Persistence of Seed Dispersal in Agroecosystems: Effects of Landscape Modification and Intensive Soil Management Practices in Avian Frugivores, Frugivory and Seed Deposition in Olive Croplands
 Pedro J. Rey1,2*
Pedro J. Rey1,2*  Francisco M. Camacho2
Francisco M. Camacho2  Rubén Tarifa2,3
Rubén Tarifa2,3  Carlos Martínez-Núñez2
Carlos Martínez-Núñez2  Teresa Salido1,2
Teresa Salido1,2  Antonio J. Pérez2
Antonio J. Pérez2  Daniel García4
Daniel García4- 1Instituto Interuniversitario de Investigación del Sistema Tierra de Andalucía, Universidad de Jaén, Jaen, Spain
- 2Departamento de Biología Animal, Biología Vegetal y Ecología, Universidad de Jaén, Jaen, Spain
- 3Estación Experimental de Zonas Áridas de Almería, EEZA, CSIC, Almería, Spain
- 4Departamento de Biología de Organismos y Sistemas e Instituto Mixto de Investigación en Biodiversidad (Univ. Oviedo-CSIC-Princ. Asturias), Universidad de Oviedo, Oviedo, Spain
Farming impacts animal-mediated seed dispersal through mechanisms operating on at least two spatial scales. First, at the landscape scale, through habitat loss and land conversion to agriculture/livestock grazing, and second, at the farm scale, via a local intensification of agricultural practices. These two scales of farming impact seed dispersal function but have rarely been integrated. In particular, studies evaluating the effect of agriculture on the seed dispersal function of frugivorous birds in Mediterranean ecosystems are lacking. This study evaluates the role of landscape transformation, from fruit-rich woodland habitats to olive grove landscapes, together with local intensive practices of soil management on the persistence of the seed dispersal function for Mediterranean fleshy-fruited plants in olive landscapes of south Spain. We used bird censuses, mist-nets, and seed traps to characterize avian frugivore assemblages, frugivory, and seed deposition in the seminatural woodland habitat (SNWH) patches and olive fields of 40 olives farms spanning 20 localities distributed across the whole range of olive cultivation in Andalusia (southern Spain). We found that despite the remarkable dispersal function of olive grove landscapes, avian frugivore abundance and diversity, frugivory, and seed arrival decreased in olive fields compared to SNWH patches. Likewise, SNWH cover loss and/or olive growing expansion decreased avian frugivory and seed arrival. Interestingly, the habitat effects in the olive farms often depended on the landscape context. In particular, less diverse fruit-eating bird assemblages pooled in SNWH patches as olive grove cover increased or SNWH decreased in the landscape, while remaining relatively invariant in the olive fields. Finally, compared to conventional intensive agriculture, low-intensity management increased frugivory and seed deposition. We conclude that olive fields are less permeable to frugivores than expected due to the agroforest-like nature of these landscapes and that the presence of SNWH patches is crucial for the maintenance of frugivory and seed dispersal in agricultural landscapes. These results evidence that woodland habitat loss by olive expansion and intensive practices seriously threaten the dispersal service in olive-dominated landscapes. Maintenance, restoration, and promotion of woodland patches should be prioritized for the conservation of seed dispersal service and for enhancing the functional connectivity in human-shaped olive landscapes.
Introduction
Seed dispersal by frugivores is an important ecosystem service that promotes spontaneous natural regeneration of vegetation and contributes to shaping natural plant communities and their dynamics in many regions (Herrera, 1985; Lázaro et al., 2005; Tomback, 2016). In particular, the seed dispersal of woody plants by frugivorous vertebrates is a major driver of the dynamics of tropical forests and Mediterranean woodlands (e.g., Herrera, 1995; Muller-Landau and Hardesty, 2005) where frugivores act as mobile links for connecting plant populations across landscape patches (Henry et al., 2007; Pérez-Méndez et al., 2017; Parejo-Farnés et al., 2020). In current real-world (human-shaped) landscapes, ensuring the persistence of seed dispersal by frugivorous vertebrates is crucial for maintaining connected plant metapopulations and metacommunities across remnant landscape patches (Mueller et al., 2014; González-Varo et al., 2017), and for promoting plant population recovery through rescue effects (García et al., 2010).
Farming is a major driver of global change and biodiversity crisis (Tilman et al., 2001; Foley et al., 2005, 2011), giving rise to the predominance of human-shaped, biologically homogenized landscapes in many regions (Green et al., 2005; Tscharntke et al., 2005, 2012; Arroyo-Rodríguez et al., 2020). Farming has also affected the ecosystem functions and the delivery of biodiversity-based ecosystem services (Letourneau et al., 2011; Gurr et al., 2017; Landis, 2017), among which seed dispersal by frugivores is not an exception. Conceptually, farming impact on animal seed dispersal is expected to operate through mechanisms, acting at least at two spatial scales (Martin et al., 2019). First, at the landscape scale, land conversion to agriculture and/or livestock grazing provokes the direct loss of natural habitats, decreasing the population sizes of frugivores and fleshy-fruited plants, and increasing isolation among remnant habitat patches, This causes plant metapopulations to collapse as it restricts animal mobility and alters seed deposition patterns. Second, at the local (farm) scale, intensification of farming practices makes lands dedicated to intensive agriculture or livestock production qualitatively inhospitable or scarcely permeable (because of pesticide application and/or drastic modification of habitat conditions and food availability) for many frugivores compared to low-intensively managed farmlands, impeding seed arrival. Reduced levels of frugivory at these two scales are also expected as fruit-bearing plants become scarce in the landscape and in the areas dedicated to crop yield. Although some of these effects have been described in both temperate and tropical systems (e.g., Lozada et al., 2007; Pejchar et al., 2008; Martínez and García, 2017), they have not been considered together (in an integrative way), which would provide a comprehensive view of the impact of farming on the persistence of animal-mediated seed dispersal. Importantly, considering the operating mechanisms at these two scales together enables us to directly associate patterns of seed dispersal decay with specific human actions (expansion of croplands, destruction or substantial modification of habitat features, intensification of agricultural practices), which allows researchers to formulate appropriate corrective measures.
The impact of farming on seed dispersal by frugivores may be expected to differ with the type of farmland, depending on how suitable the agroecosystem is for provisioning appropriate habitat structure and food resources for frugivores (Lozada et al., 2007). Namely, more structural protection is expected in woody permanent croplands than in arable lands, and more food is expected in fruit-provisioning woody croplands compared to other woody croplands. This means that fruit-provisioning woody croplands are potentially permeable to different frugivorous animals and could ease the persistence of seed dispersal function. Some of these fruit-provisioning woody croplands can be managed according to biodiversity-friendly agroforestry/agroecology principles to preserve biodiversity (including frugivorous and insectivorous vertebrates) and ecosystem services without incurring necessarily in a loss in fruit yield. Examples exist, mainly in tropical agroforest systems such as shade coffee and cocoa plantations (Lozada et al., 2007; Clough et al., 2011; De Beenhouwer et al., 2013; Maas et al., 2013; Sánchez-Clavijo et al., 2020; Araújo-Santos et al., 2021), but there are also some studies supporting the application of similar principles in temperate apple orchards (García et al., 2018), semiarid nopal agroecosystems (Mellink et al., 2016), or Mediterranean olive groves (Rey, 2011).
The olive agroecosystem is currently the most important woody crop in Europe in socio-economic and cultural terms, as well as by the extension of its cultivated area (Loumou and Giourga, 2003; European Commission, 2012). Vast areas of the Mediterranean lowlands have been progressively occupied through centuries by olive groves, rendering highly variegated natural landscapes into an olive monoculture (Weissteiner et al., 2011). Olive cropland has expanded at the expense of Mediterranean native forest, woodlands, and scrublands (seminatural woodland habitats, SNWH hereafter), particularly rich in fleshy-fruited plant species (Herrera, 1984a; Jordano, 1984) and where avian seed dispersal by birds represents a major driver of woody vegetation recovery and the long-term dynamics of ecological communities (Rey and Alcántara, 2000, 2014; Perea et al., 2021). However, unlike some notable information on avian frugivore abundance and diversity (Rey, 1993, 1995, 2011; Morgado et al., 2021), data on frugivory and seed dispersal in olive grove landscapes is anecdotal, restricted to fruit consumption during winter by the most abundant frugivores, or taken from a single or a small number of localities (Jordano and Herrera, 1981; Rey, 1992; Blanco et al., 1994; Rey et al., 1996; Rey and Valera, 1999; Delibes et al., 2012; Perea and Gutiérrez-Galán, 2016).
Besides the progressive landscape transformation, olive cultivation practices have experienced a continuous process of local (farm) intensification that has happened in two different non-mutually exclusive ways (Infante-Amate et al., 2016). A long-lasting and widespread form of olive farming intensification is based on the elimination of ground herb covers. This is typically done by using pre and post-emergence herbicides or tillage, usually accompanied by the use of other pesticides (insecticides) and fertilizers, regardless of whether the olive field is irrigated or rainfed. Ground herb cover (composed of so-called weeds) is considered to compete with the olive tree for soil nutrients and water in this form of intensive farming. However, the persistent removal of the herb cover has raised many concerns in terms of sustainability and the environment as it leaves soils persistently bare and increases soil erosion (Gómez et al., 2014), and impacts ecosystem function and services. A second, more recent, form of intensification is based on increasing the density of the planted trees. This form of intensification has raised environmental concerns, especially from the recent advent of the superintensive hedge-like plantations, because it involves important structural changes and increased input demand that affects biodiversity and sustainability. Moreover, the accelerated expansion of hedge-type plantations demands new agricultural lands and the transformation of other (typically annual) croplands and/or traditional olive groves of old trees into this novel form of plantation (Moreira et al., 2019; Morgado et al., 2020, 2021). Olive tree density based intensification is unevenly distributed in the Mediterranean region, with the traditional density frames still representing the vast majority in major olive producer countries, such as Spain, Tunisia, Italy, Greece, and Morocco (Vilar and Pereira, 2018).
Despite the relevance of olive crops, in agronomic and ecological terms, and differently to tropical agroforest systems (Lozada et al., 2007; Pejchar et al., 2008; Araújo-Santos et al., 2021), we lack information on the role of olive groves on the persistence of seed dispersal services in Mediterranean agricultural landscapes. Namely, the impact of SNWH loss derived from olive expansion and intensification is virtually unknown. Because of its savanna-like tree structure and fruit provisioning cropland nature, it could be presumed that olive groves would be relatively permeable for avian frugivore activity and seed dispersal. Olive groves are known to play a crucial role in the lifecycle of many avian frugivores that winter in the circum-Mediterranean area but that have found increasingly shortened natural wintering quarters by habitat loss during in recent centuries (Rey, 1993, 1995). This is favored by the fact that cultivated olive derives from the human selection of one of the more frequent and rewarding fruits of the Mediterranean scrublands, the wild olive (Rey, 2011). Nonetheless, evidence also exists, indicating that the avian frugivore assemblage and diet in olive groves are notably simplified compared to wild olive forests and other Mediterranean scrublands (Rey, 1993; Rey et al., 1996; Rey and Valera, 1999).
This study evaluates these effects at the regional level, focussing on Andalusia (in the south of Spain), to what extent olive expansion and the loss of SNWH—that is, landscape modification towards olive-dominated landscapes- and the intensive agriculture—that systematically removes herbaceous ground cover with herbicides and/or tillage, leaving the soils uncovered–compromise the seed dispersal function supplied by frugivorous birds. This study does not evaluate olive farming intensification based on increased tree density (which considered traditional, intensive, and superintensive tree density frames) since in Andalusia (the largest region/area devoted to olive groves in the world) traditional tree density frames still represent 75% while superintensive hedge-like plantations only reach 2.5% of the land devoted to this crop (Vilar and Pereira, 2018). The removal of ground cover has raised strong environmental, sustainability and biodiversity concerns in Andalusia (Gómez and Giráldez, 2009; Moreno et al., 2009; Paredes et al., 2013; Sánchez-Moreno et al., 2015; Parras-Alcántara et al., 2016; Martínez-Núñez et al., 2020a,b; Gómez-Rosado et al., 2021; Tarifa et al., 2021; for some examples). Awareness of the environmental impact caused by bare soils in the olive cultivation sector is reflected also in European Common Agriculture Policy (CAP) (Díaz et al., 2021). More specifically, we evaluate how landscape and the farm scale, in terms of anthropogenic impact, have affected avian frugivore abundance and diversity, frugivory and seed deposition into the productive (olive field) and non-productive (seminatural woodland) habitats of these human-shaped landscapes. We intend to provide insights for plant regeneration and enhancement of connectivity in these landscapes that could be applied by farmers and land managers. To do this, this study used information from bird censuses and mist-netting (which provide data on avian abundance and seed occurrence in bird droppings) and seed fall traps (for seed deposition data) from 20 olive grove localities widely distributed in Andalusia (southern Spain). The study considered: (i) three levels of the gradient of SNWH cover at the landscape scale (low, intermediate, and high SNWH cover); (ii) two types of habitat patch within each olive farm (seminatural woodland and olive field); and (iii) two types of agricultural practices in paired olive farms per locality (intensive practices of persistent removal of ground herb cover vs. low-intensity management, which maintains the herbaceous cover most of the year). We expected there to be a decrease of frugivore abundance and diversity with SNWH cover loss by olive grove expansion (Rey, 1993) and higher abundance and diversity in seminatural woodland remnants, especially in olive-dominated landscapes (Rey and Valera, 1999). We further expect a higher degree of frugivory in seminatural woodland remnants than in the olive fields and in landscapes with higher SNWH cover than in olive-dominated landscapes with low SNWH cover (Rey and Valera, 1999). Concomitant with these predictions, we also predicted stronger seed deposition in the woodland remnants compared to isolated perching trees or the olive fields, especially in landscapes of high SNWH cover. Finally, because of the mobility of birds at local and landscape scales, we do not expect strong effects of agricultural practices on bird abundance, frugivory and seed deposition.
