 Páraic Ó Cuív
Páraic Ó Cuív Rabina Giri
Rabina Giri Emily C. Hoedt
Emily C. Hoedt Michael A. McGuckin2†
Michael A. McGuckin2† Jakob Begun
Jakob Begun Mark Morrison
Mark Morrison- 1The University of Queensland Diamantina Institute, Translational Research Institute, The University of Queensland, Brisbane, QLD, Australia
- 2Mater Research Institute – The University of Queensland, Translational Research Institute, Brisbane, QLD, Australia
Enterococcus faecalis is an early coloniser of the human infant gut and contributes to the development of intestinal immunity. To better understand the functional capacity of E. faecalis, we constructed a broad host range RP4 mobilizable vector, pEHR513112, that confers chloramphenicol resistance and used a metaparental mating approach to isolate E. faecalis AHG0090 from a fecal sample collected from a healthy human infant. We demonstrated that E. faecalis AHG0090 is genetically tractable and could be manipulated using traditional molecular microbiology approaches. E. faecalis AHG0090 was comparable to the gold-standard anti-inflammatory bacterium Faecalibacterium prausnitzii A2-165 in its ability to suppress cytokine-mediated nuclear factor kappa B (NF-κB) activation in human gut-derived LS174T goblet cell like and Caco-2 enterocyte-like cell lines. E. faecalis AHG0090 and F. prausnitzii A2-165 produced secreted low molecular weight NF-κB suppressive peptidic bioactives. Both bioactives were sensitive to heat and proteinase K treatments although the E. faecalis AHG0090 bioactive was more resilient to both forms of treatment. As expected, E. faecalis AHG0090 suppressed IL-1β-induced NF-κB-p65 subunit nuclear translocation and expression of the NF-κB regulated genes IL-6, IL-8 and CXCL-10. Finally, we determined that E. faecalis AHG0090 is distantly related to other commensal strains and likely encodes niche factors that support effective colonization of the infant gut.
Introduction
The human gut represents the largest mucosal surface area and is the largest immune organ of the body (1). Full-term infants are born with a competent but immature immune system that must respond appropriately to the inevitable exposure to microbes that occurs following birth. The infant microbiota is derived principally from the maternal microbiota (2) and the early colonizers of the gut play a critical role in priming mucosal immunity and establishing a homeostatic relationship with the host (3, 4).
Enterococcus faecalis is one of the most abundant colonizers of the infant gastrointestinal tract (5, 6) and together with other enterococci, staphylococci, and enterobacteria helps reduce the gut environment to facilitate subsequent colonization by obligate anaerobes (7, 8). Select E. faecalis infant derived strains also possess immunomodulatory capacities (9) and exert anti-inflammatory activities by modulating the nuclear factor kappa B (NF-κB), mitogen-activated protein kinase and peroxisome proliferator-activated receptor-γ1 regulated pathways (10, 11). Some of the immunomodulatory factors produced by E. faecalis have been identified (12–14); however, the extent of immunomodulatory capacities among non-pathogenic gut strains, and the identity of the bioactives that underpin them, remains largely unknown.
Taken together, we hypothesized the infant gut microbiota would be a fertile source of immunomodulatory bioactive factors with potential prophylactic or therapeutic applications. We previously reported a method termed metaparental mating (MPM) that enables the rapid and directed isolation of genetically tractable human gut bacteria (15). In this study, we describe the isolation of E. faecalis AHG0090 and demonstrate that similar to Faecalibacterium prausnitzii A2-165 (16, 17), it produces a potent peptidic bioactive that suppresses NF-κB activation. Finally, we demonstrate that E. faecalis AHG0090 can be manipulated using traditional molecular techniques providing new opportunities to dissect the functional capacity of the human gut microbiota.
