edited the manuscript. The authors declare no competing or financial interests.

{kind=link}
{kind=link}
{kind=link}
{kind=link}