Bioprospecting Trichoderma: A Systematic Roadmap to Screen Genomes and Natural Products for Biocontrol Applications
 Tomás A. Rush1
Tomás A. Rush1  Him K. Shrestha1,2
Him K. Shrestha1,2  Muralikrishnan Gopalakrishnan Meena3
Muralikrishnan Gopalakrishnan Meena3  Margaret K. Spangler1,2
Margaret K. Spangler1,2  J. Christopher Ellis1
J. Christopher Ellis1  Jesse L. Labbé1,2*
Jesse L. Labbé1,2*  Paul E. Abraham1,2*
Paul E. Abraham1,2*- 1Oak Ridge National Laboratory, Biosciences Division, Oak Ridge, TN, United States
- 2Graduate School of Genome Science and Technology, University of Tennessee, Knoxville, Knoxville, TN, United States
- 3Oak Ridge National Laboratory, National Center for Computational Sciences, Oak Ridge, TN, United States
Natural products derived from microbes are crucial innovations that would help in reaching sustainability development goals worldwide while achieving bioeconomic growth. Trichoderma species are well-studied model fungal organisms used for their biocontrol properties with great potential to alleviate the use of agrochemicals in agriculture. However, identifying and characterizing effective natural products in novel species or strains as biological control products remains a meticulous process with many known challenges to be navigated. Integration of recent advancements in various “omics” technologies, next generation biodesign, machine learning, and artificial intelligence approaches could greatly advance bioprospecting goals. Herein, we propose a roadmap for assessing the potential impact of already known or newly discovered Trichoderma species for biocontrol applications. By screening publicly available Trichoderma genome sequences, we first highlight the prevalence of putative biosynthetic gene clusters and antimicrobial peptides among genomes as an initial step toward predicting which organisms could increase the diversity of natural products. Next, we discuss high-throughput methods for screening organisms to discover and characterize natural products and how these findings impact both fundamental and applied research fields.
Introduction
The discovery and usage of biological controls as management strategies started with astute observations of ecological niches for studying microbial interactions (Dubos, 1939; Dubos and Cattaneo, 1939; van den Bosch et al., 1982; Dias et al., 2012; Barratt et al., 2018). Later came the rise of biological control applications due to a consortium of scientists and industry partners working on multidisciplinary ideas and projects that initially seemed unrelated yet were serving a common goal. Invasive species cost around $120 billion USD yearly in crop yield losses (Pimentel et al., 2005); plant pathogens alone, primarily fungi, result in annual crop financial losses estimated at $23.5 billion USD (including control costs) (Rossman, 2009). Even more alarming are the pesticide-resistant populations that exist, which have led to the conception of several organizations, e.g., the Fungicide Resistance Action Committee (https://www.frac.info/), and the Insecticide Resistance Action Committee (https://irac-online.org/). However, not all microbes associated with crops are harmful (Stark, 2010). In fact, beneficial microbes have become an integral component of pest management strategies to control pest populations or promote plant health (Meena et al., 2017). Factors that influence the use of beneficial microbes as biological control products are stress-induced environments, nutrient-deficient areas, and known populations of plant pathogens that can be controlled (Hayat et al., 2010; Stark, 2010; Chen et al., 2018; Begum et al., 2019; Kulimushi et al., 2021). In general, the use of biological control products is preferred for many reasons, including the reduction of pesticide use, cost-effectiveness, and its efficacy against a broad range of natural pest and support services (Bale et al., 2008; Benjamin and Wesseler, 2016; Barratt et al., 2018). Yet, biological control product applications face several challenges including invasive species stemming from the fungus used as an active ingredient; increasing crop groups, cultivars and varieties; pest complexes and resistances; incompatibility with pesticides; non-targeted effects, and risk assessment strategies (Bale et al., 2008; Barratt et al., 2018; Köhl et al., 2019). Given the complexity of these challenges, herein we propose a roadmap for bioprospecting microbes, using Trichoderma species as our model organisms. The suggested roadmap integrates predictive biology, functional genomics, high-throughput analytics, and next-generation biodesign and genome engineering approaches. Moreover, because newly discovered and characterized biological controls can have various applications, we provide a summary of important advantages and drawbacks that should be considered. Using this framework, we begin to predict which species among the already sequenced Trichoderma have unique potential as valuable biocontrol agents or source of natural products.
The Growing Market for Biological Products
Biological products including biofertilizers, biostimulants, bioherbicides, and biological control products are a multi-million-dollar industry (Bale et al., 2008; Barratt et al., 2018; van Lenteren et al., 2018) and projected to become a multi-billion-dollar industry in the next few years (Figure 1). Currently, there are few studies investigating the impact that Trichoderma or its derived natural products have on the biological control market. Although Trichoderma species represent 50–60% of the fungal biological control agents (Whipps, 2001; Verma et al., 2007), their potential market value remains uncertain. Natural product–derived drugs represent 25–50% of currently marketed drugs (Kingston, 2011) and have been the source of new drugs for nearly 40 years (Newman and Cragg, 2020). The development of biological control products was predicted to be costly; however, the rate remains reasonable compared with the cost of the synthesis, toxicological evaluation, and marketing of a new pesticide (Bale et al., 2008). The main caveat is that it can take up to 10 years to complete an extensive efficacy and non-targeted effect studies before releasing a new biological control product (Bale et al., 2008).
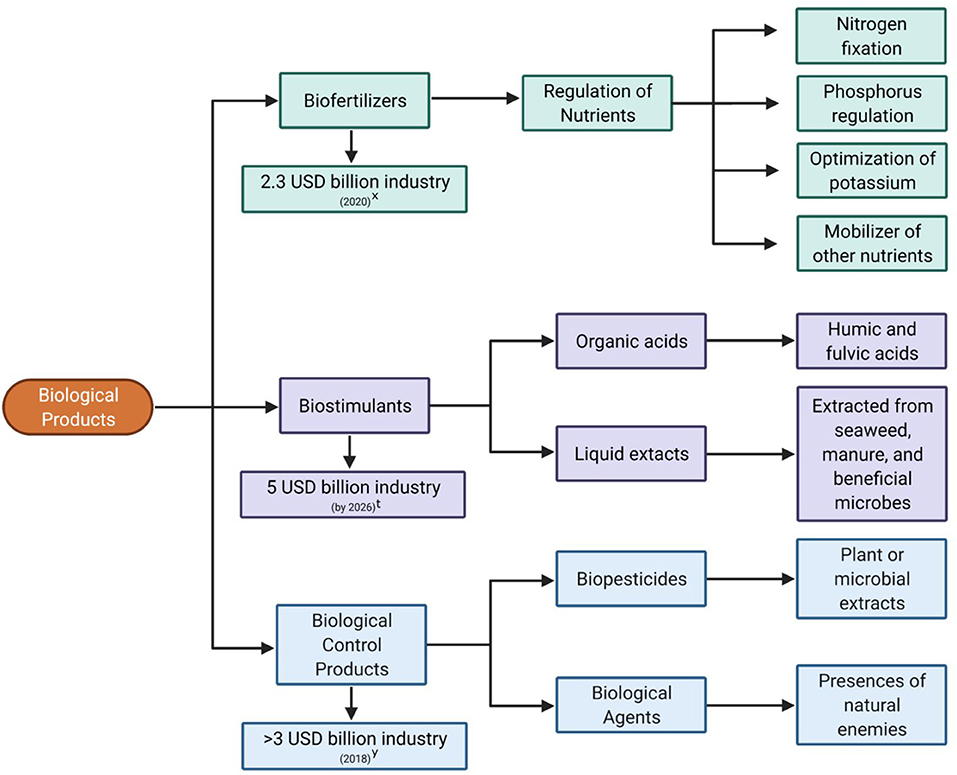
Figure 1. Summary of the various types of biological products and their market value. (x) https://markets.businessinsider.com/news/stocks/biofertilizers-market-worth-3-9-billion-by-2025-exclusive-report-by-marketsandmarkets-1029369497; (t) http://www.globenewswire.com/en/news-release/2020/12/22/2149738/0/en/Biostimulants-Market-revenue-worth-5-billion-by-2026-Says-GMI.html; (y) https://www.gminsights.com/industry-analysis/biocontrol-agents-market.
Biofertilizers consist of microorganisms that enhance the supply of adequate nutrients to crop plants (Reddy et al., 2020). Biostimulants are any substances/mixtures of natural origin or microorganisms that improve the condition of a crop without causing adverse effects (Rouphael and Colla, 2020). Biological control products comprise both biopesticides and biological control agents. Biopesticides are microbes known to produce antagonistic secreted molecules (i.e., metabolites, peptides, etc.), while biological control agents are microbes used as active ingredients owing to their ability to compete for food or space, their mycoparasitism and antibiosis capacities, or their ability to induce plant defense responses (Contreras-Cornejo et al., 2016). Characterizing the diversity of natural products among microbial protagonists, like Trichoderma, will undoubtedly add to the growing market and provide necessary resources required for a sustainable future in agriculture.
Trichoderma As a Ubiquitous Genus Worthy of Exploring for Biocontrol Product Discovery
Trichoderma are asexual, spore-producing, fungicolous ascomycete fungi that are easily isolated, culturable in substrate media, and present in nearly all soils and other diverse habitats (Harman et al., 2004; Harman, 2006; Schmoll and Schuster, 2010; Druzhinina et al., 2011; Kubicek et al., 2019; Sun et al., 2019). Trichoderma species are generalists, as they can thrive on resources provided by plants, other fungi, and animals (Kubicek et al., 2019). In some scenarios, many species of Trichoderma may act as facultative endophytes (Druzhinina et al., 2011). Trichoderma species evolved as versatile biotrophic associates that promote plant health and growth and have shown biocontrol activity against various plant pathogens (Vinale et al., 2008a,b; Lorito et al., 2010). Thus, they have become popular choices as biological control products (Howell, 2003; Benítez et al., 2004; Vinale et al., 2008a,b; Lorito et al., 2010; Kubicek et al., 2011; Kumar and Ashraf, 2017; Mukhopadhyay and Kumar, 2020). Numerous Trichoderma species are biological control agents, biofertilizers, and biostimulants, and they produce secondary metabolites with biopesticide activities (Harman et al., 2004; Harman, 2006; Vinale et al., 2008a,b; Lorito et al., 2010; Schmoll and Schuster, 2010; Druzhinina et al., 2011; Mendoza-Mendoza et al., 2018; Kubicek et al., 2019; Thambugala et al., 2020). As of July 2020, there are 375 species with valid names (Cai and Druzhinina, 2021). As of April 2021, there were 453 records of Trichoderma species found on Index Fungorum (http://www.indexfungorum.org) and 337 taxonomic species with sequences on the National Center for Biotechnology Information (NCBI), excluding confer (Cf.) names and uncharacterized isolates. However, only a handful of Trichoderma species are used for their biocontrol properties, surpassing any other fungal genera used as biological control agents. Currently, 31 fungal genera are used as active ingredients in biological control products, of which 26 belong to the Ascomycota phylum and the rest are basidiomycetes (Figure 2A). Trichoderma has by far the highest number of species used as biocontrol agents, a total of 13 (Figure 2B). A similar study showed that ascomycetes have the highest number of bioactivities against plant pathogens, and Trichoderma was the most used active ingredient (Thambugala et al., 2020).
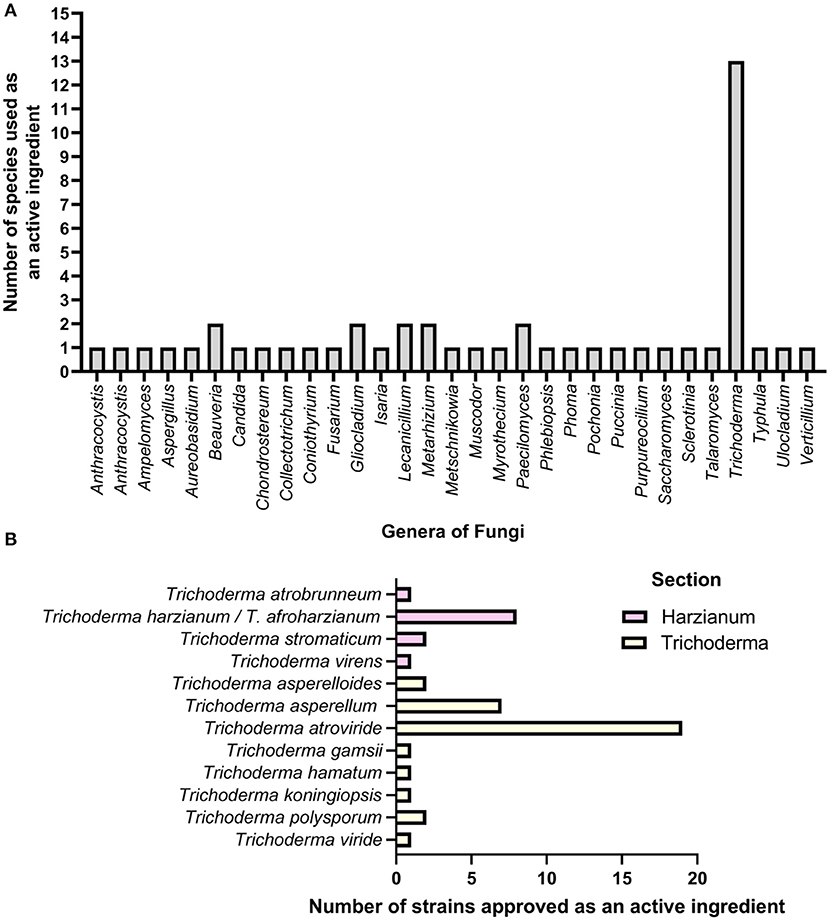
Figure 2. The application of fungi as an active ingredient in commercially available products. (A) Fungal genera used as active ingredients in commercially available products. (B) The number of Trichoderma strains approved as active ingredients in 26 commercially available pesticides produced by five separate companies. T. harzianum and T. virens were co-active ingredients in four pesticides. T. asperellum and T. gamsii were co-active ingredients in five pesticides. A combination of multiple strains of T. atroviride were used in four pesticides.
Within the species of Trichoderma, 30 strains are approved and used in current biocontrol products in Asia (Japan), European Union, North America (U.S.A.), Oceania (Australia and New Zealand), and South America (Brazil and Uruguay), as evidenced by the research conducted for this paper and the results in other literature (Kaewchai et al., 2009; van Lenteren et al., 2018; Supplementary Table 1). There are 14 root applications, 1 root and foliar application, and 2 foliar sprays available with Trichoderma species as active ingredients known to have antagonistic effects against 21 soil-borne pathogens and 7 foliar pathogens (Supplementary Table 1). This list includes only product labels that specifically indicate which Trichoderma species and strains are used.
The effects of Trichoderma species on other organisms are largely influenced by the production and secretion of metabolites, which have various established roles. Fungal metabolites have been reported to act either as communication signaling molecules between microorganisms and their hosts, or as defense agents in interactions with neighboring organisms. They were also shown to influence the development of the producing organism and to stimulate or inhibit the biosynthesis of other metabolites (Keller et al., 2005; Pusztahelyi et al., 2015; Macheleidt et al., 2016; Keller, 2019; Rokas et al., 2020). Genes responsible for the biosynthesis of secondary metabolites are often arranged into clusters (Keller, 2019). Those clusters are regulated by environmental signals and by transcriptional and epigenetics modulators (Keller, 2019). Different classes of secondary metabolites reported in fungi are indole alkaloids, non-ribosomal peptides (NRPs), polyketides, shikimic acid-derived compounds, and terpenoids (Keller et al., 2005; Pusztahelyi et al., 2015; Keller, 2019). Although Trichoderma is one of the mass producers of secondary metabolites with 23 identified families, classes, or compounds (Reino et al., 2008), and some with genetic accessibility (Schmoll and Schuster, 2010; Cardoza et al., 2011; Mukherjee et al., 2012; Keswani et al., 2014; Contreras-Cornejo et al., 2016; Zeilinger et al., 2016; Keller, 2019; Li et al., 2019; Vicente et al., 2020), little is known about the biosynthetic gene clusters responsible for the production of those metabolites. Moreover, the level of diversity among secondary metabolites produced across known Trichoderma species is still largely indefinite (Kubicek et al., 2019).
Methods
To provide a holistic view of metabolites produced by Trichoderma species, we summarized data provided by three recent reviews. An estimated 440 different molecules/non-volatile compounds/metabolites were identified and characterized from Trichoderma species (Keswani et al., 2014; Contreras-Cornejo et al., 2016; Li et al., 2019). The activities of those 440 projected compounds, reported in reviews by Keswani et al. (2014), Contreras-Cornejo et al. (2016), and Li et al. (2019), are assembled in Figure 3. If a compound has no activity reported, it is categorized as an “unknown function.” Compounds that function as biofertilizers or biostimulants are in the category “promoting plant growth or development.” Compounds with medical implications, such as being anti-tumor, anti-cancer, and so on, are placed in the “therapeutics” category. Other compounds are categorized based on their functions ascribed in previous reports. Species names were validated based on accepted Trichoderma names (Zhu and Zhuang, 2015; Li J. et al., 2018; Cai and Druzhinina, 2021) and Index Fungorum's current nomenclature (http://www.indexfungorum.org/) as of April 2021. The taxonomy of Trichoderma is cumbersome and there are multiple discrepancies in the nomenclature at the species level, thus leading to incorrect identifications of strains (Cai and Druzhinina, 2021). Trichoderma has infrageneric groups that are divided into sections or groups. Sections were assigned based on previous phylogenies data of Trichoderma (Druzhinina and Kubicek, 2005; Zhu and Zhuang, 2015; Li J. et al., 2018; Cai and Druzhinina, 2021). Pachybasium is paraphyletic (Druzhinina and Kubicek, 2005), and whether it is a section or group is indistinct. Therefore, all species belonging to Pachybasium “clade A” or “clade B” were placed in “Pachybasium.” Finally, T. harzianum strain T22, one of the commonly used active ingredient in commercial products, was determined to be T. afroharzianum (Chaverri et al., 2015; Cai and Druzhinina, 2021), the causal agent of Trichoderma ear rot on maize (Pfordt et al., 2020). Both species are part of the T. harzianum species complex (Chaverri et al., 2015). However, strain T22 still is reported in commercial products as T. harzianum or in the literature as T. harzianum or T. afroharzianum. Without further bioinformatic investigation, it is difficult to know of the reported strains used in commercial product or previous publications (other than strain T22) with biocontrol activity is T. harzianum or T. afroharzianum, so we used the term “Trichoderma harzianum/T. afroharzianum” in our analysis for Figures 2, 3 and Supplementary Table 1. However, for the antiSMASH and amPEPpy results coupled with the graph theoretic analysis, we used the genome of T. harzianum strain CBS 226.95 which is the neotype (Rifai, 1969; Druzhinina et al., 2018) and T. afroharzianum strain BFE349 (Landeis and Schmidt-Heydt, 2021).
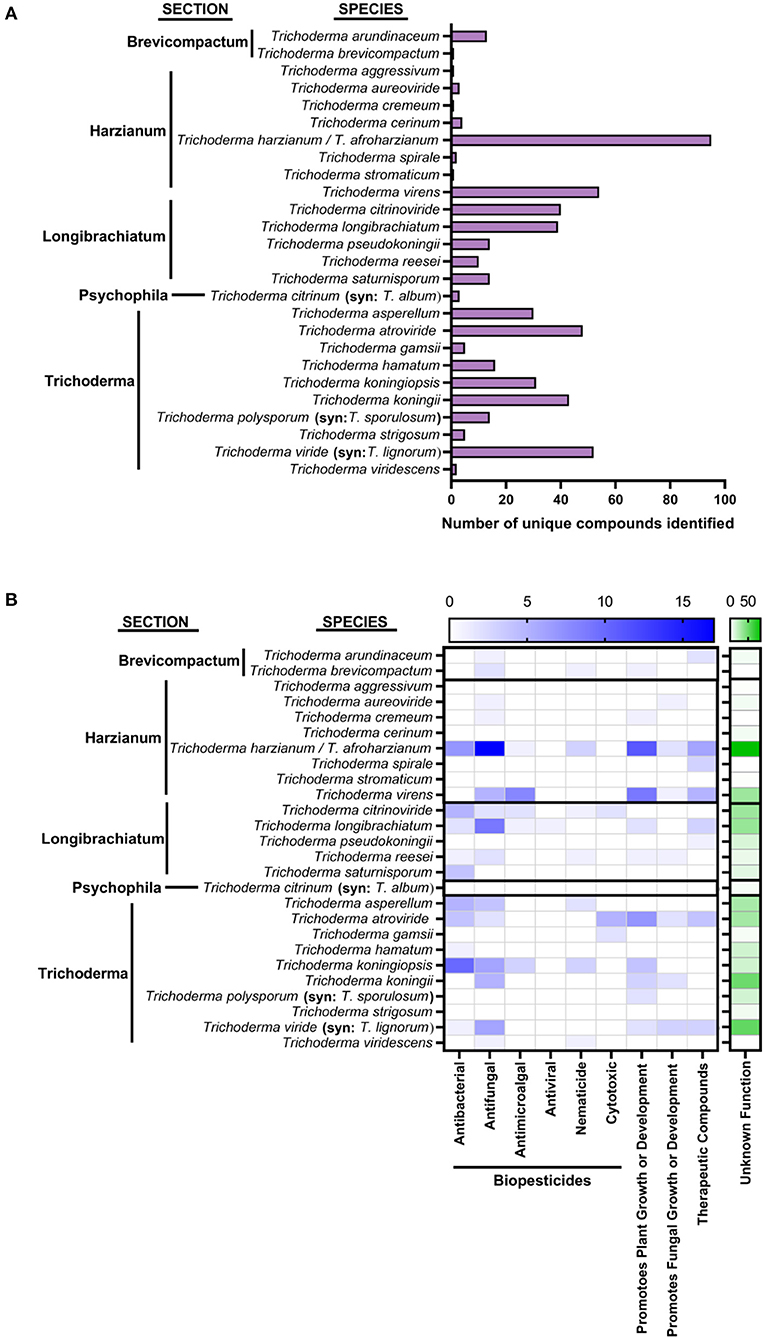
Figure 3. Number of known compounds identified in Trichoderma species (A) Species of Trichoderma organized by section, showing the number of unique compounds identified. (B) Their bioactivity refers to metabolites' activities. Metabolites with antibacterial, antifungal, antimicroalgal, antiviral, nematicide, and cytotoxic properties could be used as biopesticides. Metabolites that promote plant growth or development could be used as biofertilizers or biostimulants. Metabolites that promote the fungus' own growth and development have no assigned category. Therapeutic compounds have been identified for their potential use in the medical industry.
Results
Collectively, 27 described species of Trichoderma were reported to produce multiple compounds that were identified and characterized as (1) an enzyme that is part of a pathway to produce a secondary metabolite, (2) a described secondary metabolite, or (3) a compound with an unknown function as shown in Figure 3. Multiple species shared several of these compounds. Of these identified metabolites, several have been shown to have antibacterial or antifungal properties both in vitro and in vivo (Keswani et al., 2014; Contreras-Cornejo et al., 2016; Li et al., 2019); yet, most of these molecules' specific activity remains unknown (Contreras-Cornejo et al., 2016; Figure 3B). Many of these compounds are intermediates in the biosynthetic pathways of several Trichoderma species secondary metabolites that have agricultural and medical applications. Finally, there are biofumigants with antimicrobial and/or plant growth-promoting properties that are volatile organic compounds, as reviewed by Lee et al. (2016), Li N. et al. (2018), and Guo et al. (2019). However, they are not listed in Figure 3.
As shown in Figure 3A, the sections Harzianum, Trichoderma, and Longibrachiatum have the most identified compounds as of 2019. Harzianum has the most described functions for these identified compounds. Most of these compounds fall into the category biopesticides (as stated previously, Figure 3B). However, to our knowledge, none of these compounds have been used as active ingredients in commercial biopesticides.