Materials and Methods
Study System
Olive groves are a woody crop that occupies a large part of the Mediterranean region. With more than 1.5 million ha, Andalusia is the region with the highest area dedicated to this crop in the world. Olive growing has continued to expand in Andalusia in the last several decades, to the detriment of other crops and areas of natural vegetation. This has caused the simplification of the landscape. Over vast areas of the Guadalquivir Valley countryside, olive groves occupy the entire cultivable area and have become a monoculture, which implies a completely homogeneous human-shaped landscape. In areas where the relief is more undulating and parent rock emerges, the olive tree plantation is not viable and small patches of natural vegetation have survived clearing. This has leds to a more heterogeneous landscape in which the olive grove matrix predominates but is dotted by patches of semi-natural vegetation. Another source of heterogeneity in these landscapes are areas where the olive grove is interspersed with other crops, forming an agricultural mosaic. Finally, in the foothills and piedmont of the mountains of the region, large forest, woodland, and scrubland masses can occur along with other woody crops, and olive groves do not comprise the majority of land use. Therefore, a noticeable gradient of olive grove cover and semi-natural habitat cover can be envisaged across the whole olive growing area of the region.
Regarding crop management practices, the conventional and most widespread practice is the intensive and persistent removal of herbaceous cover using chemical methods (herbicides) (hereafter, “intensive management”). These types of practices are very aggressive for the soil, causing erosion and loss of tons of fertile soil every year (Castro et al., 2008; Gómez and Giráldez, 2009; Gómez et al., 2009; Gómez-Rosado et al., 2021). Conversely, other more environmentally respectful practices are notably increasing in the region, since the application of the most recent update to European Common Agricultural Policy (CAP), which entails the maintenance of spontaneous herbaceous cover, that is left to grow for most of the year (hereafter, “low-intensity management”). In an increasing number of cases, this maintenance of herbaceous cover is combined with organic farming, but the area dedicated to organic olive growing is still a minority in the region. Modern hedge-like plantations, although rapidly proliferating are still marginal in the region, and convey a new and different facet of the intensification of olive production that is not considered in the present study (see Morgado et al., 2021, for the effects of superintensive olive farming in avian frugivore abundance and richness). Olive trees in all the localities were more than 30-years old and were grown with a plantation frame of 7 × 8 m or frequently higher.
Experimental Design
To integrate the landscape and local (farm) scale effects of agricultural intensification, this study included 40 olive farms, which were paired in 20 localities (see details in Supplementary Table 1). Each pair of farms was composed by an intensive and a low-intensity farm that were embedded within a same landscape. The mean distance between localities was 105 ± 63.1 km (average ± 1SD). Martin et al. (2019) have shown that both crop and non-crop habitats influence functional biodiversity and agroecosystem services in Europe. We considered these two types of habitat cover for the selection of our study localities, the characterization of their landscapes, and the differentiation of habitats within each olive farm. In particular, the study localities were chosen to cover a gradient of loss of seminatural woodland habitat (SNWH) cover and land use conversion to olive groves, across the main olive growing areas of Andalusia (south Spain). We considered three categorical levels of landscape modification based on information of SNWH cover, landscapes with low (< 5%), intermediate (from 5 to 20%) and high (> 20%) SNWH cover (localities ranging between < 1% and ca. 70% of SNWH cover). In total, 8 localities fell within the Low, 8 in the Intermediate, and 4 in the High SNWH cover categories. Similar categorical approaches based on natural habitat loss (and relatively similar cutting points) have been used for ecological/agroecological conceptualization of the landscapes (e.g., McIntyre and Hobbs, 1999) or for characterizing the complexity of the agricultural landscapes (Tscharntke et al., 2005; Concepción et al., 2008). We further used the olive grove cover of each landscape as an estimator of olive expansion. We used recent land use cartography of the region, based on SIOSE data (http://www.siose.es) and a Geographic Information System software, QGIS v.2.14 (QGIS Development Team, 2018), to calculate the proportion of area occupied by forests, woodlands, scrublands (all integrating the SNWH), and olive groves in an area within a circle of 2 km radius, centered between the paired farms of each locality. We further estimated olive and SNWH covers within a 1 km radius circle around the center of each farm. A 1 km-scale is commonly used for analyses of the effects of landscape heterogeneity on birds. Available studies on seed dispersal kernels by frugivorous birds in the Iberian peninsula, including human-altered landscapes, report that most seeds are dispersed at distances notably shorter than 1 km and that seed dispersal events beyond 2 km are extremely rare (Jordano et al., 2007; Rey and Alcántara, 2014; González-Varo et al., 2017, for Mediterranean landscapes, see also Morales et al., 2013, for Cantabrian range).
Intensive and low-intensity management were defined based on the ground herb cover management, whether persistently eliminated during the whole year with herbicides or maintained for most of the year and eventually removed in late spring by mechanical mowing or livestock grazing (Rey et al., 2019; Martínez-Núñez et al., 2020a,b, for further information of characterization of these two categories of agricultural management). Finally, evaluations of the seed dispersal function (see below) were undertaken by distinguishing two types of habitats within each olive farm: the olive field, corresponding to the productive part of the farm; and semi-natural woodlands patches, corresponding to the unproductive part of the farm (see Figure 1 for the overall study design).
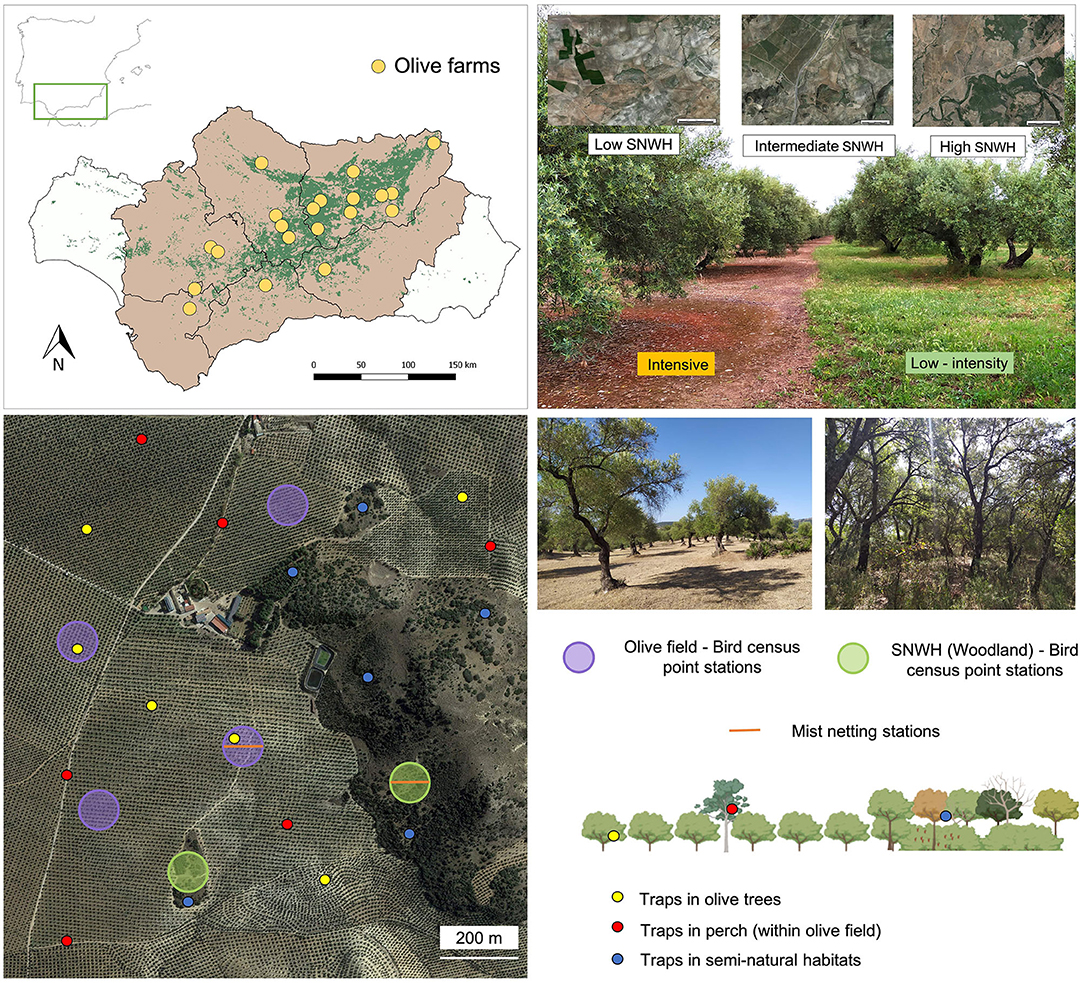
Figure 1. Study sites and sampling schemes for avian frugivore assemblages, frugivory and seed deposition in olive grove landscapes of Andalusia (south of Spain). Upper left panel shows the study area in the south of Spain and the localities of study within Andalusia (indicated by yellow circles in the right panel, each locality with a pair of olive farms differing in agricultural management). Upper right panel illustrates the difference between intensive olive farms (note the bare ground by persistent removal of the herbaceous cover) and low-intensity management farms (with a developed ground herb cover for most of the year). Insets on the upper right panel show ortophotos of olive grove landscapes. From left to right these insets depict an increase in seminatural woodland habitat (SNWH) cover in the landscape (irregular green) and an increase in olive grove cover (although in ortophoto of the low SNWH cover some polygonal cereal croplands are appreciated). Lower panels identify sampling stations within an olive farm, distinguishing between stations of bird censuses (large open circles), seed fall traps stations (small solid circles), and bird mist-netting stations (lines). Colors of stations correspond to different habitats within the olive farm as indicated in the corresponding labeling.
Evaluation of the Dispersal Function
As described in the Introduction, agriculture may filter the dispersal function by: (1) filtering the bird species pool occurring in croplands; (2) modifying the foraging activity of the frugivorous birds (frugivory); and (3) modifying the spatial behavior (mobility) of these birds, and thus affecting where they deposit their droppings (seed deposition). We evaluated several parameters that characterize each of these components of the dispersal function.
Avian Frugivore Abundance and Richness Estimate
From June 2019 to March 2020 (covering most periods of fruit availability in the Mediterranean region and the time in which birds majorly consume fruits), we conducted 5-min bird censuses at circular point census stations (with a 50 m radius), every month. We located 10 permanent census stations (6 in the olive fields and 4 in SNWH patches) in large olive farms (> 25 ha) and 6 stations (4 in olive fields and 2 in SNWH patches) in small farms (<25 ha, typically less than 10 ha). All birds seen and heard within each station were recorded, although we only considered fruit-eating birds according to classifications outlined by Storchová and Horak (2018). Censuses were conducted within 4 h of sunrise.
We obtained for each census station an estimate of fruit-eating bird abundance and species richness and characterized the frugivore guild of the olive grove landscapes. Abundance and richness were estimated as the mean number of fruit-eating birds recorded across monthly surveys in each station.
Frugivory
We intended to quantify the magnitude of frugivory of the pool of potential frugivores detected in censuses. From September 2019 to March 2020, we conducted bird mist-netting sessions in the study olive farms. Birds trapping was carried out for 3 h in each session (between mid-morning and noon). For each farm, two simultaneous capture zones, one in the olive field and the other in SNWH patches were considered, with a distance of 150 meters between zones. In each zone (habitat), we set two mist-nets of 12 × 2.5 meters and a mesh size of 16 mm (24 linear meters of the net in total per zone). In each mist-net zone, a sound call that emitted songs of the frugivorous species present was arranged at random to attract birds.
A 1-m wide strip of mosquito net was placed on the ground under the mist nets to collect the seeds excreted by the birds while they were trapped in the net (see details in Supplementary Figure 1). Once a bird was released from the net it was immediately introduced in a cloth bag with a paper cone located inside (where the excreted/regurgitated seeds were collected) until the moment of seed collection/identification. All birds were kept in the cloth bags for around 1 h and then ringed and released. The 1-h bird holding time intended to balance the time typically required for seed regurgitation/defecation of avian frugivores (temperate avian frugivores have seed retention times in their guts of less than 1 h, e.g., Herrera, 1984b; Sorensen, 1984; Fukui, 2003) while trying not to compromise the birds' daily energy requirements and food provisioning. For each individual captured, the total number of seeds of each species collected under the net and in the collector was recorded.
Bird trapping, handling, and banding were carried out by two expert ringers (FMC and RT) who had ringing licenses and authorization for the scientific banding of wild birds in Andalusia.
Logistic constraints hindered sampling in each olive farm of each locality every month. Therefore, we decided to conduct a monthly trapping session in one of the farms of each pair (all the localities thus having a bird trapping session per month). Consequently, the statistical analysis on frugivory will be based only on these 20 farms where birds were more actively trapped (see statistical analyses section). Among the farms sampled extensively for frugivory, there was a relatively even distribution between intensive and low-intensive farms and among SNWH categories. In the other member of each pair of farms, we set mist-nets just twice during the whole study period. Data from these last farms were used only for completing a general description of fruit species consumed by the frugivore assemblage in olive groves.
From this information, we reported three components/descriptors of the frugivory intensity: the proportion of the mist-netted frugivore species with seeds in their droppings, the abundance of seeds per dropping (including droppings without seeds) of fruit-eating birds, and the seed species richness per dropping. The first elucidates to what extent the potential frugivores can disperse seeds in olive grove landscapes, the second captures the magnitude of this activity, and the third provides information on the diversity of their fruit foraging activity.
Parallel to the sessions of bird mist-netting, we exhaustively surveyed the number of fruiting species around a 50 m radius circular plot. This allows us to gain an idea of the fruiting species richness in the neighborhood of the nets. Fruit species richness other than cultivated olive was frequently null around nets in the olive field while in the forest woodland patches decreased with the level of SNWH loss: average of 1.6 species (range: 0–7) in landscapes with low SNWH cover, 3.7 species (range 1–12) in intermediate landscapes, 6.8 species (range 4–8) in landscapes with high SNWH cover. Given the high avian mobility, 50 m-radius scales around nets may be too small to fully characterize fruit availability for avian frugivores in a locality. Consequently, we further assessed the fleshy fruit species detected in all 50 m radius census stations used for surveying frugivorous bird assemblages. We show the fruits available at each locality in Supplementary Figure 2. Again, the richness of the fruit species available at farm scale decreased with the level of SNWH loss in the landscape: average of 4.4 species (range: 0–16) in landscapes with low SNWH cover, 10.0 species (range 4–20) in intermediate landscapes, 12.0 species (range 9–16) in landscapes with high SNWH cover.
