





Abstract
Premature infants now have an improved chance of survival, but the impact of respiratory therapies on the brain, particularly the cerebellum, remains unclear. We examined the effects of early nasal continuous positive airway pressure (EnCPAP) ventilation and delayed (Dn) CPAP on the development of the cerebellum in prematurely delivered baboons. The baboons were delivered at 125 ± 2days of gestation and ventilated for 28 days with either EnCPAP commencing at 24 hours (n = 5) or DnCPAP commencing at 5 days (n = 5). Gestational controls (n = 4) were delivered at 153 days. Cerebella were assessed histologically, and an ontogeny study (90 days to term) was performed to establish values for key cerebellar developmental indicators. Cerebellar weight was reduced in DnCPAP but not EnCPAP animals versus controls; cerebellar/total brain weight ratio was increased in EnCPAP (p < 0.05) versus control and DnCPAP animals. There was no overt damage in the cerebella of any animals, but a microstructural alteration index based on morphological developmental parameters and microglial immunoreactivity was increased in both prematurely delivered cohorts versus controls (p < 0.001) and was higher in DnCPAP than EnCPAP animals (p < 0.05). These results indicate that respiratory regimens can influence cerebellar development and that early compared with delayed extubation to nCPAP seems to be beneficial.
Introduction
There is increasing recognition that premature birth can affect not only the developing cerebral hemispheres but also the cerebellum (1-4). Moreover, poor cerebellar development occurs in infants who also suffer supratentorial damage (3, 4). It has been suggested that there might be a trophic interaction between these structures during development that could account for this relationship in preterm infants (2). The long-term impairments in motor skills and coordination, cognition, language, and social function that occur in some premature individuals may originate in part from this adverse cerebellar development. The human cerebellum undergoes a significant growth spurt from midgestation to term followed by a 3.5-fold increase in mass from term to 9 months of age (5). After this time, it grows more slowly and in tandem with the cerebrum. During these periods of rapid growth, the cerebellum may be particularly vulnerable to environmental influences such as alterations in oxygen, nutrient, and hormone levels, all factors that can be affected in association with preterm birth.
Although neuroimaging studies have provided evidence of cerebellar injury and reduced cerebellar volume in premature infants, an animal model is required to explore the impact of preterm birth and neonatal therapies. We previously investigated the impact of neonatal therapies on the cerebral hemispheres and identified the presence of subtle brain injury and alterations to growth in the absence of potentiating factors such as hypoxia or infection in a unique baboon model of premature delivery (6-8). To date, we have not examined the cerebellum in these animals. The aim of the present study was to evaluate the influence of a specific respiratory therapy, nasal continuous positive airway pressure (nCPAP), on the extent of injury and altered development in the cerebellum. Animals were delivered at 125 ± 2 days of gestation (dg), which is equivalent to 26 to 28 weeks of gestation in humans, and maintained for 28 days in intensive care. They were randomized to receive either early or delayed extubation to nCPAP preceded by positive pressure ventilation. We examined parameters including volumetric measurements, neuronal somal size, astrocyte and oligodendrocyte development, and markers of cell proliferation and inflammation. The developmental parameters in the experimental animals were compared with those in gestational control animals delivered at 153 dg. To establish criteria for altered development in the baboon, we also carried out a parallel ontogeny study for several key developmental features of the cerebellum from 90 dg to term.
Materials and Methods
Animal studies were performed at the Southwest Foundation for Biomedical Research in San Antonio, TX. Animal husbandry, handling, and procedures were reviewed and approved to conform to the American Association for Accreditation of Laboratory Animal Care.
Ontogeny Study
An ontogeny study of key features of cerebellar development was performed to determine whether development had been disrupted or delayed in the prematurely delivered animals. Cerebella were collected from control animals at 90 (n = 1), 125 (n = 3), 140 (n = 3), and 160 (n = 2) dg and term (n = 2). A midsagittal block of vermis was processed in paraffin, and 8-μm-thick sections were used to assess growth using hematoxylin and eosin, Purkinje cell development using calbindin-immunoreactivity (IR), peak periods of cell proliferation (Ki67-IR), ontogeny of astrocytes and radial glial fibers (glial fibrillary acidic protein [GFAP-IR]), and the sequence of myelination (Black-Gold histochemistry). The DeadEnd Colorimetric TdT-mediated dUTP nick-end labeling (TUNEL) system (Promega, Madison, WI) was used according to the manufacturer's instructions to identify cell death. Immunohistochemistry was performed as described below for the experimental study.
Black-Gold Histochemistry
A 5-mm midsagittal block of vermis was collected from each cerebellum, and serial frozen sections (100 μm thick) were collected and reacted with 0.2% Black-Gold (a gift from Dr Larry Schmued, National Center for Toxicological Research/Food and Drug Administration, Jefferson, AR) for 20 to 30 minutes at 60°C. Staining was enhanced with 0.2% potassium tetrachloroaurate (Sigma, St Louis, MO) to identify myelinated axons (9).