Not surprisingly, the top five compounds frequently found, as shown in Figure 3, are mostly shared among various Trichoderma species (Reino et al., 2008). These are Koninginins, Trichorovins, Trichokonins, 6-Pentyl-2H-pyran-2-one, and Trichocaranes. Koninginins have structural similarities to compounds like flavonoids and vitamin E. They can inhibit phospholipase A2 (PLA2) and have been shown to have mycotoxic capabilities (Souza et al., 2008) as well as have antimicrobial properties (Reino et al., 2008). Koninginins have been found in T. koningii, T. aureoviride, T. harzianum, T. kongingiopsis, and a brown mutant of T. viride exposed to ultraviolet light (Reino et al., 2008). Recently they were identified from Phomopsis stipata (Biasetto et al., 2020). Trichorovins is an 11-residue peptaibol, originally described from T. viride (Fujita et al., 1994), that forms voltage-dependent and cation-selective ion channels in planar lipid bilayer membranes (Wada et al., 1996). Trichorovins have been found in Trichoderma longibrachiatum; T. lixii; T. harzianum, and T. viride. Trichokonins were described from T. koningii (Huang et al., 1995) and are broad-spectrum antimicrobial peptaibols with bioactivity over a wide pH and temperature range. They have no loss of activity even after autoclaving and are insensitive to proteolytic enzymes (Xiao-Yan et al., 2006). Trichokonins were found in T. koningii, T. longibrachiatum, and T. pseudokoningii. The category 6-Pentyl-2H-pyran-2-one are 2-pyranones, antifungal agents, with phytotoxic activity and antagonistic effects against multiple pathogenic fungi (Reino et al., 2008). This compound gives Trichoderma spp. a coconut aroma (Reino et al., 2008). They have been found in T. atroviride, T. harzianum, T. koningii, T. viride, T. viridescens, T. asperellum, and peaches (Parker et al., 1997; El-Sayed et al., 2014). Finally, Trichocaranes are metabolites with carotene skeletons that inhibit the growth of etiolated wheat coleoptiles (Macias et al., 2000). They appear to be unique to T. virens.
Besides secondary metabolites, antimicrobial peptides (AMPs) are another resource for biological products. AMPs, a cell defense mechanism produced by many organisms, are short and generally positively charged peptides that can directly kill microbial pathogens by modulating the host defense system (Mahlapuu et al., 2016; De Cesare et al., 2020). There has been increased AMP research over the years because of concerns regarding the advent of a “post-antibiotic era” (Mahlapuu et al., 2016). In addition, bacterial resistance to AMPs has been shown to be low or potentially negligible (Spohn et al., 2019). To date, there are more than 3,000 characterized AMPs based on their source, activity, structural characteristics, and amino acid composition (Wang et al., 2016; Huan et al., 2020). Many AMPs interact with membranes, causing cell wall inhibition and nucleic acid binding (De Cesare et al., 2020). Among other types, Trichoderma has a unique class of AMPs called peptaibols that include rare amino acids in their sequences, which provide resistance to the host or pathogen proteases and induce programmed cell death in plant fungal pathogens (Montesinos, 2007; Shi et al., 2012; Arinbasarova et al., 2017; Dotson et al., 2018; De Cesare et al., 2020; Sood et al., 2020). While the discovery of AMPs is not new, recent technological and computational advancements are expected to improve their classification, exploration, and characterization (De Cesare et al., 2020; Huan et al., 2020).
Roadmap
As previously mentioned by Bale et al. (2008), it can be as much as 10 years before a newly discovered biological control agent is released. Therefore, we provide a roadmap to guide researchers with a thorough experimental plan to discover a novel product and implement it into the market. Our roadmap details two starting points, an omics road or biodesign road, to predict and identify putative natural products. Using reference genomes, the omics road queries candidate species for predicted backbone enzymes, putative metabolites, or annotated proteins relevant to biocontrol. Given the dynamic nature of genome expression, computational approaches, like machine-learning or graph theoretical methods, benefit greatly from the addition of functional genomics data (e.g., transcriptomics, proteomics, and metabolomics). In parallel or separately, the challenges of linking predictable gene clusters to their corresponding compounds (Kenshole et al., 2021) is addressed by following the biodesign road to extract putative metabolites, isolate them, and test for bioactivity. Both roads merge at the implementation step, where the metabolite characterized for specific bioactivity can be used as a biological control product. The implementation will determine the compound, its bioactivity, the gene(s) or biosynthetic pathways responsible for its production, and its potential use in a greenhouse or field setting. Collectively, the roadmap provides insightful experimental planning that might allow for faster approval of a novel biological control product into the market (Figure 10).
Genomic Prediction Tools Highlight Natural Product Diversity Across Trichoderma Species
Advancements in high-throughput sequencing technologies have greatly reduced the cost of genotyping organisms, which has expanded genomic libraries for numerous fungi. With increased fungal genome data now available, advances in computing and bioinformatic algorithms are improving our understanding of fungal biology and evolution (Ma and Fedorova, 2010; Aguilar-Pontes et al., 2014; Grigoriev et al., 2014; Stajich, 2017) and new machine learning-based gene prediction tools are beginning to address the problem of gene function discovery (Chavali and Rhee, 2018; Mahood et al., 2020).
When using genomic prediction tools, it's important to note that several genes, which are often clustered together on the chromosomes, can be required for the biosynthesis of a single compound (Smedsgaard and Nielsen, 2005). Moreover, many gene clusters associated with the production of natural products respond to specific stimuli, such as environmental cues, nutrients, signaling compounds, or other stress factors (Hertweck, 2009) to become active (Gupta et al., 2014; Khan et al., 2020) (Figure 4). Hence, genomic predictions are often complemented with other omics measurements (e.g., transcriptome and proteome) to better understand what biological conditions are required for production of select natural products.
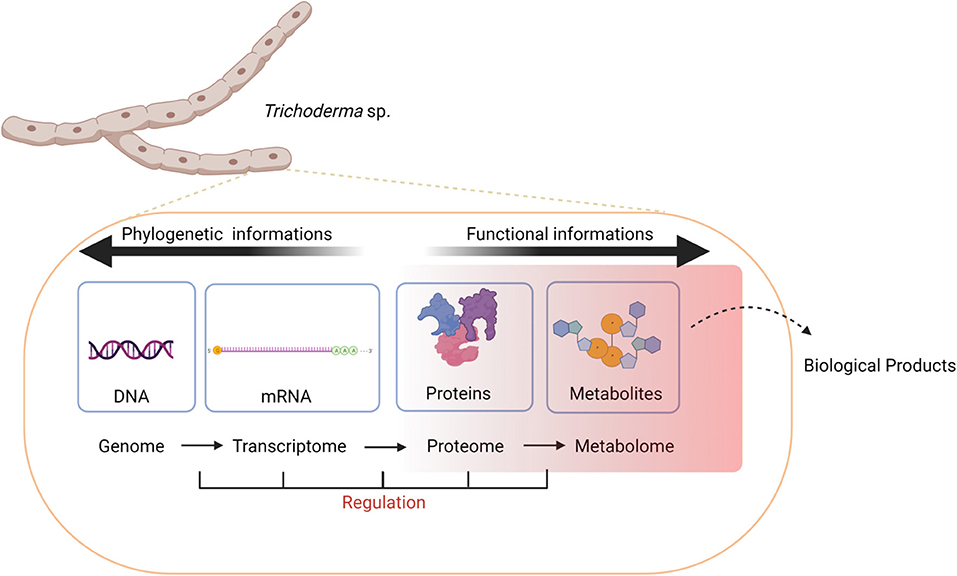
Figure 4. Natural products are the result of layers of regulation transcending the genome, transcriptome, proteome, and metabolome levels that are altered in response to changes in endogenous or environmental cues.
As of April of 2021, there were 25 species of Trichoderma available for download from the NCBI database (ncbi.nlm.nih.gov; Table 1). Trichoderma cyanodichotomus, T. longibrachiatum, T. parareesei, and T. reesei are type specimens; T. asperellum and T. oligosporum are holotype specimens; and T. harzianum is a neotype specimen. It is worth mentioning that the species T. atrobrunneum strain ITEM 908 and T. atroviride strain IMI 206040 are registered active ingredients in biological products (Supplementary Table 1). The rest of the sequenced strains are characterized species with no assigned type.
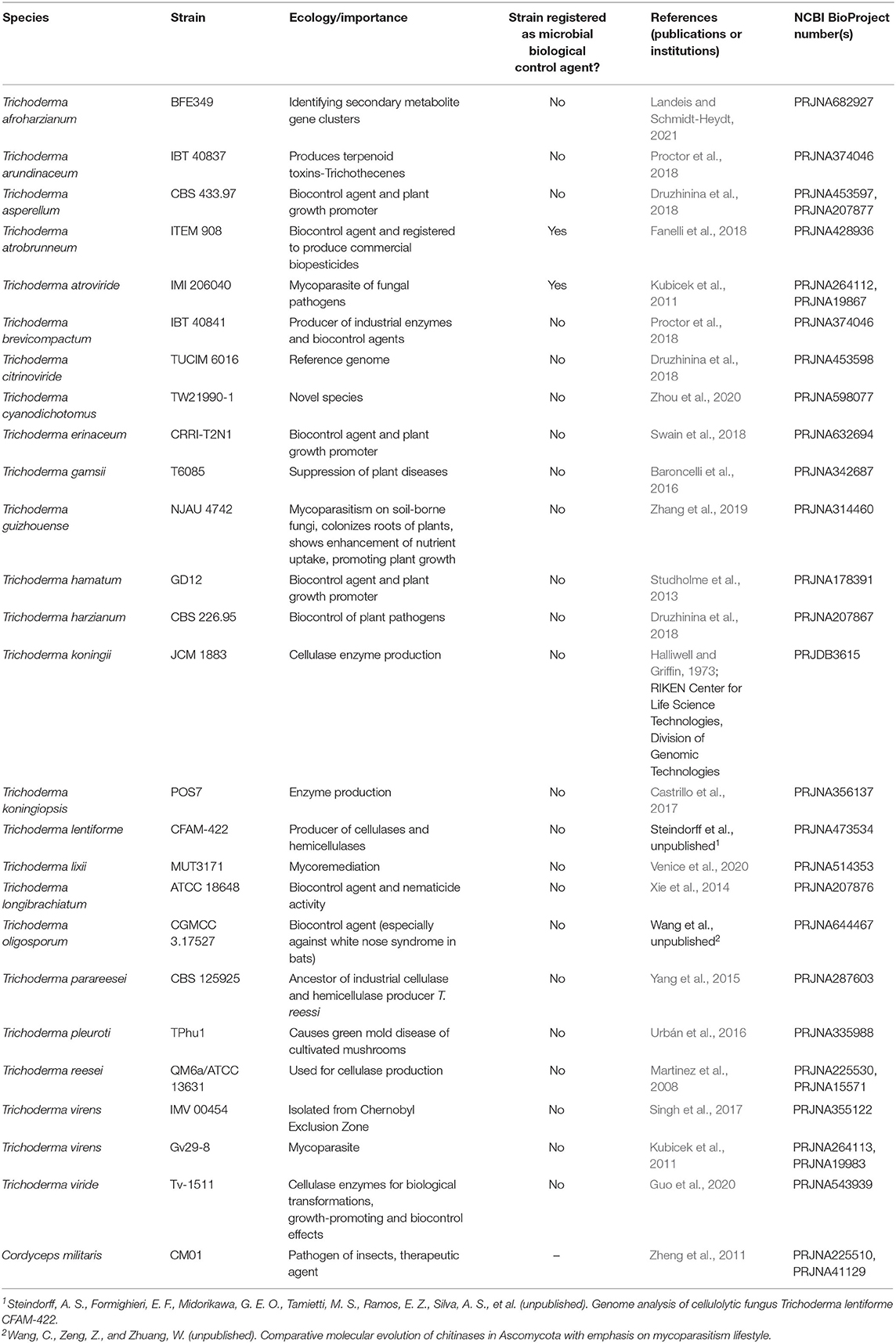
Table 1. List of Trichoderma species examined in this study.
Herein, we used machine-learning tools to examine the diversity of predicted natural products across available Trichoderma species' genomes and graph theory to interpret and qualify the results to highlight organisms having intriguing potential for natural product discovery. Currently, there are several tools available to mine genomic data for the presence of biosynthetic pathways associated with the production of natural product, which are often referred to as secondary metabolites (Fedorova et al., 2012; Chavali and Rhee, 2018), and the origin of specialized molecules like antimicrobial peptides (Xu et al., 2021). Using the widely-used antiSMASH (https://antismash.secondarymetabolites.org/#!/start) tool, we analyzed each publicly available Trichoderma genome for genome-wide identification, annotation, and analysis of secondary metabolite biosynthetic gene clusters (BGCs) (Blin et al., 2019). Additionally, each genome was interrogated by the software tool amPEPpy, which uses a random forest classifier to predict putative antimicrobial peptides based on protein sequence characteristics often attributed to antimicrobials (e.g., small open reading frames, positively charged, etc.) (Lawrence et al., 2020).
Secondary Metabolite Exploration Through Genomic Mining and Computational Analysis
Using antiSMASH v5.0, we estimated the total number of backbone enzymes and putative metabolites predicted for each genome, as shown in the heatmap generated in GraphPad Prism v 9.1.0 (221) (Figure 5). The backbone enzyme is the first enzyme in the metabolic pathway that catalyze the synthesis of the core structure, which later undergo several modifications by tailoring enzymes leading to the final product. We have organized the heatmap data (Figure 5) according to the circular phylogeny (Figure 6) produced based on genomes with protein sequences available and based on previous publications for taxonomic resolution within the Trichoderma genus for species not included in our phylogenic analysis. However, the taxonomic resolution for each section within Trichoderma is an ongoing investigation (Druzhinina and Kubicek, 2005; Cai and Druzhinina, 2021).
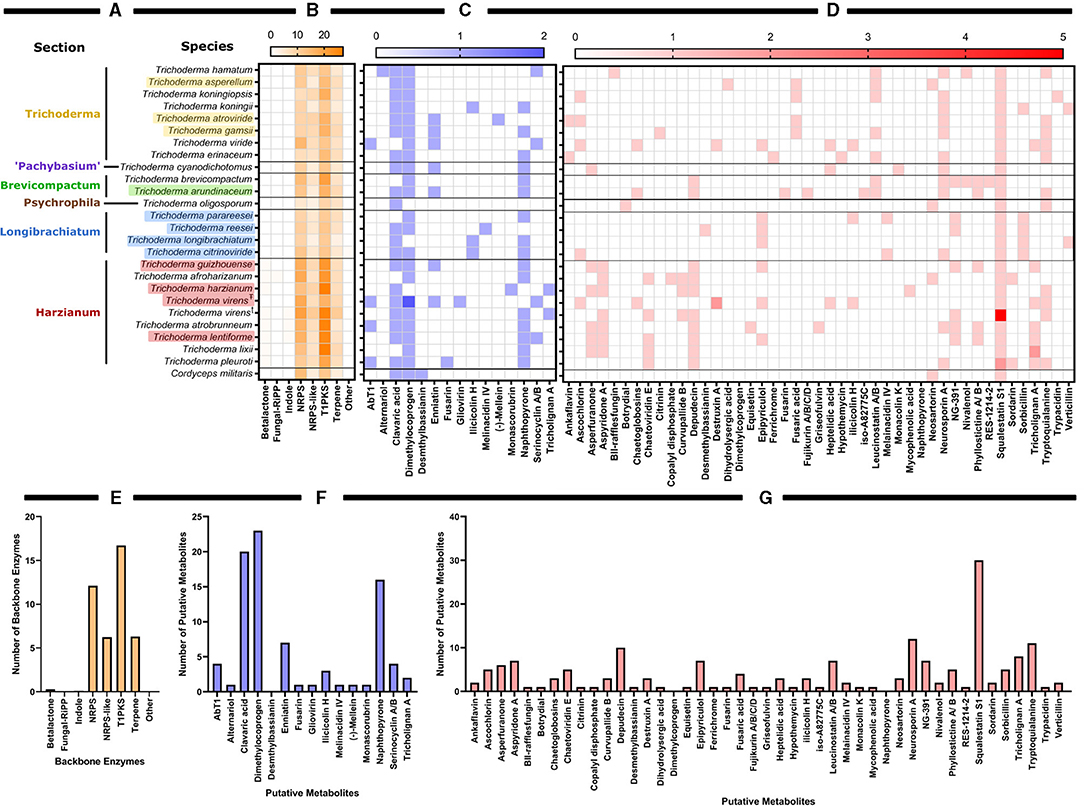
Figure 5. Predictive backbone enzymes an putative metabolites identified by antiSMASH for the available whole genomes of Trichoderma species. (A) Nucleotide genomes of Trichoderma spp. organized by their sections as determined by protein-based genome alignment as shown in Figure 6 and corresponding published data from Druzhinina and Kubicek (2005); Zhu and Zhuang (2015); Li J. et al. (2018), and Cai and Druzhinina (2021). Colored species names correspond with Figure 6. Black text species names correspond with the previous publications on Trichoderma taxonomy. Genomes are aligned with corresponding heat map showing (B) the number of backbone enzymes and (C) putative metabolites with a ≥75% or (D) <75% sequence match with known metabolites. (E) The total number of backbone enzymes found in all species examined. (F) The total number of putative metabolites with a sequence match ≥75% with known metabolites found in all species examined. (G) The total number of putative metabolites with a sequence match <75% with known metabolites found in all species examined. Strains of T. virens noted as (T) are strain Gv29-8 and as (Ŧ) are strain IMV 00454.
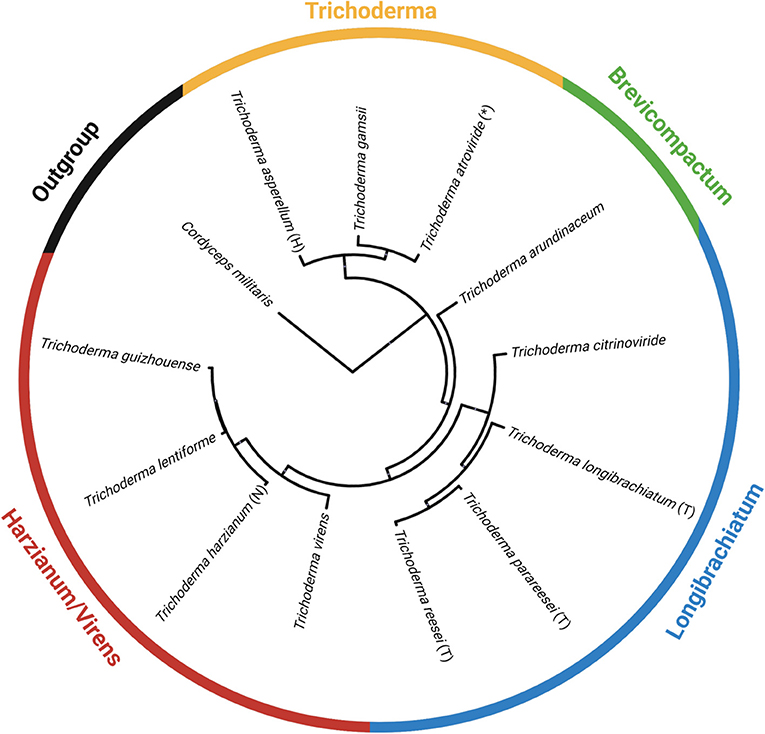
Figure 6. A circular phylogeny of Trichoderma spp. based on genomes available on NCBI as of April 2021. The phylogenetic tree is constructed from the identified orthologous protein groups from each Trichoderma spp. and one outgroup, Cordyceps militaris. The tree was exported and visualized in the interactive Tree of Life—iTOL (https://itol.embl.de/). An asterisk (*) means this strain is used as an active ingredient in commercially available biocontrol products. (T) is the type species; (H) is the holotype; and (N) is the neotype. Data are comparable to the chronogram orthologous protein alignment shown in Kubicek et al. (2019), with T. arundinaceum, a Brevicompactum representative as described in Degenkolb et al. (2008a).
Based on Trichoderma comparative genomic screening, the Harzianum section has the highest number of backbone enzymes and putative metabolites predicted in those species. However, in commercial biocontrol products, Trichoderma is better represented than Harzianum, despite having fewer predicted backbone enzymes and putative metabolites (as shown in Figure 2B). Type 1 polyketide synthases (T1PKSs) followed by NRP synthetases (NRPSs), were the backbone enzymes most often predicted in Trichoderma. NRP is a type of antimicrobial peptide that will be further discussed in this review. This group of metabolites includes peptaibols, which are linear peptides containing between 7 and 20 amino acid residues. In general, polyketides are poorly investigated as biocontrol agents (Daguerre et al., 2017). Specifically, T1PKS have rarely been examined in Trichoderma and are proposed to be mostly orthologous groups (Baker et al., 2012). To further investigate these highly anticipated backbone enzymes, machine-learning tools like the DDAP database for screening of biosynthetic T1PKS pathways (https://tylii.github.io/ddap/) can be used. Additionally, NRP discovery and the comprehensive peptaibiotics database can be applied to uncover the identities of these NRPs in Trichoderma (Caboche et al., 2008; Stoppacher et al., 2013; Neumann et al., 2015; Flissi et al., 2020). Finally, the data shown in Figure 5 are comparable to those from a previous review in which polyketides, NRPs, and terpenes were largely represented in the genomes of T. reesei, T. atroviride, and T. virens (Zeilinger et al., 2016).
The top identified putative metabolites with sequence matches ≥75% were dimethyl bassianin (23 matches), clavaric acid (20 matches), napththopyrone (16 matches), enniatin (7 matches), and AbT1 and Serinocyclin A/B with 4 matches each. Dimethyl bassianin (PubChem name is Bassianin) is a pyridine that provides yellow pigment (Lagashetti et al., 2019). Clavaric acid is a triterpenoidal inhibitor with antitumor and antionocogenic activities (Jayasuriya et al., 1998). Napththopyrone is an aromatic polyketide with cytotoxic, antitumor, antimicrobial, and tyrosine kinase properties (Bokesch et al., 2010; Lu et al., 2014; Venice et al., 2020). It has been shown to have a moderate ability to inhibit multidrug transporters and is a pigment that protects fungi from a wide range of predators (Bokesch et al., 2010; Lu et al., 2014; Venice et al., 2020). Enniatin is a mycotoxin discovered in Fusarium. Enniatin B was reported to have antifungal activity toward T. afroharzianum strain T22 (Meca et al., 2010). AbT1 is a precursor of cyclic peptide antibiotic Aureobasidin A from the crucial industrial yeast, Aureobasidium pullulans (Slightom et al., 2009). Serinocyclin A/B is a cyclic heptapeptide reported to cause a sublethal locomotory defect in mosquito larvae (Krasnoff et al., 2007). The top five putative metabolites with sequence matches <75% were squalestatin S1 (syn: zaragozic acid, 30 matches), neurosporin A (12 matches), tryptoquialanine (11 matches), depudecin (10 matches), and tricholignan A (8 matches). Squalestatin S1 controls cholesterol biosynthesis. It targets squalene synthases and has a broad spectrum of antifungal properties (Bonsch et al., 2016; Lebe and Cox, 2019). Neurosporin A is involved in fungal sexual development, provides chemo-resistance to arthropod predation, and demonstrates significant insecticidal activity (Zhao et al., 2017). Tryptoquialanine is an indole alkaloid and a tremorgenic mycotoxin that can elicit intermittent or sustained tremors in vertebrates (Gao et al., 2011). Depudecin is a small linear polyketide known to be a histone deacetylase inhibitor. Moreover, this compound contributes to pathogenesis, has antiprotozoal activity, and provides a more significant fitness benefit (Reynolds et al., 2017). Tricholignan A is a redox-active ortho-hydroquinone that facilitates reductive iron assimilation and plays a potential role in promoting plant growth under iron-deficient conditions (Chen et al., 2019).