Seed Deposition
To determine the probability of seed deposition into different habitats within the olive farm and to characterize seed rain, we used plastic plant pots of 40 cm diameter (0.1257 m2 of collecting area) and 20 cm depth as seed fall traps. Traps were covered with a 1 × 1 cm wire mesh to avoid seed predation by micro mammals and perforated on their base to drain rainwater. To avoid seed drag by water drainage, we glued a mosquito mesh (1 cm above the trap button) that retained the seeds (e.g., Ficus carica seeds) to the trap wall. Eighteen seed fall traps were placed per farm in three different types of habitats (6 traps per habitat): beneath the olive tree canopy, beneath the canopy of isolated overtopping non-olive trees serving as perches to birds within the olive field, and within SNWH remnants. Traps were always set hanging from branches of trees and/or tall scrubs (Supplementary Figure 1).
The traps were active for 17 months, between October 2018 and March 2020 in 9 localities (3 in each of the landscape categories) out of the 20 study localities (18 olive farms). We collected the trapped seeds periodically (every 3 months, with monthly-bimonthly checks to make sure that the traps were active). All seeds collected were pooled at the trap level for these analyses.
From this data, we obtained for each trap three components of the seed deposition by birds: the event of seed arrival (1 vs. 0) as the trap collecting or not collecting seeds; the number of seeds collected, and the number of seed species collected.
Statistical Analyses
For each response variable considered for frugivore assemblage (abundance and richness), frugivory, and seed deposition, we conducted Generalized Linear Mixed Models (GLMMs), considering habitat loss (i.e., level of SNWH cover: low, intermediate, and high SNWH), agricultural management (intensive vs. low-intensity), and habitat type (SNWH habitat vs. olive field) as fixed categorical effects. The olive grove cover at the 1-km circular radius was additionally considered as a continuous covariable since, although negatively correlated with SNWH (r = −0.59; N = 40 farms), such correlation was not strong enough to generate serious variance inflation factors in the models. Note that SNWH and olive cover, although related, describe different processes, habitat loss (not exclusively due to land conversion to olive grove), and olive expansion, respectively. We also considered all the possible interactions among these four fixed effects. The locality was considered a random blocking effect in all these analyses. In the case of bird abundance and richness per census station, we pooled data for all the study period (as a mean per census station). In the case of models of frugivory, the monthly trapping sessions conducted in the single station of each habitat within the farm were nested within a locality. We used Gaussian family distribution for a mean number of bird abundance (log-transformed) and richness per census station provided the non-negative continuous nature of the data averaged per point census station. Visual inspections of the residuals support that these response variables were distributed normally. In the case of frugivory, due to the frequency of zeros, we used negative binomial family models both for the number of seeds and number of seed species per dropping, and binomial models and logit link function for the proportion of frugivore species with seeds in their droppings. Finally, in the case of seed deposition, we used binomial family models and logit link function for probability of seed deposition, negative binomial for the total number of seeds collected, and Poisson distribution and log link function for the number of species collected per trap. All models were fitted using the R package glmmTMB (Brooks et al., 2017) and their R values (the marginal R2, that is, the variance explained by the fixed effects only) were obtained with the R package MuMIn (Barton, 2015). A model selection procedure based on AIC was used to choose the best model for each response variable among all models that were better than the null model (including only the random factor). When two or more models performed well but were indistinguishable (i.e., when differing in AIC < 2), we typically opted for the most parsimonious (attending to the behavior of the residuals of each model), or used the example that contained a biologically significant or marginally significant effect. The comparison of each competing model against the null model was conducted using the dredge function from MuMIn (Barton, 2015).
Results
Variation in the Frugivore Abundance and Richness Across Olive Grove Landscapes and Agricultural Management Practices
Thirty five species of birds classified as frugivores occurred in the studied olive groves (Table 1) throughout the whole period of fruit availability considered (June 2019–March 2020). Among these species, some were resident, occurring in olive groves throughout all the year (Sardinian warbler, the European blackbird, or the European robin among the most common), while others occurred temporally, mainly as wintering (for example several thrushes, Blackcap or Black redstart), or during the post-reproductive period and migratory pass (several Curruca and Sylvia species, flycatchers, and the Common redstart), during summer and autumn. The wintering Blackcap and Song thrush and the resident Sardinian warbler, European blackbird, and European robin (the latter leaving the olive groves in the majority of localities during the reproductive season) were by far the most abundant and ubiquitous species. Overall, fruit-eating birds were more than two-fold more abundant in olive grove landscapes during the winter period compared to summer and autumn (Table 1), though the richness of frugivorous species was higher during summer-autumn (32 species in summer-autumn vs. 24 in winter).
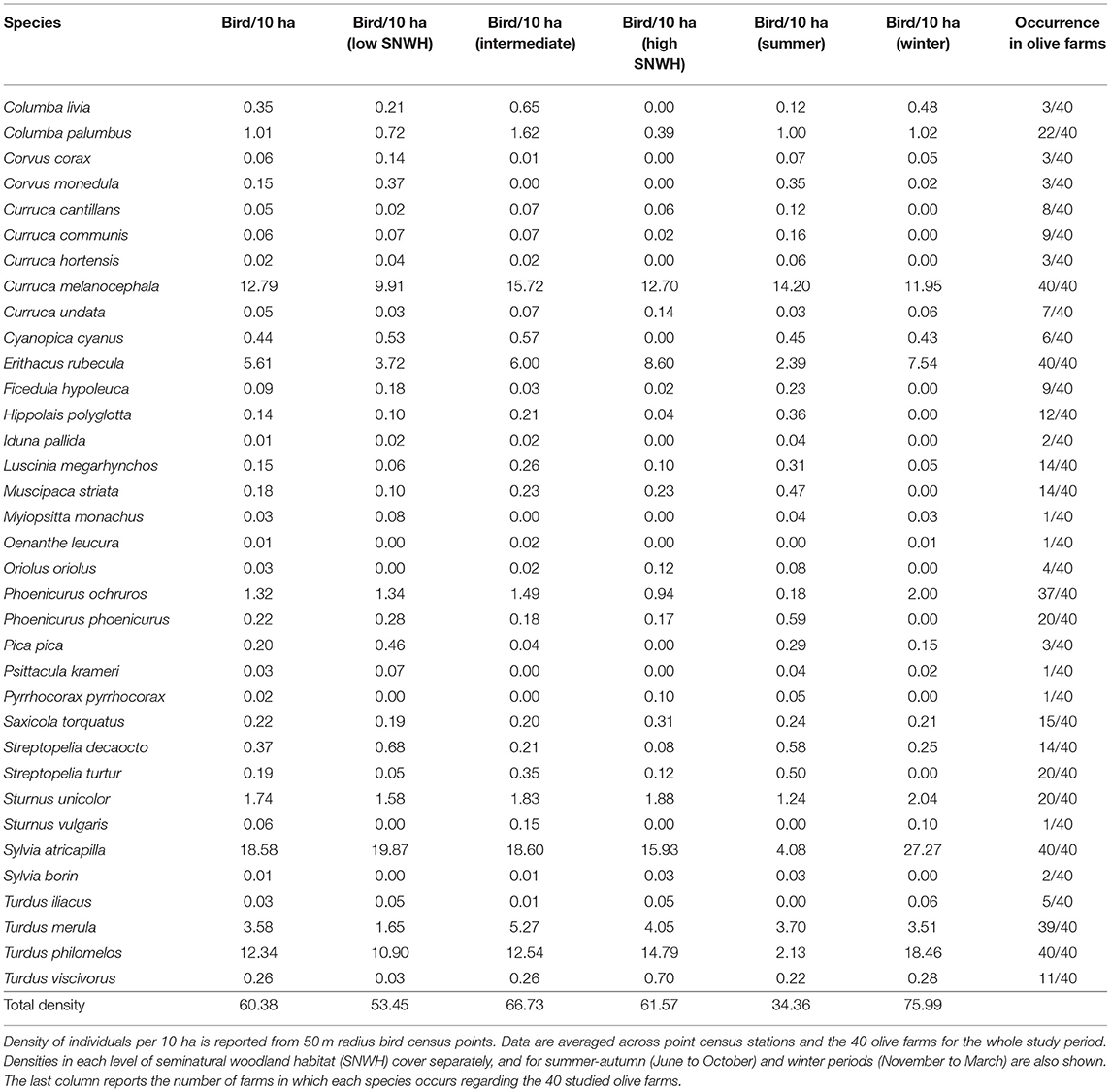
Table 1. List of frugivorous birds recorded in bird censuses in olive farms and their respective abundances during the period of fruit availability (June 2019 to March 2020).
The abundance of fruit-eating birds varied notably among olive groves (ranging from 6.6 individuals/10 ha to 24.1 individuals/10 ha). Our model selection procedure, considering effects of agricultural management (M), SNWH and olive grove cover (OGC) at the landscape level, and habitat within the olive farms (H), showed that only a single model was better than the null model (i.e., the model considering only the random factor). The selected model (Table 2) showed significant effects of H on avian frugivore abundance, being consistently higher in SNWH patches than in the olive fields (Figure 2A). Furthermore, we found an interdependent effect of H x OGC, meaning that frugivores increased their abundance in seminatural woodland patches but remained invariant in the olive field as olive grove cover increased in the landscape (Figure 2A).
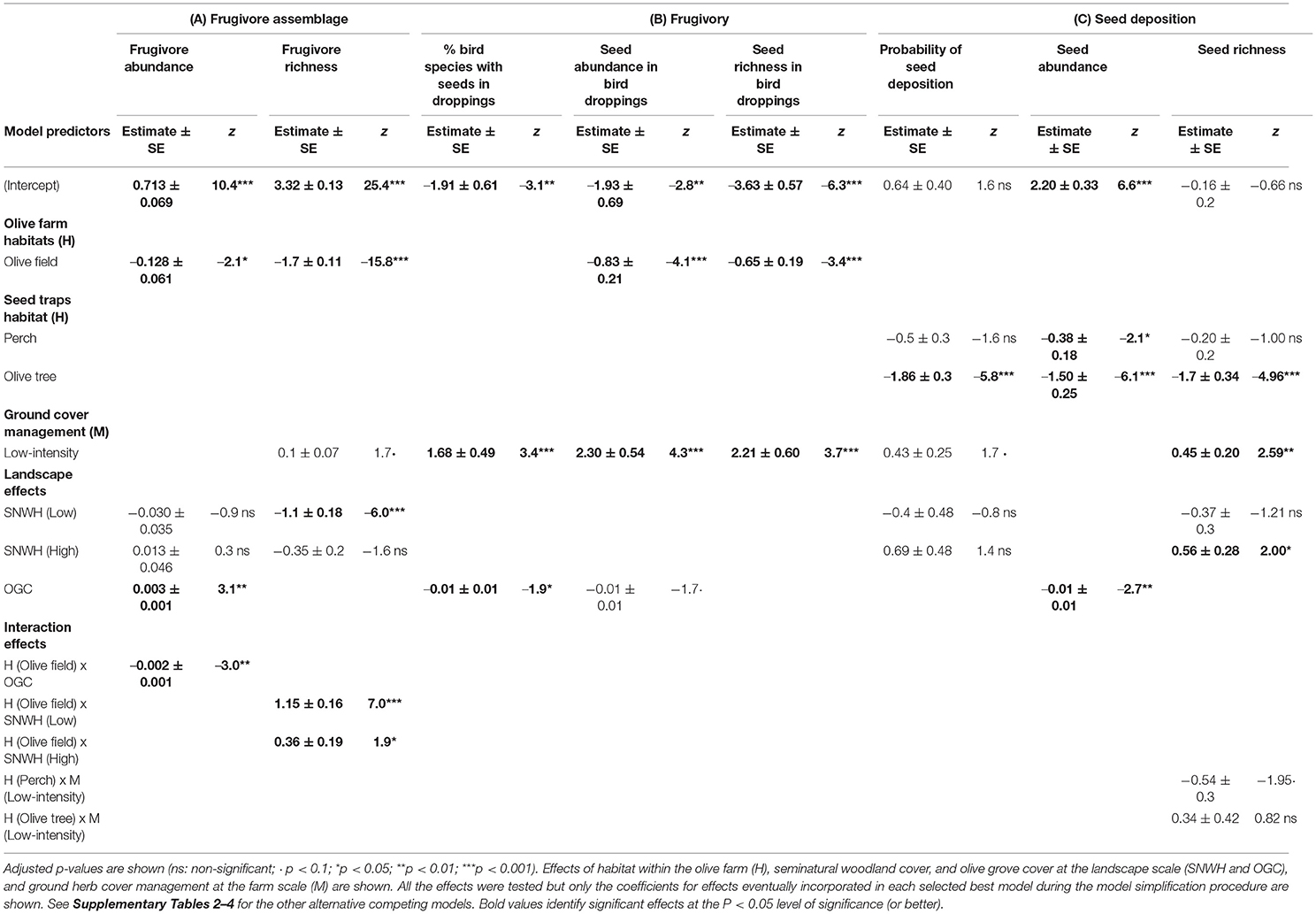
Table 2. Selected best model parameters after a selection procedure based in AIC for all the descriptors for avian frugivore assemblage abundance and richness (A), Degree of avian frugivory (B), and seed deposition (C) used as response variables in this study.