Materials and Methods
Growth and Culture Conditions
The recipient cultures for MPM were prepared by inoculating Brain Heart Infusion (BHI, Difco™) supplemented with colistin sulfate with a raw stool sample collected from a healthy 2-year-old female child. The donor had not taken antibiotics during the 3-month period prior to collection. The child was recruited as part of a study into the link between the gut microbiota and type 1 diabetes susceptibility. All study samples were collected in accordance with the recommendations of the Mater Health Services Human Research Ethics Committee (HREC/13/MHS/21/AM02). All subjects gave written informed consent in accordance with the Declaration of Helsinki, with written consent provided from parents or legal guardians for all subjects <13 years. The protocol was approved by the Mater Health Services HREC. E. faecalis was cultured in BHI and the Escherichia coli ST18 donor strain for MPM was cultured in BHI supplemented with δ-aminolevulinic acid (100 µg ml−1). The E. coli cloning strains were grown in LB and F. prausnitzii A2-165 was cultured in anaerobic Reinforced Clostridial Medium (RCM, Oxoid™) buffered with salt solutions 2 and 3 (18). F. prausnitzii cultures were routinely manipulated in a Coy vinyl anaerobic chamber with an oxygen free atmosphere (85% N2:10% CO2:5% H2). Both E. coli ST18 and JM109 competent cells were prepared by the rubidium chloride method (19) while Invitrogen™ E. coli DH5 α competent cells were purchased from ThermoFisher Scientific. The bacterial growth media were supplemented with erythromycin (100 µg ml−1), chloramphenicol (20 µg ml−1) or colisitin sulfate (20 µg ml−1) as appropriate.
Bacterial growth was measured by the increase in optical density at 600 nm (OD600 nm). Specific growth rates [μ (h−1)] were calculated by log10 transformation of the OD600 nm measurements and plotting a trendline (R2 > 0.97) for the linear phase of growth corresponding to exponential (EX) growth phase. Then μ was calculated using the equation: μ = (slope of the line × 2.3).
Vector Construction and Manipulation
To construct pEHR513111, catP was PCR amplified from pJIR1456 (20) with primers (Pf 5′ GAT CGT TTA AAC AGT GGG CAA GTT GAA AAA TTC AC; Pr 5′ GAT CCC TGC AGG TTA GGG TAA CAA AAA ACA CCG TAT TTC TAC) that introduced unique PmeI and SbfI restriction sites. The digested catP was then cloned into pEHR512111, replacing erm, and generating pEHR513111. The pEHR513112 vector was subsequently constructed by cloning cphy_3290-evolglow-C-Bs2 from pEHR512112 (15) into the multiple cloning site of pEHR513111 as an EcoRI-HindIII fragment. The pEHR513111 and pEHR513112 vectors were confirmed by restriction digest analysis and the sequences compiled using publicly available sequences. When appropriate, the pEHR513112 vector was cured from E. faecalis AHG0090 by overnight growth in BHI medium supplemented with acridine orange (1–8 µg ml−1) and then plating on BHI medium. Single colonies were replica plated onto BHI medium with or without chloramphenicol to identify naïve E. faecalis AHG0090 isolates.
Plasmid DNA was extracted from E. faecalis cultures using a modified alkaline lysis method (21). Briefly, 1 ml of E. faecalis culture was centrifuged and the cells were resuspended in 200 µl of solution 1. The cell suspension was supplemented with 1 µl of mutanolysin (20 U/μl) and 10 µl lysozyme (200 mg ml−1) and incubated at 37°C for up to 1 h. Next, 200 µl of solution 2 was added, and the mixture was incubated on ice for 5 min. Then, 200 µl of solution 3 was added, and the mixture was incubated on ice for 10 min. The cell debris were removed by centrifugation and the clarified cell lysate was recovered and extracted with phenol:chloroform:isoamyalcohol (25:24:1). The aqueous phase was transferred to a fresh tube and plasmid DNA was precipitated using isopropanol, washed with 70% (v/v) ethanol and then resuspended in TE buffer. Plasmid DNA was similarly prepared from E. coli except that the treatments with mutanolysin and lysozyme, and the incubations on ice were not performed.
Mating Procedures
The E. coli ST18 donor strain for metaparental and biparental matings was grown in BHI medium supplemented with δ-aminolevulinic acid and chloramphenicol. The MPM experiments were performed essentially as previously described (15) with two exceptions. First, the mating mix and controls were spotted directly onto the surface of BHI agar rather than plating on a nylon filter. Second, 100 µl of mating mix and controls were transferred to fresh BHI broth and grown for 5 h before selecting for transconjugants. Biparental matings were similarly performed except that the 5 h outgrowth was not done. Transconjugants were recovered on BHI medium supplemented with chloramphenicol and colisin sulfate.