A list of the putative metabolites predicted in the genomes of Trichoderma species is presented in Table 2. A caveat for associating putative or known metabolites with Trichoderma species in Table 2 is that numerous strains have been misidentified at the species level for several reasons: taxonomic identification is missing, non-specific analytical methods were used, species no longer exist in culture, or they do not have DNA sequences available (Nielsen et al., 2005; Degenkolb et al., 2008b; Tijerino et al., 2011; Cai and Druzhinina, 2021). Altogether, this caveat has been problematic for predictive metabolomics in Trichoderma. Those putative metabolites, excluding ferrichrome, were described in 31 separate genera, of which 30 belong to the Ascomycota and 1 belongs to the Basidiomycota. Ferrichrome is found in numerous ascomycete and basidiomycete fungi (Haas, 2014). Considering that Trichoderma is an ascomycete fungus, one can speculate that these predicted putative metabolites found in other ascomycete fungi could be produced by Trichoderma species as well, particularly since they all share a most recent common ancestor (Schoch et al., 2009; Spatafora et al., 2017). Moreover, horizontal gene transfer of secondary metabolite gene clusters has been demonstrated between ascomycete species (Kroken et al., 2003; Patron et al., 2007; Khaldi et al., 2008; Cardoza et al., 2011; Slot and Rokas, 2011; Sieber et al., 2014; Dhillon et al., 2015; Tran et al., 2019; Rokas et al., 2020) and between ascomycete species and other microbes (Schmitt and Lumbsch, 2009; Lawrence et al., 2011). To investigate these putative metabolites in Trichoderma, it should be considered that sometimes antiSMASH does not accurately predict gene cluster borders. Therefore, researchers should consider looking at some neighboring genes when performing knockout or overexpression studies to characterize a biosynthetic gene cluster. Besides antiSMASH, there are other computational programs for secondary metabolite gene mining that are less commonly used, e.g., SMURF and ClusterFinder (Tran et al., 2019). Using other available programs for comparison might be useful to obtain more accurate biosynthetic gene cluster (BGC) border predictions. Finally, inconsistency in the nomenclature of metabolites in different biochemical databases used for genome-scale metabolic screening occurs and should be considered in the experimental design (Pham et al., 2019).
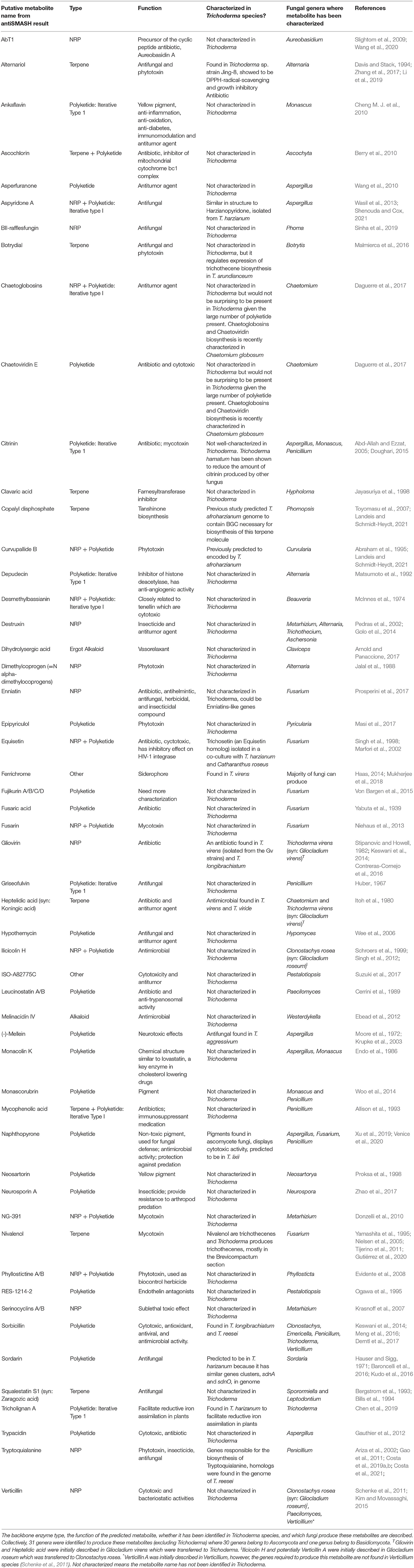
Table 2. A descriptive list of putative metabolites predicted from antiSMASH based on screening Trichoderma genomes.
To quantify the uniqueness and predictive diversity of natural products across the Trichoderma sections or functional categories of secondary metabolites, we used graph theory as a robust mathematical platform to represent the relationships between or influence of antiSMASH results across the different Trichoderma sections. In graph theory analysis, a graph is comprised of nodes and edges (Newman, 2018) and, in this context, the sections and backbone enzymes represent the nodes, and the edges are weighted by the average number of times a category is observed by all the species in a section (Figure 7A). We used two different graph-theoretic measures to quantify and rank the importance of sections and backbone enzymes. These are (1) the strength and (2) PageRank of the nodes. The strength of the node is determined by the summation of all the edges from (out-strength) or to (in-strength) a node, representing the same values reported previously in Figure 5E. The PageRank measure (Page and Brin, 1998), which is the underlying method used by Google search engine to rank web pages based on the links between web pages, quantifies the relative importance of a node (section or backbone enzyme) based on the connections it has. We rank the sections and the enzymes for being the most influential and influenced nodes, respectively, using the directed PageRank measures broadcasting and receiving measures (Grindrod et al., 2011) as shown in Figures 7B,C. These values are min-max normalized (between 0 and 1). The above graph-theoretic analysis was performed separately for the interaction among the sections and putative metabolites with ≥75% and <75% matched sequences with known metabolites. We only report the min-max normalized broadcasting and receiving PageRank values for these analyses, as shown in Figures 7D–G.
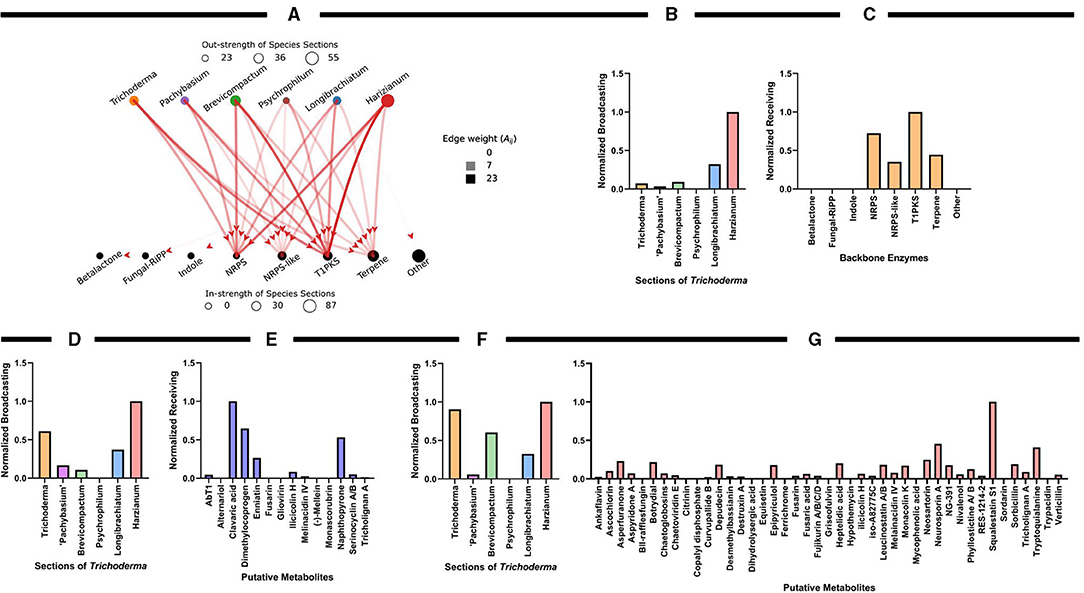
Figure 7. Graph theoretical analysis to characterize influential species sections of Trichoderma for identifying putative compounds or metabolites. (A) The weighted-directed graph representation of interactions among the sections and backbone enzymes. The edge transparency represents the average number of times an enzyme is identified by a section and the node size (circle radius) represents the total interaction strength of the sections and enzymes. (B,C) PageRank centrality measures for the sections and backbone enzymes, quantified by the min-max normalized (values between 0 and 1) broadcasting and receiving values, respectively. (D,E) Normalized broadcasting and receiving PageRank measures computed using the interactions among the sections and putative metabolites with a ≥75% match sequence; (F,G) represents the same for <75% matched sequences with known metabolites.
Overall, the graph theory analysis highlighted notable interactions between organisms and secondary metabolite predictions that can assist with selecting organisms or natural products for further analyses. Interestingly, the Harzianum section has the most prevalence of predicted backbone enzymes and putative metabolites compared to all the other sections examined. This data supports what has been observed in several publications in identifying bioactive compounds (Figure 3) and why several biological products use species from this section are used as active ingredients (Figure 2B). The Trichoderma section showed a higher prediction of putative metabolites found in their genomes (Figures 7D,F), which may explain why species from the Trichoderma section have the most used active ingredient in commercial agricultural products (Figure 2B). While the Longibrachiatum section was shown to have a relatively high number of backbone enzymes found in their genomes (Figure 7B), species from Longibrachiatum are not listed as active ingredients in commercial agricultural labels (Figure 2B). It is interesting, though, that three well-studied species, T. reesei, T. parareesei, and T. longibrachiatum (Supplementary Table 1) are used in other industries. For example, T. reesei strain DSM 32338 was genetically modified to produce muramidase enzyme used as an additive for chickens and other minor poultry for fattening (Rychen et al., 2018). Trichoderma reesei is often genetically manipulated to overexpressed genes responsible for cellulases that are then used in the biotechnology industries (Druzhinina and Kubicek, 2017; Hinterdobler et al., 2021). Trichoderma reesei has been reported to sexually reproduce which can provide tools for fast and improved development of strains for industrial use (Seidl et al., 2009). However, there can be a pitfall in releasing these species in the environment as an active ingredient because it has the potential to sexually recombine with a native population. But alternatives like the sympatric species, T. longibrachiatum, and T. parareesei have a clonal lifestyle (Druzhinina et al., 2008, 2010; Atanasova et al., 2010), therefore could be suitable species to use as biological control products. Trichoderma parareesei have been shown to produce cellulases like, T. reesei, and have biostimulant activities by increasing seedling lateral root development on tomatoes and produced antagonistic effects against fungal foliar pathogen, Botrytis cinerea (Rubio et al., 2014). Several publications show the effects of T. longibrachiatum as a fungicide and nematicide on several crops, yet the efficacy is unknown (Migheli et al., 1998; Rojo et al., 2007; Zhang et al., 2015, 2017, 2018b). Our data suggest that by using genome-mining companioned with computational analysis, there is evidence of several known Trichoderma species that have not been fully explored for their use as a biological product in agriculture. This untapped resource could be part of a solution to overcoming pathogen resistance or other issues growers face today, mainly if the environmental conditions and origins of species isolation are known.
Antimicrobial Peptides Exploration Through Genomic Mining and Computational Analysis
To date, there were more than 30 computational methods for AMP prediction and identification as of 2021 (Xu et al., 2021). Of those computational methods, amPEPpy is a user-friendly, open-source, portable, multi-threaded command-line application for predicting AMPs through genome-based screening using a random forest classifier (Lawrence et al., 2020). amPEPpy was validated as predicting AMPs more accurately than other approaches (Xu et al., 2021). We used amPEPpy to predict promising antimicrobial agents (Lawrence et al., 2020) within the Trichoderma genomes. We compared predicted AMPs sequences to sequences available on NCBI as of April 2021 because there are no online tools that cross-link data generated from amPEPpy to numerous known AMPs, as antiSMASH does with secondary metabolites. Accession numbers generated by amPEPpy are found in Supplementary Table 2 and functional annotations were summarized by major keywords into categories. Each functional annotation categories represent the percentage of amino acid sequences that match the predicted AMP motif: 100, 99.9–95.0, 94.9–90.0, 89.9–80.0, 79.9–70, and 69.9–51.0%. Anything below 50.9% was not considered for further analysis. A heatmap was generated to reflect these results in GraphPad Prism v 9.1.0 (221), indicating the number of times each category was predicted.
Based on Trichoderma comparative genomic screening, the Harzianum section has the highest number of predicted AMPs. Interestingly, among all organisms a large percentage of AMPs were annotated as hypothetical proteins, and the large number of observed hypothetical proteins in T. harzianum, T. virens, T. reesei, T. atroviride, and T. asperellum is intriguing. In some instances, these hypothetical proteins have a high sequence homology between species. In fact, T. lentiforme (accession number KAF3073950) and T. arundinaceum (accession number RFU77176) were predicted to have a 100% match for an unknown AMP and currently are described as “hypothetical protein” on NCBI.
Although hypothetical proteins dominated the annotations in the NCBI accession keyword search (Figure 8D), we listed the top five annotated proteins from each species. Among these, proteins annotated as non-ribosomal peptide synthetases (NRPSs) is a notable annotation because these proteins are known to produce AMPs in bacteria and fungi (Finking and Marahiel, 2004; Felnagle et al., 2008; De Cesare et al., 2020). Apart from this annotation, it was interesting to observed “glycoside hydrolase family” and “cytochrome P450 family” annotations as abundant keyword annotated protein names (Figure 8C). Interestingly, glycoside hydrolase family are found in many microbes and plants and are frequently associated with microbe-host interactions (Faure, 2002; Xu et al., 2004; Berlemont and Martiny, 2016; Snarr et al., 2017; Mendoza-Mendoza et al., 2018). Additionally, they have been shown to be required for needed for virulence in oomycetes and fungi triggering pathogen-associated molecular patterns (Mendoza-Mendoza et al., 2018; Tan et al., 2020; Zhang et al., 2021), formation of biofilms in bacteria (Szymańska et al., 2020), as a therapeutic for antibiofilm activity from pathogens across taxonomic kingdoms (Snarr et al., 2017), and in chemical defenses against herbivory (Xu et al., 2004). Glycoside hydrolases have also been shown to be involved in the production of secondary metabolites and peptaibols that have been found in 21 genomes of Trichoderma spp. (Fanelli et al., 2018; Mendoza-Mendoza et al., 2018). In a study to target activity-specific genetic markers for biocontrol from genomes of Trichoderma spp., Fanelli et al. (2018) compared protein family (Pfam) database, domains associated with stress tolerance and antagonistic activities, and found that 7 out of 15 Pfam domains with specific roles were CAZyme, containing hundreds of CAZymes proteins (Mistry et al., 2021). Two other Pfam domains with specific roles were the ABC transporter region and peptidase, which were also found in the results of this study (Fanelli et al., 2018). Like the glycoside hydrolases, cytochrome P450 superfamily have also been associated with antimicrobials (Geisler et al., 2013) and the production of primary secondary metabolites (Shin et al., 2018).
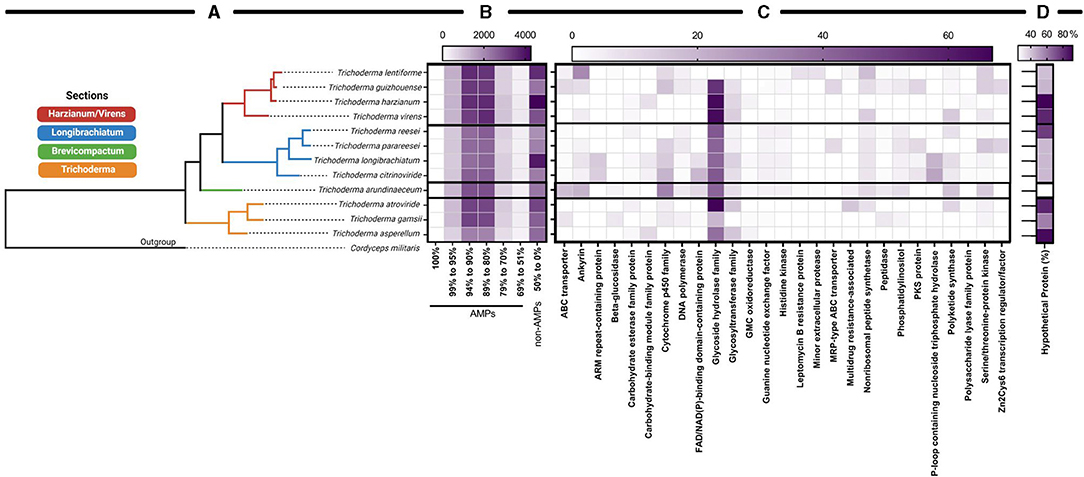
Figure 8. Predictive AMPs found in available genomes of species in Trichoderma. (A) A dendrogram based on the genomes of Trichoderma spp. with reference protein sequences and (B) organized by their sections and the corresponding heat map showing the numbers having a 100, 99–95, 94–90, 89–80, 79–70, 69–51, and ≤ 50% match with AMP identifying characters shared by species of Trichoderma within their sections. (C) The top five annotated accession numbers found across all species from 95 to 100% match of AMPs. (D) Percentage of hypothetical proteins from 95 to 100% match of AMPs. The outgroup is Cordyceps militaris.
Like the antiSMASH analysis, we used graph-theoretic analysis to further interrogate the predictions from amPEPpy to quantify the importance (or ranking) of the sections with respect to their probability of having AMP-related genes or keyword protein annotations. Two separate analyses were performed using the average values for the species of each section from the data in Figures 8A,B. We measured the min-max normalized broadcasting PageRank of the sections and the receiving PageRank of their probability of having AMP-related genes (Figures 9A,B) and, correspondingly, the measures related to the keyword protein annotations (Figures 9C,D).
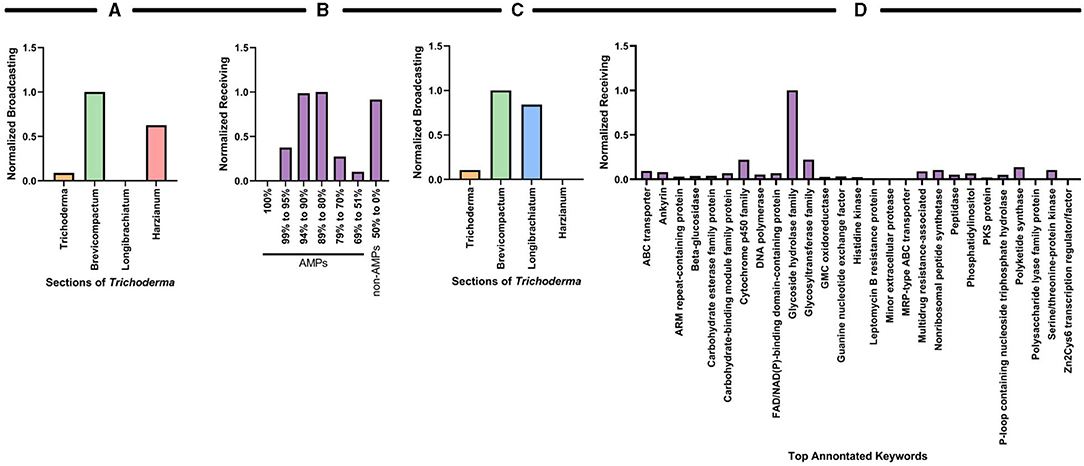
Figure 9. Graph theoretical analysis to characterize influential species sections of Trichoderma based on their probability of having AMP-related genes and keyword protein annotations. The PageRank measure for the sections with respect to the probable AMP-related gene are quantified by the normalized (A) broadcasting and (B) receiving values, respectively. The PageRank measure for the sections with respect to the keyword proteins are quantified by the normalized (C) broadcasting and (D) receiving values, respectively.
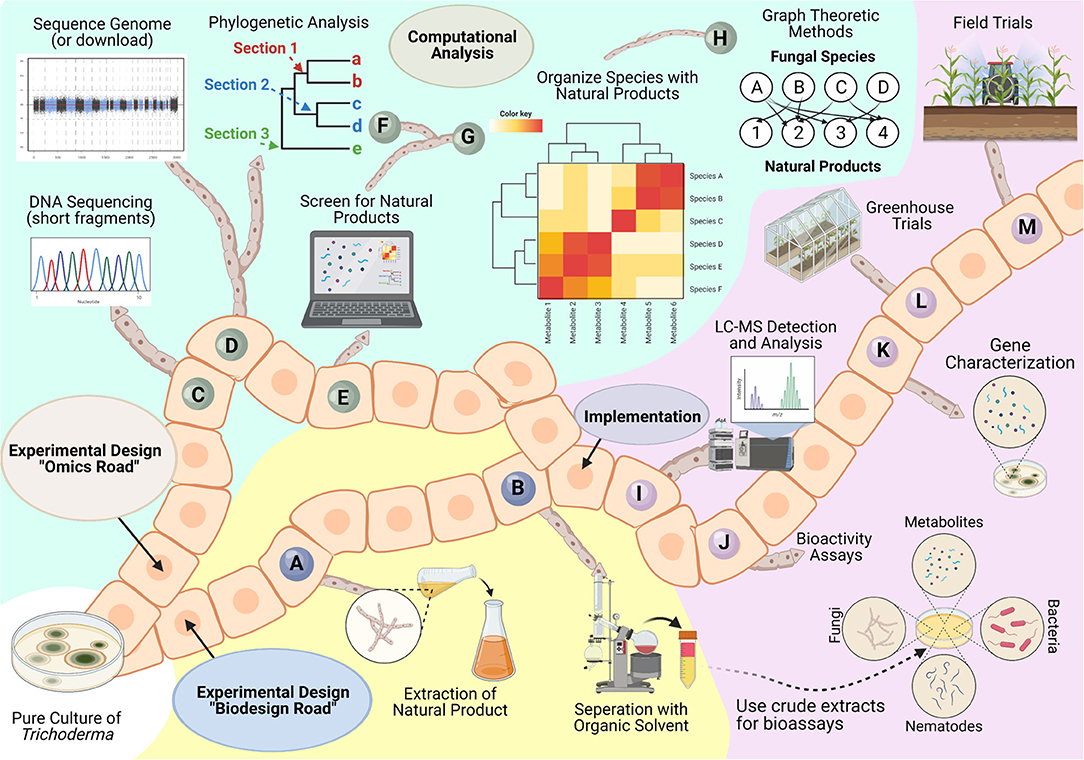
Figure 10. Roadmap for bioprospecting Trichoderma for natural products. Isolation of an axenic Trichoderma isolate is needed to initiate the roadmap. The biodesign road details the following general steps to extract and experimentally screen bioactive compounds: (A) Grow the culture in a liquid broth medium and collect the supernatant by filtration; (B) Use an appropriate organic solvent (e.g., ethyl acetate, methanol, etc.) to extract metabolites present in the filtrate. This crude exudate is composed of a mixture of natural products that can be further interrogated to identify targeted bioactivity properties. To predict putative natural products before extensive laboratory studies, then the “omics” road can be followed using the following steps: (C) Extract genomic DNA from the isolate and determine the species identity by amplifying and sequencing genetic markers like the internal transcribed spacer region (ITS), elongation factor 1-alpha (tef1), and RNA polymerase II subunit (rbp2) (Cai and Druzhinina, 2021); (D) Sequence reference genome of isolate; (E) Predict natural products using tools such as antiSMASH or amPEPpy. With this information, (F) comparative genomic analyses can be performed to determine which taxonomic section the target isolate belongs to, and (G,H) can be used to relate predictions across organisms to highlight the uniqueness and diversity of putative natural products across available Trichoderma by using graph theory. Both roads are led to an implementation step where experimental design aims to (I) selectively characterize predicted or isolated compound(s) by liquid or gas chromatography coupled to mass spectrometry, (J) evaluate bioactivity for selected compound(s), (K) determine the genes and proteins responsible for the production of the compound(s), (L) conduct trials to determine its potential usefulness in greenhouse conditions, and (M) test the compound effects in field trials.