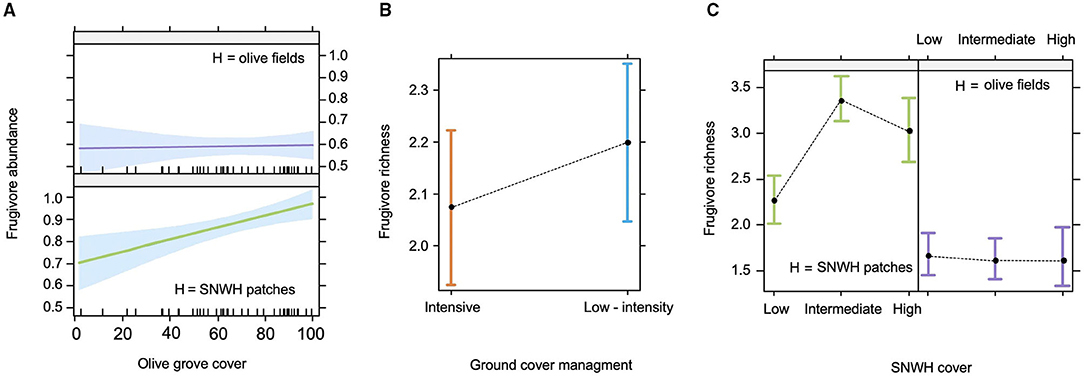
Figure 2. Significant effects in the selected models for avian frugivore abundance (A) and species richness (B,C). Note that in (A) the Y axis is in log10 scale, and that frugivore abundance ranges from 10(0.5+1) = 30 individuals, to 10(1+1) = 100 individuals/10 ha. Data represented are predicted least-squares means and 95% confidence intervals. Estimates of fixed effects are shown at the reference mean value for all the covariables. H refers to habitat (olive field and SNWH patches) where censuses and other samplings were conducted. SNWH refers to seminatural woodland habitat and SNWH cover refers to categories (low, intermediate, and high) of seminatural woodland cover in the landscape. OGC means olive grove cover at the landscape level and M is the local agricultural management (intensive and low-intensity ground herb cover management).
In the case of frugivore species richness, two models were significantly better than the null model (Supplementary Table 2A). Both models showed that frugivore species richness was affected by H and by SNWH cover, but these effects were fundamentally interdependent. While frugivore richness was higher in seminatural woodland patches in all landscapes, it was invariantly low in the olive field (Figure 2C), that is, more SNWH cover did not increase frugivore richness in the olive field. Interestingly, this interaction effect also shows that frugivore richness in remnant SNWH patches was lower in landscapes with low SNWH cover compared to landscapes of intermediate or high SNWH. In addition to these effects, we also found a marginally significant effect of M (Table 2A). In particular, species richness was higher in low-intensive farms than in intensive ones (Figure 2B). In any case, this latter effect should be considered with caution since the alternative more parsimonious model, not including M, was not significantly worse (although it had higher AIC, ΔAIC < 2).
Variation in Frugivory
Thirteen out of 17 mist-netted frugivorous bird species defecated/regurgitated seeds of fleshy fruit species in the whole set of olive groves studied (Table 3). They defecated/regurgitated 2,063 seeds of 27 different fleshy-fruited plant species (Table 4). Blackcap (Sylvia atricapilla) was by far the most important seed disperser, disseminating seeds of 26 out of the 27 species recorded (except Juniperus oxycedrus) in 31 olive groves and accounting for 90.4 % of total collected seeds in droppings. The other two major seed dispersers were the resident Sardinian warbler (Curruca melanocephala, dispersing seeds of 7 fleshy-fruited species in 18 olive groves), and the migrant Garden warbler (Sylvia borin, dispersing seeds of 4 species in 4 olive groves). Wild fruits accounted for a notable amount (62% in total) of seeds in bird droppings while cultivated olives represented 31% and other cultivated or naturalized species 7%. The plant species whose seeds were more frequent in bird droppings in olive groves were the cultivated olive (Olea europaea var. europaea), Pistacia lentiscus, Solanum spp., Rhamnus lycioides, and the cultivated/naturalized Ficus carica (> 100 seeds in birds droppings for each of them) and Phillyrea angustifolia. The seed species occurring in bird droppings in more different localities (i.e., more ubiquitous in the orchards under study) were the cultivated olive (found in droppings in 29 olive groves), Asparagus spp. (15 olive groves), F. carica (15), Pistacia lentiscus (14), Solanum spp. (13) and Rubus ulmifolius (10). Sixteen fleshy fruit species appeared only in droppings of Blackcap, while one species (Juniperus oxycedrus) occurred only in droppings of Mistle Thrush (Turdus viscivorus). Apart from the cultivated olive (dispersed by 7 bird species) and F. carica (dispersed by 5 species), the fleshy-fruit species attracting a higher number of frugivorous birds were P. lentiscus and R. lycioides (occurring in droppings of 6 and 4 bird species, respectively).
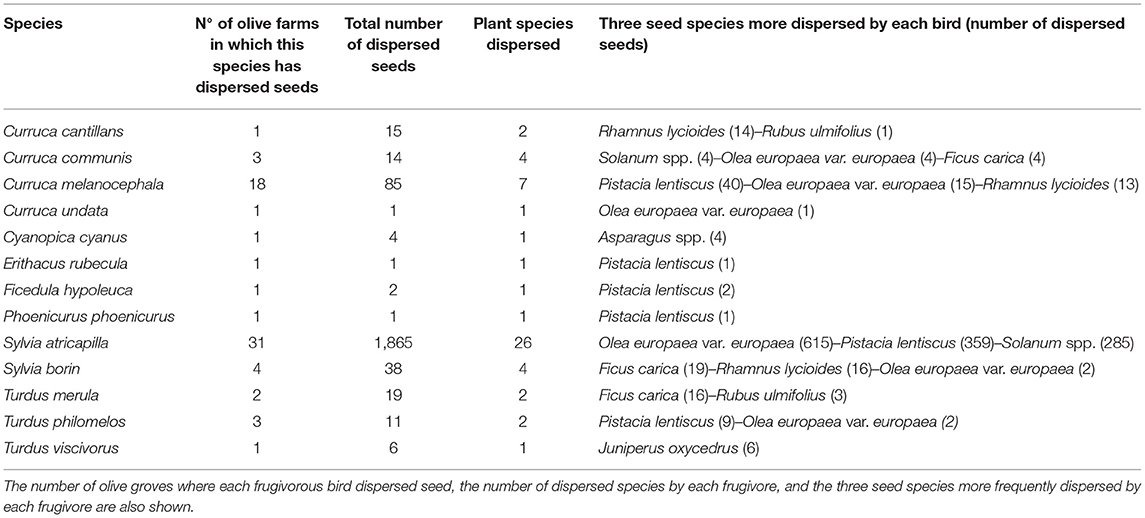
Table 3. Frugivorous bird species dispersing seeds in olive groves and number of seeds occurring in their droppings/regurgitations.
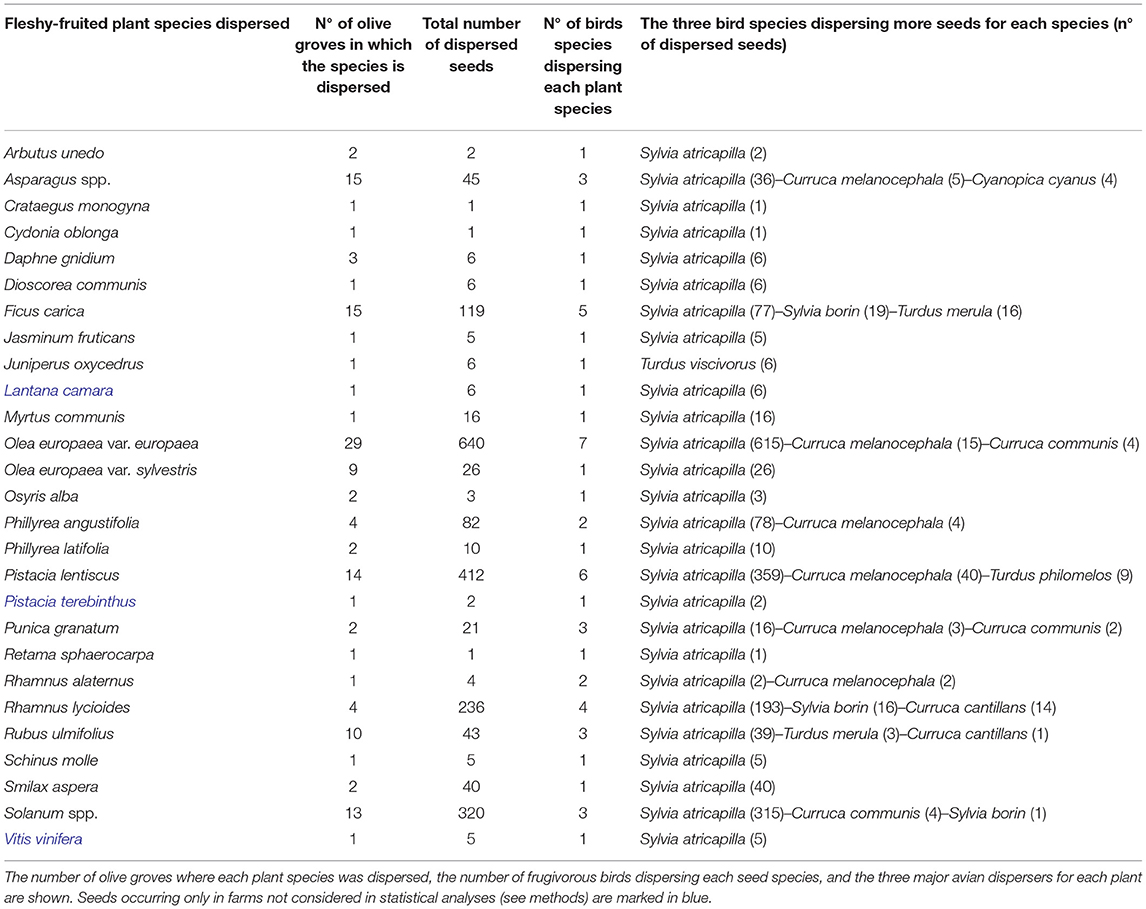
Table 4. Fleshy fruit species dispersed by birds in olive groves and number of dispersed seeds (i.e., collected in droppings) of each plant species.
Our models showed that frugivory varies fundamentally among habitats within olive farms and management regimes with some influence also of the olive grove cover, while the level of SNWH cover per se did not improve these models. Thus, in the case of the proportion of frugivore species dispersing seeds (i.e., with seed in droppings) no effect was detected (i.e., no model was better than the null model) when seeds of cultivated olives were considered (results not shown). Nonetheless, the exclusion of the seeds of cultivated olives from the analyses renders multiple models better than the null model. Supplementary Table 3 shows the five models with the lowest AIC. The selected model (PFS3) incorporated significant effects of M (with low-intensity farms with ca. 4 times more proportion of frugivorous bird species dispersing non-cultivated olives, Figure 3A) and OGC in the landscape (Table 2B), with the frequency of species dispersing seeds decreasing with the olive cover (Figure 3B). This model was substantially similar to model PFS1 (with lower AIC, Supplementary Table 3) but was preferred because the latter incorporated the non-statistically significant effect of H.
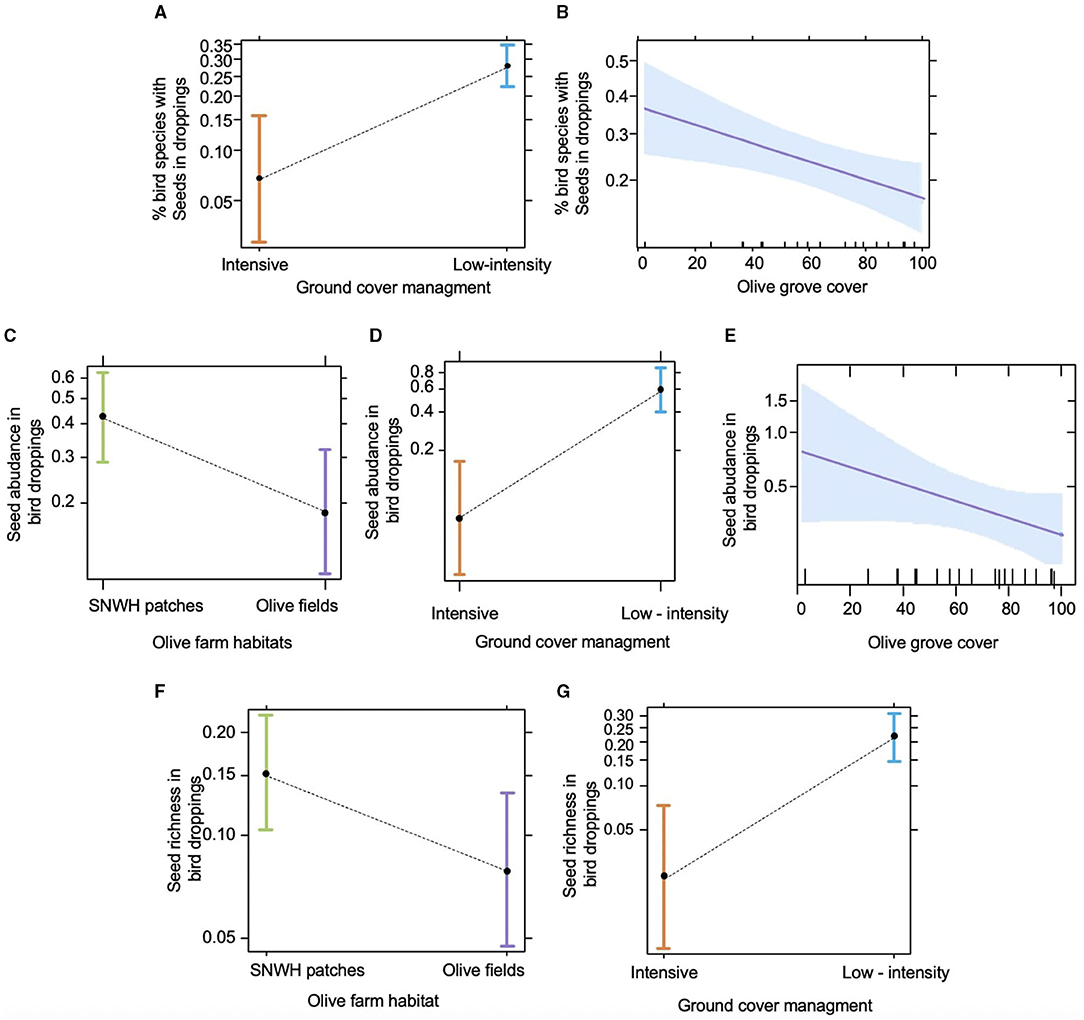
Figure 3. Significant effects in the selected models for frugivory using as descriptors proportion of trapped frugivorous species with seeds in droppings (seed disperser probability in A,B), the abundance of seeds per dropping (C–E), and seed species richness in droppings (species richness per dropping in F,G). Data represented are predicted least-squares means and 95% confidence intervals. Estimates of fixed effects are shown at the reference mean value for all the covariables. See Figure 2 caption for code description.