Analysis
Hematoxylin and eosin-stained sections were assessed as described below (see quantitative analysis), to assess the total cross-sectional area, area of the inner granule layer (IGL) and white matter (WM), and the widths of the external granule layer (EGL) and molecular layer (ML). The density of astrocytes (GFAP-IR) in the deep WM was assessed qualitatively. Glial fibrillary acidic protein-IR sections were also examined to determine the distribution of Bergmann glial processes in the ML. The distribution of Black-Gold staining was assessed qualitatively at each age. Ki67-IR cell counts were performed in the deep WM as described below (see quantitative analysis). Analysis of TUNEL-positive cells was performed using the same methodology. All analyses were carried out in lobules 1 (early developing [10]) and 8 (later developing [10]) and in the deep WM.
Premature Baboon Model Treated With nCPAP
Pregnant baboon dams (Papio papio) were treated with 6 mg of intramuscular betamethasone, 48 and 24 hours before elective hysterotomy under general anesthesia. Study animals were delivered at 125 ± 2 days (67% of gestation; term ∼185 days), intubated and treated with 200 mg/kg Curosurf (Chiesi Farmaceutica S.p.A., Parma, Italy) before the initiation of ventilator support.
Respiratory Management
The respiratory management and treatment of both early CPAP (EnCPAP) and delayed (Dn) CPAP animals were initially identical and have been described previously (6). Extubation to nCPAP was attempted at either 24 hours (EnCPAP; n = 5) or 5 days (120 hours) of age (DnCPAP; n = 5). In relation to early nCPAP, we are aware that since this paradigm was devised, extubation occurs earlier than 24 hours in some neonatal intensive care units, but 24 hours is still a common time frame when administration of 1 to 2 doses of surfactant is required. All animals were maintained on the Infant Flow Generator nCPAP delivery device (ElectroMedical Equipment, Ltd, Brighton, UK) via nasal prongs and occasionally nasal mask with an initial pressure of 7 cm H2O until the conclusion of the experiment at 28 days after birth. The management of nutrition, drugs, patent ductus arteriosus, and hypotension has been described previously (11).
Physiological Data
PaO2, PaCO2, pH, FIO2, systolic, diastolic and mean arterial blood pressure (MAP), and heart rate were monitored continuously throughout the experimental period. We also examined the relationship between physiological instability and measurements of brain growth and microstructural alterations in the newborn baboons (see statistical analysis below) by calculating the “interval flux” of physiological variables as a surrogate measure of the physiological instability. We first determined the maximum and minimum values of each variable during a time interval (6 hourly time periods for the first 48 hours and daily periods thereafter); the interval flux was the difference between these values during the specified time period. We then identified the maximum flux and calculated the mean of the interval fluxes during the entire experimental time period for each animal.
Tissue Collection and Processing
At 153 dg, all prematurely delivered animals were euthanized with sodium pentobarbitone (130 mg/kg intravenously). To study gestational control brains, additional pregnant baboon dams were treated with 6 mg of intramuscular betamethasone at 123 and 124 dg and animals (n = 4) delivered by elective hysterotomy at 153 dg; animals were then killed immediately. All brains were immersed in 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). A midsagittal block of the cerebellar vermis was collected from each brain and processed to paraffin, and 10 (8 μm) sections were collected. With the exception of 1 DnCPAP animal, which was excluded due to suboptimal preservation of the cerebellum, all analyses were performed on all nCPAP and control animals.
Morphological Analysis
All measurements were made on coded slides. Areas and widths were assessed using a digitizing program (Sigma Scan Pro v4; SPSS Science, Chicago, IL) and counts performed using an image analysis system (Image Pro Plus v4.1; Media Cybernetics, Bethesda, MD).
Histology
Two sections from each block were stained with hematoxylin and eosin and qualitatively assessed for gross morphological changes, including the presence of subarachnoid or intraparenchymal hemorrhages, infarcts, axonal injury, gliosis, or other lesions.
Immunohistochemistry
Purkinje cells were identified using rabbit anti-rat calbindin (1:1000; Swant, Bellinzona, Switzerland); rabbit anti-cow GFAP (1:500; DakoCytomation, Glostrup, Denmark) was used to identify astrocytes; mouse anti-chicken myelin basic protein (MBP; 1:100; Chemicon, International, Inc, Temecula, CA) was used to assess the extent of myelination; rabbit anti-ionized calcium-binding adapter molecule 1 (Iba1; 1:100; Wako Chemicals, Osaka, Japan) was used to identify activated macrophages/microglia; and mouse anti-human Ki67 clone MIB-1 (1:100; DakoCytomation) was used to identify proliferating cells. The avidin-biotin-peroxidase complex (Vector Laboratories, Burlingame, CA) (7) was used to visualize IR. Antigen retrieval was performed for MBP-IR, Iba1-IR, and Ki67-IR using 0.01 M sodium citrate buffer (pH 6.0) and microwaved for 10 minutes at boiling temperature; sections were cooled in citrate buffer for 45 minutes before commencing immunohistochemistry. Control and experimental material was stained simultaneously to avoid procedural variation. Control experiments were performed by omitting the primary antibodies, whereupon staining failed to occur.