The graph-theoretic analysis suggests that the Brevicompactum section, followed by Harzianum, had the most influence on the data (Figure 9A). Interestingly, there were several AMPs identified by amPEPpy that fall into the categories of 94–80% match based on current definitions of AMPs (Figure 9B). The graph-theoretic analysis also suggests that section Brevicompactum, closely followed by Longibrachiatum, have the most prevalence of predicted proteins potentially involved in AMP biosynthesis and production compared to all the other sections examined (Figure 9C). As illustrated in Figure 9D, graph theory results further substantiate the prevalence of “glycoside hydrolase family,” “cytochrome P450 family,” and “glycosyltransferase.”
Overall, by using whole-genome sequencing data available on NCBI, we were able to demonstrate the utility of genome prediction tools to identify putative secondary metabolites and AMPs that warrant further interrogation. Next, we describe how these inferred natural products can be experimentally interrogated with or without a priori knowledge.
Discussion
Experimental Framework to Characterize Natural Products in Trichoderma Species
In general, competition assays and choice or no-choice assays are often used as a first step for comparing the effects of a fungus on other microbes (Köhl et al., 2019) and would be useful in identifying negative association between an organism and the metabolic substances produced by another microbe (Fravel, 1988; Köhl et al., 2019). It is necessary to also understand how a particular fungus or natural product behaves in greenhouse conditions, which can be used to explore host-microbe interactions (Gibson et al., 1999) and efficacy against pathogens, host range, and environmental conditions before field trials. Next, with appropriate permits in-hand, field trial can be conducted to know how a biological product will perform in natural conditions, that are inherently more complex and less predictable. Often, off-target effects against non-target species, like insects, vertebrates, and weeds, as well as potential toxicity and carcinogenic effects on humans can be examined (Headrick and Goeden, 2001).
Herein, we limit the scope of this entire process to an experimental framework for the rapid discovery and identification of natural product and therefore will not be describing topics related to greenhouse or field trials (Figure 11).
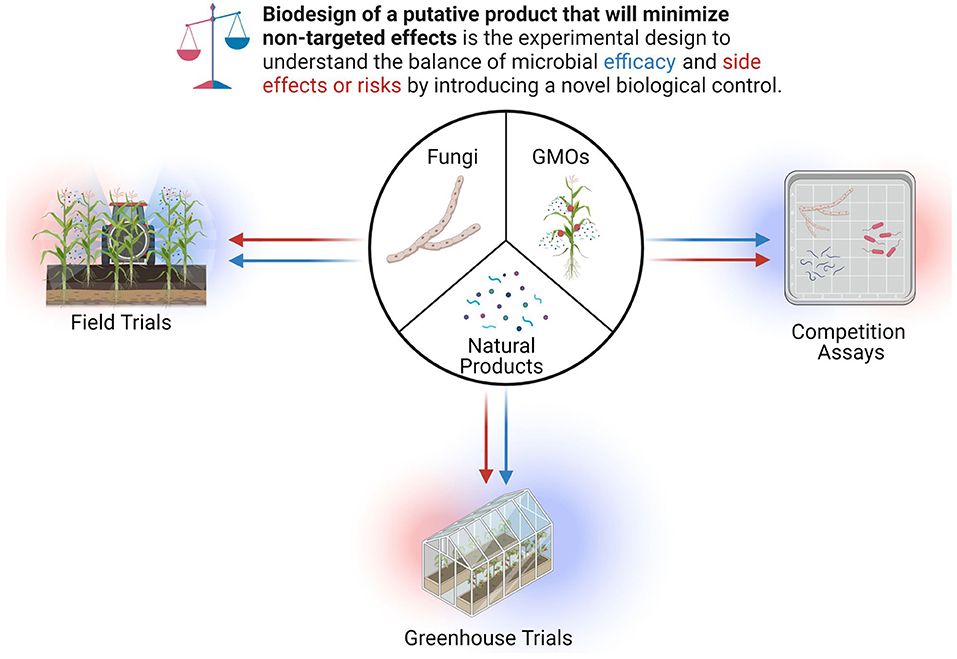
Figure 11. Biodesign of putative products to minimize untargeted effects.
Extraction of Natural Products From Trichoderma Species
Most Trichoderma isolates have been recovered from soils, from the fruiting bodies of other fungi, or from dead wood colonized by them (Druzhinina et al., 2011, Kubicek et al., 2019). The soil suspension, dilution, and aliquot methods are recommended for isolation of individual colonies. The most efficient substrate medium for the isolation and culture of Trichoderma isolates is potato dextrose agar supplemented with chloramphenicol, streptomycin, and rose bengal (Vargas Gil et al., 2009). Following the isolation and growth, the crude extract is collected to separate the desired natural products from the raw materials. The most common extraction methods are solvent extraction, distillation, pressing and sublimation (Zhang et al., 2018a) (Figure 12).
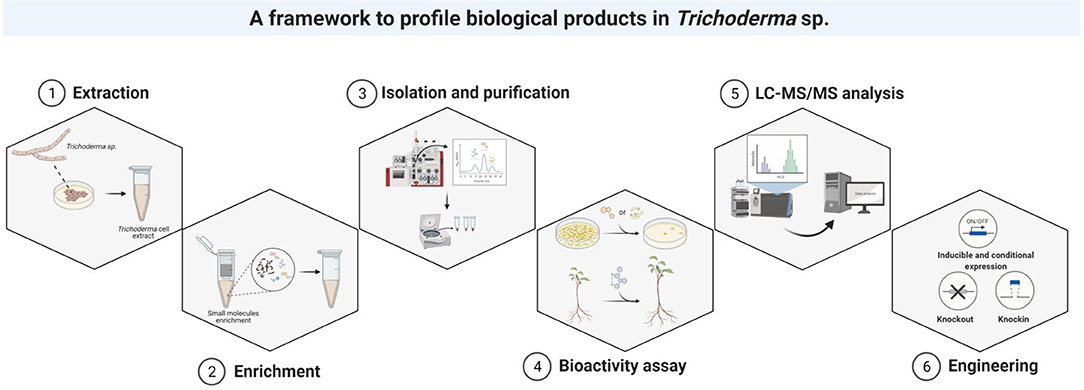
Figure 12. The framework for the identification of natural products in Trichoderma species.
Enrichment, Isolation, and Purification of Natural Products
Trichoderma crude extracts contain a complex mixture of biological molecules, with the bioactive products representing <1% of the crude extract (Odendaal et al., 2011). The low abundance of naturally occurring biological products represents a major bottleneck in their discovery (Hanson and Howell, 2004; McMullin et al., 2017). To selectively isolate low-abundance biological products from the complex mixture, various enrichment strategies have been developed. Size exclusion ultrafiltration strategies, such as molecular weight cutoff spin column filters, represent common methods of enriching the low-molecular-weight fraction of a crude extract (Hanson and Howell, 2004; McMullin et al., 2017). In addition to molecular weight cutoff, gel-based separations, solvent extractions, and size exclusion chromatography are all frequently used for biological product enrichment (Rivera-Chávez et al., 2017; Song et al., 2018; Zhang et al., 2018a).
Although the enrichment step helps to increase the representation of low-abundance biological products, the resultant fraction may still have impurities and diverse biological molecules. Thus, this fraction can be further fractionated based on physicochemical properties (e.g., polarity, hydrophobicity, stability, solubility) to isolate and purify the specific target product with bioactivity (Rivera-Chávez et al., 2017). The fractionation step starts with separation and detection of the natural products via chromatographic methods followed by detection methods such as ultraviolet light detectors. Based on a specific retention time or window, natural products can be selectively or broadly collected, lyophilized, and resolubilized in the appropriate solvent and concentration. The detailed technologies and techniques for isolation and purification of natural products from microbes have been extensively reviewed and explained by previous methodology and review publications (Bucar et al., 2013; Joana Gil-Chávez et al., 2013; Gomes et al., 2017; Zhang et al., 2018a; Atanasov et al., 2021).
Bioactivity Assays of Natural Products
There are two primary routes that can be taken to apply the active ingredient of interest: using the whole fungus as the active ingredient or applying the natural product of the fungus, whether biosynthesized by the organism or chemically synthesized in vitro. In deciding between using the fungus or using the compound alone, it is useful to consider the mechanism by which it will carry out its effect.
Using the fungus itself, one mode of action is for the beneficial fungus to outcompete the pathogenic organisms for nutrients and space on the plant by colonizing the mutual areas (Ghorbanpour et al., 2018). For this to be effective, the fungus must have strong colonization capabilities and persist long enough to out-compete the pathogenic population. Trichoderma excels at root colonization because of its production of compounds such as hydrophobins, which allow its hyphal filaments to attach to a multitude of surfaces, aiding its ability to survive in the presence of other unwanted organisms (Guzmán-Guzmán et al., 2017). Furthermore, some Trichoderma species produce siderophores, which chelate iron, making it unavailable in the environment and potentially inhibiting the growth of competing organisms (Harman et al., 2004). In addition to the competition, Trichoderma can use mycoparasitic interactions to recognize and attack the parasite (Błaszczyk et al., 2014). One mechanism Trichoderma uses during its attack is to secrete various cell wall–degrading enzymes such as cellulase, pectinase, glucanase, lipase, and protease, which degrade several compounds, including chitin and glucan polysaccharides (Błaszczyk et al., 2014). Initially, lectins are used to recognize the pathogen and the hyphae coil around the pathogen and then penetrate the cell and use the enzymes previously mentioned to degrade the cell wall (Ghorbanpour et al., 2018). In addition to mycoparasitism, these cell wall degrading enzymes are involved in antibiosis, an antagonistic interaction in which Trichoderma produces these and other antimicrobial compounds, such as antibiotics and secondary metabolites, that exhibit a lethal effect on the parasitic organism (Błaszczyk et al., 2014); however, mycoparasitism and antibiosis may not be the primary mechanisms of biocontrol (Lorito et al., 2010). Included in this list of antimicrobials are compounds such as trichodermin, herizanolide, peptabiols, and epipolythiodioxopiperazines (Ghorbanpour et al., 2018). Two additional mechanisms include mycovirus-mediated cross-protection and induced systemic resistance. While these are different mechanisms—one is an infection with a non-virulent mycovirus and the other a beneficial organism inducing an immune response—they have similar overarching effects of priming/preparing the plant defense system for infection with pathogens (Ghorbanpour et al., 2018).
The other alternative is to use a natural product of the fungus as the active ingredient, which may allow for a more targeted outcome and circumvent the issues that may come with introducing a non-native organism to a new environment (Butt et al., 2001). These natural products, such as secondary metabolites and antimicrobial peptides, can also assist in the plant's defense mechanisms, function in an antibiotic manner, and compete against pathogens for nutrients such as metals (Keswani et al., 2019).
Once either the compound or the entire fungus is selected, a series of trials is necessary to validate its efficacy. For antimicrobial efficacy, it's important to establish a minimum inhibitory concentration with respect to the pathogenic organism targeted by the biocontrol agent and the commensal organisms that the product will encounter, determining if and how detrimental these are to them. To determine the minimum inhibitory concentration, the reconstituted samples from the fraction collectors in an appropriate solvent can be serially diluted and tested. Alternatively, a micro-titer broth dilution can be carried out (Aboobaker et al., 2019). This relatively easy screening method is a valuable tool, as any potential compounds can be immediately weeded out if they require an extremely high concentration to be successful. Additionally, dual-culture assays can be performed to determine the suppression of pathogenic growth directly (Lahlali and Hijri, 2010). Once initial screening has taken place, larger-scale experiments such as greenhouse trials can be carried out. During a greenhouse trial, data can be gathered on the effects of inoculating a plant of interest with the biocontrol agent; and preliminary conclusions will be reached regarding the overall health of the plant and any antagonistic effects (Perelló et al., 2003). Upon completion of greenhouse trials, larger-scale field experiments could be done to more closely mimic the conditions biocontrol agents encounter once they are deployed in nature. In these field experiments, larger-scale data, such as any effects on growth, seed production, and pathogen reduction, will be excellent indicators of market viability (Abdel-Fattah et al., 2007). Once the vetting process is completed and the biocontrol agent has been thoroughly assessed, mass production issues must be explored to ensure the product is effective and lucrative, including growth conditions of the fungus, synthetic synthesis of individual compounds vs. extraction from the fungus, formula optimization for prolonged shelf life, and determination of a dispersal method.
Natural Product Identification and Characterization
Following the detection and isolation of fractions with relevant bioactivity, biological product identification can be carried out with chromatography-coupled mass spectrometry (MS) for complex samples or direct injection/infusion for pure or low-complex mixtures. For chromatographic techniques, gas chromatography (GC) and liquid chromatography (LC) represent the common choices. GC is commonly used for volatile and polar small compounds, while LC allows the identification of mid-polar to non-polar metabolites. The choice of technique used can dictate what types of small molecules can be identified and this needs to be made on a case-by-case basis. The biological products predicted using prediction tools like antiSMASH and amPEPpy from the Trichoderma genome can help provide information needed to select suitable techniques for biological product profiling.
Alongside the chromatographic techniques, MS represents a key experimental platform that is applied in the discovery, and characterization of natural products. Generally, MS-based approaches used to measure natural products is broadly categorized as metabolomics. Metabolomics, either targeted or untargeted, can be used to identify the natural products in Trichoderma. The advantages and disadvantages of these approaches have been reviewed extensively (Roberts et al., 2012; Schrimpe-Rutledge et al., 2016). For the identification, quantification, and structural elucidation of natural products, MS data are searched against the spectral libraries.
A significant challenge in the sensitive identification of biological products via MS includes the lack of representative databases or spectral libraries. Thus, the characterization of biological products with the use of de novo search strategies (Cheng Q. et al., 2010). De novo sequencing algorithms derive biological product structures, molecular formulas, or sequences using only fragmented ionic information from the tandem mass spectra, thus providing the complete profiling of all biological products. For metabolomics, SIRIUS 4 is one of the leading de novo software, using deep neural networks for metabolite identification from high-resolution tandem MS data (Dührkop et al., 2019). It integrates high-resolution isotope pattern analysis and fragmentation trees with structural elucidation to provide a combined and coherent assessment of molecular structures from MS/MS data for large datasets (Dührkop et al., 2019). This tool allows the identification of the molecular formula of a query compound with very high accuracy; no spectral or structural databases are required for this step of the analysis because all theoretically possible molecular formulas are considered, allowing one to overcome the limitations of the current structural databases (Dührkop et al., 2019). For peptide-based biological products, PEAKS is well-established search software that also utilized machine-learning and combines de novo sequencing with traditional database searching (Ma et al., 2003; Zhang et al., 2012).
Genetic Engineering
Several transformation systems, relying on effective selectable markers, have been developed to genetically modify different Trichoderma species. The most widely used and optimized procedures for doing so are based on protoplasts (Gruber et al., 1990; Li et al., 2017) and Agrobacterium tumefaciens-mediated transformation (Zhong et al., 2007; Yang et al., 2011). Other less common genetic approaches are reported to be used in studying Trichoderma, such as Biolistics (gene gun) (Te'o et al., 2002), and electroporation (Wanka, 2021). The most investigated species in genetic engineering studies to produce enzymes or metabolites is T. reesei (Liu et al., 2015). Genetic transformation systems have also been developed for other Trichoderma species, including T. harzianum (Goldman et al., 1990; Zeilinger, 2004), T. viride (Manczinger et al., 1997), T. atroviride (Zeilinger, 2004; Calcáneo-Hernández et al., 2020), and T. longibrachiatum (Sánchez-Torres et al., 1994). In recent years, the CRISPR/Cas9-mediated genome editing system has been successfully employed in T. reesei and delivered in the format of either plasmid (Liu et al., 2015) or ribonucleoprotein complexes (Hao and Su, 2019). Finally, the transformation methods developed for Trichoderma species can be used for overexpression experiments on putative backbone genes or transcription factors as a strategy to increase the production and allow the characterization of specific metabolites produced in low amounts or not produced under standard culture conditions.
Methods to Implement Computational Analysis Into Biological Products for Integrated Pest Management System
Fungi as an Active Ingredient
For numerous biological products, the active ingredient uses reactivated lyophilized cells of the fungus itself. To propose new Trichoderma species as active ingredients requires a fundamental understanding of the fungus behavior within an environment. Therefore, the approach should consider the fungus lifecycle, its interactions with other microbes and organisms, its responses to abiotic and biotic stresses, its pathogenicity behavior on cash crops, and its ability to promote the growth of competing weeds that can serve as vectors for other plant pathogens. Regarding Trichoderma species, several were shown to promote plant growth under abiotic and biotic stress conditions. Although all studies reviewed concentrated on how the fungus helped a plant in stress conditions, few or none focused on how stress affected the survivability of the fungus. The benefits of establishing a new species or strain as an active ingredient reside in getting more specific and long-lasting effects, cost-efficient disease management, and higher yields of cash crops. Particularly in Trichoderma, there is a strong history of species used as successful active ingredients (Figure 2, Supplementary Table 1). Another consideration is the effectiveness of a strain within a species or the efficacy of mixing different strains or species of Trichoderma and fungicides as formulations and active ingredients.
Importantly, due to the genetic diversity of Trichoderma strains, they can be unpredictable in terms of mechanisms and biocontrol improvement in different environmental settings (Benítez et al., 2004) and therefore should not be generalized at the species level. For this reason, companies use a particular strain in their products. Combinations of Trichoderma strains or species have been shown to better promote growth of cash crops in agriculture and forestry than a single strain or species (Chirino-Valle et al., 2016; Halifu et al., 2019; Chen et al., 2021). As such, there are several commercially available products with mixed strains or species of Trichoderma for better biocontrol, like Bioten® WP, BioWorks® Rootshield® Plus+ WP; Kiwivax®, Vinevax™ Bio-Dowel, and Vinevax™ Pruning Wound Dressing, to name a few (Supplementary Table 1). In addition, there are genetically altered hybrid strains between closely related Trichoderma species created by protoplast fusion (Stasz et al., 1988). These mutant strains had shown better biocontrol efficacy by selecting secondary metabolites or enzymes used for antimicrobial activity than individual species (Stasz, 1990; Hanson and Howell, 2002). However, to our knowledge, it is unknown if there are any commercial products or field experiments to test the efficacy of these modified fungal strains. Finally, mixing biological control agents (like Trichoderma) with fungicides (or derivatives of natural products) has been shown to reduce disease pressure (Ons et al., 2020). Since biological controls are dependent on environmental conditions, combining a biological product with fungicides can increase efficacy (Ons et al., 2020). However, researchers should examine if the Trichoderma strains are susceptible to the fungicide before using them in a mixed application. Moreover, there are few studies investigating biological products durability to control plant pathogens (Bardin et al., 2015), and it is known that there are pathogens resistant to some fungicides (Hahn, 2014; Hollomon, 2015). Therefore, further studies are needed to determine the durability and environmental sustainability by combining both biological products with fungicides.
Some disadvantages to applying active fungal strains in a field is that a fungus could sexually recombine with a native population, introducing unknown ecological consequences, or simply be incapable of surviving in the newly introduced environment. Thus, understanding the fungal lifecycle is critical to designing a new biological product. Fungal clones can expand their populations but still lack adaptability because of short-term survival mechanisms (Drenth et al., 2019). Sexual recombination is suitable for long-term survival and adaptability to a changing environment (Drenth et al., 2019). However, when non-native fungi capable of sexually reproducing are introduced into an environment, they can reproduce with a native population, creating hybridization and potentially having non-target effects. Hybridization has been shown in numerous pathogenic fungi and has led to the decline of several host crops or ornamental plants and targeting of new hosts (Schardl and Craven, 2003; Depotter et al., 2016; Stukenbrock, 2016). Alternatively, hybridization in symbiotic fungi has been beneficial for plant adaptation (particularly in grasses) for nutrient acquisition and stress conditions (Hamilton et al., 2010; Saari and Faeth, 2012; Oberhofer et al., 2014; Jia et al., 2016). Therefore, a screen of potential mating type genes within a species should be considered with the proposition of knocking out the mating genes if possible.
Fungi, in particular Trichoderma, have been used as active ingredients in bioherbicides to control weeds; however, there are no reports of their natural products being used (Kremer, 2005; Triolet et al., 2020). There are ongoing investigations to understand the mechanisms involved in controlling weed populations without affecting the crop of interest (Harding and Raizada, 2015). The main limiting factors for using fungi as bioherbicides are their narrow host range, their specific biotic requirements for good efficacy, and their possible non-targeted effects (Kremer, 2005). Therefore, the extraction of compounds possessing herbicide properties from these fungi can be a fruitful endeavor for overcoming these limitations (Kremer, 2005; Harding and Raizada, 2015; Triolet et al., 2020).
In the promising bioactive ingredient from a Trichoderma species (Figure 3), much bioactivity has been observed in the laboratory. However, the potential for transferring these species from the lab to the field is vague.
Natural Products as an Active Ingredient
Natural products deriving from microbes have also been used as active ingredients (Singh and Yadav, 2020) primarily those released during antibiosis interactions. Antibiosis is the secretion of natural products (e.g., secondary metabolites, AMPs) that have antimicrobial properties in the vicinity of other microbes (Howell, 1998). Secreted molecules might be the key to triggering a response from a host plant or Trichoderma itself. For example, the production of peptides or small molecules from a pathogenic fungus and its perception through G protein-coupled receptors might trigger a mycoparasitism response in Trichoderma species (Druzhinina et al., 2011). Several classes of molecules secreted by Trichoderma, like xylanases, peptaibols, swollenin, and cerato-platanins, have been reported to act as microbe-associated molecular patterns that trigger immunity in plants (Howell, 2003; Druzhinina et al., 2011; Newman et al., 2013). Owing to the difficulty of mass-producing biological products using fungi as active ingredients, and the agrochemical effects on the environment, natural products appear as promising solutions for implementation in integrated pest management (Dayan et al., 2009; Yan et al., 2018; Singh and Yadav, 2020; Triolet et al., 2020). To propose a new natural product as an active ingredient, a fundamental understanding of the molecule chemistry and behavior within an environment is needed. The main benefits of this approach are that these natural products could be used in combination with existing pesticides with known identities, bioactivities, and effects on plant growth promotion and defenses. An example of a natural product used in pesticides is the quinone outside inhibitor (QoI) family of fungicides derived from the group of natural products called “strobilurin” (Balba, 2007; Vincelli, 2012). This group was first isolated from the wood-rotting fungus Strobilurus tenacellus (Anke et al., 1977). QoI fungicides inhibit mitochondrial responses by binding at the Qo site of the cytochrome bc1 enzyme complex. QoI fungicides are a preventative measure through translaminar movement and are known to result in plant growth enhancements (Balba, 2007; Vincelli, 2012). QoI fungicides pose less risk to human health than alternative pesticides (Balba, 2007; Vincelli, 2012). Unfortunately, there are resistant QoI fungicide populations because of disease pressure in growers' fields, creating a need for alternative sources of natural products (https://www.frac.info/). As shown in Figure 5 and Table 2, Trichoderma citrinoviridie, T. virens, and T. viride all have BGCs with a 50% sequence match, and T. atroviride and T. koningiopsis both have BGCs with a 37% match with the cluster encoding the putative metabolite Ascochlorin. Berry et al. (2010) have demonstrated that Ascoholorin, isolated from the pathogenic fungus Ascochyta viciae, is a cyctochrome bc1 inhibitor acting on both Qi and Qo sites. Currently, Ascoholorin has not been identified or characterized in Trichoderma species. However, based on our bioinformatic results, there might possibly be an Ascoholorin-like compound in Trichoderma species, which could be investigated as a biopesticide.