Microscopy
Naïve and transconjugants strains of E. faecalis AHG0090 were examined with an Olympus BX 63 microscope fitted with an Xcite LED light source and fluorescence filter cube U-FBN (excitation 470–495 nm, emission 510 nm). Images were captured using a DP80 camera and the Olympus cellSens modular imaging software platform and ImageJ software package1 were used for visualization and processing.
E. faecalis rrs Sequencing
The rrs gene was PCR amplified using the primers 27F and 1492R (22) as previously described (23), and sequenced at the Australian Genomic Research Facility (Brisbane, Australia) using primers 530F and 907R (22). The individual sequence reads were trimmed to remove low quality bases and assembled using deFUME (24). The assembled sequence was then aligned against the Ribosomal Database Project (25) core set of aligned rrs sequences and E. faecalis AHG0090 was identified using the Classifier and SeqMatch functions.
Genome Sequencing and Analysis
High molecular weight DNA was prepared from the E. faecalis AHG0090 MPM isolate as previously described (23). The DNA was then quantified using the QuantiFluor ONE dsDNA system according to manufacturer’s instructions (Promega, Australia) and the integrity of the DNA was determined by agarose gel electrophoresis. The genome was shotgun sequenced using the Illumina NextSeq 500 system (2 × 150bp High Output kit) with v2 chemistry. The sequence data were quality checked, filtered, and then de novo assembled using the SPAdes assembler v 3.10.1 (26). Genome sequencing quality was evaluated with CheckM, which estimates the input files for completeness and contamination based on the phylogenetic assignment of a broad set of marker genes (27). The E. faecalis AHG0090 contigs were ordered using Mauve (28) with the closed E. faecalis V583 genome sequence used as a reference. The Mauve generated assembly was submitted to the RAST annotation pipeline and the results were examined in the SEED Viewer (29). Genome based phylogeny was obtained using GTDB,2 built from the concatenation of 120 universal bacterial-specific marker genes (30). Tree inference was performed with FastTree v2.1.7 (31) and included all genomes in IMG v4.510 (32). The resulting tree was imaged using ARB v6.0.6 (33). To identify candidate plasmids, the fastq files were mapped to the E. faecalis AHG0090 genome assembly using BamM v1.7.33 to determine the coverage profiles for each contig. The average coverage was then calculated and contigs with >1,000× coverage were identified as candidate plasmids. In addition, we used PlasmidSPAdes to assemble plasmids from whole genome sequencing data (34). The candidate plasmids were manually curated to determine if they could be closed and compared to other plasmids using Blastn. This Whole Genome Shotgun project was deposited at DDBJ/EMBL/GenBank under the accession PDUN00000000. The version described in this paper is the first version, PDUN01000000.
Measurement of E. faecalis Immunomodulatory Activities
The immunomodulatory potential of E. faecalis was assessed using LS174T-NF-κBluc goblet cell like and Caco-2-NF-κBluc enterocyte-like reporter cell lines (35). Briefly, three individual E. faecalis AHG0090 colonies were established as independent cultures with BHI broth. Following overnight growth, each individual culture was used to initiate duplicate 50 ml BHI broth cultures at a starting OD600 nm of 0.01 (n = 6 cultures, consisting of n = 3 independent biological replicates with n = 2 technical replicates each). The OD600 nm of the cultures was monitored longitudinally and 5 ml of cultures was harvested from each broth culture at early EX, mid-exponential (MX), early stationary (ES), and late stationary (LS) phase of growth. At each collection, 1.5 ml of each culture was centrifuged at 16,000 × g for 5 min and 0.5 ml of the cell-free supernatant fraction was transferred to fresh tubes and stored at −30°C as single-use aliquots.