Colocalization Studies
Sections were prepared for immunohistochemistry as described above and incubated in mouse anti-human Ki67 with either rabbit anti-cow GFAP or rabbit anti-Iba1 overnight at 4°C. Sections were washed and incubated for 60 minutes at room temperature in the dark with secondary antibodies coupled to either of the following: Alexa goat anti-rabbit 594 (red fluorescence; Invitrogen Corporation, Carlsbad, CA) and Alexa goat anti-mouse 488 (green fluorescence; Invitrogen) for Ki67/GFAP colocalization; goat anti-rabbit 488 and donkey anti-mouse 594 for Ki67/Iba1 colocalization; and goat anti-rabbit 488 and donkey anti-mouse 594 for Ki67/Olig2 (rabbit anti-Olig-2 polyclonal antibody; 1:200; Chemicon) colocalization, (1:500; Molecular Probes, Eugene, OR). After washing and mounting, sections were examined with a Zeiss Axioskop Fluorescence Microscope (Zeiss, Oberkochen, Germany).
Quantitative Analysis
Measurements were made in lobules 1 and 8 where appropriate; cells/layers were randomly sampled in 2 sections per animal for each analysis. For all parameters, means were calculated for each animal and then a group mean was determined.
Area Measurements
Areas of the total cross-section, the WM and the IGL were determined in hematoxylin and eosin-stained sections through the vermis.
Layer Widths
The widths of the EGL and ML were measured at 50 sites per section. Somal areas of 25 cells from randomly selected regions within the IGL were measured (50 cells/cerebellum; 1,300×).
Purkinje Cell Somal Area and Areal Density
In calbindin-IR sections, somal areas were measured in 50 Purkinje cells from randomly selected regions (100 cells/cerebellum; 1,300×). To estimate the Purkinje cell density, cells with a clearly defined nucleus were counted in 10 randomly selected lengths of transected Purkinje line (5 lengths per section), and the results were expressed as Purkinje cells per unit surface area (in millimeter squared) of Purkinje cell layer (12).
Counts of Proliferating Cells
In 5 regions of the EGL (75-μm length of EGL; 600×), all Ki67-IR cells were counted and the number expressed as the proportion of total cells in the EGL. To estimate the number of proliferating Bergmann glial cells located just beneath the layer of Purkinje cell perikarya (13), 5 regions (150-μm length of Purkinje cell layer; 600×) were sampled; counts were expressed as the mean number of Ki67-IR Bergmann glia/mm2 of Purkinje cell layer. In the deep WM, 5 regions were sampled (600×), and the mean number of Ki67-IR cells (in cells per millimeter squared) was determined.
MBP-IR Oligodendrocytes
Myelin basic protein-IR oligodendrocytes (in cells per millimeter squared) were counted in the WM in 10 regions (0.023 mm2; 600×).
GFAP-IR Optical Density
Average optical densities of astrocytes in the deep WM were determined using an image analysis system (14) in 5 sampled regions (0.07 mm2; 600×); the high density of astrocytes precluded cell counting.
Iba1-IR Macrophages/Microglia Counts
Ionized calcium-binding adapter molecule 1-IR cells were densely distributed in the WM in all animals making counts in this region impractical. The numbers of Iba1-IR cells/mm2 in the EGL, ML/Purkinje cell region and IGL were assessed in 5 sampled regions in each EGL (0.007 mm2; ×600), ML/Purkinje cell region (0.012 mm2; ×600) and IGL (0.08 mm2; ×600) per section.
Semiquantitative Analysis
Myelination (MBP-IR) was most advanced in gestational control brains, and this was given a score of 3. The projection of myelinated fibers into the IGL in the 2 nCPAP groups was scored against this standard (0-no myelination; 1-a few myelinated fibers; 2-bundles of myelinated fibers present; and 3-similar extent of myelination to gestational controls). Qualitative assessment was made of the Bergmann glial morphology in GFAP-IR sections.
Microstructural Alterations Index
The cerebellar microstructural alteration index was devised to assess overall damage and altered development for each animal over a number of parameters. Development parameters included the volumes of WM and IGL, width of the ML, IGL and Purkinje cell somal areas (data was ranked from 13 for lowest values to 1 for highest), and density of MBP-IR oligodendrocytes (13 for lowest to 1 for highest). Damage parameters were the densities of invading microglia/macrophages (Iba1-IR cells) in the Purkinje cell/ML region and IGL (13 highest density to 1 for lowest). Scores were added, and a mean was calculated for each group, with a higher score indicating more microstructural alterations. We acknowledge that equal weighting has been given to all parameters in constructing this index. At this stage of gestation, it is difficult to predict which parameters of development might be the most relevant predictors of long-lasting deficits.
Statistical Analysis
Linear regression analysis was carried out to determine if there were correlations between the cerebellar microstructural alterations index and forebrain damage index calculated previously (6), the physiological variables previously reported (6), and quantitative parameters. A probability of p < 0.05 was considered significant. For all histological parameters, the statistical significance of differences between control, EnCPAP, and DnCPAP groups was tested using a 1-way analysis of variance with post hoc analysis (Tukey test); a probability of p < 0.05 was considered to be significant. Results are expressed as mean ± SEM for weights and mean of means ± SEM for histological parameters.
Results
Ontogeny of the Baboon Cerebellum
Growth Rate
At 90 dg, all 10 lobules are present; the cerebellum undergoes rapid growth between 90 and 160 dg; it increases in weight ∼7-fold between 90 and 125 dg and 3.3-fold between 125 and 160 dg, as a consequence of cell proliferation, growth of processes, and myelination. After this time, growth begins to slow (Table 1).