To our knowledge, there are minimal caveats for introducing natural products into the environment; yet they might present some unknown non-targeted effects on beneficial microbes, livestock, and humans. Natural products are potentially unstable in extremely cold temperatures; therefore, they may not overwinter in the field, allowing more pathogenic inoculum to increase. They might also become denatured after long exposure to environmental factors, such as ultraviolet radiation.
Genetically Modified Crops With Biosynthetic Gene Clusters
Genetically modified organisms (GMOs) are engineered to produce a natural product by inserting a gene of interest into the target plant genome. This can enhance a plant's ability to fight against pathogenic insects and resist herbicides. The implementation of GMO crops with natural products derived from microbes started over 30 years ago. Currently, several GMO crops engineered with natural products, such as corn, soybeans, papaya, tomatoes, rice, and cotton, are available. The world's first GMO crop, FLAVR SAVR, was a virus-resistant tomato variety that suppressed the enzyme polygalacturonase, which dissolves cell-wall pectin and causes the fruit to become soft (Bruening and Lyons, 2000). Currently, three GMO crops are commonly used: Bacillus thuringiensis corn (Bt corn), Roundup Ready crops, and the Rainbow papaya, all approved by the United States Environmental Protection Agency and the United States Department of Agriculture. Bt corn is a hybrid plant bioengineered with a broad-spectrum insecticide in which the active ingredient is crystal (Cry) protein toxins (Gewin, 2003; Koch et al., 2015). Roundup Ready crops (soybeans, corn, alfalfa, cotton, and sorghum), genetically modified to contain a protein-coding for a glyphosate-insensitive enzyme, can kill competing weeds and reduce the source of inoculum for many plant pathogens. The Rainbow papaya has a gene from a papaya ringspot virus inserted into the fruit to make it resistant to the same virus, using the same logic as vaccination in mammalians.
Previous successes have shown that producing natural biological products directly from GMO crops is a feasible procedure. To our knowledge, there are no reports of inserting BGCs from Trichoderma into GMO crops. Thus, this idea may represent a new niche to explore in agriculture. However, caution is essential in creating GMO crops, as there are many rules and regulations to follow depending on the agricultural system for each country.
Conclusions
The success of any biological product will rely on its efficacy, cost efficiency, stability, and profitability. Our roadmap outlines the multifaceted abilities of Trichoderma as the prominent biological control agent. In addition, we demonstrate how to use omics and machine learning to identify new or existing products that can be used in the market and how to extract and implement those products while listing the benefits and caveats (Figure 13).
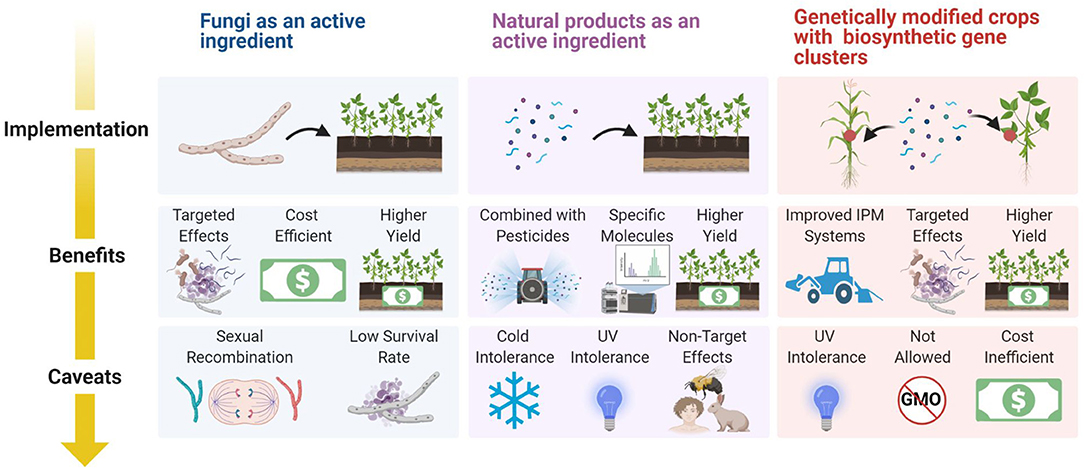
Figure 13. Considerations for how to introduce biological products from Trichoderma into field applications.
However, efficiently, and effectively translating laboratory observations to field applications should be linked to a purpose tied to a specific market or reason. For example, renewable and sustainable biofuels and bioproducts from plant biomass are a steadily growing industry representing alternative fuel sources to petroleum. Identifying novel or existing Trichoderma species or natural products to help promote terrestrial plants for higher biofuel and bioproduct production or the promotion of associated beneficial microbes can improve efforts to advance the use of bioenergy. Experiments have shown that Trichoderma species can improve the growth parameters of bioenergy crops like hybrid grasses (Chirino-Valle et al., 2016). In addition, there is a need to understand how using biocontrol products to influence environmental microbiome processes that drive global carbon cycling will impact climate change. Biological control could be a way to biologically mitigate heavy metals, radionuclides, or agrochemicals present in the soil, coastal sediments, freshwater, and subsurfaces.
Another example is sustainable agriculture, where the results should address farmers' and industry partners' needs and concerns. Farmers will adopt sustainable programs when there is a short-term economic outcome and if doing so benefits their farms, the environment, or both (Piñeiro et al., 2020). Ultimately, any biological control implementation should follow the goals of integrated pest management—economic viability, environmental safety, and social acceptability. These goals address the needs and concerns of the producer, the seller, and the consumer (Dara, 2019). For some engineered products, like GMOs, an understanding of the geopolitical climate is necessary for developing research and economic plans.
As outlined in this systematic review, new issues will arise as more biological products are developed, marketed, and implemented into agricultural or environmental settings. We have observed that several known compounds have been identified from Trichoderma species. However, these results do not translate into active ingredients available on the market. We suggest that the implementation team involved in developing products should be multidisciplinary. For example, for a promising biological product to be marketed, an ideal implementation team should include, but not be limited to, researchers that know the market/economy, chemistry, computational analysis, and the integral dynamics of an agricultural microcosm (e.g., plants, microbes, insects, etc.). Lastly, there are numerous new publications that are made yearly based on novel features from global studies on Trichoderma isolates. With this extensive dataset, there is need for additional high-quality genomes and representation of the various sections within Trichoderma. Altogether, this comprehensive roadmap can serve as guidance for the future production of biological products from Trichoderma and could be used for other fungal systems.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding authors.
Author Contributions
TR, HS, MS, JL, and PA designed research. TR, HS, MG, JE, MS, JL, and PA performed research and wrote the manuscript. MG conducted graph-theory analysis. JE and TR conducted phylogeny analysis. TR and HS analyzed antiSMASH results. TR and MS analyzed amPEPpy results. All authors contributed to the article and approved the submitted version.
Funding
This research was sponsored by the Genomic Science Program, US Department of Energy (DOE), Office of Science, Biological and Environmental Research, as part of the Secure Ecosystem Engineering and Design and the Plant Microbe Interfaces Scientific Focus Areas at the Oak Ridge National Laboratory (ORNL). ORNL was managed by UT-Battelle LLC for DOE under contract DE-AC05-00OR22725. To perform the graph-theoretic analysis, MG used the resources of the Oak Ridge Leadership Computing Facility at ORNL, which was supported by the DOE Office of Science under Contract No. DE-AC05-00OR22725.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Licenses and Permissions
This manuscript has been authored by UT-Battelle, LLC, under contract DE-AC05-00OR22725 with the US Department of Energy (DOE). The US government retains and the publisher, by accepting the article for publication, acknowledges that the US government retains a nonexclusive, paid-up, irrevocable, worldwide license to publish or reproduce the published form of this manuscript, or allow others to do so, for US government purposes. DOE will provide public access to these results of federally sponsored research in accordance with the DOE Public Access Plan (http://energy.gov/downloads/doe-public-access-plan).
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors thank Dr. Joanna Tannous for her critical comments and the reviewers for their contribution to the peer review of this work. Figures 1, 4, 6, 8A, 10–13 were created or edited with www.BioRender.com.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffunb.2021.716511/full#supplementary-material
Supplementary Table 1. List of products that uses Trichoderma strains used as an active ingredient.
Supplementary Table 2. List of accession numbers and probability to be an AMP for each Trichoderma species examined with amPEPpy.
References
Abd-Allah, E. F., and Ezzat, S. M. (2005). Natural occurrences of citrinin in rice grains and its biocontrol by Trichoderma hamatum. Phytoparasitica 33, 73–84. doi: 10.1007/BF02980928
Abdel-Fattah, G. M., Shabana, Y. M., Ismail, A. E., and Rashad, Y. M. (2007). Trichoderma harzianum: a biocontrol agent against Bipolaris oryzae. Mycopathologia 164, 81–89. doi: 10.1007/s11046-007-9032-9
Aboobaker, Z., Viljoen, A., Chen, W., Crous, P. W., Maharaj, V. J., and van Vuuren, S. (2019). Endophytic fungi isolated from Pelargonium sidoides DC: antimicrobial interaction and isolation of a bioactive compound. South African J. Bot. 122, 535–542. doi: 10.1016/j.sajb.2019.01.011
Abraham, W. R., Meyer, H., and Abate, D. (1995). Curvupallides, a new class of alkaloids from the fungus Curvularia pallescens. Tetrahedron 51, 4947–4952. doi: 10.1016/0040-4020(95)98692-B
Aguilar-Pontes, M. V., de Vries, R. P., and Zhou, M. (2014). (Post-)Genomics approaches in fungal research. Brief. Funct. Genomics 13, 424–439. doi: 10.1093/bfgp/elu028
Allison, A. C., Kowalski, W. J., Muller, C. D., and Eugui, E. M. (1993). Mechanisms of action of mycophenolic acid. Ann. N. Y. Acad. Sci. 696, 63–87. doi: 10.1111/j.1749-6632.1993.tb17143.x
Anke, T., Oberwinkler, F., Steglich, W., and Schramm, G. (1977). The strobilurins - new antifungal antibiotics from the basidiomycete Strobilurus tenacellus. J. Antibiot. 30, 806–810. doi: 10.7164/antibiotics.30.806
Arinbasarova, A. Y., Baskunov, B. P., and Medentsev, A. G. (2017). A low-molecular mass antimicrobial peptide from Trichoderma cf. aureoviride Rifai VKM F-4268D. Microbiol. Russian Fed. 86, 289–291. doi: 10.1134/S0026261717020059
Ariza, M. R., Larsen, T. O., Petersen, B. O., Duus, J., and Barrero, A. F. (2002). Penicillium digitatum metabolites on synthetic media and citrus fruits. J. Agric. Food Chem. 50, 6361–6365. doi: 10.1021/jf020398d
Arnold, S. L., and Panaccione, D. G. (2017). Biosynthesis of the pharmaceutically important fungal ergot alkaloid dihydrolysergic acid requires a specialized allele of cloA. Appl. Environ. Microbiol. 83. doi: 10.1128/AEM.00805-17
Atanasov, A. G., Zotchev, S. B., Dirsch, V. M., Orhan, I. E., Banach, M., Rollinger, J. M., et al. (2021). Natural products in drug discovery: advances and opportunities. Nat. Rev. Drug Discov. 20, 200–216. doi: 10.1038/s41573-020-00114-z
Atanasova, L., Jaklitsch, W. M., Komoń-Zelazowska, M., Kubicek, C. P., and Druzhinina, I. S. (2010). Clonal species Trichoderma parareesei sp. nov. likely resembles the ancestor of the cellulase producer Hypocrea jecorina/T. reesei. Appl. Environ. Microbiol. 76, 7259–7267. doi: 10.1128/AEM.01184-10
Baker, S. E., Perrone, G., Richardson, N. M., Gallo, A., and Kubicek, C. P. (2012). Phylogenomic analysis of polyketide synthase-encoding genes in Trichoderma. Microbiology 158, 147–154. doi: 10.1099/mic.0.053462-0
Balba, H. (2007). Review of strobilurin fungicide chemicals. J. Environ. Sci. Heal. Part B Pestic. Food Contam. Agric. Wastes 42, 441–451. doi: 10.1080/03601230701316465
Bale, J. S., Van Lenteren, J. C., and Bigler, F. (2008). Biological control and sustainable food production. Philos. Trans. R. Soc. B Biol. Sci. 363, 761–776. doi: 10.1098/rstb.2007.2182
Bardin, M., Ajouz, S., Comby, M., Lopez-Ferber, M., Graillot, B., Siegwart, M., et al. (2015). Is the efficacy of biological control against plant diseases likely to be more durable than that of chemical pesticides? Front. Plant Sci. 6:566. doi: 10.3389/fpls.2015.00566
Baroncelli, R., Zapparata, A., Piaggeschi, G., Sarrocco, S., and Vannacci, G. (2016). Draft whole-genome sequence of Trichoderma gamsii T6085, a promising biocontrol agent of Fusarium head blight on wheat. Genome Announc. 4:e01747-15. doi: 10.1128/genomeA.01747-15
Barratt, B. I. P., Moran, V. C., Bigler, F., and van Lenteren, J. C. (2018). The status of biological control and recommendations for improving uptake for the future. Biocontrology 63, 155–167. doi: 10.1007/s10526-017-9831-y
Begum, N., Qin, C., Ahanger, M. A., Raza, S., Khan, M. I., Ashraf, M., et al. (2019). Role of arbuscular mycorrhizal fungi in plant growth regulation: implications in abiotic stress tolerance. Front. Plant Sci. 10:1068. doi: 10.3389/fpls.2019.01068
Benítez, T., Rincón, A. M., Limón, M. C., and Codón, A. C. (2004). Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 7, 249–260. doi: 10.2436/im.v7i4.9480
Benjamin, E. O., and Wesseler, J. H. H. (2016). A socioeconomic analysis of biocontrol in integrated pest management: a review of the effects of uncertainty, irreversibility and flexibility. NJAS Wageningen J. Life Sci. 77, 53–60. doi: 10.1016/j.njas.2016.03.002
Bergstrom, J. D., Kurtz, M. M., Rew, D. J., Amend, A. M., Karkas, J. D., Bostedor, R. G., et al. (1993). Zaragozic acids: a family of fungal metabolites that are picomolar competitive inhibitors of squalene synthase. Proc. Natl. Acad. Sci. U.S.A. 90, 80–84. doi: 10.1073/pnas.90.1.80
Berlemont, R., and Martiny, A. C. (2016). Glycoside hydrolases across environmental microbial communities. PLoS Comput. Biol. 12:e1005300. doi: 10.1371/journal.pcbi.1005300
Berry, E. A., Huang, L., Lee, D. W., Daldal, F., Nagai, K., and Minagawa, N. (2010). Ascochlorin is a novel, specific inhibitor of the mitochondrial cytochrome bc1 complex. Biochim. Biophys. Acta - Bioenerg. 1797, 360–370. doi: 10.1016/j.bbabio.2009.12.003
Biasetto, C. R., Somensi, A., Sordi, R., Chapla, V. M., Ebrahimi, S. N., Silva, G. H., et al. (2020). The new koninginins T-U from Phomopsis stipata, an endophytic fungus isolated from Styrax camporum Pohl. Phytochem. Lett. 36, 106–110. doi: 10.1016/j.phytol.2020.01.019
Bills, G. F., Peláez, F., Polishook, J. D., Diez-Matas, M. T., Harris, G. H., Clapp, W. H., et al. (1994). Distribution of zaragozic acids (squalestatins) among filamentous ascomycetes. Mycol. Res. 98, 733–739. doi: 10.1016/S0953-7562(09)81046-0
Błaszczyk, L., Siwulski, M., Sobieralski, K., Lisiecka, J., and Jedryczka, M. (2014). Trichoderma spp. - application and prospects for use in organic farming and industry. J. Plant Prot. Res. 54, 309–317. doi: 10.2478/jppr-2014-0047
Blin, K., Shaw, S., Steinke, K., Villebro, R., Ziemert, N., Lee, S. Y., et al. (2019). AntiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 47, W81–W87. doi: 10.1093/nar/gkz310
Bokesch, H. R., Cartner, L. K., Fuller, R. W., Wilson, J. A., Henrich, C. J., Kelley, J. A., et al. (2010). Inhibition of ABCG2-mediated drug efflux by naphthopyrones from marine crinoids. Bioorganic Med. Chem. Lett. 20, 3848–3850. doi: 10.1016/j.bmcl.2010.05.057
Bonsch, B., Belt, V., Bartel, C., Duensing, N., Koziol, M., Lazarus, C. M., et al. (2016). Identification of genes encoding squalestatin S1 biosynthesis and: in vitro production of new squalestatin analogues. Chem. Commun. 52, 6777–6780. doi: 10.1039/c6cc02130a
Bucar, F., Wube, A., and Schmid, M. (2013). Natural product isolation-how to get from biological material to pure compounds. Nat. Prod. Rep. 30, 525–545. doi: 10.1039/c3np20106f
Butt, T. M., Jackson, C., and Magan, N. (2001). Fungi as Biocontrol Agents: Progress, Problems, and Potential. Wallingford: CABI.
Caboche, S., Pupin, M., Leclère, V., Fontaine, A., Jacques, P., and Kucherov, G. (2008). NORINE: a database of nonribosomal peptides. Nucleic Acids Res. 36:D326. doi: 10.1093/nar/gkm792
Cai, F., and Druzhinina, I. S. (2021). In honor of John Bissett: authoritative guidelines on molecular identification of Trichoderma. Fungal Divers 107, 1–69. doi: 10.1007/s13225-020-00464-4
Calcáneo-Hernández, G., Rojas-Espinosa, E., Landeros-Jaime, F., Cervantes-Chávez, J. A., and Esquivel-Naranjo, E. U. (2020). An efficient transformation system for Trichoderma atroviride using the pyr4 gene as a selectable marker. Brazil. J. Microbiol. 51, 1631–1643. doi: 10.1007/s42770-020-00329-7
Cardoza, R. E., Malmierca, M. G., Hermosa, M. R., Alexander, N. J., McCormick, S. P., Proctor, R. H., et al. (2011). Identification of loci and functional characterization of trichothecene biosynthesis genes in filamentous fungi of the genus Trichoderma. Appl. Environ. Microbiol. 77, 4867–4877. doi: 10.1128/AEM.00595-11
Castrillo, M. L., Bich, G., ángel Modenutti, C., Turjanski, A., Zapata, P. D., and Villalba, L. L. (2017). First whole-genome shotgun sequence of a promising cellulase secretor, Trichoderma koningiopsis strain POS7. Genome Announc. 5:e00823-17. doi: 10.1128/genomeA.00823-17
Cerrini, S., Lamba, D., Scatturin, A., Rossi, C., and Ughetto, G. (1989). The crystal and molecular structure of the α-helical nonapeptide antibiotic leucinostatin A. Biopolymers 28, 409–420. doi: 10.1002/bip.360280138
Chavali, A. K., and Rhee, S. Y. (2018). Bioinformatics tools for the identification of gene clusters that biosynthesize specialized metabolites. Brief. Bioinformatics 19, 1022–1034. doi: 10.1093/bib/bbx020
Chaverri, P., Branco-Rocha, F., Jaklitsch, W., Gazis, R., Degenkolb, T., and Samuels, G. J. (2015). Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia 107, 558–590. doi: 10.3852/14-147
Chen, D., Hou, Q., Jia, L., and Sun, K. (2021). Combined use of two Trichoderma strains to promote growth of pakchoi (Brassica chinensis L.). Agronomy 11:726. doi: 10.3390/agronomy11040726
Chen, M., Arato, M., Borghi, L., Nouri, E., and Reinhardt, D. (2018). Beneficial services of arbuscular mycorrhizal fungi – from ecology to application. Front. Plant Sci. 9:1270. doi: 10.3389/fpls.2018.01270
Chen, M., Liu, Q., Gao, S. S., Young, A. E., Jacobsen, S. E., and Tang, Y. (2019). Genome mining and biosynthesis of a polyketide from a biofertilizer fungus that can facilitate reductive iron assimilation in plant. Proc. Natl. Acad. Sci. U.S.A. 116, 5499–5504. doi: 10.1073/pnas.1819998116
Cheng, M. J., Wu, M., Der Chen, I. S., Chen, C. Y., Lo, W. L., and Yuan, G. F. (2010). Secondary metabolites from the red mould rice of Monascus purpureus BCRC 38113. Nat. Prod. Res. 24, 1719–1725. doi: 10.1080/14786410902941477
Cheng, Q., Cao, Y., Jiang, C., Xu, L., Wang, M., Zhang, S., et al. (2010). Identifying secreted proteins of Marssonina brunnea by degenerate PCR. Proteomics 10, 2406–2417. doi: 10.1002/pmic.200900844
Chirino-Valle, I., Kandula, D., Littlejohn, C., Hill, R., Walker, M., Shields, M., et al. (2016). Potential of the beneficial fungus Trichoderma to enhance ecosystem-service provision in the biofuel grass Miscanthus × giganteus in agriculture. Sci. Rep. 6, 1–8. doi: 10.1038/srep25109
Contreras-Cornejo, H. A., Macías-Rodríguez, L., Del-Val, E., and Larsen, J. (2016). Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: interactions with plants. FEMS Microbiol. Ecol. 92:fiw036. doi: 10.1093/femsec/fiw036
Costa, J. H., Bazioli, J. M., Barbosa, L. D., Dos Santos Júnior, P. L. T., Reis, F. C. G., Klimeck, T., et al. (2021). Phytotoxic tryptoquialanines produced in vivo by Penicillium digitatum are exported in extracellular vesicles. MBio 12, 1–16. doi: 10.1128/mBio.03393-20
Costa, J. H., Bazioli, J. M., de Vilhena Araújo, E., Vendramini, P. H., de Freitas Porto, M. C., Eberlin, M. N., et al. (2019a). Monitoring indole alkaloid production by Penicillium digitatum during infection process in citrus by Mass Spectrometry Imaging and molecular networking. Fungal Biol. 123, 594–600. doi: 10.1016/j.funbio.2019.03.002
Costa, J. H., Wassano, C. I., Angolini, C. F. F., Scherlach, K., Hertweck, C., and Pacheco Fill, T. (2019b). Antifungal potential of secondary metabolites involved in the interaction between citrus pathogens. Sci. Rep. 9, 1–11. doi: 10.1038/s41598-019-55204-9
Daguerre, Y., Edel-Hermann, V., and Steinberg, C. (2017). “Fungal genes and metabolites associated with the biocontrol of soil-borne plant pathogenic fungi,” in Fungal Metabolites, eds J. M. Mérillon and K. G. Ramawat (New York, NY: Springer International Publishing), 33–104.