In the case of the abundance of seeds per frugivore dropping, all five models that were better than the null model incorporated H and M as significant predictors, and four of them also included OGC. The best model (SAD2; see Supplementary Table 3) showed that the abundance of seeds in droppings was significantly higher in the seminatural patches than in the olive fields (a more than two-fold increase, Figure 3C), increased substantially in low-intensity management compared to intensive farms (Figure 3D) and decreased with OGC (Figure 3E), though this last trend was marginally significant (Table 2B). Likewise, all significant models for variation in seed species richness in the bird droppings incorporated H and M (some of them also OGC) (Table 2B), and the selected best model (SRD1) determined a significant increase in the diversity of seeds in droppings in seminatural patches compared olive fields (Figure 3F) and in farms under low-intensity management compared intensive farming (Figure 3G).
Variation in Seed Deposition
In total, we collected 1,160 seeds of 34 fleshy fruit species in the 18 olive groves where traps were set up (see Table 5). The rate of seed deposition steadily increased over time: in 14 months (between 2018 and 2019) we collected 523 seeds in 186 (40.2%) out of 462 active seed fall traps (mean number of seeds collected per trap capturing seeds = 2.82 seeds), while in 3 months (winter) in 2020 we collected 637 seeds in 205 (47.4%) out of 432 active seed fall traps (mean number of seeds collected per trap capturing seeds = 3.11 seeds). Thirty traps were lost during the period of study. Referred to the total active traps in each period these figures represent 9.0 and 11.7 seeds/m2 each period. We were unable to classify 9 out of the 34 species collected (Table 5). Unclassified seeds accounted for less than 1% of the collected seeds. Sixteen species were only collected in a single olive grove.
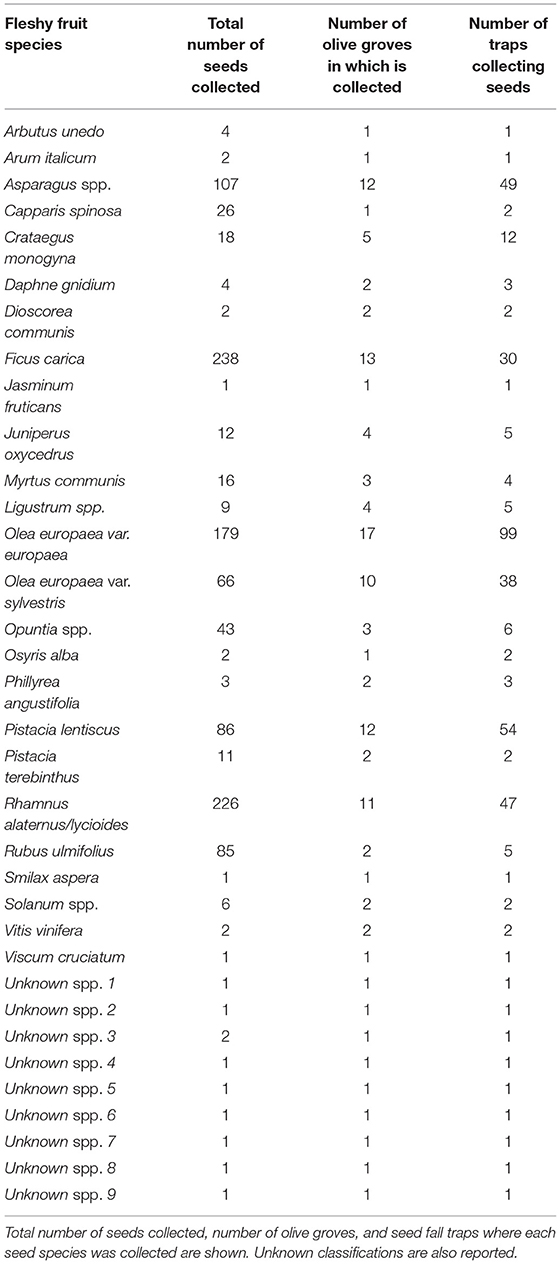
Table 5. Collected seeds in the seed fall traps of 18 olive farms of 9 localities of study.
The most ubiquitous dispersed seeds were cultivated olive (collected in 17 out of 18 olive groves) and F. carica (13), and among the native species, Asparagus spp. and Pistacia lentiscus (both collected in 12 olive groves), Rhamnus alaternus/lycioides (11) and Olea europaea var. sylvestris (wild olives; 10). The cultivated and wild olives, P. lentiscus, Asparagus spp., Rhamnus alaternus/lycioides and F. carica seeds fell at least in 30 seed traps. In contrast, R. ulmifolius, although abundantly collected (85 seeds), was registered only in 2 olive groves and 5 traps. Notably, wild plant species accounted for 60% of the seed rain while cultivated olives accounted only for 15% of the deposited seeds and other naturalized or cultivated species (Ficus, Opuntia, Vitis) represented the remaining 25%. These figures were, overall, congruent with data on bird droppings.
A set of models was significantly better than the null model for capturing the variation in each descriptor of seed deposition (Supplementary Table 4). Thus, in the case of the probability of seed deposition into seed fall traps, the models incorporated different combinations of H, M, and SNWH cover (Supplementary Table 4). The model with lower AIC (PSA1) showed significantly higher seed arrival to natural/seminatural patches and perches than the olive field (Figure 4A) and marginally significant differences between local management (Table 2C), with higher seed arrival in low-intensity farms (Figure 4B). This model also incorporated the effect of SNWH cover (Table 2C), with an increased gradient of seed deposition as SNWH cover increases in the landscape; although the corresponding estimated values were not statistically significant (Figure 4C). The effects of M and SNWH cover should be taken with caution since this model was not better than the more parsimonious PSA5, which only incorporated the effect of H but had higher AIC. Regarding the abundance of seeds collected by each trap, better models than the null model included combinations of H, M, SNWH cover, and OGC (see models SAT1 to SAT5 in Supplementary Table 3). The selected best model (SAT4) showed a significant increase in the number of seeds collected by traps in the SNWH patches and isolated perch trees compared to under the canopy of olive trees within the olive field (Figure 4D). The number of seeds per trap also significantly decreased as olive cover in the landscape increased (Figure 4E). Finally, for the number of seed species collected per trap, we achieved 3 models better than the null model.

Figure 4. Significant effects in the selected models of the seed deposition variability considering as descriptor seed arrival probability (A–C) seed abundance (D,E), and species richness (F,G) in the seed fall traps. Data represented are predicted least-squares means and 95% confidence intervals. Estimates of fixed effects are shown at the reference mean value for all the covariables. See Figure 2 caption for code description.
The two models with less AIC (SRT1 and SRT2) were substantially equivalent. Thus, they showed both simple and interactive effects of H and M (Table 2C), with more diversity of seeds deposited into SNWH patches and isolated perch trees within the olive fields than under the canopy of olive trees, with low-intensive farms receiving more seed species than intensive farms only in the olive field (Figure 4G). The significant effect of SNWH cover (significant in the case of the best model based on AIC) and OGC (in the second model) in these two models were interchangeable, with this equivalence showing that SNWH loss and the associated expansion of olive groves reduced the diversity of seeds in the seed rain (Figure 4F for SNWH cover effect). The coefficient for OGC was negative in the alternative model (not shown).
Discussion
Avian-mediated seed dispersal is a fundamental function in the dynamics of Mediterranean woodland ecosystems (Herrera, 1995). In the Mediterranean lowlands, woodland ecosystems have been largely replaced by olive, vineyard, and cereal croplands throughout a millenary history of cultivation in the region. However, comprehensive studies of the effect of agriculture on seed dispersal by frugivorous birds in the Mediterranean are lacking. The present study shows the extent to which the landscape transformation from woodland habitats to olive grove landscapes (the large scale anthropogenic impact of agriculture) and the local (farm) scale of intensification of the agriculture practices (i.e., ground herb cover removal that leaves soil uncovered), combine their effects to filter the assemblages of frugivorous birds and simplify their frugivory activity and seed deposition patterns. The data of the present study represents the most comprehensive and large-scale survey conducted on the effects of agriculture (specifically olive agriculture) on frugivory and seed dispersal in Mediterranean ecosystems.
The results show the remarkable persistence of the dispersal function of frugivorous birds for Mediterranean fruits in olive grove landscapes since more than 60% of seeds found in bird droppings and the seed rain were from wild native species, accounting for a relevant fraction of the regional flora with fleshy fruits. The results of this study also show that agriculture filters avian frugivore richness and abundance, causing a severe decay of the seed dispersal function. We confirmed our prediction on the important and pervasive role of seminatural woodland habitat remnants within farms for conserving the dispersal function through enhancing frugivore abundance and diversity, increasing frugivory, and attracting seed deposition. Agriculture impacted the seed dispersal function fundamentally by causing changes at the landscape scale, involving woodland habitat loss and conversion to olive fields (and other agricultural uses), and ultimately threatening this function in olive-dominated landscapes. Thus, in parallel to the habitat effect, we confirmed that an impoverished frugivore assemblage (in terms of diversity and abundance), decreased levels of avian frugivory, and decay in the abundance and species diversity of the seed rain are to be expected as seminatural woodland cover decreases or olive grove cover increases in these landscapes. Although we did not expect to find important effects of intensification of the local agricultural practices of ground cover removal, negative effects arose frequently, warning of the additional consequences of local agricultural management on frugivore activity and on the seed rains they generate in this agroecosystem. These results have important implications for the management and preservation of seed dispersal services at several scales.
The Assemblage of Frugivorous Birds in Olive Grove Landscapes Is Functionally Heterogeneous but Locally Impoverished, Which Contributes to Bias Seed Dispersal, Limiting It to a Few Fruit Species
The role of olive groves as winter quarters for birds and for maintaining their frugivory activity in the Mediterranean Basin has been remarked on in several studies (Rey, 1993, 1995, 2011). These studies focused on the local scale of the olive fields rather than at the landscape scale and showed that olive groves host an impoverished assemblage of frugivorous birds that found this agroecosystem suboptimal. This is because olive groves are simplified compared to natural scrublands and forests, both structurally and in terms of fruit and other food resources, which affect the diet, the foraging behavior, and body condition of the frugivorous birds still able to settle in this agroforest-like agroecosystem (Rey and Gutiérrez, 1996, 1997; Rey et al., 1996, 1997; Rey and Valera, 1999). Our results confirmed previous suggestions. Thus, practically, the totality of avian seed dispersers in the lowlands of the region were represented in the olive grove landscapes considered as a whole (Table 1). They consumed and dispersed a substantial number of the fruiting species found in the region (Tables 3–5). However, frugivore assemblages became impoverished locally, as did seed dispersal function.
The frugivore assemblage of the olive grove landscapes seems to be characterized by its functional heterogeneity and the potential complementarity of seed dispersal services, which are fundamental for ensuring seed dispersal quality and resilience in real human-shaped landscapes (García and Martínez, 2012; García et al., 2013; Escribano-Ávila et al., 2014). Frugivores that contribute more significantly to seed dispersal in olive groves are either wintering birds (Blackcap) or resident species (Sardinian warbler and European blackbird), which consume fruit during all seasons and contribute to seed dispersal of the whole set of species available. Along with these species, some other bird species occur in olive groves dispersing fruits during short periods. They typically consume fruits and disperse seeds during a migratory pass, and include flycatchers and redstarts and especially, several Sylvia and Curruca species (Jordano, 1982, 1984, 1988; Herrera, 1984a).
The frugivore assemblage inhabiting olive grove landscapes varies considerably in body size with small, medium-sized, and large frugivores (Herrera, 1984b; Jordano, 1987), allowing complementary seed dispersal of all wild and cultivated fruits in the region (the latter pecked rather than swallowed in many cases, Rey and Gutiérrez, 1997). Finally, among common frugivores in olive groves, avian mobility and the ability to track fruit resources in human shaped landscapes is especially pronounced in Blackcap and Song thrush (Rey, 1995; Tellería et al., 2008). At the landscape scale, long distance flights between fragments of natural habits across olive groves are frequent in some large frugivorous birds of genus Turdus, Columba, and among Corvidae (authors pers. obs.; see also Perea and Gutiérrez-Galán, 2016).
These results indicate that the frugivore assemblage inhabiting olive grove landscapes has the potential to adequately disperse most fruit-bearing species in the region. However, the seeds in the droppings collected (Tables 3, 4) and findings in the seed traps show that the fruits of many species are being consumed and dispersed extremely infrequently by most frugivores in olive grove landscapes, and that, in many cases, their dispersal is carried out by only one or two frugivorous bird species at most. Moreover, although our results illustrate remarkable frugivore activity, the low ratio of dispersed and available species (Supplementary Figure 2) indicates that a notable number of species remain undispersed at each locality (% of available but not dispersed species ranging from 13 to 100%). In addition, some species were not (or rarely) dispersed regionally, despite being present in these olive grove landscapes. Interestingly, these results also illustrate that some seeds were dispersed over long distances since they were found to be dispersed at the farm scale but seemed to be absent in the farms (this last phenomenon is identified as gray-colored empty cells in Supplementary Figure 2).