Cellular Characteristics and Layers
The cross-sectional area of the vermis, WM, and IGL increased commensurately with gestational age. The thickness of the EGL seemed to peak at approximately ∼125 dg in lobule 1 and between 125 and 140 dg in lobule 8. At 90 dg, Purkinje cells were multilayered (2-3 cells per layer); they were predominantly organized into a single layer from 125 dg onward. Purkinje cell somal areas increased steadily until 160 dg, after which there was a dramatic increase during the 3 weeks until term, particularly in lobule 1. The areal density of Purkinje cells decreased markedly between 125 and 140 dg, with increasing dendritic arborization. The width of the ML in both lobules increased steadily from 90 dg until term. The somal area of IGL cells in both lobules increased by approximately 28% between 125 and 140 dg, with small increases thereafter (Table 1).
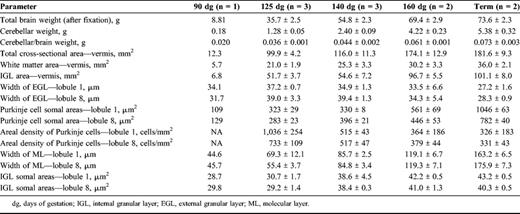
Myelinated fibers were evident in the deep cerebellar WM at 125 dg, and a few fibers were seen in the deep regions of the developing folia (Figs. 1A, D). By 140 dg, myelination had increased in all folia with intense staining in the deep WM. Some myelinated axons were evident in the IGL (Figs. 1B, E). At 160 dg, myelination had extended further into the IGL but had not yet reached the Purkinje cell layer (Figs. 1C, F).
Ontogeny of myelination in the cerebellum by Black-Gold histochemistry. (A) Myelinated fibers are confined to the white matter at 125 days of gestation (dg). (B) At 140 dg, they begin to enter the IGL. (C) Myelinated fibers are abundant in the IGL at 160 dg. (D-F) Boxed areas are shown at higher magnification. (G-I) Glial fibrillary acidic protein (GFAP)-immunoreactivity in Bergmann glial fibers (arrows) in the IGL with increasing gestational age: 125 dg (G), 140 dg (H), and 160 dg (I). Arrowheads indicate migrating granule cells. EGL, external granule layer; IGL, inner granule layer; ML, molecular layer; PCL, Purkinje cell layer, WM, white matter. Scale bars = (A-C) 2.5 mm; (D-F) 150 μm; (G-I) 50 μm.
At 125 dg, GFAP-IR astrocytes (not shown) densely populated the deep WM; they were present at a lower density throughout the IGL (Fig. 1G). Bergmann glia were GFAP-IR at 125 dg (Fig. 1G), 140 dg (Fig. 1H), and 160 dg (Fig. 1I) and persisted to term. Cell proliferation in the deep cerebellar WM was high at 90 dg, but it was not possible to determine if this was the peak because earlier time points were not assessed (Fig. 2A). The peak period of cell death (TUNEL-positive cells) in the cerebellar WM seemed to be 125 dg; numbers declined steeply to 140 dg and then more gradually to term (Fig. 2B). It is possible that TUNEL-positive cells included both astrocytes and oligodendrocytes.
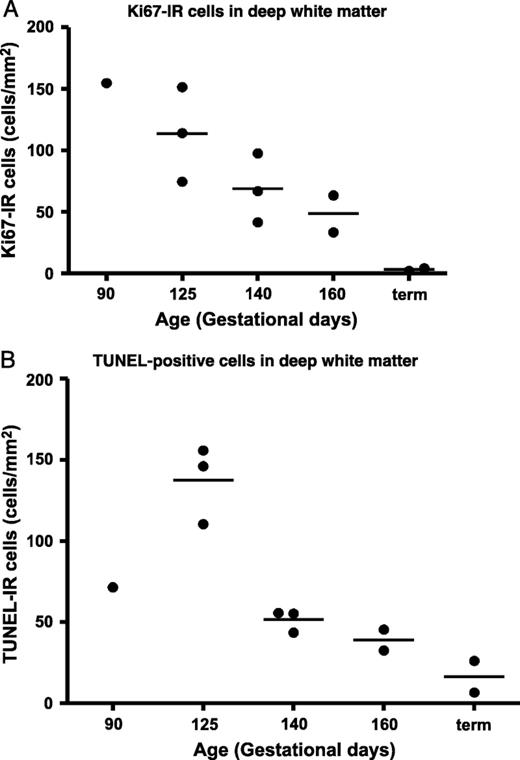
(A) Ontogeny of Ki-67-immunoreactive (IR) cells in the deep cerebellar white matter showing a decrease from 90 days of gestation (dg) to term (in cells per millimeter squared). (B) TdT-mediated dUTP nick-end labeling (TUNEL)-positive cells in the deep cerebellar white matter; the peak period of cell death seems to be at ∼125 dg.