Dara, S. K. (2019). The new integrated pest management paradigm for the modern age. J. Integr. Pest Manag. 10, 1–9. doi: 10.1093/jipm/pmz010
Davis, V. M., and Stack, M. E. (1994). Evaluation of alternariol and alternariol methyl ether for mutagenic activity in Salmonella typhimurium. Appl. Environ. Microbiol. 60, 3901–3902. doi: 10.1128/aem.60.10.3901-3902.1994
Dayan, F. E., Cantrell, C. L., and Duke, S. O. (2009). Natural products in crop protection. Bioorganic Med. Chem. 17, 4022–4034. doi: 10.1016/j.bmc.2009.01.046
De Cesare, G. B., Cristy, S. A., Garsin, D. A., and Lorenz, M. C. (2020). Antimicrobial peptides: a new frontier in antifungal therapy. MBio 11, 1–21. doi: 10.1128/mBio.02123-20
Degenkolb, T., Dieckmann, R., Nielsen, K. F., Gräfenhan, T., Theis, C., Zafari, D., et al. (2008a). The Trichoderma Brevicompactum clade: a separate lineage with new species, new peptaibiotics, and mycotoxins. Mycol. Prog. 7, 177–219. doi: 10.1007/s11557-008-0563-3
Degenkolb, T., von Döhren, H., Nielsen, K. F., Samuels, G. J., and Brückner, H. (2008b). Recent advances and future prospects in peptaibiotics, hydrophobin, and mycotoxin research, and their importance for chemotaxonomy of Trichoderma and Hypocrea. Chem. Biodivers. 5, 671–680. doi: 10.1002/cbdv.200890064
Depotter, J. R. L., Seidl, M. F., Wood, T. A., and Thomma, B. P. H. J. (2016). Interspecific hybridization impacts host range and pathogenicity of filamentous microbes. Curr. Opin. Microbiol. 32, 7–13. doi: 10.1016/j.mib.2016.04.005
Derntl, C., Guzmán-Chávez, F., Mello-de-Sousa, T. M., Busse, H. J., Driessen, A. J. M., Mach, R. L., et al. (2017). In vivo study of the sorbicillinoid gene cluster in Trichoderma reesei. Front. Microbiol. 8:2037. doi: 10.3389/fmicb.2017.02037
Dhillon, B., Feau, N., Aerts, A. L., Beauseigle, S., Bernier, L., Copeland, A., et al. (2015). Horizontal gene transfer and gene dosage drives adaptation to wood colonization in a tree pathogen. Proc. Natl. Acad. Sci. U.S.A. 112, 3451–3456. doi: 10.1073/pnas.1424293112
Dias, D. A., Urban, S., and Roessner, U. (2012). A historical overview of natural products in drug discovery. Metabolites 2, 303–336. doi: 10.3390/metabo2020303
Donzelli, B. G. G., Krasnoff, S. B., Churchill, A. C. L., Vandenberg, J. D., and Gibson, D. M. (2010). Identification of a hybrid PKS-NRPS required for the biosynthesis of NG-391 in Metarhizium robertsii. Curr. Genet. 56, 151–162. doi: 10.1007/s00294-010-0288-0
Dotson, B. R., Soltan, D., Schmidt, J., Areskoug, M., Rabe, K., Swart, C., et al. (2018). The antibiotic peptaibol alamethicin from Trichoderma permeabilises Arabidopsis root apical meristem and epidermis but is antagonised by cellulase-induced resistance to alamethicin. BMC Plant Biol. 18:165. doi: 10.1186/s12870-018-1370-x
Doughari, J. (2015). The occurrence, properties and significance of citrinin mycotoxin. J. Plant Pathol. Microbiol. 6:321. doi: 10.4172/2157-7471.1000321
Drenth, A., McTaggart, A. R., and Wingfield, B. D. (2019). Fungal clones win the battle, but recombination wins the war. IMA Fungus 10, 1–6. doi: 10.1186/s43008-019-0020-8
Druzhinina, I., and Kubicek, C. P. (2005). Species concepts and biodiversity in Trichoderma and Hypocrea: from aggregate species to species clusters. J. Zhejiang Univ. Sci. 6B, 100–112. doi: 10.1631/jzus.2005.B0100
Druzhinina, I. S., Chenthamara, K., Zhang, J., Atanasova, L., Yang, D., Miao, Y., et al. (2018). Massive lateral transfer of genes encoding plant cell wall-degrading enzymes to the mycoparasitic fungus Trichoderma from its plant-associated hosts. PLoS Genet. 14:e1007322. doi: 10.1371/journal.pgen.1007322
Druzhinina, I. S., Komoń-Zelazowska, M., Atanasova, L., Seidl, V., and Kubicek, C. P. (2010). Evolution and ecophysiology of the industrial producer Hypocrea jecorina (Anamorph Trichoderma reesei) and a new sympatric agamospecies related to it. PLoS ONE 5:e9191. doi: 10.1371/journal.pone.0009191
Druzhinina, I. S., Komoń-Zelazowska, M., Kredics, L., Hatvani, L., Antal, Z., Belayneh, T., et al. (2008). Alternative reproductive strategies of Hypocrea orientalis and genetically close but clonal Trichoderma Iongibrachiatum, both capable of causing invasive mycoses of humans. Microbiology 154, 3447–3459. doi: 10.1099/mic.0.2008/021196-0
Druzhinina, I. S., and Kubicek, C. P. (2017). Genetic engineering of Trichoderma reesei cellulases and their production. Microb. Biotechnol. 10, 1485–1499. doi: 10.1111/1751-7915.12726
Druzhinina, I. S., Seidl-Seiboth, V., Herrera-Estrella, A., Horwitz, B. A., Kenerley, C. M., Monte, E., et al. (2011). Trichoderma: the genomics of opportunistic success. Nat. Rev. Microbiol. 9, 749–759. doi: 10.1038/nrmicro2637
Dubos, R. J. (1939). Studies on a bactericidal agent extracted from a soil bacillus: I. Preparation of the agent. Its activity in vitro. J. Exp. Med. 70, 1–10. doi: 10.1084/jem.70.1.1
Dubos, R. J., and Cattaneo, C. (1939). Studies on a bactericidal agent extracted from a soil bacillus iii. Preearation and activity of a protein-free fraction. J. Exp. Med. 70, 249–256. doi: 10.1084/jem.70.3.249
Dührkop, K., Fleischauer, M., Ludwig, M., Aksenov, A. A., Melnik, A. V., Meusel, M., et al. (2019). SIRIUS 4: a rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 16, 299–302. doi: 10.1038/s41592-019-0344-8
Ebead, G. A., Overy, D. P., Berrué, F., and Kerr, R. G. (2012). Westerdykella reniformis sp. nov., producing the antibiotic metabolites melinacidin IV and chetracin B. IMA Fungus 3, 189–201. doi: 10.5598/imafungus.2012.03.02.11
El-Sayed, A. M., Mitchell, V. J., and Suckling, D. M. (2014). 6-Pentyl-2H-pyran-2-one: a potent peach-derived kairomone for New Zealand Flower Thrips, Thrips obscuratus. J. Chem. Ecol. 40, 50–55. doi: 10.1007/s10886-014-0379-3
Endo, A., Hasumi, K., Yamada, A., Shimoda, R., and Takeshima, H. (1986). The synthesis of compactin (ml-236b) and monacolin k in fungi. J. Antibiot. 39, 1609–1610. doi: 10.7164/antibiotics.39.1609
Evidente, A., Cimmino, A., Andolfi, A., Vurro, M., Zonno, M. C., Cantrell, C. L., et al. (2008). Phyllostictines A-D, oxazatricycloalkenones produced by Phyllosticta cirsii, a potential mycoherbicide for Cirsium arvense biocontrol. Tetrahedron 64, 1612–1619. doi: 10.1016/j.tet.2007.12.010
Fanelli, F., Liuzzi, V. C., Logrieco, A. F., and Altomare, C. (2018). Genomic characterization of Trichoderma atrobrunneum (T. harzianum species complex) ITEM 908: insight into the genetic endowment of a multi-target biocontrol strain. BMC Genomics 19:662. doi: 10.1186/s12864-018-5049-3
Faure, D. (2002). The family-3 glycoside hydrolases: from housekeeping functions to host-microbe interactions. Appl. Environ. Microbiol. 68, 1485–1490. doi: 10.1128/AEM.68.4.1485-1490.2002
Fedorova, N. D., Moktali, V., and Medema, M. H. (2012). Bioinformatics approaches and software for detection of secondary metabolic gene clusters. Methods Mol. Biol. 944, 23–45. doi: 10.1007/978-1-62703-122-6_2
Felnagle, E. A., Jackson, E. E., Chan, Y. A., Podevels, A. M., Berti, A. D., McMahon, M. D., et al. (2008). Nonribosomal peptide synthetases involved in the production of medically relevant natural products. Mol. Pharm. 5, 191–211. doi: 10.1021/mp700137g
Finking, R., and Marahiel, M. A. (2004). Biosynthesis of nonribosomal peptides. Annu. Rev. Microbiol. 58, 453–488. doi: 10.1146/annurev.micro.58.030603.123615
Flissi, A., Ricart, E., Campart, C., Chevalier, M., Dufresne, Y., Michalik, J., et al. (2020). Norine: update of the nonribosomal peptide resource. Nucleic Acids Res. 48, D465–D469. doi: 10.1093/nar/gkz1000
Fravel, D. R. (1988). Role of antibiosis in the biocontrol of plant diseases. Annu. Rev. Phytopathol. 26, 75–91.
Fujita, T., Wada, S. I., Iida, A., Nishimura, T., Kanai, M., and Toyama, N. (1994). Fungal metabolites. XIII. Isolation and structural elucidation of new peptaibols, Trichodecenins-I and -II, from Trichoderma viride. Chem. Pharm. Bull. 42, 489–494. doi: 10.1248/cpb.42.489
Gao, X., Chooi, Y. H., Ames, B. D., Wang, P., Walsh, C. T., and Tang, Y. (2011). Fungal indole alkaloid biosynthesis: genetic and biochemical investigation of the tryptoquialanine pathway in Penicillium aethiopicum. J. Am. Chem. Soc. 133, 2729–2741. doi: 10.1021/ja1101085
Gauthier, T., Wang, X., Dos Santos, J., Fysikopoulos, A., Tadrist, S., Canlet, C., et al. (2012). Trypacidin, a spore-borne toxin from Aspergillus fumigatus, is cytotoxic to lung cells. PLoS ONE 7:e0029906. doi: 10.1371/journal.pone.0029906
Geisler, K., Hughes, R. K., Sainsbury, F., Lomonossoff, G. P., Rejzek, M., Fairhurst, S., et al. (2013). Biochemical analysis of a multifunctional cytochrome P450 (CYP51) enzyme required for synthesis of antimicrobial triterpenes in plants. Proc. Natl. Acad. Sci. U.S.A. 110, E3360–E3367. doi: 10.1073/pnas.1309157110
Gewin, V. (2003). Genetically modified corn - environmental benefits and risks. PLoS Biol. 1:e8. doi: 10.1371/journal.pbio.0000008
Ghorbanpour, M., Omidvari, M., Abbaszadeh-Dahaji, P., Omidvar, R., and Kariman, K. (2018). Mechanisms underlying the protective effects of beneficial fungi against plant diseases. Biol. Control 117, 147–157. doi: 10.1016/j.biocontrol.2017.11.006
Gibson, D. J., Connolly, J., Hartnett, D. C., and Weidenhamer, J. D. (1999). Designs for greenhouse studies of interactions between plants. J. Ecol. 87, 1–16. doi: 10.1046/j.1365-2745.1999.00321.x
Goldman, G. H., Van Montagu, M., and Herrera-Estrella, A. (1990). Transformation of Trichoderma harzianum by high-voltage electric pulse. Curr. Genet. 17, 169–174. doi: 10.1007/BF00312863
Golo, P. S., Gardner, D. R., Grilley, M. M., Takemoto, J. Y., Krasnoff, S. B., Pires, M. S., et al. (2014). Production of destruxins from Metarhizium spp. fungi in artificial medium and in endophytically colonized cowpea plants. PLoS ONE 9:e104946. doi: 10.1371/journal.pone.0104946
Gomes, A. R., Duarte, A. C., and Rocha-Santos, T. A. P. (2017). “Analytical techniques for discovery of bioactive compounds from marine fungi,” in Fungal Metabolites, eds J. M. Mérillon and K. G. Ramawat (New York, NY: Springer International Publishing), 415–434.
Grigoriev, I. V., Nikitin, R., Haridas, S., Kuo, A., Ohm, R., Otillar, R., et al. (2014). MycoCosm portal: gearing up for 1000 fungal genomes. Nucleic Acids Res. 42, D699–704. doi: 10.1093/nar/gkt1183
Grindrod, P., Parsons, M. C., Higham, D. J., and Estrada, E. (2011). Communicability across evolving networks. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 83:046120. doi: 10.1103/PhysRevE.83.046120
Gruber, F., Visser, J., Kubicek, C. P., and de Graaff, L. H. (1990). The development of a heterologous transformation system for the cellulolytic fungus Trichoderma reesei based on a pyrG-negative mutant strain. Curr. Genet. 18, 71–76. doi: 10.1007/BF00321118
Guo, K., Sui, Y., Li, Z., Huang, Y., and Zhang, H. (2020). Trichoderma viride Tv-1511 colonizes arabidopsis leaves and promotes arabidopsis growth by modulating the MAP kinase 6-mediated activation of plasma membrane H+-ATPase. J. Plant Growth Regul. 39, 1261–1276. doi: 10.1007/s00344-019-10063-6
Guo, Y., Ghirardo, A., Weber, B., Schnitzler, J. P., Philipp Benz, J., and Rosenkranz, M. (2019). Trichoderma species differ in their volatile profiles and in antagonism toward ectomycorrhiza Laccaria bicolor. Front. Microbiol. 10:891. doi: 10.3389/fmicb.2019.00891
Gupta, S., Ellis, S. E., Ashar, F. N., Moes, A., Bader, J. S., Zhan, J., et al. (2014). Transcriptome analysis reveals dysregulation of innate immune response genes and neuronal activity-dependent genes in autism. Nat. Commun. 5, 1–8. doi: 10.1038/ncomms6748
Gutiérrez, S., McCormick, S. P., Cardoza, R. E., Lindo, L., Alexander, N. J., and Proctor, R. H. (2020). “Trichoderma trichothecenes,” in New and Future Developments in Microbial Biotechnology and Bioengineering, eds V. G. Gupta and S. Rodriguez-Couto (New York, NY: Elsevier), 281–301.
Guzmán-Guzmán, P., Alemán-Duarte, M. I., Delaye, L., Herrera-Estrella, A., and Olmedo-Monfil, V. (2017). Identification of effector-like proteins in Trichoderma spp. and role of a hydrophobin in the plant-fungus interaction and mycoparasitism. BMC Genet. 18:16. doi: 10.1186/s12863-017-0481-y
Haas, H. (2014). Fungal siderophore metabolism with a focus on Aspergillus fumigatus. Nat. Prod. Rep. 31, 1266–1276. doi: 10.1039/c4np00071d
Hahn, M. (2014). The rising threat of fungicide resistance in plant pathogenic fungi: botrytis as a case study. J. Chem. Biol. 7, 133–141. doi: 10.1007/s12154-014-0113-1
Halifu, S., Deng, X., Song, X., and Song, R. (2019). Effects of two Trichoderma strains on plant growth, rhizosphere soil nutrients, and fungal community of Pinus sylvestris var. mongolica annual seedlings. Forests 10, 1–17. doi: 10.3390/f10090758
Halliwell, G., and Griffin, M. (1973). The nature and mode of action of the cellulolytic component C1 of Trichoderma koningii on native cellulose. Biochem. J. 135, 587–594. doi: 10.1042/bj1350587
Hamilton, C. E., Dowling, T. E., and Faeth, S. H. (2010). Hybridization in endophyte symbionts alters host response to moisture and nutrient treatments. Microb. Ecol. 59, 768–775. doi: 10.1007/s00248-009-9606-9
Hanson, L. E., and Howell, C. R. (2002). Biocontrol efficacy and other characteristics of protoplast fusants between Trichoderma koningii and T. virens. Mycol. Res. 106, 321–328. doi: 10.1017/S0953756202005592
Hanson, L. E., and Howell, C. R. (2004). Elicitors of plant defense responses from biocontrol strains of Trichoderma virens. Phytopathology 94, 171–176. doi: 10.1094/PHYTO.2004.94.2.171
Hao, Z., and Su, X. (2019). Fast gene disruption in Trichoderma reesei using in vitro assembled Cas9/gRNA complex. BMC Biotechnol. 19. doi: 10.1186/s12896-018-0498-y
Harding, D. P., and Raizada, M. N. (2015). Controlling weeds with fungi, bacteria and viruses: a review. Front. Plant Sci. 6:659. doi: 10.3389/fpls.2015.00659
Harman, G. E. (2006). Overview of mechanisms and uses of Trichoderma spp. Phytopathology 96, 190–194. doi: 10.1094/PHYTO-96-01
Harman, G. E., Howell, C. R., Viterbo, A., Chet, I., and Lorito, M. (2004). Trichoderma species - Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2, 43–56. doi: 10.1038/nrmicro797
Hauser, D., and Sigg, H. P. (1971). Isolierung und Abbau von Sordarin. 1. Mitteilung über Sordarin. Helv. Chim. Acta 54, 1178–1190. doi: 10.1002/hlca.19710540427
Hayat, R., Ali, S., Amara, U., Khalid, R., and Ahmed, I. (2010). Soil beneficial bacteria and their role in plant growth promotion: a review. Ann. Microbiol. 60, 579–598. doi: 10.1007/s13213-010-0117-1
Headrick, D. H., and Goeden, R. D. (2001). Biological control as a tool for ecosystem management. Biol. Control 21, 249–257. doi: 10.1006/bcon.2001.0939
Hertweck, C. (2009). Hidden biosynthetic treasures brought to light. Nat. Chem. Biol. 5, 450–452. doi: 10.1038/nchembio0709-450
Hinterdobler, W., Li, G., Spiegel, K., Basyouni-Khamis, S., Gorfer, M., and Schmoll, M. (2021). Trichoderma reesei isolated from Austrian soil with high potential for biotechnological application. Front. Microbiol. 12:552301. doi: 10.3389/fmicb.2021.552301
Hollomon, D. W. (2015). “Fungicide resistance: 40 years on and still a major problem,” in Fungicide Resistance in Plant Pathogens, eds H. Ishii and D. W. Hollomon (Tokyo: Springer Japan), 3–11.
Howell, C. R. (2003). Mechanisms employed by Trichoderma species in the biological control of plant diseases: the history and evolution of current concepts. Plant Dis. 87, 4–10. doi: 10.1094/PDIS.2003.87.1.4
Huan, Y., Kong, Q., Mou, H., and Yi, H. (2020). Antimicrobial peptides: classification, design, application and research progress in multiple fields. Front. Microbiol. 11:2559. doi: 10.3389/fmicb.2020.582779
Huang, Q., Tezuka, Y., Hatanaka, Y., Kikuchi, T., Nishi, A., and Tubaki, K. (1995). Studies on metabolites of mycoparasitic fungi. IV. minor peptaibols of Trichoderma koningii. Chem. Pharm. Bull. 43, 1663–1667. doi: 10.1248/cpb.43.1663
Huber, F. M. (1967). “Griseofulvin,” in Mechanism of Action, eds D. Gottlieb and P. D. Shaw (Berlin: Springer Berlin Heidelberg), 181–189.
Itoh, Y., Kodama, K., Furuya, K., Takahashi, S., Haneishi, T., Takiguchi, Y., et al. (1980). A new sesquiterpene antibiotic, heptel1dic acid producing organisms, fermentation, isolation and characterization. J. Antibiot. 33, 468–473. doi: 10.7164/antibiotics.33.468
Jalal, M. A. F., Love, S. K., and van der Helm, D. (1988). Nα-Dimethylcoprogens: three novel trihydroxamate siderophores from pathogenic fungi. Biol. Met. 1, 4–8. doi: 10.1007/BF01128011
Jayasuriya, H., Silverman, K. C., Zink, D. L., Jenkins, R. G., Sanchez, M., Pelaez, F., et al. (1998). Clavaric acid: a triterpenoid inhibitor of farnesyl-protein transferase from Clavariadelphus truncatus. J. Nat. Prod. 61, 1568–1570. doi: 10.1021/np980200c
Jia, T., Oberhofer, M., Shymanovich, T., and Faeth, S. H. (2016). Effects of hybrid and non-hybrid Epichloë endophytes and their associated host genotypes on the response of a native grass to varying environments. Microb. Ecol. 72, 185–196. doi: 10.1007/s00248-016-0743-7
Joana Gil-Chávez, G., Villa, J. A., Fernando Ayala-Zavala, J., Basilio Heredia, J., Sepulveda, D., Yahia, E. M., et al. (2013). Technologies for extraction and production of bioactive compounds to be used as nutraceuticals and food ingredients: an overview. Compr. Rev. Food Sci. Food Saf. 12, 5–23. doi: 10.1111/1541-4337.12005
Kaewchai, S., Soytong, K., and Hyde, K. D. (2009). Mycofungicides and fungal biofertilizers. Fungal Divers. 38, 25–50.
Keller, N. P. (2019). Fungal secondary metabolism: regulation, function and drug discovery. Nat. Rev. Microbiol. 17, 167–180. doi: 10.1038/s41579-018-0121-1
Keller, N. P., Turner, G., and Bennett, J. W. (2005). Fungal secondary metabolism - from biochemistry to genomics. Nat. Rev. Microbiol. 3, 937–947. doi: 10.1038/nrmicro1286
Kenshole, E., Herisse, M., Michael, M., and Pidot, S. J. (2021). Natural product discovery through microbial genome mining. Curr. Opin. Chem. Biol. 60, 47–54. doi: 10.1016/j.cbpa.2020.07.010
Keswani, C., Mishra, S., Sarma, B. K., Singh, S. P., and Singh, H. B. (2014). Unraveling the efficient applications of secondary metabolites of various Trichoderma spp. Appl. Microbiol. Biotechnol. 98, 533–544. doi: 10.1007/s00253-013-5344-5
Keswani, C., Singh, H. B., Hermosa, R., García-Estrada, C., Caradus, J., He, Y. W., et al. (2019). Antimicrobial secondary metabolites from agriculturally important fungi as next biocontrol agents. Appl. Microbiol. Biotechnol. 103, 9287–9303. doi: 10.1007/s00253-019-10209-2
Khaldi, N., Collemare, J., Lebrun, M. H., and Wolfe, K. H. (2008). Evidence for horizontal transfer of a secondary metabolite gene cluster between fungi. Genome Biol. 9:R18. doi: 10.1186/gb-2008-9-1-r18
Khan, I., Xie, W. L., Yu, Y. C., Sheng, H., Xu, Y., Wang, J. Q., et al. (2020). Heteroexpression of Aspergillus nidulans laeA in marine-derived fungi triggers upregulation of secondary metabolite biosynthetic genes. Mar. Drugs 18:652. doi: 10.3390/md18120652
Kim, J., and Movassaghi, M. (2015). Biogenetically-inspired total synthesis of epidithiodiketopiperazines and related alkaloids. Acc. Chem. Res. 48, 1159–1171. doi: 10.1021/ar500454v
Kingston, D. G. I. (2011). Modern natural products drug discovery and its relevance to biodiversity conservation. J. Nat. Prod. 74, 496–511. doi: 10.1021/np100550t
Koch, M. S., Ward, J. M., Levine, S. L., Baum, J. A., Vicini, J. L., and Hammond, B. G. (2015). The food and environmental safety of Bt crops. Front. Plant Sci. 6:283. doi: 10.3389/fpls.2015.00283
Köhl, J., Kolnaar, R., and Ravensberg, W. J. (2019). Mode of action of microbial biological control agents against plant diseases: relevance beyond efficacy. Front. Plant Sci. 10:845. doi: 10.3389/fpls.2019.00845
Krasnoff, S. B., Keresztes, I., Gillilan, R. E., Szebenyi, D. M. E., Donzelli, B. G. G., Churchill, A. C. L., et al. (2007). Serinocyclins A and B, cyclic heptapeptides from Metarhizium anisopliae. J. Nat. Prod. 70, 1919–1924. doi: 10.1021/np070407i
Kroken, S., Glass, N. L., Taylor, J. W., Yoder, O. C., and Turgeon, B. G. (2003). Phylogenomic analysis of type I polyketide synthase genes in pathogenic and saprobic ascomycetes. Proc. Natl. Acad. Sci. U.S.A. 100, 15670–15675. doi: 10.1073/pnas.2532165100
Krupke, O. A., Castle, A. J., and Rinker, D. L. (2003). The North American mushroom competitor, Trichoderma aggressivum cf. aggressivum, produces antifungal compounds in mushroom compost that inhibit mycelial growth of the commercial mushroom Agaricus bisporus. Mycol. Res. 107, 1467–1475. doi: 10.1017/S0953756203008621
Kubicek, C. P., Herrera-Estrella, A., Seidl-Seiboth, V., Martinez, D. A., Druzhinina, I. S., Thon, M., et al. (2011). Comparative genome sequence analysis underscores mycoparasitism as the ancestral life style of Trichoderma. Genome Biol. 12, 1–15. doi: 10.1186/gb-2011-12-4-r40
Kubicek, C. P., Steindorff, A. S., Chenthamara, K., Manganiello, G., Henrissat, B., Zhang, J., et al. (2019). Evolution and comparative genomics of the most common Trichoderma species. BMC Genomics 20:485. doi: 10.1186/s12864-019-5680-7
Kudo, F., Matsuura, Y., Hayashi, T., Fukushima, M., and Eguchi, T. (2016). Genome mining of the sordarin biosynthetic gene cluster from Sordaria araneosa Cain ATCC 36386: Characterization of cycloaraneosene synthase and GDP-6-deoxyaltrose transferase. J. Antibiot. 69, 541–548. doi: 10.1038/ja.2016.40
Kulimushi, S. M., Muiru, W. M., and Mutitu, E. W. (2021). Potential of Trichoderma spp., Bacillus subtilis and Pseudomonas fluorescens in the management of early blight in tomato. Biocontrol Sci. Technol. doi: 10.1080/09583157.2021.1900784. [Epub ahead of print].