The frugivory activity was strongly dominated by two generalist frugivores, Blackcap and Sardinian warbler, which are also major seed dispersers in the native vegetation of the Mediterranean region (Jordano and Herrera, 1981; Herrera, 1984a). Frugivory activity and dispersal were biased toward some plant species, particularly the cultivated and the wild olives and Pistacia lentiscus, two lipid-rich winter fruits that account for more than 50% of the seeds in bird droppings and ca. one-third in the seed traps, despite both are single-seeded fruits. These two fruit species are known to support the frugivorous diet of wintering birds in the lowlands of the Mediterranean region (Herrera, 1984a; Jordano, 1984; González-Varo et al., 2017; Parejo-Farnés et al., 2020) and in olive groves and wild olive scrublands (Rey, 1992; Rey et al., 1997; Rey and Valera, 1999). Besides these species, some summer fruits (F. carica, R. ulmifolius, Solanum spp., and R. lycioides, the latter common in many habitat remnants) that typically serve as fruit sources for resident and migrant frugivores (Jordano, 1982, 1988; Herrera, 1984a) were abundantly dispersed locally during summer-autumn (Tables 4, 5).
This bias in frugivory and seed dispersal is in part due to the lack of some frugivores in olive dominated landscapes, where for example, some migrant Curruca and Sylvia and Oriolus species or resident Turdus viscivorus, were virtually absent, while others like Turdus merula and Erithacus rubecula decay sharply with the loss of woodlands. In particular, the seed dispersal of all but one fruit species involves the Blackcap, even though some fruits were available mainly when this species has left most olive groves. Blackcap was by far the most abundant frugivore in winter, and the one most captured in mist nets, while large birds, even those that were relatively abundant, were rarely captured if any with mist-nets, meaning their contribution to frugivory and seed rains may be underestimated. Although Blackcap leaves olive groves during spring and summer, some individuals remain in olive groves in the piedmont of the mountain systems of the region during summer-autumn periods and consume summer-autumn fruits, which explains its ample fruit diet. Even though some frugivores, such as European robin, Song thrush, and European blackbird, are known to disperse many seed species in Mediterranean forests and scrublands (Herrera, 1984a; Jordano, 1984); they dispersed a relatively low number of fruit species in the olive grove landscapes. Although they were captured less than Blackcaps (compared to its abundance in censuses) their frequency of occurrence in seeds in droppings (proportion of droppings with seeds) was rather low in our study, for example, European robin (0%), Song thrush (9%) and European blackbird (7.4%) (see also Rey, 1992; Tarifa et al., 2021). It seems that these species are slower to consume fruits in olive grove landscapes than in natural habitats, perhaps because in many olive grove landscapes these fruits are too scarce to attract these birds or to make profitable the tracking of their availability.
In short, these results suggest that olive expansion and concomitant loss of forest/scrubland patches constrain local fruit and frugivore assemblages, limiting their fruit consumption, and disrupting the seed dispersal and seed rain of many plant species.
Habitat and Landscape Effects: Woodland Habitat Loss and Olive Grove Expansion Filter Frugivore Assemblage, and Simplify Frugivory and Seed Deposition Patterns
We found an important pervasive effect of the type of habitat within the farm on frugivore abundance and diversity, frugivory, and the seed abundance and diversity in the seed rain. Avian frugivores tended to concentrate in forest/scrubland patches and their activity of fruit consumption was higher compared to those in the olive field, as revealed both by the number of seeds and species richness in droppings, which subsequently translated into the concentration of the seed deposition in the natural habitat remnants. This is related to the fact that within the olive fields most fruiting species are actively eliminated by agricultural practices, with only some species, such as Asparagus spp., Solanum spp., Capparis spinosa, persisting (Rey, 2011; Tarifa et al., 2021) in the olive fields of low-intensity farms. This is in sharp contrast to the availability of non-cultivated fruits in the remnant forest patches (Tarifa et al., 2021). This type of effect of patch quality on avian abundance and richness, and on frugivory and seed arrival, has been reported in other landscapes of the Iberian Peninsula, which were fragmented or structurally degraded by annual croplands and livestock grazing (Santos and Tellería, 1994; Alcántara et al., 2000; García and Chacoff, 2007; García et al., 2010; Herrera and García, 2010; Rey and Alcántara, 2014). Similar local or patch effects are also typical in tropical agroforest systems, such as cocoa and coffee plantations (for instance, Lozada et al., 2007) that, as olive groves, are permanent croplands. In these agroforest systems, the practice of growing the crop under a more or less diverse canopy of tropical trees (shade plantations), compared to the most common and productive sun plantations, conditions the farm for frugivores and affects their frugivory activity and seed deposition patterns, which have been shown to approach those reported on intact forests (Lozada et al., 2007; Araújo-Santos et al., 2021). As in other agricultural landscapes, isolated trees, that overtop olive tree canopy serve as perches for birds and provide alternative resources (food or mating and nesting sites) in the olive fields (e.g., Hoi-Leitner et al., 1999), introducing a source of heterogeneity for the patterns of frugivore activity and seed deposition. They typically have a focal effect, attracting frugivore activity within inhospitable matrices and generating seed nucleation (Herrera and García, 2009), especially if they bear fruit. We confirmed the nucleation effect of isolated perching trees, with findings revealing that, even if they did not bear fruit, they attracted seeds to the olive field. The probability of seed deposition under the canopy of isolated perching trees in the olive field, and the number and the diversity of arrived seeds, were to some extent comparable to those of habitat remnant patches and much higher than under the canopy of olive trees (Figures 4A,D,G).
Interestingly, the habitat effects in olive groves interact frequently with SNWH cover and/or olive grove cover (i.e., expansion) in landscapes. These types of interacting effects have been reported in literature on avian frugivores in landscapes fragmented by agroforest croplands (Araújo-Santos et al., 2021). While the abundance and diversity of frugivores in the seminatural patches of the olive farms varied in relation to olive grove cover in the landscape and/or across levels of SNWH (augmenting the abundance but decreasing the diversity as olive grove cover is higher and the patches of natural habitat smaller and infrequent), they remain invariantly low in the olive field (see Figures 2A,C). In turn, frugivory decreased with the olive grove cover in the landscape (Figures 3B,E). On the one hand, this means that as seminatural woodland habitat patches become smaller and infrequent in landscapes, they host fewer fruiting species (a typical patch cover effect, Arroyo-Rodríguez et al., 2020), and attract a less diverse group (fewer species) of frugivores that concentrate in the small patches still present and build less diverse fruit diets. On the other hand, these results suggest that independently of the landscape context in which olive farms are located, olive fields in their current state are of very low attractiveness, and are even inhospitable for many species of frugivores. This is unexpected since they supply them with a rewarding and abundant fruit (the cultivated olive) and their agroforest-like nature could be relatively suitable for forest/scrubland birds. These results also suggest that the spillover of avian frugivores and seeds of fruit-bearing plants from the remnant patches of natural habitat toward the olive field matrix is rare, with several species of frugivores concentrating their abundance and activity exclusively in the seminatural patches, this effect will become even more pronounced with the expansion of the olive groves and the homogenization of olive-dominated landscapes. Therefore, unlike the permeability typically found in tropical agroforest systems to animal-mediated seed dispersal (for instance, shade coffee or cocoa plantations, Lozada et al., 2007; Araújo-Santos et al., 2021), olive fields are less permeable than we initially presumed to frugivorous birds, and to their dispersal function.
In this scenario, the persistence of a diverse frugivore guild and its dispersal function in the olive grove landscapes strongly depends not only on the presence but also on the cover provided by seminatural woodland habitats. Maintenance of forest remnants has been shown to be fundamental to maintaining frugivory and seed dispersal services and potential forest regeneration in agricultural landscapes, both in temperate and Mediterranean regions and in the tropics. A number of forest remnants in such landscapes promotes local (within patch) and regional (between patches) seed dispersal by medium-sized and large avian frugivores capable of long-distance flight, which to some extent counteracts the effects of forest fragmentation on fruit consumption and removal from focal plant species (Breitbach et al., 2010; González-Varo, 2010; Parejo-Farnés et al., 2020). This ultimately should enable abundant and diverse seed rains and the persistence of species at landscape scale through long-distance seed dispersal (García et al., 2010; González-Varo et al., 2017; Araújo-Santos et al., 2021).
Impact of Local Intensive Ground Herb Cover Removal on Frugivory and Seed Dispersal
Intensification of agricultural practices may affect the abundance and diversity of farmland birds (Donald et al., 2001) and birds thriving in woody and agroforest croplands (for instance, Assandri et al., 2016, 2017, for vineyards; Rey et al., 2019; Morgado et al., 2020 in olive groves; Bennet et al., 2021, in cocoa agroforest, Araújo-Santos et al., 2021 in coffee plantations). Among the most susceptible avian guilds to intensive agricultural practices in permanent croplands are insectivores and frugivores (Rey, 2011; Martínez-Núñez et al., 2020a; Bennet et al., 2021). It has been suggested that intensive agriculture affects pest biocontrol and seed dispersal services (Johnson et al., 2010; Maas et al., 2013; Araújo-Santos et al., 2021). The different methods adopted and levels of intensification that take place in local agriculture and how they affect avian frugivory and seed deposition patterns in these croplands have only been investigated in tropical agroforest systems (Lozada et al., 2007; Araújo-Santos et al., 2021) to date, and there is no previous information on olive groves. We found that the effects of intensification of local agricultural practices to be more frequent in olive groves than we initially expected.
Different from other woody and agroforest croplands, such as cocoa and coffee agroecosystems, where intensification mainly concerns the removal of a canopy of tropical forest tree species, the form of intensification in olive groves involves the persistent application of herbicide to remove ground herbaceous cover (which are considered weeds in olive fields) (Rey et al., 2019; Martínez-Núñez et al., 2020b; Tarifa et al., 2021). We confirmed that local intensive practices of ground cover removal notably impacted the frugivory. This practice has consistent negative effects on the proportion of frugivorous species dispersing seeds, and the seed number and diversity in their droppings. This effect eventually translated to decay in the probability of seed deposition and the diversity of seed rains, especially in the olive field. Active practices of ground cover removal in intensive farms much probably hampered the occurrence of fruit species in the olive fields, affecting the frugivory.
Frugivorous birds also use insects in their diets in olive groves (Rey, 1992; Rey et al., 1996; Rey and Valera, 1999). Intensive farms more frequently use pesticides against olive pests, which together with the removal of herbaceous cover substantially decreases insect availability in intensive farms (Carpio et al., 2019) and could contribute to making these farms inhospitable for some birds. This may explain our findings, which indicate the negative (although only marginally significant) effects of local intensification practices on avian frugivore richness.
Synthesis and Applications
The olive agroecosystem plays an important role in the conservation of numerous frugivorous birds that winter in the circum-Mediterranean region and of their seed dispersal function (Rey, 2011). However, although information about their abundance and species composition is extensive, we still know very little about the consequences of agriculture on the seed dispersal service that frugivores provide. Our study was conducted at the farm, landscape, and regional levels, and suggests that olive cultivation threatens the seed dispersal service delivered by avian frugivores for many Mediterranean plant species. Importantly, all components of the seed dispersal function (abundance and diversity of avian frugivores, intensity of frugivory, and seed deposition) are threatened in most olive grove landscapes due to olive expansion and loss of woodland habitats. Moreover, this phenomenon is further aggravated by predominant intensive agricultural practices that remove ground herb covers with herbicides.
The increasing area occupied by olive grove-dominated landscapes not only threatens frugivorous/insectivorous bird communities, it also lessens their functional complementarity and diminishes the connectivity, and potential regeneration of, the persisting remnant patches of seminatural woodland habitat within these landscapes. There is a current expansion of olive groves, toward superintensive olive growing in hedges, which could further aggravate the ongoing loss of frugivore diversity and frugivory in olive groves (but see Morgado et al., 2021) due to the deep structural modification of this agroforest-like agroecosystem. These threats could be generalized to most olive growing areas of the Mediterranean Basin since land conversion to olive cultivation by intensive and superintensive practices is being pursued throughout the entire region.
Our findings have important applications for the management and recovery of the seed dispersal function. We have shown that the presence and cover of fruit-rich SNWH patches in olive grove landscapes are critical for the conservation of avian frugivores and their dispersal services and that intensive agriculture further impacts this function. Management actions should take these effects into account, and address this increase in SNWH cover, especially in fruit-rich woodlands, and undertake maintenance of ground herb cover, working at several spatial scales.
At a large scale, we recommend:
- Compulsory maintenance of woodland habitats within agricultural landscapes.
- Avoidance of property concentration (land consolidation), which frequently entails the elimination of edges, field margins, and hedgerow.
At the farm scale, the following should be prioritized:
- Maintenance of the still present woodland habitat patches within the farm, and further enriching them with native fleshy-fruited species, since many of them have already been internally cleared (through historical practices).
- Creation of new woodland patches that are rich in fleshy fruits and located in unproductive zones of the farm, especially in olive-dominated landscapes.
- Reforestation of the edges and field margins and installation of hedges rich in fleshy fruits.
- Conservation of isolated perching trees and promotion of new perches for birds by planting trees or installing artificial perches (e.g., wooden poles).
- Fomenting the maintenance of herbaceous ground cover, at least between rows of olive trees and in strips around woodland patches, and decreasing the levels of pesticide application.
These actions could be implemented within the frame of the new European Union Common Agriculture Policy (CAP post-2020), through its different environmental instruments, which include enhanced conditionality, eco-schemes, agri-environmental, and climate measures targeted specificaly and regionally to the agroecosystem (Díaz et al., 2021) of the Mediterranean region.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
This animal study was approved under licenses authorized by Bird Migration Center of Spain (dependent of SEO-BirdLife) and the Regional Government of Andalucía.
Author Contributions
PR and DG design this study and provided funding from their projects. FC, RT, CM-N, and AP conducted most field and lab work. RT, FC, and CM-N conducted the statistical analyses supervised by PR. TS carried out SIG data processing and analyses. PR, DG, RT, and FC wrote several drafts of this manuscript with feedback from CM-N. All authors critically revised the final version and give approval for its submission.