Effects of Respiratory Regimes
Details regarding the clinical and respiratory characteristics of the animals have been described previously (6). Briefly, the PaCO2 minimum and maximum, the FiO2 maximum, and the MAP minimum values were increased (p < 0.05) in DnCPAP compared with EnCPAP. There was no difference in pH, PaO2, or heart rate maximum and minimum values between groups. The FIO2 maximum and mean interval fluxes as well as the MAP mean interval flux values were increased (p < 0.05) in DnCPAP compared with EnCPAP animals. There was no difference between DnCPAP and EnCPAP groups in relation to the maximum or mean interval flux values for PaO2, pH, or heart rate.
Body and Brain Measurements
Body and brain weights were reduced in both nCPAP groups of animals compared with age-matched gestational controls (p < 0.05; Table 2). The mean cerebellar weight and total cross-sectional area (Figs. 3A-C) were reduced in DnCPAP (p < 0.01) but not in EnCPAP (p > 0.05) animals compared with controls. The cerebellar-to-brain weight ratio was increased (p < 0.05) in EnCPAP compared with control and DnCPAP animals. No infarcts, hemorrhages, or overt damage were observed in the cerebellar vermis of any animal.
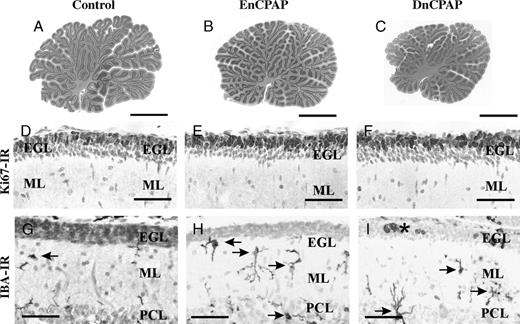
(A-C) The cross-sectional areas in hematoxylin and eosin stain of the vermis are reduced in delayed nasal continuous positive airway pressure (DnCPAP) animals (C) compared with controls (A) and early CPAP (EnCPAP) (B) animals. (D-F) The numbers of Ki67-immunoreactive (IR) proliferating cells in lobule 8 of the external granule layer (EGL) are not different among control (D), EnCPAP (E), and DnCPAP (F) animals. (G-I) Ionized calcium-binding adapter molecule 1 (Iba1)-IR macrophage/microglial cells (arrows) are increased in the molecular layer (ML) and Purkinje cell layer (PCL) in EnCPAP (H) and DnCPAP (I) animals compared with controls (G). Amoeboid cells in the EGL are indicated by an asterisk. Scale bars = (A-C) 3 mm; (D-F) 20μm; (G-I) 15 μm.
Effects of Respiratory Regimes on Body and Brain Weights and Cerebellar Volume Measurements
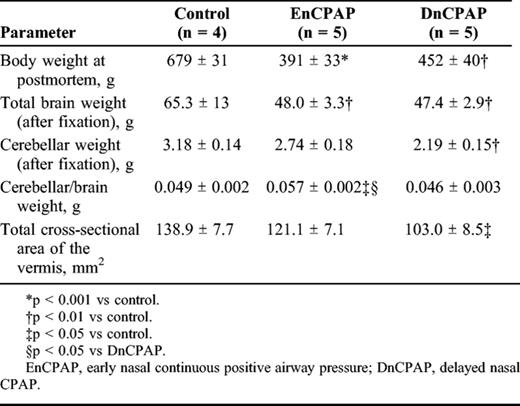
Effects of Respiratory Regimes on Body and Brain Weights and Cerebellar Volume Measurements
External Granule Cell Layer
No differences (p > 0.05) were observed between groups in the width of the EGL in lobule 1 (control, 31.4 ± 1.5 µm; EnCPAP, 29.6 ± 1.9 µm; DnCPAP, 30.7 ± 0.9 µm) or lobule 8 (control, 34 ± 2 µm; EnCPAP, 33 ± 1 µm; DnCPAP, 34 ± 3 µm), nor in the number (lobule 1: control, 633 ± 34 µm; EnCPAP, 607 ± 49 µm; DnCPAP 580 ± 18 µm; lobule 8: control, 721 ± 15 µm; EnCPAP, 661 ± 26 µm; DnCPAP, 687 ± 18 µm or proportion (data not shown) of Ki67-IR cells/mm EGL between groups. This is illustrated for lobule 8 in Figures 3D-F. Ionized calcium-binding adapter molecule 1-immunoreactivity cells were located throughout the EGL, ML (Figs. 3G-I), and IGL in all groups. Ionized calcium-binding adapter molecule 1-immunoreactivity macrophage/microglial cells were predominantly ramified; however, amoeboid cells were observed in the EGL in 1 of 4 DnCPAP (Fig. 3I) and 1 of 5 EnCPAP animals but not in any of the control animals. Aggregates of Iba1-IR cells predominantly associated with blood vessels were occasionally observed in the EGL, ML (Fig. 4C), and IGL of DnCPAP (2/4) and EnCPAP (3/5) animals.