Kumar, M., and Ashraf, S. (2017). “Role of Trichoderma spp. as a biocontrol agent of fungal plant pathogens,” in Probiotics and Plant Health, eds V. Kumar, M. Kumar, S. Sharma, and R. Prasad (Singapore: Springer), 497–506.
Lagashetti, A. C., Dufossé, L., Singh, S. K., and Singh, P. N. (2019). Fungal pigments and their prospects in different industries. Microorganisms 7:604. doi: 10.3390/microorganisms7120604
Lahlali, R., and Hijri, M. (2010). Screening, identification and evaluation of potential biocontrol fungal endophytes against Rhizoctonia solani AG3 on potato plants. FEMS Microbiol. Lett. 311, 152–159. doi: 10.1111/j.1574-6968.2010.02084.x
Landeis, A., and Schmidt-Heydt, M. (2021). Sequencing and analysis of the entire genome of the mycoparasitic fungus Trichoderma afroharzianum. Microbiol. Resour. Announc. 10:e00211-21. doi: 10.1128/mra.00211-21
Lawrence, D. P., Kroken, S., Pryor, B. M., and Arnold, A. E. (2011). Interkingdom gene transfer of a hybrid NPS/PKS from bacteria to filamentous ascomycota. PLoS ONE 6:e28231. doi: 10.1371/journal.pone.0028231
Lawrence, T. J., Carper, D. L., Spangler, M. K., Carrell, A. A., Rush, T. A., Minter, S. J., et al. (2020). amPEPpy 1.0: a portable and accurate antimicrobial peptide prediction tool. Bioinformatics 3, 1–3. doi: 10.1093/bioinformatics/btaa917
Lebe, K. E., and Cox, R. J. (2019). Oxidative steps during the biosynthesis of squalestatin S1. Chem. Sci. 10, 1227–1231. doi: 10.1039/c8sc02615g
Lee, S., Yap, M., Behringer, G., Hung, R., and Bennett, J. W. (2016). Volatile organic compounds emitted by Trichoderma species mediate plant growth. Fungal Biol. Biotechnol. 3, 1–14. doi: 10.1186/s40694-016-0025-7
Li, D., Tang, Y., Lin, J., and Cai, W. (2017). Methods for genetic transformation of filamentous fungi. Microb. Cell Fact. 16:168. doi: 10.1186/s12934-017-0785-7
Li, J., Wu, Y., Chen, K., Wang, Y., Hu, J., Wei, Y., et al. (2018). Trichoderma cyanodichotomus sp. nov., a new soil-inhabiting species with a potential for biological control. Can. J. Microbiol. 64, 1020–1029. doi: 10.1139/cjm-2018-0224
Li, N., Alfiky, A., Wang, W., Islam, M., Nourollahi, K., Liu, X., et al. (2018). Volatile compound-mediated recognition and inhibition between Trichoderma biocontrol agents and Fusarium oxysporum. Front. Microbiol. 9:2614. doi: 10.3389/fmicb.2018.02614
Li, M. F., Li, G. H., and Zhang, K. Q. (2019). Non-volatile metabolites from Trichoderma spp. Metabolites 9:58. doi: 10.3390/metabo9030058
Liu, R., Chen, L., Jiang, Y., Zhou, Z., and Zou, G. (2015). Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discov. 1:15007. doi: 10.1038/celldisc.2015.7
Lorito, M., Woo, S. L., Harman, G. E., and Monte, E. (2010). Translational research on Trichoderma: from'omics to the field. Annu. Rev. Phytopathol. 48, 395–417. doi: 10.1146/annurev-phyto-073009-114314
Lu, S., Tian, J., Sun, W., Meng, J., Wang, X., Fu, X., et al. (2014). Bis-naphtho-γ-pyrones from fungi and their bioactivities. Molecules 19, 7169–7188. doi: 10.3390/molecules19067169
Ma, B., Zhang, K., Hendrie, C., Liang, C., Li, M., Doherty-Kirby, A., et al. (2003). PEAKs: Powerful software for peptide de novo sequencing by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 17, 2337–2342. doi: 10.1002/rcm.1196
Ma, L. J., and Fedorova, N. D. (2010). A practical guide to fungal genome projects: strategy, technology, cost and completion. Mycology 1, 9–24. doi: 10.1080/21501201003680943
Macheleidt, J., Mattern, D. J., Fischer, J., Netzker, T., Weber, J., Schroeckh, V., et al. (2016). Regulation and role of fungal secondary metabolites. Annu. Rev. Genet. 50, 371–392. doi: 10.1146/annurev-genet-120215-035203
Macias, F. A., Varela, R. M., Simonet, A. M., Cutler, H. G., Cutler, S. J., Eden, M. A., et al. (2000). Bioactive carotanes from Trichoderma virens. J. Nat. Prod. 63, 1197–1200. doi: 10.1021/np000121c
Mahlapuu, M., Håkansson, J., Ringstad, L., and Björn, C. (2016). Antimicrobial peptides: an emerging category of therapeutic agents. Front. Cell. Infect. Microbiol. 6:194. doi: 10.3389/fcimb.2016.00194
Mahood, E. H., Kruse, L. H., and Moghe, G. D. (2020). Machine learning: a powerful tool for gene function prediction in plants. Appl. Plant Sci. 8. doi: 10.1002/aps3.11376
Malmierca, M. G., Izquierdo-Bueno, I., Mccormick, S. P., Cardoza, R. E., Alexander, N. J., Moraga, J., et al. (2016). Botrydial and botcinins produced by Botrytis cinerea regulate the expression of Trichoderma arundinaceum genes involved in trichothecene biosynthesis. Mol. Plant Pathol. 17, 1017–1031. doi: 10.1111/mpp.12343
Manczinger, L., Komonyi, O., Antal, Z., and Ferenczy, L. (1997). A method for high-frequency transformation of Trichoderma viride. J. Microbiol. Methods 29, 207–210. doi: 10.1016/S0167-7012(97)00026-2
Marfori, E. C., Kajiyama, S., Fukusaki, E. I., and Kobayashi, A. (2002). Trichosetin, a novel tetramic acid antibiotic produced in dual culture of Trichoderma harzianum and Catharanthus roseus callus. Zeitschrift Naturforsch. Sect. C J. Biosci. 57, 465–470. doi: 10.1515/znc-2002-5-611
Martinez, D., Berka, R. M., Henrissat, B., Saloheimo, M., Arvas, M., Baker, S. E., et al. (2008). Genome sequencing and analysis of the biomass-degrading fungus Trichoderma reesei (syn. Hypocrea jecorina). Nat. Biotechnol. 26, 553–560. doi: 10.1038/nbt1403
Masi, M., Meyer, S., Górecki, M., Mandoli, A., Di Bari, L., Pescitelli, G., et al. (2017). Pyriculins A and B, two monosubstituted hex-4-ene-2,3-diols and other phytotoxic metabolites produced by Pyricularia grisea isolated from buffelgrass (Cenchrus ciliaris). Chirality 29, 726–736. doi: 10.1002/chir.22744
Matsumoto, M., Matsutani, S., Sugita, K., Yoshida, H., Hayashi, F., Terui, Y., et al. (1992). Depudecin: a novel compound inducing the flat phenotypi of nih3t3 cells doubly transformed by ras-and src-oncogene, produced by Alternaria brassicicola. J. Antibiot. 45, 879–885. doi: 10.7164/antibiotics.45.879
McInnes, A. G., Smith, D. G., Wat, C. K., Vining, L. C., and Wright, J. L. C. (1974). Tenellin and bassianin, metabolites of Beauveria species. Structure elucidation with 15N- and doubly 13C-enriched compounds using 13C nuclear magnetic resonance spectroscopy. J. Chem. Soc. Chem. Commun. 8, 281–282. doi: 10.1039/C39740000281
McMullin, D. R., Renaud, J. B., Barasubiye, T., Sumarah, M. W., and Miller, J. D. (2017). Metabolites of Trichoderma species isolated from damp building materials. Can. J. Microbiol. 63, 621–632. doi: 10.1139/cjm-2017-0083
Meca, G., Soriano, J. M., Gaspari, A., Ritieni, A., Moretti, A., and Mañes, J. (2010). Antifungal effects of the bioactive compounds enniatins A, A1, B, B1. Toxicon 56, 480–485. doi: 10.1016/j.toxicon.2010.04.013
Meena, M., Swapnil, P., Zehra, A., Aamir, M., Dubey, M. K., Goutam, J., et al. (2017). “Beneficial microbes for disease suppression and plant growth promotion,” in Plant-Microbe Interactions in Agro-Ecological Perspectives, eds D. Singh, H. Singh, and R. Prabha (Singapore: Springer), 395–432.
Mendoza-Mendoza, A., Zaid, R., Lawry, R., Hermosa, R., Monte, E., Horwitz, B. A., et al. (2018). Molecular dialogues between Trichoderma and roots: role of the fungal secretome. Fungal Biol. Rev. 32, 62–85. doi: 10.1016/j.fbr.2017.12.001
Meng, J., Wang, X., Xu, D., Fu, X., Zhang, X., Lai, D., et al. (2016). Sorbicillinoids from fungi and their bioactivities. Molecules 21:715. doi: 10.3390/molecules21060715
Migheli, Q., González-Candelas, L., Dealessi, L., Camponogara, A., and Ramón-Vidal, D. (1998). Transformants of Trichoderma longibrachiatum overexpressing the β-1,4-endoglucanase gene egl1 show enhanced biocontrol of Pythium ultimum on cucumber. Phytopathology 88, 673–677. doi: 10.1094/PHYTO.1998.88.7.673
Mistry, J., Chuguransky, S., Williams, L., Qureshi, M., Salazar, G. A., Sonnhammer, E. L. L., et al. (2021). Pfam: the protein families database in 2021. Nucleic Acids Res. 49, D412–D419. doi: 10.1093/nar/gkaa913
Montesinos, E. (2007). Antimicrobial peptides and plant disease control. FEMS Microbiol. Lett. 270, 1–11. doi: 10.1111/j.1574-6968.2007.00683.x
Moore, J. H., Davis, N. D., and Diener, U. L. (1972). Mellein and 4-hydroxymellein production by Aspergillus ochraceus Wilhelm. Appl. Microbiol. 23, 1067–1072. doi: 10.1128/aem.23.6.1067-1072.1972
Mukherjee, P. K., Horwitz, B. A., and Kenerley, C. M. (2012). Secondary metabolism in Trichoderma- a genomic perspective. Microbiology 158, 35–45. doi: 10.1099/mic.0.053629-0
Mukherjee, P. K., Hurley, J. F., Taylor, J. T., Puckhaber, L., Lehner, S., Druzhinina, I., et al. (2018). Ferricrocin, the intracellular siderophore of Trichoderma virens, is involved in growth, conidiation, gliotoxin biosynthesis and induction of systemic resistance in maize. Biochem. Biophys. Res. Commun. 505, 606–611. doi: 10.1016/j.bbrc.2018.09.170
Mukhopadhyay, R., and Kumar, D. (2020). Trichoderma: a beneficial antifungal agent and insights into its mechanism of biocontrol potential. Egypt. J. Biol. Pest Control 30, 1–8. doi: 10.1186/s41938-020-00333-x
Neumann, N. K. N., Stoppacher, N., Zeilinger, S., Degenkolb, T., Brückner, H., and Schuhmacher, R. (2015). The peptaibiotics database - a comprehensive online resource. Chem. Biodivers. 12, 743–751. doi: 10.1002/cbdv.201400393
Newman, D. J., and Cragg, G. M. (2020). Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 83, 770–803. doi: 10.1021/acs.jnatprod.9b01285
Newman, M. A., Sundelin, T., Nielsen, J. T., and Erbs, G. (2013). MAMP (microbe-associated molecular pattern) triggered immunity in plants. Front. Plant Sci. 4:139. doi: 10.3389/fpls.2013.00139
Niehaus, E. M., Kleigrewe, K., Wiemann, P., Studt, L., Sieber, C. M. K., Connolly, L. R., et al. (2013). Genetic manipulation of the Fusarium fujikuroi fusarin gene cluster yields insight into the complex regulation and fusarin biosynthetic pathway. Chem. Biol. 20, 1055–1066. doi: 10.1016/j.chembiol.2013.07.004
Nielsen, K. F., Gräfenhan, T., Zafari, D., and Thrane, U. (2005). Trichothecene production by Trichoderma brevicompactum. J. Agric. Food Chem. 53, 8190–8196. doi: 10.1021/jf051279b
Oberhofer, M., Güsewell, S., and Leuchtmann, A. (2014). Effects of natural hybrid and non-hybrid Epichloë endophytes on the response of Hordelymus europaeus to drought stress. New Phytol. 201, 242–253. doi: 10.1111/nph.12496
Odendaal, A. Y., Trader, D. J., and Carlson, E. E. (2011). Chemoselective enrichment for natural products discovery. Chem. Sci. 2, 760–764. doi: 10.1039/c0sc00620c
Ogawa, T., Ando, K., Aotani, Y., Shinoda, K., Tanaka, T., Tsukuda, E., et al. (1995). RES-1214-1 and-2, novel non-peptidic endothelin type A receptor antagonists produced by Pestalotiopsis sp. J. Antibiot. 48, 1401–1406. doi: 10.7164/antibiotics.48.1401
Ons, L., Bylemans, D., Thevissen, K., and Cammue, B. P. A. (2020). Combining biocontrol agents with chemical fungicides for integrated plant fungal disease control. Microorganisms 8, 1–19. doi: 10.3390/microorganisms8121930
Page, L., and Brin, S. (1998). The anatomy of a large-scale hypertextual Web search engine. Comput. Networks 30, 107–117. doi: 10.1016/s0169-7552(98)00110-x
Parker, S. R., Cutler, H. G., Jacyno, J. M., and Hill, R. A. (1997). Biological activity of 6-pentyl-2H-pyran-2-one and its analogs. J. Agric. Food Chem. 45, 2774–2776. doi: 10.1021/jf960681a
Patron, N. J., Waller, R. F., Cozijnsen, A. J., Straney, D. C., Gardiner, D. M., Nierman, W. C., et al. (2007). Origin and distribution of epipolythiodioxopiperazine (ETP) gene clusters in filamentous ascomycetes. BMC Evol. Biol. 7, 1–15. doi: 10.1186/1471-2148-7-174
Pedras, M. S. C., Irina Zaharia, L. I., and Ward, D. E. (2002). The destruxins: synthesis, biosynthesis, biotransformation, and biological activity. Phytochemistry 59, 579–596. doi: 10.1016/S0031-9422(02)00016-X
Perelló, A., Mónaco, C., Simón, M. R., Sisterna, M., and Dal Bello, G. (2003). Biocontrol efficacy of Trichoderma isolates for tan spot of wheat in Argentina. Crop Prot. 22, 1099–1106. doi: 10.1016/S0261-2194(03)00143-1
Pfordt, A., Schiwek, S., Karlovsky, P., and von Tiedemann, A. (2020). Trichoderma afroharzianum Ear Rot–a new disease on maize in Europe. Front. Agron. 2, 547758. doi: 10.3389/fagro.2020.547758
Pham, N., van Heck, R. G. A., van Dam, J. C. J., Schaap, P. J., Saccenti, E., and Suarez-Diez, M. (2019). Consistency, inconsistency, and ambiguity of metabolite names in biochemical databases used for genome-scale metabolic modelling. Metabolites 9:28. doi: 10.3390/metabo9020028
Pimentel, D., Zuniga, R., and Morrison, D. (2005). Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 52, 273–288. doi: 10.1016/j.ecolecon.2004.10.002
Piñeiro, V., Arias, J., Dürr, J., Elverdin, P., Ibáñez, A. M., Kinengyere, A., et al. (2020). A scoping review on incentives for adoption of sustainable agricultural practices and their outcomes. Nat. Sustain. 3, 809–820. doi: 10.1038/s41893-020-00617-y
Proctor, R. H., McCormick, S. P., Kim, H. S., Cardoza, R. E., Stanley, A. M., Lindo, L., et al. (2018). Evolution of structural diversity of trichothecenes, a family of toxins produced by plant pathogenic and entomopathogenic fungi. PLoS Pathog. 14:e1006946. doi: 10.1371/journal.ppat.1006946
Proksa, B., Uhrín, D., Liptaj, T., and Šturdíková, M. (1998). Neosartorin, an ergochrome biosynthesized by Neosartorya fischeri. Phytochemistry 48, 1161–1164. doi: 10.1016/S0031-9422(98)00169-1
Prosperini, A., Berrada, H., Ruiz, M. J., Caloni, F., Coccini, T., Spicer, L. J., et al. (2017). A review of the mycotoxin enniatin B. Front. Public Heal. 5:304. doi: 10.3389/fpubh.2017.00304
Pusztahelyi, T., Holb, I. J., and Pócsi, I. (2015). Secondary metabolites in fungus-plant interactions. Front. Plant Sci. : 573. doi: 10.3389/fpls.2015.00573
Reddy, G. C., Goyal, R. K., Puranik, S., Waghmar, V., Vikram, K. V., and Sruthy, K. S. (2020). “Biofertilizers toward sustainable agricultural development,” in Plant Microbe Symbiosis, eds A. Varma, S. Tripathi, and R. Prasad (Cham: Springer International Publishing), 115–128.
Reino, J. L., Guerrero, R. F., Hernández-Galán, R., and Collado, I. G. (2008). Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 7, 89–123. doi: 10.1007/s11101-006-9032-2
Reynolds, H. T., Slot, J. C., Divon, H. H., Lysøe, E., Proctor, R. H., and Brown, D. W. (2017). Differential retention of gene functions in a secondary metabolite cluster. Mol. Biol. Evol. 34, 2002–2015. doi: 10.1093/molbev/msx145
Rivera-Chávez, J., Raja, H. A., Graf, T. N., Gallagher, J. M., Metri, P., Xue, D., et al. (2017). Prealamethicin F50 and related peptaibols from Trichoderma arundinaceum: validation of their authenticity via in situ chemical analysis. RSC Adv. 7, 45733–45741. doi: 10.1039/c7ra09602j
Roberts, L. D., Souza, A. L., Gerszten, R. E., and Clish, C. B. (2012). Targeted metabolomics. Curr. Protoc. Mol. Biol. 1:Unit30.2. doi: 10.1002/0471142727.mb3002s98
Rojo, F. G., Reynoso, M. M., Ferez, M., Chulze, S. N., and Torres, A. M. (2007). Biological control by Trichoderma species of Fusarium solani causing peanut brown root rot under field conditions. Crop Prot. 26, 549–555. doi: 10.1016/j.cropro.2006.05.006
Rokas, A., Mead, M. E., Steenwyk, J. L., Raja, H. A., and Oberlies, N. H. (2020). Biosynthetic gene clusters and the evolution of fungal chemodiversity. Nat. Prod. Rep. 37, 868–878. doi: 10.1039/c9np00045c
Rossman, A. Y. (2009). “The impact of invasive fungi on agricultural ecosystems in the United States,” in Ecological Impacts of Non-Native Invertebrates and Fungi on Terrestrial Ecosystems, eds D. W. Langor, and J. Sweeney (Dordrecht: Springer Netherlands), 97–107.
Rouphael, Y., and Colla, G. (2020). Editorial: biostimulants in agriculture. Front. Plant Sci. 11:40. doi: 10.3389/fpls.2020.00040
Rubio, M. B., Quijada, N. M., Pérez, E., Domínguez, S., Monte, E., and Hermosa, R. (2014). Identifying beneficial qualities of 831 Trichoderma parareesei for plants. Appl. Environ. Microbiol. 80, 1864–1873. doi: 10.1128/AEM.03375-13
Rychen, G., Aquilina, G., Azimonti, G., Bampidis, V., Bastos, M., de, L., et al. (2018). Safety and efficacy of muramidase from Trichoderma reesei DSM 32338 as a feed additive for chickens for fattening and minor poultry species. EFSA J. 16:5342. doi: 10.2903/j.efsa.2018.5342
Saari, S., and Faeth, S. H. (2012). Hybridization of Neotyphodium endophytes enhances competitive ability of the host grass. New Phytol. 195, 231–236. doi: 10.1111/j.1469-8137.2012.04140.x
Sánchez-Torres, P., González, R., Pérez-González, J. A., González-Candelas, L., and Ramón, D. (1994). Development of a transformation system for Trichoderma longibrachiatum and its use for constructing multicopy transformants for theegl1 gene. Appl. Microbiol. Biotechnol. 41, 440–446. doi: 10.1007/bf01982533
Schardl, C. L., and Craven, K. D. (2003). Interspecific hybridization in plant-associated fungi and oomycetes: a review. Mol. Ecol. 12, 2861–2873. doi: 10.1046/j.1365-294X.2003.01965.x
Schenke, D., Böttcher, C., Lee, J., and Scheel, D. (2011). Verticillin A is likely not produced by Verticillium sp. J. Antibiot. 64, 523–524. doi: 10.1038/ja.2011.36
Schmitt, I., and Lumbsch, H. T. (2009). Ancient horizontal gene transfer from bacteria enhances biosynthetic capabilities of fungi. PLoS ONE 4:e4437. doi: 10.1371/journal.pone.0004437
Schmoll, M., and Schuster, A. (2010). Biology and biotechnology of Trichoderma. Appl. Microbiol. Biotechnol. 87, 787–799. doi: 10.1007/s00253-010-2632-1
Schoch, C. L., Sung, G. H., López-Giráldez, F., Townsend, J. P., Miadlikowska, J., Hofstetter, V., et al. (2009). The ascomycota tree of life: a phylum-wide phylogeny clarifies the origin and evolution of fundamental reproductive and ecological traits. Syst. Biol. 58, 224–239. doi: 10.1093/sysbio/syp020
Schrimpe-Rutledge, A. C., Codreanu, S. G., Sherrod, S. D., and McLean, J. A. (2016). Untargeted metabolomics strategies—challenges and emerging directions. J. Am. Soc. Mass Spectrom. 27, 1897–1905. doi: 10.1007/s13361-016-1469-y
Schroers, H. J., Samuels, G. J., Seifert, K. A., and Gams, W. (1999). Classification of the mycoparasite Gliocladium roseum in Clonostachys as C. rosea, its relationship to Bionectria ochroleuca, and notes on other Gliocladium-like fungi. Mycologia 91, 365–385. doi: 10.2307/3761383
Seidl, V., Seibel, C., Kubicek, C. P., and Schmoll, M. (2009). Sexual development in the industrial workhorse Trichoderma reesei. Proc. Natl. Acad. Sci. U.S.A. 106, 13909–13914. doi: 10.1073/pnas.0904936106
Shenouda, M. L., and Cox, R. J. (2021). Molecular methods unravel the biosynthetic potential of: Trichoderma species. RSC Adv. 11, 3622–3635. doi: 10.1039/d0ra09627j
Shi, M., Chen, L., Wang, X. W., Zhang, T., Zhao, P. B., Song, X. Y., et al. (2012). Antimicrobial peptaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 158, 166–175. doi: 10.1099/mic.0.052670-0
Shin, J., Kim, J. E., Lee, Y. W., and Son, H. (2018). Fungal cytochrome p450s and the p450 complement (Cypome) of Fusarium graminearum. Toxins 10:112. doi: 10.3390/toxins10030112
Sieber, C. M. K., Lee, W., Wong, P., Münsterkötter, M., Mewes, H. W., Schmeitzl, C., et al. (2014). The Fusarium graminearum genome reveals more secondary metabolite gene clusters and hints of horizontal gene transfer. PLoS ONE 9:e110311. doi: 10.1371/journal.pone.0110311
Singh, J., and Yadav, A. N. (2020). Natural Bioactive Products in Sustainable Agriculture. Singapore: Springer Nature.