Funding
This study was funded by the projects AGRABIES (CGL2015-68963-C2 funded by MCIN/AEI/10.13039/501100011033/ FEDER Una manera de hacer Europa) and RECOVECOS (PID2019-108332GB-I00, MICIN/AEI/10.13039/501100011033). It was conducted under the general framework and associated farms of the LIFE Program project OLIVARES VIVOS (ref. LIFE14 NAT/ES/001094). CM-N received a grant as part of the FPI fellowship (BES-2016-078736) of the National Research Plan. DG was awarded a grant by the project ClaveSER (044-2019; Fundación BBVA).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are grateful to Gemma Calvo and Enrique Muñoz for their assistance in the field and our colleagues at the OLIVARES VIVOS LIFE project, Francisco Valera, J. Eugenio Gutiérrez, and Carlos Ruiz for continuous support in our research on olive groves. We are also in debt to the owners and workers at the olive farms that participated, who facilitated many aspects of the work.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.782462/full#supplementary-material
References
Alcántara, J. M., Rey, P. J., Valera, F., and Sánchez-Lafuente, A. M. (2000). Factors shaping the seedfall patternof a bird-dispersed plant. Ecology 81, 1937–1950. doi: 10.1890/0012-9658(2000)081<1937:FSTSPA>2.0.CO;2
Araújo-Santos, I., Morante-Filho, J. C., Oliveira, S., Cabral, J. P., Rocha-Santos, L., Cassano, C. R., et al. (2021). Seed rain in cocoa agroforests is induced by effects of forest loss on frugivorous birds and management intensity. Agric. Ecosyst. Environ. 313:107380. doi: 10.1016/j.agee.2021.107380
Arroyo-Rodríguez, V., Fahrig, L., Tabarelli, M., Watling, J. I., Tischendorf, L., Benchimol, L., et al. (2020). Designing optimal human-modified landscapes for forest biodiversity conservation. Ecol. Lett. 23, 1404–1420. doi: 10.1111/ele.13535
Assandri, G., Bogliani, G., Pedrini, P., and Brambilla, M. (2016). Diversity in the monotony? Habitat traits and management practices shape avian communities in intensive vineyards. Agric. Ecosyst. Environ. 223, 250–260. doi: 10.1016/j.agee.2016.03.014
Assandri, G., Bogliani, G., Pedrini, P., and Brambilla, M. (2017). Assessing common birds' ecological requirements to address nature conservation in permanent crops: lessons from Italian vineyards. J. Environ. Manag. 191, 145–154. doi: 10.1016/j.jenvman.2016.12.071
Barton, K. (2015). Package ‘MuMIn’. Multi-model inference, Ver. 1.15.6: 18. Available online at: https://cran.r-project.org/web/packages/MuMIn/index.html
Bennet, R. E., Sillett, T. E., Rice, R. A., and Marra, P. P. (2021). Impact of cocoa agricultural intensification on bird diversity and community composition. Conserv. Biol. doi: 10.1111/cobi.13779
Blanco, G., Fargallo, J. A., and Cuevas, J. A. (1994). Consumption rates of olives by choughs in central Spain. Variations and importance. J. Field Ornithol. 65, 482–489.
Breitbach, N., Laube, I., Steffan-Dewenter, I., and Böhning-Gaese, K. (2010). Bird diversity and seed dispersal along a human land-use gradient: high seed removal in structurally simple farmland. Oecologia 162, 965–976. doi: 10.1007/s00442-009-1547-y
Brooks, M. E., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R. J. 9, 378–400. doi: 10.32614/RJ-2017-066
Carpio, A. J., Castro, J., and Tortosa, F. S. (2019). Arthropod biodiversity in olive groves under two soil management systems: presence versus absence of herbaceous cover crop. Agric. For. Entomol. 21, 58–68. doi: 10.1111/afe.12303
Castro, J., Fernández-Ondoño, E., Rodríguez, C., Lallena, A. M., Sierra, M., and Aguilar, J. (2008). Effects of different olive-grove management systems on the organic carbon and nitrogen content of the soil in Jaén (Spain). Soil Tillage Res. 98, 56–67 doi: 10.1016/j.still.2007.10.002
Clough, Y., Barkmann, J., Juhrbandt, J., Kessler, M., Wanger, T. H., Anshary, A., et al. (2011). Combining high biodiversity with high yields in tropical agroforests. Proc. Natl. Acad. Sci. U.S.A. 108, 8311–8316. doi: 10.1073/pnas.1016799108
Concepción, E. D., Díaz, M., and Baquero, R. (2008). Effects of landscape complexity on the ecological effectiveness of agri-environment schemes. Landscape Ecol. 23, 135–148. doi: 10.1007/s10980-007-9150-2
De Beenhouwer, M., Aerts, R., and Honnay, O. (2013). A global meta-analysis of the biodiversity and ecosystem service benefits of coffee and cacao agroforestry. Agric. Ecosyst. Environ. 175, 1–7. doi: 10.1016/j.agee.2013.05.003
Delibes, M., Corbacho, C., Calvo, G., and Fedriani, J. M. (2012). Agriculture as matchmaker of an unexpected mutualism: great bustard disperses and enhances emergence of domestic olive seeds. Basic Appl. Ecol. 13, 125–131. doi: 10.1016/j.baae.2011.11.003
Díaz, M., Concepción, E. D., Morales, M. B., Alonso, J. C., Azcárate, F. M., Bartomeus, I., et al. (2021). Environmental objectives of Spanish agriculture: scientific guidelines for their effective implementation under the common agricultural policy 2023-2030. Ardeola 68, 445–460 doi: 10.13157/arla.68.2.2021.fo1
Donald, P. F., Gree, R. E., and Heath, M. F. (2001). Agricultural intensification and the collapse of Europe's farmland bird populations. Proc. R. Soc. B Biol. Sci. 268, 25.a−29.a. doi: 10.1098/rspb.2000.1325
Escribano-Ávila, G., Calviño-Cancela, M., Pías, B., Virgós, E., Valladares, F., and Escudero, A. (2014). Diverse guilds provide complementary dispersal services in a woodland expansion process after land abandonment. J. Appl. Ecol. 51, 1701–1711. doi: 10.1111/1365-2664.12340
European Commission (2012). “Economic analysis of the olive sector,” Directorate-General for Agriculture and Rural Development, 1–10. Available online at: http://ec.europa.eu/agriculture/olive-oil/economic-analysis_en.pdf
Foley, J. A., Defries, R., Asner, G., Barford, C., Bonan, G., Carpenter, S. R., et al. (2005). Global consequences of land use. Science 309, 570–574. doi: 10.1126/science.1111772
Foley, J. A., Ramankutty, N., Brauman, K. A., Cassidy, E. S., Gerber, J. S., Johnston, M., et al. (2011). Solutions for a cultivated planet. Nature 478, 337–342. doi: 10.1038/nature10452
Fukui, A., (2003). Relationship between seed retention time in bird's gut and fruit characteristics. Ornithol. Sci. 2, 41–48. doi: 10.2326/osj.2.41
García, D., and Chacoff, N. P. (2007). Scale-dependent effects of habitat fragmentation on hawthorn pollination, frugivory, and seed predation. Conserv. Biol. 21, 400–411. doi: 10.1111/j.1523-1739.2006.00593.x
García, D., and Martínez, D. (2012). Species richness matters for the quality of ecosystem services: a test using seed dispersal by frugivorous birds. Proc. R. Soc. B 279. 3106–3113. doi: 10.1098/rspb.2012.0175
García, D., Martínez, D., Herrera, J. M., and Morales, J. M. (2013). Functional heterogeneity in a plant–frugivore assemblage enhances seed dispersal resilience to habitat loss. Ecography 35, 1–12. doi: 10.1111/j.1600-0587.2012.07519.x
García, D., Miñarro, M., and Martínez-Sastre, R. (2018). Birds as suppliers of pest control in cider apple orchards: avian biodiversity drivers and insectivory effect. Agric. Ecosyst. Environ. 254, 233–243. doi: 10.1016/j.agee.2017.11.034
García, D., Zamora, R., and Amico, G. C. (2010). Birds as suppliers of seed dispersal in temperate ecosystems: conservation guidelines from real-world landscapes. Conserv. Biol. 24, 1070–1079. doi: 10.1111/j.1523-1739.2009.01440.x
Gómez, J. A., and Giráldez, J. V. (2009). “Erosión y degradación de suelos,” in Sostenibilidad de la Producción de Olivar en Andalucía. Junta de Andalucía, ed J. A. Gómez (Sevilla), 68–125.
Gómez, J. A., Infante-Amate, J., González de Molina, M., Vanwalleghem, T., Taguas, E. V., and Lorite, I. (2014). Olive cultivation, its impact on soil erosion and its progression into yield impacts in Southern Spain in the past as a key to a future of increasing climate uncertainty. Agriculture 4, 170–198. doi: 10.3390/agriculture4020170
Gómez, J. A., Sobrinho, T. A., Giráldez, J. V., and Fereres, E. (2009). Soil management effects on runoff, erosion and soil properties in an olive grove of Southern Spain. Soil Tillage Res. 102, 5–13. doi: 10.1016/j.still.2008.05.005
Gómez-Rosado, M., Parras-Quesada, L., Aguilera-Huertas, J., and Lozano-García, B. (2021). Soil productivity degradation in a long-term eroded olive orchard under semiarid mediterranean conditions. Agronomy 11, 812. doi: 10.3390/agronomy11040812
González-Varo, J. P. (2010). Fragmentation, habitat composition and the dispersal/predation balance in interactions between the Mediterranean myrtle and avian frugivores. Ecography. 33, 185–197 doi: 10.1111/j.1600-0587.2009.06021.x
González-Varo, J. P., Carvalho, C. S., Arroyo, J. M., and Jordano, P. (2017). Unravelling seed dispersal through fragmented landscapes: Frugivore species operate unevenly as mobile links. Mol. Ecol. 26, 4309–4321. doi: 10.1111/mec.14181
Green, R. E., Cornell, S. J., Scharlemann, J. P. W., and Balmford, A. (2005). Farming and the fate of wild nature. Science. 307, 550–555. doi: 10.1126/science.1106049
Gurr, G. M., Wratten, S. D., Landis, D. A., and You, M. (2017). Habitat management to suppress pest populations: progress and prospects. Annu. Rev. Entomol. 62, 91–109. doi: 10.1146/annurev-ento-031616-035050
Henry, M., Pons, J. M., and Cosson, J. F. (2007). Foraging behaviour of a frugivorous bat helps bridge landscape connectivity and ecological processes in a fragmented rainforest. J. Anim. Ecol. 76, 801–813. doi: 10.1111/j.1365-2656.2007.01258.x
Herrera, C. M. (1984a). A study of avian frugivores, bird-dispersed plants, and their interaction in Mediterranean scrublands. Ecol. Monogr. 54, 1–23. doi: 10.2307/1942454
Herrera, C. M. (1984b). Adaptation to frugivory of Mediterranean avian seed dispersers. Ecology 65, 609–617. doi: 10.2307/1941423
Herrera, C. M. (1985). “Habitat-consumer interactions in frugivorous birds,” in Habitat Selection in Birds, ed M. L. Cody (Orlando, FL: Academic Press), 341–365.
Herrera, C. M. (1995). Plant-vertebrate dispersal systems in the Mediterranean: ecological, evolutionary, and historical determinants. Annu. Rev. Ecol. Syst. 26, 705–727. doi: 10.1146/annurev.es.26.110195.003421
Herrera, J. M., and García, D. (2009). The role of remnant trees in seed dispersal through the matrix: being alone is not always so sad. Biol. Conserv. 142, 149–158. doi: 10.1016/j.biocon.2008.10.008
Herrera, J. M., and García, D. (2010). Effects of forest fragmentation on dispersal and seedling establishment in ornithochorous trees. Conserv. Biol. 24, 1089–1098. doi: 10.1111/j.1523-1739.2010.01459.x
Hoi-Leitner, M., Hoi, H., Romero-Pujante, M., and Valera, F. (1999), Multi-male display sites in serins (Serinus serinus L.). Ecosciencie 6, 143–147. doi: 10.1080/11956860.1999.11682514
Infante-Amate, J., Villa, I., Aguilera, E., Torremocha, E., Guzmán, G., Cid, A., et al. (2016). “The Making of Olive Landscapes in the South of Spain. A History of Continuous Expansion and Intensification,” in Biocultural Diversity in Europe. Environmental History, Vol. 5, eds M. Agnoletti and F. Emanueli (Cham: Springer).
Johnson, M. D., Kellerman, J. L., and Stercho, A. M. (2010). Pest reduction services by birds in shade and sun coffee in Jamaica. Anim. Conserv. 13, 140–147. doi: 10.1111/j.1469-1795.2009.00310.x
Jordano, P. (1982). Migrant birds are the main seed dispersers of blackberries in southern Spain. Oikos 38, 183–193. doi: 10.2307/3544018
Jordano, P. (1984). Relaciones entre plantas y aves frugívoras en el matorral mediterráneo del área de Doñana (Ph.D. Dissertation). University of Seville. Available online at: https://dialnet.unirioja.es/servlet/tesis?codigo=23313
Jordano, P. (1987). Frugivory, external morphology and digestive system in mediterranean sylviid warblers Sylvia spp. Ibis 129, 175–189. doi: 10.1111/j.1474-919X.1987.tb03199.x
Jordano, P. (1988). Diet, fruit choice and variation in body condition of frugivorous warblers in mediterranean scrubland. Ardea 76, 193–209.