For the immunomodulatory assays, 96-well microtiter plates were seeded with 20,000 LS174T-NF-kBluc or Caco-2-NF-kBluc reporter cells per well as previously described (35). The ability of the cell-free bacterial supernatants to suppress NF-κB activation in LS174T-NF-kBluc was assessed by adding supernatant (10% v/v in complete DMEM medium) to the cells and incubating for 30 min at 37°C. The LS174T and Caco-2 reporter cell lines were then stimulated with TNFα (25 ng ml−1) or IL-1β (50 ng ml−1), respectively, in the presence of 10% v/v supernatant for 4 h before assessing luciferase activity. The ability of the supernatants to suppress activation was compared to the NF-κB inhibitor indole-3-carbinol (I3C, 5 µM). NF-κB driven luciferase expression was assessed using the Pierce™ Firefly Luc One-Step Glow Assay Kit (ThermoFisher Scientific) according to the manufacturer’s instructions. The cytotoxicity of the supernatants was assessed using Cell Proliferation Reagent WST-1 (Sigma Aldrich) according to the manufacturer’s instructions.
Nuclear Translocation Immunofluorescence Assays
Glass coverslips in a 12 well-plate were seeded with 20,000 Caco-2 cells per well and cultured overnight. Cell-free culture supernatants harvested from MX phase cultures were added (10% v/v) to the Caco-2 cells and incubated for 30 min and then stimulated with IL-1β (25 ng ml−1) for 1 h. Cells were also treated with BHI medium and I3C alone. The cells were then processed and analyzed as previously described (35).
Quantitative Reverse Transcriptase PCR (qRT-PCR) Assays
A 12-well plate was seeded with 50,000 Caco-2 cells per well and cultured overnight. Cell-free culture supernatants harvested from MX phase cultures were added (10% v/v) to the Caco-2 cells and incubated for 30 min and then stimulated with IL-1β (25 ng ml−1) for 6 h. Cells were also treated with I3C and BHI medium alone. Total RNA was isolated and the expression of the NF-κB-dependent genes IL-6, IL-8 and CXCL10 was assessed as previously described (35), except that different primers were used for IL-6 (Pf 5′ CCA CTC ACC TCT TCA GAA CG; Pr 5′ CAT CTT TGG AAG GTT CAG GTT G) (36).
Results
Isolation of E. faecalis AHG0090
To recover genetically tractable facultative anaerobic Firmicutes bacteria from the healthy infant human gut, we produced a microbial enrichment culture from human stool and determined that chloramphenicol completely inhibited growth of candidate recipients on BHI medium under aerobic conditions. Consequently, we constructed a vector, pEHR513111, carrying catP, and this vector was further modified by cloning the evoglow-C-Bs2 under the control of the Clostridium phyofermentans ISDg cphy_3290 promoter generating pEHR513112 (Figure 1A).
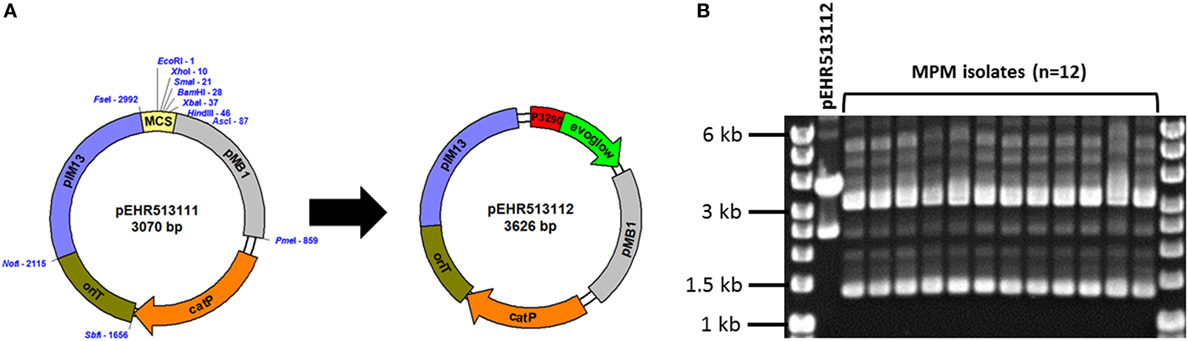
Figure 1. (A) The pEHR513111 plasmid carrying catP confers chloramphenicol resistance on its host. The pEHR513112 plasmid carries evoglow-C-Bs2 under the control of the Clostridium phyofermentans ISDg cphy_3290 promoter. (B) Plasmid profiling of the transconjugants recovered by metaparental mating (MPM isolates). The pEHR513112 plasmid carried by the transconjugants was identified by comparison with plasmid DNA prepared from Escherichia coli (pEHR513112).