Singh, S. B., Liu, W., Li, X., Chen, T., Shafiee, A., Card, D., et al. (2012). Antifungal spectrum, in vivo efficacy, and structure-activity relationship of ilicicolin H. ACS Med. Chem. Lett. 3, 814–817. doi: 10.1021/ml300173e
Singh, S. B., Zink, D. L., Goetz, M. A., Dombrowski, A. W., Polishook, J. D., and Hazuda, D. J. (1998). Equisetin and a novel opposite stereochemical homolog phomasetin, two fungal metabolites as inhibitors of HIV-1 integrase. Tetrahedron Lett. 39, 2243–2246. doi: 10.1016/S0040-4039(98)00269-X
Singh, N. K., Blachowicz, A., Romsdahl, J., Wang, C., Torok, T., and Venkateswaran, K. (2017). Draft genome sequences of several fungal strains selected for exposure to microgravity at the international space station. Genome Announc. 5:e01602-16. doi: 10.1128/genomeA.01602-16
Sinha, S., Nge, C. E., Leong, C. Y., Ng, V., Crasta, S., Alfatah, M., et al. (2019). Genomics-driven discovery of a biosynthetic gene cluster required for the synthesis of BII-Rafflesfungin from the fungus Phoma sp. F3723. BMC Genomics 20, 1–18. doi: 10.1186/s12864-019-5762-6
Slightom, J. L., Metzger, B. P., Luu, H. T., and Elhammer, A. P. (2009). Cloning and molecular characterization of the gene encoding the Aureobasidin A biosynthesis complex in Aureobasidium pullulans BP-1938. Gene 431, 67–79. doi: 10.1016/j.gene.2008.11.011
Slot, J. C., and Rokas, A. (2011). Horizontal transfer of a large and highly toxic secondary metabolic gene cluster between fungi. Curr. Biol. 21, 134–139. doi: 10.1016/j.cub.2010.12.020
Smedsgaard, J., and Nielsen, J. (2005). Metabolite profiling of fungi and yeast: from phenotype to metabolome by MS and informatics. J. Exp. Botany 56, 273–286. doi: 10.1093/jxb/eri068
Snarr, B. D., Baker, P., Bamford, N. C., Sato, Y., Liu, H., Lehoux, M., et al. (2017). Microbial glycoside hydrolases as antibiofilm agents with cross-kingdom activity. Proc. Natl. Acad. Sci. U.S.A. 114, 7124–7129. doi: 10.1073/pnas.1702798114
Song, Y. P., Miao, F. P., Fang, S. T., Yin, X. L., and Ji, N. Y. (2018). Halogenated and nonhalogenated metabolites from the marine-alga-endophytic fungus Trichoderma asperellum CF44-2. Mar. Drugs 16:266. doi: 10.3390/md16080266
Sood, M., Kapoor, D., Kumar, V., Sheteiwy, M. S., Ramakrishnan, M., Landi, M., et al. (2020). Trichoderma: the “secrets” of a multitalented biocontrol agent. Plants 9, 1–25. doi: 10.3390/plants9060762
Souza, A. D. L., Rodrigues-Filho, E., Souza, A. Q. L., Pereira, J. O., Calgarotto, A. K., Maso, V., et al. (2008). Koninginins, phospholipase A2 inhibitors from endophytic fungus Trichoderma koningii. Toxicon 51, 240–250. doi: 10.1016/j.toxicon.2007.09.009
Spatafora, J. W., Aime, M. C., Grigoriev, I. V., Martin, F., Stajich, J. E., and Blackwell, M. (2017). The fungal tree of life: from molecular systematics to genome-scale phylogenies. Microbiol. Spectr. 5. doi: 10.1128/microbiolspec.funk-0053-2016
Spohn, R., Daruka, L., Lázár, V., Martins, A., Vidovics, F., Grézal, G., et al. (2019). Integrated evolutionary analysis reveals antimicrobial peptides with limited resistance. Nat. Commun. 10, 1–13. doi: 10.1038/s41467-019-12364-6
Stajich, J. E. (2017). Fungal genomes and insights into the evolution of the Kingdom. Microbiol. Spectr. 5:2016. doi: 10.1128/microbiolspec.funk-0055-2016
Stark, L. A. (2010). Beneficial microorganisms: countering microbephobia. CBE Life Sci. Educ. 9, 387–389. doi: 10.1187/cbe.10-09-0119
Stasz, A. T. E., Harman, G. E., and Weeden, N. F. (1988). Protoplast preparation and fusion in two biocontrol strains of Trichoderma harzianum. Mycologia 80, 141–150. doi: 10.2307/3807788
Stasz, T. E. (1990). Genetic improvement of fungi by protoplast fusion for biological control of plant pathogens. Can. J. Plant Pathol. 12, 322–327. doi: 10.1080/07060669009501007
Stipanovic, R. D., and Howell, C. R. (1982). The structure of gliovirin, a new antibiotic from Gliocladium virens. J. Antibiot. 35, 1326–1330. doi: 10.7164/antibiotics.35.1326
Stoppacher, N., Neumann, N. K. N., Burgstaller, L., Zeilinger, S., Degenkolb, T., Brückner, H., et al. (2013). The comprehensive peptaibiotics database. Chem. Biodivers. 10, 734–743. doi: 10.1002/cbdv.201200427
Studholme, D. J., Harris, B., Le Cocq, K., Winsbury, R., Perera, V., Ryder, L., et al. (2013). Investigating the beneficial traits of Trichoderma hamatum GD12 for sustainable agriculture-insights from genomics. Front. Plant Sci. 4:258. doi: 10.3389/fpls.2013.00258
Stukenbrock, E. H. (2016). The role of hybridization in the evolution and emergence of new fungal plant pathogens. Phytopathology 106, 104–112. doi: 10.1094/PHYTO-08-15-0184-RVW
Sun, J. Z., Liu, X. Z., McKenzie, E. H. C., Jeewon, R., Liu (Jack), J. K., Zhang, X. L., et al. (2019). Fungicolous fungi: terminology, diversity, distribution, evolution, and species checklist. Fungal Divers. 95, 337–430. doi: 10.1007/s13225-019-00422-9
Suzuki, T., Watanabe, S., Kobayashi, S., and Tanino, K. (2017). Enantioselective total synthesis of (+)-Iso-A82775C, a proposed biosynthetic precursor of chloropupukeananin. Org. Lett. 19, 922–925. doi: 10.1021/acs.orglett.7b00085
Swain, H., Adak, T., Mukherjee, A. K., Mukherjee, P. K., Bhattacharyya, P., Behera, S., et al. (2018). Novel Trichoderma strains isolated from tree barks as potential biocontrol agents and biofertilizers for direct seeded rice. Microbiol. Res. 214, 83–90. doi: 10.1016/j.micres.2018.05.015
Szymańska, M., Karakulska, J., Sobolewski, P., Kowalska, U., Grygorcewicz, B., Böttcher, D., et al. (2020). Glycoside hydrolase (PelAh) immobilization prevents Pseudomonas aeruginosa biofilm formation on cellulose-based wound dressing. Carbohydr. Polym. 246:116625. doi: 10.1016/j.carbpol.2020.116625
Tan, X., Hu, Y., Jia, Y., Hou, X., Xu, Q., Han, C., et al. (2020). A conserved glycoside hydrolase family 7 cellobiohydrolase PsGH7a of Phytophthora sojae is required for full virulence on soybean. Front. Microbiol. 11:1285. doi: 10.3389/fmicb.2020.01285
Te'o, V. S. J., Bergquist, P. L., and Nevalainen, K. M. H. (2002). Biolistic transformation of Trichoderma reesei using the Bio-Rad seven barrels Hepta Adaptor system. J. Microbiol. Methods 51, 393–399. doi: 10.1016/S0167-7012(02)00126-4
Thambugala, K. M., Daranagama, D. A., Phillips, A. J. L., Kannangara, S. D., and Promputtha, I. (2020). Fungi vs. fungi in biocontrol: an overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 10:718. doi: 10.3389/fcimb.2020.604923
Tijerino, A., Elena Cardoza, R., Moraga, J., Malmierca, M. G., Vicente, F., Aleu, J., et al. (2011). Overexpression of the trichodiene synthase gene tri5 increases trichodermin production and antimicrobial activity in Trichoderma brevicompactum. Fungal Genet. Biol. 48, 285–296. doi: 10.1016/j.fgb.2010.11.012
Toyomasu, T., Tsukahara, M., Kaneko, A., Niida, R., Mitsuhashi, W., Dairi, T., et al. (2007). Fusicoccins are biosynthesized by an unusual chimera diterpene synthase in fungi. Proc. Natl. Acad. Sci. U.S.A. 104, 3084–3088. doi: 10.1073/pnas.0608426104
Tran, P. N., Yen, M. R., Chiang, C. Y., Lin, H. C., and Chen, P. Y. (2019). Detecting and prioritizing biosynthetic gene clusters for bioactive compounds in bacteria and fungi. Appl. Microbiol. Biotechnol. 103, 3277–3287. doi: 10.1007/s00253-019-09708-z
Triolet, M., Guillemin, J. P., Andre, O., and Steinberg, C. (2020). Fungal-based bioherbicides for weed control: a myth or a reality? Weed Res. 60, 60–77. doi: 10.1111/wre.12389
Urbán, P., Miao, Y., Fekete, C., Hatvani, L., Büchner, R., Vágvölgyi, C., et al. (2016). “Complete genome sequence of the green mould pathogen, Trichoderma pleuroti,” in 18th Danube-Kris-Mures-Tisa (DMKT) Euroregional Conference on Environment and Health, Book of Abstracts, ed B. Škrbić (Novi Sad: University of Novi Sad), 51–52.
van den Bosch, R., Messenger, P. S., and Gutierrez, A. P. (1982). “The history and development of biological control,” in An Introduction to Biological Control, eds A. P. Gutierrez, P. S. Messenger, and R. van den Bosch (New York, NY: Springer US), 21–36.
van Lenteren, J. C., Bolckmans, K., Köhl, J., Ravensberg, W. J., and Urbaneja, A. (2018). Biological control using invertebrates and microorganisms: plenty of new opportunities. BioControl 63, 39–59. doi: 10.1007/s10526-017-9801-4
Vargas Gil, S., Pastor, S., and March, G. J. (2009). Quantitative isolation of biocontrol agents Trichoderma spp., Gliocladium spp. and actinomycetes from soil with culture media. Microbiol. Res. 164, 196–205. doi: 10.1016/j.micres.2006.11.022
Venice, F., Davolos, D., Spina, F., Poli, A., Prigione, V. P., Varese, G. C., et al. (2020). Genome sequence of Trichoderma lixii mut3171, a promising strain for mycoremediation of pah-contaminated sites. Microorganisms 8, 1–15. doi: 10.3390/microorganisms8091258
Verma, M., Brar, S. K., Tyagi, R. D., Surampalli, R. Y., and Valéro, J. R. (2007). Antagonistic fungi, Trichoderma spp.: panoply of biological control. Biochem. Eng. J. 37, 1–20. doi: 10.1016/j.bej.2007.05.012
Vicente, I., Baroncelli, R., Morán-Diez, M. E., Bernardi, R., Puntoni, G., Hermosa, R., et al. (2020). Combined comparative genomics and gene expression analyses provide insights into the terpene synthases inventory in Trichoderma. Microorganism 8, 1–20. doi: 10.3390/microorganisms8101603
Vinale, F., Sivasithamparam, K., Ghisalberti, E. L., Marra, R., Barbetti, M. J., Li, H., et al. (2008a). A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol. Mol. Plant Pathol. 72, 80–86. doi: 10.1016/j.pmpp.2008.05.005
Vinale, F., Sivasithamparam, K., Ghisalberti, E. L., Marra, R., Woo, S. L., and Lorito, M. (2008b). Trichoderma-plant-pathogen interactions. Soil Biol. Biochem. 40, 1–10. doi: 10.1016/j.soilbio.2007.07.002
Vincelli, P. (2012). Q(o)I (strobilurin) fungicides: benefits and risks. Plant Heal. Instr. doi: 10.1094/phi-i-2002-0809-02. [Epub ahead of print].
Von Bargen, K. W., Niehaus, E. M., Krug, I., Bergander, K., Würthwein, E. U., Tudzynski, B., et al. (2015). Isolation and structure elucidation of Fujikurins A-D: products of the PKS19 Gene Cluster in Fusarium fujikuroi. J. Nat. Prod. 78, 1809–1815. doi: 10.1021/np5008137
Wada, S., ichi Iida, A., Asami, K., and Fujita, T. (1996). Ion channel-forming property of trichorovin-XII, an 11-residue peptaibol from the fungus Trichoderma viride, in planar lipid bilayer membranes. Bioorganic Med. Chem. Lett. 6, 2275–2278. doi: 10.1016/0960-894X(96)00410-6
Wang, C. C. C., Chiang, Y. M., Praseuth, M. B., Kuo, P. L., Liang, H. L., and Hsu, Y. L. (2010). Asperfuranone from Aspergillus nidulans inhibits proliferation of human non-small cell lung cancer A549 cells via blocking cell cycle progression and inducing apoptosis. Basic Clin. Pharmacol. Toxicol. 107, 583–589. doi: 10.1111/j.1742-7843.2010.00545.x
Wang, G., Li, X., and Wang, Z. (2016). APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 44, D1087–D1093. doi: 10.1093/nar/gkv1278
Wang, X., Peng, J., Sun, L., Bonito, G., Guo, Y., Li, Y., et al. (2020). Genome sequencing of Paecilomyces penicillatus provides insights into its phylogenetic placement and mycoparasitism mechanisms on morel mushrooms. Pathogens 9, 1–12. doi: 10.3390/pathogens9100834
Wanka, F. (2021). “Open the pores: electroporation for the transformation of Trichoderma reesei,” in Methods in Molecular Biology, eds A. R. Mach-Aigner and R. Martzy (New York, NY: Humana Press Inc.), 73–78.
Wasil, Z., Pahirulzaman, K. A. K., Butts, C., Simpson, T. J., Lazarus, C. M., and Cox, R. J. (2013). One pathway, many compounds: heterologous expression of a fungal biosynthetic pathway reveals its intrinsic potential for diversity. Chem. Sci. 4, 3845–3856. doi: 10.1039/c3sc51785c
Wee, J. L., Sundermann, K., Licari, P., and Galazzo, J. (2006). Cytotoxic hypothemycin analogues from Hypomyces subiculosus. J. Nat. Prod. 69, 1456–1459. doi: 10.1021/np060258o
Whipps, J. M. (2001). Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 52, 487–511. doi: 10.1093/jexbot/52.suppl_1.487
Woo, P. C. Y., Lam, C. W., Tam, E. W. T., Lee, K. C., Yung, K. K. Y., Leung, C. K. F., et al. (2014). The biosynthetic pathway for a thousand-year-old natural food colorant and citrinin in Penicillium marneffei. Sci. Rep. 4, 1–8. doi: 10.1038/srep06728
Xiao-Yan, S., Qing-Tao, S., Shu-Tao, X., Xiu-Lan, C., Cai-Yun, S., and Yu-Zhong, Z. (2006). Broad-spectrum antimicrobial activity and high stability of Trichokonins from Trichoderma koningii SMF2 against plant pathogens. FEMS Microbiol. Lett. 260, 119–125. doi: 10.1111/j.1574-6968.2006.00316.x
Xie, B. B., Qin, Q. L., Shi, M., Chen, L. L., Shu, Y. L., Luo, Y., et al. (2014). Comparative genomics provide insights into evolution of Trichoderma nutrition style. Genome Biol. Evol. 6, 379–390. doi: 10.1093/gbe/evu018
Xu, Y., Vinas, M., Alsarrag, A., Su, L., Pfohl, K., Rohlfs, M., et al. (2019). Bis-naphthopyrone pigments protect filamentous ascomycetes from a wide range of predators. Nat. Commun. 10, 1–12. doi: 10.1038/s41467-019-11377-5
Xu, Z., Escamilla-Treviño, L. L., Zeng, L., Lalgondar, M., Bevan, D. R., Winkel, B. S. J., et al. (2004). Functional genomic analysis of Arabidopsis thaliana glycoside hydrolase family 1. Plant Mol. Biol. 55, 343–367. doi: 10.1007/s11103-004-0790-1
Xu, J., Li, F., Leier, A., Xiang, D., Shen, H.-H., Marquez Lago, T. T., et al. (2021). Comprehensive assessment of machine learning-based methods for predicting antimicrobial peptides. Brief. Bioinform. doi: 10.1093/bib/bbab083. [Epub ahead of print].
Yabuta, T., Sumiki, Y., Aso, K., Tamura, T., Igarashi, H., and Tamari, K. (1939). Biochemical studies on the bakanae fungus. IV. The culture conditions for producing gibberellin or fusaric acid. J. Agric. Chem. Soc. Jpn. 15, 1209–1220.
Yamashita, A., Yoshizawa, T., Aiura, Y., Sanchez, P. C., Dizon, E. I., Arim, R. H., et al. (1995). Fusarium mycotoxins (Fumonisins, Nivalenol, and Zearalenone) and aflatoxins in corn from Southeast Asia. Biosci. Biotechnol. Biochem. 59, 1804–1807. doi: 10.1271/bbb.59.1804
Yan, Y., Liu, Q., Jacobsen, S. E., and Tang, Y. (2018). The impact and prospect of natural product discovery in agriculture. EMBO Rep. 19:e46824. doi: 10.15252/embr.201846824
Yang, D., Pomraning, K., Kopchinskiy, A., Aghcheh, R. K., Atanasova, L., Chenthamara, K., et al. (2015). Genome sequence and annotation of Trichoderma parareesei, the ancestor of the cellulase producer Trichoderma reesei. Genome Announc. 3, 885–900. doi: 10.1128/genomeA.00885-15
Yang, L., Yang, Q., Sun, K., Tian, Y., and Li, H. (2011). Agrobacterium tumefaciens mediated transformation of ChiV gene to Trichoderma harzianum. Appl. Biochem. Biotechnol. 163, 937–945. doi: 10.1007/s12010-010-9097-7
Zeilinger, S. (2004). Gene disruption in Trichoderma atroviride via Agrobacterium-mediated transformation. Curr. Genet. 45, 54–60. doi: 10.1007/s00294-003-0454-8
Zeilinger, S., Gruber, S., Bansal, R., and Mukherjee, P. K. (2016). Secondary metabolism in Trichoderma- chemistry meets genomics. Fungal Biol. Rev. 30, 74–90. doi: 10.1016/j.fbr.2016.05.001
Zhang, L., Yan, J., Fu, Z., Shi, W., Ninkuu, V., Li, G., et al. (2021). FoEG1, a secreted glycoside hydrolase family 12 protein from Fusarium oxysporum, triggers cell death and modulates plant immunity. Mol. Plant Pathol. 22, 522–538. doi: 10.1111/mpp.13041
Zhang, Q. W., Lin, L. G., and Ye, W. C. (2018a). Techniques for extraction and isolation of natural products: a comprehensive review. Chinese Med. 13:20. doi: 10.1186/s13020-018-0177-x
Zhang, S., Gan, Y., Ji, W., Xu, B., Hou, B., and Liu, J. (2017). Mechanisms and characterization of Trichoderma longibrachiatum T6 in suppressing nematodes (Heterodera avenae) in wheat. Front. Plant Sci. 8:1491. doi: 10.3389/fpls.2017.01491
Zhang, S., Gan, Y., and Xu, B. (2015). Biocontrol potential of a native species of Trichoderma longibrachiatum against Meloidogyne incognita. Appl. Soil Ecol. 94, 21–29. doi: 10.1016/j.apsoil.2015.04.010
Zhang, S., Xu, B., Zhang, J., and Gan, Y. (2018b). Identification of the antifungal activity of Trichoderma longibrachiatum T6 and assessment of bioactive substances in controlling phytopathgens. Pestic. Biochem. Physiol. 147, 59–66. doi: 10.1016/j.pestbp.2018.02.006
Zhang, Y., Wang, X., Pang, G., Cai, F., Zhang, J., Shen, Z., et al. (2019). Two-step genomic sequence comparison strategy to design Trichoderma strain-specific primers for quantitative PCR. AMB Express 9:179. doi: 10.1186/s13568-019-0904-4
Zhang, J., Xin, L., Shan, B., Chen, W., Xie, M., Yuen, D., et al. (2012). PEAKS DB: de novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteomics 11:M111.010587. doi: 10.1074/mcp.M111.010587
Zhao, Y., Ding, J., Yuan, W., Huang, J., Huang, W., Wang, Y., et al. (2017). Production of a fungal furocoumarin by a polyketide synthase gene cluster confers the chemo-resistance of Neurospora crassa to the predation by fungivorous arthropods. Environ. Microbiol. 19, 3920–3929. doi: 10.1111/1462-2920.13791
Zheng, P., Xia, Y., Xiao, G., Xiong, C., Hu, X., Zhang, S., et al. (2011). Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional chinese medicine. Genome Biol. 12:R116. doi: 10.1186/gb-2011-12-11-r116
Zhong, Y. H., Wang, X. L., Wang, T. H., and Jiang, Q. (2007). Agrobacterium-mediated transformation (AMT) of Trichoderma reesei as an efficient tool for random insertional mutagenesis. Appl. Microbiol. Biotechnol. 73, 1348–1354. doi: 10.1007/s00253-006-0603-3
Zhou, Y., Wang, Y., Chen, K., Wu, Y., Hu, J., Wei, Y., et al. (2020). Near-complete genomes of two Trichoderma species: a resource for biological control of plant pathogens. Mol. Plant Microbe Interact. 33, 1036–1039. doi: 10.1094/MPMI-03-20-0076-A
Keywords: predictive biology, functional genomics, secondary metabolites, antimicrobials, integrated pest management, plant-microbe interactions, graph theory—graph algorithms, drug discoveries
Citation: Rush TA, Shrestha HK, Gopalakrishnan Meena M, Spangler MK, Ellis JC, Labbé JL and Abraham PE (2021) Bioprospecting Trichoderma: A Systematic Roadmap to Screen Genomes and Natural Products for Biocontrol Applications. Front. Fungal Biol. 2:716511. doi: 10.3389/ffunb.2021.716511
Received: 28 May 2021; Accepted: 10 August 2021;
Published: 16 September 2021.
Edited by:
Artemio Mendoza-Mendoza, Lincoln University, New ZealandReviewed by:
Birinchi Kumar Sarma, Banaras Hindu University, IndiaRosa Hermosa, University of Salamanca, Spain
Copyright © 2021 Rush, Shrestha, Gopalakrishnan Meena, Spangler, Ellis, Labbé and Abraham. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jesse L. Labbé, labbejj@ornl.gov; Paul E. Abraham, abrahampe@ornl.gov