Jordano, P., García, C., Godoy, J. A., and Gracía-Castaño, J. L. (2007). Differential contribution of frugivores to complex seed dispersal patterns. Proc. Natl. Acad. Sci. U.S.. 104, 3278–3282. doi: 10.1073/pnas.0606793104
Jordano, P., and Herrera, C. M. (1981). The frugivorous diet of Blackcap populations Sylvia atricapilla wintering in southern Spain. Ibis 123, 502–507. doi: 10.1111/j.1474-919X.1981.tb04055.x
Landis, D. A. (2017). Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 18, 1–12. doi: 10.1016/j.baae.2016.07.005
Lázaro, A., Mark, S., and Olesen, J. M. (2005). Bird-made fruit orchards in northern Europe: nestedness and network properties. Oikos 110, 321–329. doi: 10.1111/j.0030-1299.2005.13500.x
Letourneau, D. K., Armbrecht, I., Rivera, B. S., Lerma, J. M., Carmona, E. J., Daza, M. C., et al. (2011). Does plant diversity benefit agroecosystems? A synthetic review. Ecol. Appl. 21, 9–21. doi: 10.1890/09-2026.1
Loumou, A., and Giourga, C. (2003). Olive groves: “The life and identity of the Mediterranean”. Agric. Hum. Values 20, 87–95. doi: 10.1023/A:1022444005336
Lozada, T., de Koning, G. H. J., Marché, R., Klein, A., and Tscharntke, T. (2007). Tree recovery and seed dispersal by birds: comparing forest, agroforestry and abandoned agroforestry in coastal Ecuador. Perspect. Plant Ecol. Evolut. Syst. 8, 131–140. doi: 10.1016/j.ppees.2006.10.001
Maas, B., Clough, Y., and Tscharntke, T. (2013). Bats and birds increase crop yield in tropical agroforestry Landscapes. Ecol. Lett. 16, 1480–1487. doi: 10.1111/ele.12194
Martin, E. A., Dainese, M., Clough, Y., Báldi, A., Bommarco, R., Gagic, V., et al. (2019). The interplay of landscape composition and configuration: new pathways to manage functional biodiversity and agroecosystem services across Europe. Ecol. Lett. 22, 1083–1094. doi: 10.1111/ele.13265
Martínez, D., and García, D. (2017). Role of avian seed dispersers in tree recruitment in woodland pastures. Ecosystems. 20, 616–629. doi: 10.1007/s10021-016-0043-6
Martínez-Núñez, C., Manzaneda, A. J., Isla, J., Tarifa, R., Calvo, G., Molina, J. L., et al. (2020b). Low-intensity management benefits solitary bees in olive groves. J. Appl. Ecol. 57, 111–120. doi: 10.1111/1365-2664.13511
Martínez-Núñez, C., Rey, P. J., Manzaneda, A. J., Tarifa, R., Salido, T., Isla, J., et al. (2020a). Direct and indirect effects of agricultural practices, landscape complexity and climate on insectivorous birds, pest abundance and damage in olive groves. Agric. Ecosyst. Environ. 304:107145. doi: 10.1016/j.agee.2020.107145
McIntyre, S., and Hobbs, R. (1999). A framework for conceptualizing human effects on landscapes and its relevance to management and research models. Conserv. Biol. 13, 1282–1292. doi: 10.1046/j.1523-1739.1999.97509.x
Mellink, E., Riojas-Lopez, M. E., and Giraudoux, P. (2016). A neglected opportunity for bird conservation: the value of a perennial, semiarid agroecosystem in the Llanos de Ojuelos, central Mexico. J. Arid Environ. 124, 1–9. doi: 10.1016/j.jaridenv.2015.07.005
Morales, J. M., García, D., Martínez, D., Rodríguez-Pérez, J., and Herrera, J. M. (2013). Frugivore behavioural details matter for seed dispersal: a multi-species model for cantabrian thrushes and trees. PLoS ONE 8:e65216doi: 10.1371/journal.pone.0065216
Moreira, F., Herrera, J. M., and Beja, P. (2019). Making olive oil sustainable. Science 365, 873. doi: 10.1126/science.aay7899
Moreno, B., García-Rodríguez, S., Cañizares, R., Castro-Rodríguez, J., and Benítez, E. (2009). Rainfed olive farming in south-eastern Spain: long-term effect of soil management on biological indicators of soil quality. Agric. Ecosyst. Environ. 131, 333–339. doi: 10.1016/j.agee.2009.02.011
Morgado, R., Pedroso, R., Porto, M., Herrera, J. M., Rego, P., Moreira, F., et al. (2021). Preserving wintering frugivorous birds in agro-ecosystems under land use change: lessons from intensive and super-intensive olive orchards. J. Appl. Ecol. doi: 10.1111/1365-2664.14029
Morgado, R., Santana, J., Porto, M., Sánchez-Oliver, J. S., Reino, L., Herrera, J. M., et al. (2020). A Mediterranean silent spring? The effects of olive farming intensification on breeding bird communities. Agric. Ecosyst. Environ 288:106694. doi: 10.1016/j.agee.2019.106694
Mueller, T., Lenz, J., Caprano, T., Fiedler, W., Böhning-Gaese, K., et al. (2014). Large frugivorous birds facilitate functional connectivity of fragmented landscapes. J. Appl. Ecol. 51, 684–692. doi: 10.1111/1365-2664.12247
Muller-Landau, H. C., and Hardesty, B. D. (2005). “Seed dispersal of woody plants in tropical forests: concepts, examples and future directions” in Biotic Interactions in the Tropics: Their Role in the Maintenance of Species Diversity, eds D. F. R. P. Burslem, M. A. Pinard and S. E. Hartley (Cambridge: Cambridge University Press), 267–309.
Paredes, D., Cayuela, L., and Campos, M. (2013). Synergistic effects of ground cover and adjacent vegetation on natural enemies of olive insect pests. Agric. Ecosyst. Environ. 173, 72–80. doi: 10.1016/j.agee.2013.04.016
Parejo-Farnés, C., Aguilar, R., Herrera, J. M., Aparicio, A., and Albadalejo, R. (2020). Effects of habitat fragmentation on frugivorous birds and on seed removal from Pistacia lentiscus in two contrasting fruiting seasons. Perspect. Plant Ecol. Evol. Syst. 45:125541. doi: 10.1016/j.ppees.2020.125541
Parras-Alcántara, L., Lozano-García, B., Keesstra, S., Cerdà, A., and Brevik, E. C. (2016). Long-term effects of soil management on ecosystem services and soil loss estimation in olive grove top soils. Science of the Total Environment 571, 498–506. doi: 10.1016/j.scitotenv.2016.07.016
Pejchar, L., Pringle, R. M., Ranganathan, J., Zook, J. R., Duran, G., Oviedo, F., et al. (2008). Birds as agents of seed dispersal in a human-dominated landscape in southern Costa Rica. Biol. Conserv. 141, 536–544. doi: 10.1016/j.biocon.2007.11.008
Perea, A. J., Wiegand, T., Garrido, J. L., Rey, P. J., and Alcántara, J. M. (2021). Legacy effects of seed dispersal mechanisms shape the spatial interaction network of plant species in Mediterranean forests. J. Ecol. 109, 3670–3684. doi: 10.1111/1365-2745.13744
Perea, R., and Gutiérrez-Galán, A. (2016). Introducing cultivated trees into the wild: wood pigeons as dispersers of domestic olive seeds Acta Oecologica 71, 73–79. doi: 10.1016/j.actao.2015.09.005
Pérez-Méndez, N., Jordano, P., and Valido, A. (2017). Persisting in defaunated landscapes: reduced plant population connectivity after seed dispersal collapse. J. Ecol. 106, 936–947. doi: 10.1111/1365-2745.12848
QGIS Development Team (2018). QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online at: http://qgis.osgeo.org
Rey, P. J., (1992). Preadaptación de la avifauna frugívora invernante al cultivo del olivar (Ph. D. Thesis). Universidad de Granada.
Rey, P. J. (1993). The role of olive orchards in the wintering of frugivorous birds in Spain. Ardea 81, 151–160.
Rey, P. J. (1995). Spatio-temporal variation in fruit and frugivorous bird abundance in olive orchards. Ecology 76, 1625–1635. doi: 10.2307/1938163
Rey, P. J. (2011). Preserving frugivorous birds in agro-ecosystems: lessons from Spanish olive orchards. J. Appl. Ecol. 48, 228–237. doi: 10.1111/j.1365-2664.2010.01902.x
Rey, P. J., and Alcántara, J. M. (2000). Recruitment dynamics of a fleshy-fruited plant (Olea europaea): connecting patterns of seed dispersal to seedling establishment. J. Ecol. 88, 622–633. doi: 10.1046/j.1365-2745.2000.00472.x
Rey, P. J., and Alcántara, J. M. (2014). Effects of habitat alteration on the effectiveness of plant-avian seed dispersal mutualisms: consequences for plant regeneration. Perspect. Plant Ecol. Evol. Syst. 16, 21–31. doi: 10.1016/j.ppees.2013.11.001
Rey, P. J., Alcántara, J. M., and Sánchez-Lafuente, A. M. (1996). Temporal variation in food availability and diet of blackcaps in olive orchards. J. Field Ornithol. 67, 592–603.
Rey, P. J., and Gutiérrez, J. E. (1996). Pecking of olives by frugivorous birds: a shift in feeding behaviour to overcome gape limitation. J. Avian Biol. 27, 327–333. doi: 10.2307/3677264
Rey, P. J., and Gutiérrez, J. E. (1997). Elección de fruto y conducta de alimentación de aves frugívoras en olivares y acebuchares: una estrategia óptima basada en la razón beneficio/tiempo de manipulación. Ardeola 44, 27–39.
Rey, P. J., Gutiérrez, J. E., Alcántara, J. M., and Valera, F. (1997). Fruit size in wild olives: implications for avian seed dispersal. Funct. Ecol. 11, 611–618 doi: 10.1046/j.1365-2435.1997.00132.x
Rey, P. J., Manzaneda, A. J., Valera, F., Alcántara, J. M., Tarifa, R., Isla, J., et al. (2019). Landscape-moderated bio- diversity effects of ground herb cover in olive groves: implications for regional biodiversity conservation. Agric. Ecosyst. Environ. 277, 61–73. doi: 10.1016/j.agee.2019.03.007
Rey, P. J., and Valera, F. (1999). Diet plasticity in blackcap (Sylvia atricapilla): the ability to overcome -nutritional constraints imposed by agricultural intensification. Ecoscience 6, 429–438. doi: 10.1080/11956860.1999.11682539
Sánchez-Clavijo, L. M., Bayly, N. J., and Quintana-Ascencio, P. F. (2020). Habitat selection in transformed landscapes and the role of forest remnants and shade coffee in the conservation of resident birds. J. Anim. Ecol. 89, 553–564. doi: 10.1111/1365-2656.13108
Sánchez-Moreno, S., Castro, J, Alonso-Prados, E, Alonso-Prados, J. L., García-Baudín, J. M., Talavera, M., and Durán-Zuazo, V. H. (2015). Tillage and herbicide decrease soil biodiversity in olive orchards. Agron. Sustain. Dev. 35, 691–700. doi: 10.1007/s13593-014-0266-x
Santos, T., and Tellería, J. L. (1994). Influence of forest fragmentation on seed consumption and dispersal of spanish juniper Juniperus thurifera. Biol. Conserv. 70, 129–134. doi: 10.1016/0006-3207(94)90280-1
Sorensen, A. E. (1984). Nutrition, energy and passage time: experiments with fruit preference in European blackbirds. J. Anim. Ecol. 53, 545–557. doi: 10.2307/4533
Storchová, L., and Horak, D. (2018). Life-history characteristics of European birds. Glob. Ecol. Biogeogr. 27, 400–406. doi: 10.1111/geb.12709
Tarifa, R., Martínez-Núñez, C., Valera, F., González-Varo, J. P., Salido, T., and Rey, P. J. (2021). Agricultural intensification erodes taxonomic and functional diversity in Mediterranean olive groves by filtering out rare species. J. Appl. Ecol. doi: 10.1111/1365-2664.13970
Tellería, J. L., Ramírez, A., and Pérez-Trís, J. (2008). Fruit tracking between sites and years by birds in Mediterranean wintering grounds. Ecography 31, 381–388. doi: 10.1111/j.0906-7590.2008.05283.x
Tilman, D., Fargione, J., Wolff, B., D'Antonio, C., Dobson, A., Howarth, R., et al. (2001). Forecasting agriculturally driven global environmental change. Science 292, 281–284. doi: 10.1126/science.1057544
Tomback, D. F. (2016). “Seed dispersal by corvids: birds that build forests” in Why Birds Matter: Avian Ecological Function and Ecosystem Services, ed Ç. H. Sekercioglu, D. G. Wenny, and C. J. Whelan (Chicago: The University of Chicago Press), 196–234.
Tscharntke, T., Klein, A. M., Kruess, A., Steffan-Dewenter, I., and Thies, C. (2005). Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecol. Lett. 8, 857–874. doi: 10.1111/j.1461-0248.2005.00782.x
Tscharntke, T., Tylianakis, J. M., Rand, T. A., Didham, R. K., Fahrig, L., Batáry, P., et al. (2012). Landscape moderation of biodiversity patterns and processes - eight hypotheses. Biol. Rev. 87, 661–685. doi: 10.1111/j.1469-185X.2011.00216.x
Vilar, J., and Pereira, J. E. (2018). International Olive Growing. Worldwide Analysis and Summary. Jaén: Ed. Fundación Caja Rural de Jaén.
Keywords: seed dispersal, frugivorous bird, olive grove, agriculture intensification, landscape transformation, habitat loss, agroforest ecosystem, frugivory
Citation: Rey PJ, Camacho FM, Tarifa R, Martínez-Núñez C, Salido T, Pérez AJ and García D (2021) Persistence of Seed Dispersal in Agroecosystems: Effects of Landscape Modification and Intensive Soil Management Practices in Avian Frugivores, Frugivory and Seed Deposition in Olive Croplands. Front. Ecol. Evol. 9:782462. doi: 10.3389/fevo.2021.782462
Received: 24 September 2021; Accepted: 10 November 2021;
Published: 16 December 2021.
Edited by:
Anna Traveset, Institut Mediterrani d'Estudis Avançats, SpainReviewed by:
T. Rodd Kelsey, The Nature Conservancy, United StatesMiguel Porto, Centro de Investigacao em Biodiversidade e Recursos Geneticos (CIBIO-InBIO), Portugal
Copyright © 2021 Rey, Camacho, Tarifa, Martínez-Núñez, Salido, Pérez and García. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pedro J. Rey, prey@ujaen.es