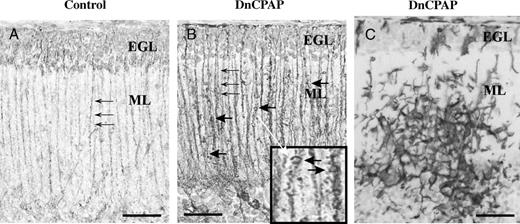
Glial cell immunohistochemistry. (A, B) Glial fibrillary acidic protein-immunoreactive Bergmann glial fibers (small arrows) in control (A) and delayed nasal continuous positive airway pressure (DnCPAP) (B) animals. (B) Excrescences (large arrows) along fibers and at higher power (inset indicated by white arrow) are seen. (C) An aggregate of ionized calcium-binding adapter molecule 1-immunoreactive cells in the ML layer in a DnCPAP animal. EGL, external granule layer; ML, molecular layer. Scale bars = (A, B) 30 μm; (B, inset) 75 μm; (C) 30 μm.
Purkinje Cells and ML
There was a reduction in the width of the ML (p < 0.05) in DnCPAP animals compared with controls in lobule 1; no differences (p > 0.05) were observed between groups in lobule 8 (Table 3). There was no difference (p > 0.05) in Purkinje cell somal areas nor in the density of Purkinje cells in either lobule 1 or 8 between groups. There was an increase in the number of Iba1-IR macrophages/microglial cells in the Purkinje cell/ML in lobule 1 in EnCPAP (p < 0.05) and DnCPAP (p < 0.01) animals compared with controls (Figs. 3G-I). There was no difference between groups in the number of Ki67-IR Bergmann glia in lobule 1 or 8. There seemed to be morphological alterations in GFAP-IR Bergmann glial fibers, which were more prominent in DnCPAP than in EnCPAP animals and showed tapering toward the pial surface with dilations and excrescences in 4 of 4 Dn (Fig. 4B) and 1 of 5 EnCPAP animals; these morphological alterations were not observed in controls (Fig. 4A).
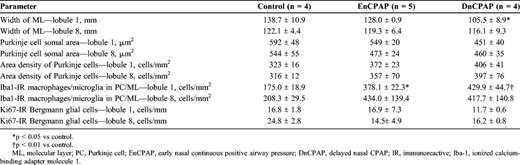
Inner Granule Cell Layer
The area of the IGL was reduced (p < 0.05) in DnCPAP animals compared with controls (Table 4). There was a decrease in the somal area of granule cells in the IGL in lobules 1 and 8 in DnCPAP compared with control (p < 0.01) and EnCPAP (p < 0.05) animals (Figs. 5A-C). There was an increase in the number of Iba1-IR macrophages/microglial cells in the IGL in lobule 1 of EnCPAP (p < 0.01) and DnCPAP (p < 0.01) animals compared with controls, with a similar trend in lobule 8.
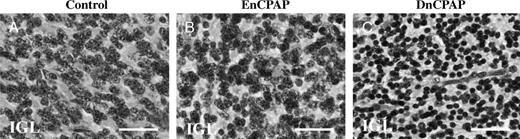
The somal areas of internal granule layer (IGL) neurons are reduced in delayed nasal CPAP (DnCPAP) (C) compared with control (A) and early nasal (EnCPAP) (B) animals. Scale bars = A-C = 28 μm.
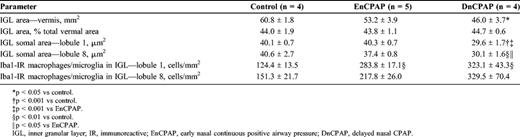
White Matter
The cross-sectional area of the cerebellar WM did not differ between groups (control, 24 ± 1 mm2; EnCPAP, 21 ± 1 mm2; DnCPAP, 22 ± 1 mm2), neither was there a difference when expressed as a percentage of total vermal area. There was no difference between groups in the number of MBP-IR oligodendrocytes in the deep WM in either lobule 1 (control, 638 ± 33 cells/mm2; EnCPAP, 658 ± 33 cells/mm2; DnCPAP, 730 ± 97 cells/mm2) or 8 (control, 844 ± 103 cells/mm2; EnCPAP, 919 ± 118 cells/mm2; DnCPAP, 930 ± 160 cells/mm2). Ki67-IR cells were present throughout the deep WM, with an increase (p < 0.05) in DnCPAP but not in EnCPAP animals compared with controls (control, 40 ± 16 cells/mm2; EnCPAP, 98 ± 33 cells/mm2; DnCPAP, 166 ± 16 cells/mm2; Figs. 6A-C). Colocalization of Ki67-IR with Olig2-IR, Iba1-IR, or GFAP-IR was carried out to determine the phenotype of proliferating cells. Most cells were oligodendrocytes, with very few astrocytes or microglia observed. Colocalization of Ki67 and Olig2-IR is illustrated in Figures 6D-F in a DnCPAP animal. There was no difference between groups in the optical density of GFAP-IR astrocytes in the deep WM (control, 0.180 ± 0.008; EnCPAP, 0.197 ± 0.022; DnCPAP, 0.181 ± 0.007).
(A-C) There is an increase in the number of Ki67-immunoreactive (IR) cells in the deep white matter (DWM) in a delayed nasal continuous positive airway pressure (DnCPAP) animal (C) compared with control (A) and early nasal CPAP (EnCPAP) (B) animals. (D-F) Colocalization of Olig2-IR and Ki67-IR in the deep cerebellar white matter in a DnCPAP animal. D, Olig2-IR; E, Ki67-IR; F, merge. Scale bars = (A-F) 30 μm.