Following MPM, we recovered 14 transconjugants and assessed the clonality of 12 isolates by plasmid profiling. All 12 isolates carried a plasmid of comparable molecular weight to pEHR513112 in addition to at least two other plasmids (~1.4 kb closed covalent circular form, ~3.2 kb closed covalent circular form) (Figure 1B). However, the plasmid DNA profiles of all 12 isolates were virtually identical suggesting they were clonal. Based on these observations, we chose one transconjugant for further analysis and produced 1,473 bp of 16S rRNA sequence. Based on this sequence, we determined the isolate was affiliated with the E. faecalis taxon, and hereafter it is referred to as E. faecalis AHG0090.
E. faecalis AHG0090 is Genetically Tractable
We examined whether E. faecalis AHG0090 can be genetically manipulated using traditional techniques in molecular microbiology. E. faecalis AHG0090 was grown in BHI broth supplemented with acridine orange to cure pEHR513112. The addition of acridine orange up to 8 µg ml−1 did not affect growth; however, all the colonies recovered on BHI medium were sensitive to chloramphenicol suggesting they had lost pEHR513112. Plasmid DNA prepared from E. faecalis AHG0090 recovered by MPM carried a plasmid with the same molecular weight as pEHR513112, however, this plasmid was absent from naïve E. faecalis AHG0090 (Figure 2A). We next confirmed that naïve E. faecalis AHG0090 was genetically tractable by using it as the recipient in a biparental mating with E. coli ST18 carrying pEHR513112. Using our standard biparental mating protocol, we recovered transconjugants and achieved a conjugation efficiency of 3.83 × 10−7 transconjugants per recipient. As expected, the biparental mating-derived transconjugant carried the pEHR513112 plasmid band (Figure 2A) and plasmid recovery experiments from the MPM and re-transformed isolates confirmed the plasmids were stably maintained (Figure 2B). Consistent with these observations the re-transformed but not naïve strain was fluorescent (Figure 2C). Notably, the endogenous plasmids were unaffected by the acridine orange treatment suggesting that they are stably maintained.
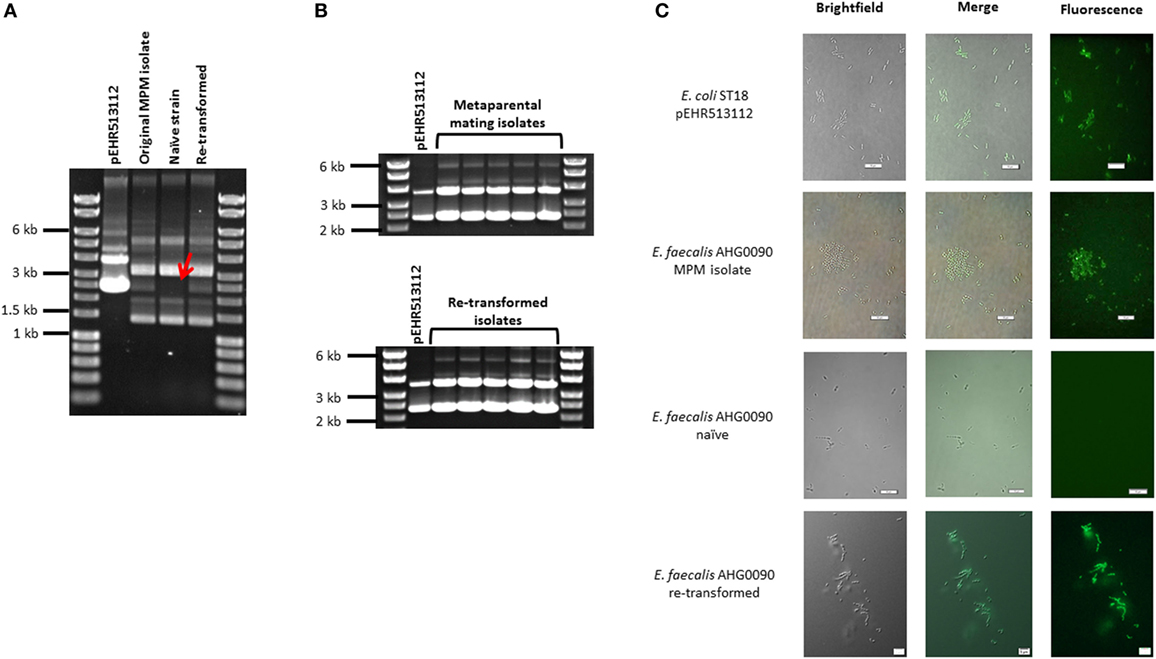
Figure 2. (A) Enterococcus faecalis AHG0090 is genetically tractable and can be manipulated using standard molecular microbiology approaches. The pEHR513112 plasmid was readily identifiable in E. faecalis AHG0090 recovered by metaparental mating (original MPM isolate) and following the re-introduction of the plasmid by biparental mating (re-transformed), but was lost following treatment with acridine orange (naïve strain). The absence of pEHR513112 in the naïve strain is indicated by a red arrow. (B) The pEHR513112 plasmid is stably maintained in E. faecalis AHG0090. pEHR513112 plasmids recovered from E. faecalis AHG0090 transconjugants produced by MPM and following re-transformation of the naïve strain were examined by agarose gel electrophoresis to identify any major structural deletions and re-arrangements. (C) E. faecalis strains carrying pEHR513112 are fluorescent. Escherichia coli ST18 carrying pEHR513112 and naïve and transconjugant E. faecalis AHG0090 strains were analyzed using brightfield and fluorescence microscopy. The scale bars represent 10 µm.
E. faecalis AHG0090 Produces an NF-κB Suppressive Peptidic Bioactive
Given the immunomodulatory activity previously ascribed to E. faecalis isolates, we examined the ability of E. faecalis AHG0090 to suppress cytokine-mediated epithelial NF-κB activation using our LS174T and Caco-2 reporter cell lines. E. faecalis AHG0090 was grown in BHI medium and achieved a specific growth rate of 1.63 ± 0.14 h−1 (growth rate ± SD) during EX growth phase and a maximum recorded yield of 3.74 ± 0.07 (OD600 ± SD) following 8 h of growth (Figure 3A). Cell-free supernatants were harvested from early EX, MX, ES and LS phase cultures as the closely related bacterium Lactobacillus plantarum produces immunomodulins that inhibit IFNγ production in a growth phase dependent manner (37). Culture supernatant harvested from all four-time points suppressed NF-κB activation in both cell lines although the extent of suppression was greatest with supernatants harvested from MX phase onward (Figure 3B).
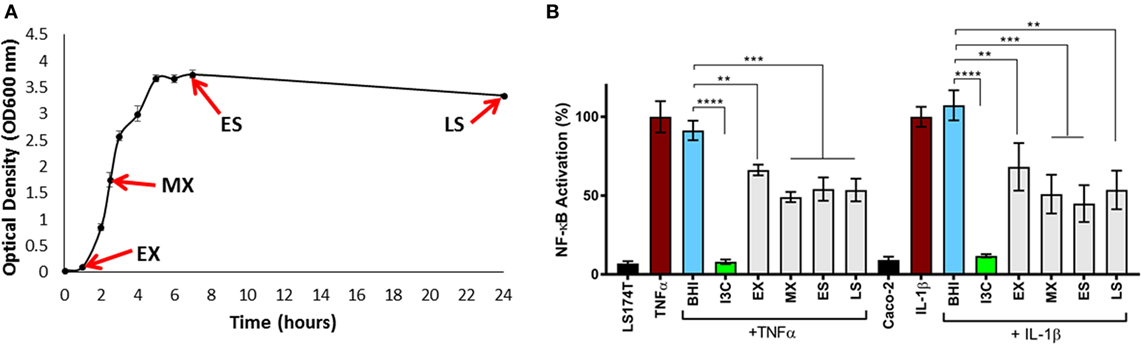
Figure 3. (A) Harvesting of Enterococcus faecalis AHG0090 cell-free culture supernatants. E. faecalis AHG0090 was cultured in Brain Heart Infusion (BHI) medium and culture supernatants were harvested in early exponential (EX), mid-exponential (MX), early stationary (ES) and late stationary (LS) phase. Growth was recorded as the change in optical density over time (hours). (B) Characterization of the nuclear factor kappa B (NF-κB) suppressive capacity of E. faecalis AHG0090. The effects of the E. faecalis AHG0090 culture supernatants on NF-κB activation in the LS174T-NF-κBluc and Caco-2-NF-κBluc reporter cell lines were measured by the luciferase assay. The extent of NF-κB activation was assessed after 6 h stimulation with TNFα/IL-1β and baseline suppression of the reporter gene was assessed using sterile BHI medium (**p < 0.01, ***p < 0.001 as determined by one-way ANOVA with Dunnett’s multiple comparison test).