Cerebellar Microstructural Alteration Index
There was an increase in this index for both EnCPAP (90 ± 4; p < 0.05) and DnCPAP (114 ± 10; p < 0.001) animals compared with controls (66 ± 1) and an increase in DnCPAP compared with EnCPAP animals (p < 0.05).
Relationship Between Indices of Forebrain Damage and Cerebellar Microstructural Alterations
Across all cohorts, there was a positive correlation between the indices of forebrain damage and cerebellar microstructural alterations (r2 = 0.52; p < 0.005; Fig. 7).
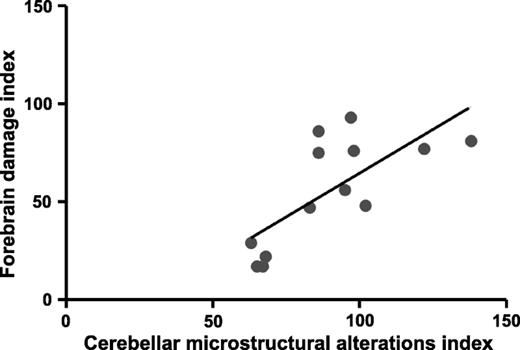
Regression analysis for all animals. There is a positive correlation between the forebrain damage (data from previous study 6) and cerebellar microstructural indices (B; r2= 0.52; p < 0.005). EnCPAP, early nasal continuous positive airway pressure; DnCPAP, delayed nasal continuous positive airway pressure; DWM, deep white matter.
Relationship Between Physiological Parameters and Cerebellar Growth and Microstructural Alterations
Negative correlations were observed between the mean interval flux in FIO2 and 1) the width of the ML in lobule 1 (r2 = 0.50; p < 0.03); 2) IGL somal areas in lobules 1 (r2 = 0.82; p < 0.007) and 8 (r2 = 0.62; p < 0.02); and 3) the Purkinje cell somal area in lobule 1 (r2 = 0.48; p < 0.04). Negative correlations were also observed between maximum flux in MAP and the total WM volume (r2 = 0.72; p < 0.009) and maximum flux in PaCO2 and the ML width in lobule 8 (r2 = 0.48; p < 0.04). These correlations indicate that greater fluxes in blood gases and blood pressure result in poorer growth of cells and processes.
Discussion
The major finding of this study is that baboons delivered prematurely at 67% of gestation and nursed in an intensive care setting for ∼28 days exhibited significant adverse effects on multiple aspects of cerebellar growth and structure; overall, these effects are more pronounced when weaning from positive pressure ventilation to nCPAP is delayed from 1 to 5 days. This finding is in accord with our previous study on the same cohort of animals in which we reported that delayed weaning to nCPAP increased adverse effects on brain growth in general and on the incidence of cerebral abnormalities (6).
To inform the present experimental study, we initially assessed the ontogeny of key features of cerebellar development from 90 dg through term because this information was not previously available for the baboon. Consistent with other primate data (5), we confirmed that the cerebellum undergoes rapid growth during later gestation (125-160 dg), the time frame of the present study; this corresponds to 27 to 40 weeks of human gestation.
In the deep WM, the density of proliferating cells was elevated from 90 to 125 dg. Cell death was also apparent in this region, with the peak occurring at 125 dg, which coincides with the onset of myelination. In rodents, it has been reported that 50% of cells in the developing WM die between birth and postnatal day 7 and that 50% to 70% of these cells are astrocytes (15). The authors proposed that astrocytes might play a key role in supporting the development of afferent and efferent tracts in the developing cerebellum before the differentiation of oligodendrocytes and the onset of myelination (15). Although we did not identify the phenotype of the proliferating or dying cells in the baboon, our findings are consistent with this hypothesis.
Premature Delivery Alters Cerebellar Growth
The lower cerebellar weight in DnCPAP animals compared with controls was reflected in a reduction in the cross-sectional area of the vermis. Extrapolating from the ontogeny data, it is evident that growth since delivery had been markedly slowed in these animals. Our findings are consistent with reports in humans in which the rapid growth occurring in the cerebellum in late gestation is impeded by preterm birth and associated with brain injury (2). Although there was a trend for the growth of the cerebellum to be compromised with EnCPAP, the higher ratio of cerebellar-to-brain weight compared with both controls and DnCPAP suggests that growth is somewhat spared with this regimen.
Differential Effect of EnCPAP and DnCPAP on the Cerebellar Microstructure
In the prematurely delivered cohorts, we found that the growth of granule cells was affected by delayed but not early weaning to nCPAP in both lobules 1 and 8. Comparison with data from the ontogeny study indicated that the somal size had not increased since the time of delivery, presumably contributing to the smaller IGL area in the vermis. Because somal size is correlated with the extent of dendritic (16) and axonal arborization (17), a reduction in somal size might have marked effects on connectivity and have functional implications in later life. In assessing whether granule cell proliferation was affected in DnCPAP animals, we found that the EGL was not thinner than that of controls, nor was there any evidence of a reduction in the number or proportion of proliferating to postmitotic granule cells within a unit area of EGL. Because cerebellar weight and cross-sectional area were reduced, however, the total areal volume of the EGL is also likely to be reduced, resulting in an overall reduction in absolute numbers of proliferating neurons. There was no evidence of increased cell death within the EGL.
Further evidence of a differential effect of the length of ventilation and weaning to CPAP was demonstrated with alterations in the morphology of Bergmann glial fibers in 100% of DnCPAP animals but in only 20% of EnCPAP animals; the basis for this abnormal morphology is unknown. There were no quantifiable differences in the density of Bergmann glial fibers in either cohort compared with controls. It has been suggested that Bergmann glia can engulf and phagocytose apoptotic granule cells (13), but we did not find evidence for this in our study. Because Bergmann glia are involved in neuronal migration and neuron-glia signaling events, an abnormal morphology of these cells could affect granule cell migration and contribute to the reduction in IGL width.
We found no specific evidence that Purkinje cells were affected in either DnCPAP or EnCPAP animals; no cell death was observed, and when compared with controls, there was no difference in somal size, areal density, or width of the ML, a major component of which are the dendritic trees of Purkinje cells. With the few exceptions already noted, there was no marked differential effect of the specific respiratory regimen on lobules 1 and 8.
The ontogeny study demonstrated that myelination was in its early stages at 125 dg; this is a time at which preoligodendrocytes are reportedly vulnerable to adverse conditions including hypoxia (18). We therefore predicted that there might be a demonstrable effect on myelination in the cerebellum of prematurely delivered animals. In fact, neither delayed nor early CPAP seemed to alter either the proportion of WM in the vermis or the density of MBP-IR cells in the deep WM compared with controls. Cell proliferation in the deep WM was, however, actually elevated in DnCPAP animals; most of these cells were Olig2-positive, indicating an oligodendrocyte lineage. We speculate that this might reflect early growth arrest followed by rebound proliferation as inhibitory influences due to delayed weaning to nCPAP (described below) are removed.
Mechanisms Underlying the Effects of Respiratory Intervention on the Cerebellum
The question arises as to why premature delivery causes mild cerebellar damage and delayed growth and, further, why delaying extubation to nCPAP might exacerbate this response. In the baboon model, as in the preterm infant, attempts were made to reduce hypoxic episodes, but despite this, there was variation in respiratory stability in both cohorts. The DnCPAP group experienced more apnea requiring reintubation (11); they also exhibited higher fluxes in the fraction of inspired oxygen required to maintain appropriate blood gases. Regression analysis revealed that animals with higher fluxes had poorer cerebellar growth overall, particularly in relation to inner granule and Purkinje cell development. Studies of hypoxic insults to the ovine brain at a similar stage of cerebellar development have demonstrated reductions in cerebellar weight and dendritic growth (12). There are many other potential contributing factors to the risk of altered cerebellar growth and injury in addition to hypoxemia, including poor nutrition, infection, exposure to drugs, and stress. Stress has been shown to have specific effects on the developing cerebellum in rats, including reductions in granule cell size and interneuronal connectivity (19). Although every effort was made to provide infant baboons with adequate nutrition (20), the slow body and cerebellar weight gain might indicate a reduced supply of nutrients to the cerebellum which could have contributed to the observed alterations. The DnCPAP group clearly experienced an extended time required on positive pressure ventilation, and this may have activated inflammatory cascades. In relation to inflammation, macrophages/microglia in both cohorts were elevated compared with controls; this elevation was more pronounced in the DnCPAP animals in both the molecular and IGLs. Consistent with this, serum cytokines levels have also been shown to be elevated in the DnCPAP animals (11). Because there was little evidence of cell death at the time of this study, we are unable to determine the underlying etiology of the increased number of inflammatory cells.
Relationship Between Cerebral Injury and Cerebellar Alterations
Taking all animals into consideration, there was a positive correlation between the indices of cerebral injury from our previous study (6) and altered cerebellar microstructure, indicating that with increasing forebrain injury, there is a concomitant increase in cerebellar microstructural alterations.
It should be noted that although astrogliosis was an important finding in the forebrain WM of nCPAP animals, the high density of astrocytes in the developing cerebellum in all animals precluded us from carrying out astrocyte density counts. Although there was no difference in optical density for GFAP-IR, it is possible that any alterations might have been below the level of detection for this technique.
In summary, baboons delivered prematurely at 67% of gestation and nursed for 28 days exhibit significant adverse effects on aspects of cerebellar growth and microstructure; overall, these effects are more pronounced when intubation to nCPAP is delayed from 1 to 5 days. These findings demonstrate that respiratory regimens in neonatal intensive care can indeed influence cerebellar development and hence possibly functional outcomes in the long term. It has already been established that premature infants with impaired cerebellar development are likely to have more cognitive and motor deficits in preschool years (3) and in adolescence (1).
Acknowledgments
We are grateful to Vicki Winter for organizing the shipment of brains to Prof Rees' laboratory and to the personnel at the Bronchopulmonary Dysplasia Resource Center, San Antonio, TX, particularly the animal husbandry group and neonatal intensive care unit technicians. Sarah Cain provided valuable assistance with histology.
References
Author notes
This study was supported by Grant R01 HL074942-OIAI from the National Institutes of Health.
Online-only color figures are available at http://www.jneuropath.com.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}