We compared the NF-κB suppressive capacity of E. faecalis AHG0090 to the model anti-inflammatory gut bacterium F. prausnitzii A2-165 and determined that both strains suppressed NF-κB activation to a similar extent in the LS174T and Caco-2 reporter cell lines (Figure 4A). Critically, the E. faecalis AHG0090 and F. prausnitzii A2-165 cell-free supernatants did not exert cytotoxic effects. The E. faecalis AHG0090 culture supernatant was fractionated by passing it through a 3 kDa molecular weight cut-off filter and the NF-κB suppressive activity of the flow-through but not the retentate was similar to that of the unfractionated culture supernatant (Figure 4A). We next assessed the impact of heat and proteinase K treatments on the F. prausnitzii A2-165 and E. faecalis AHG0090 NF-κB suppressive bioactives. The activity of the F. prausnitzii A2-165 <3 kDa culture supernatant fraction was not significantly different to the Reinforced Clostridial Medium control following heat (57 and 97°C) or proteinase K treatment (Figure 4B), consistent with the NF-κB suppressive capacity of this bacterium being mediated by Mam-derived peptides (16). The E. faecalis AHG0090 <3 kDa culture supernatant fraction displayed similar characteristics but still retained activity following treatment at 57°C for 30 min or proteinase K digestion for 1 h when compared to the BHI control (Figure 4B, p < 0.05). This activity was lost at higher temperatures or following longer heat treatment and following extended proteinase K treatment (Figure 4B). Taken together, these data suggest E. faecalis AHG0090 secretes a low molecular weight NF-κB suppressive peptidic bioactive with differing properties to the F. prausnitzii Mam peptides.
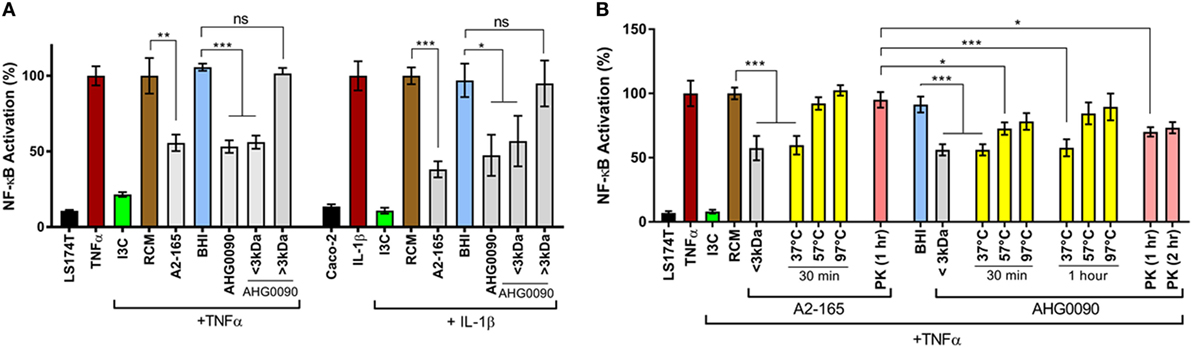
Figure 4. (A) Faecalibacterium prausnitzii A2-165 and Enterococcus faecalis AHG0090 produce low molecular weight nuclear factors kappa B (NF-κB) suppressive bioactives. The extent of NF-κB activation was assessed after 6 h stimulation of the LS174T-NF-kBluc and Caco-2-NF-kBluc reporter cell lines with TNFα and IL-1β, respectively. Baseline suppression of the reporter gene was assessed using sterile Reinforced Clostridial Medium (RCM) or Brain Heart Infusion (BHI) medium. The suppressive effects of the supernatants were assessed against the appropriate medium control. (B) F. prausnitzii A2-165 and E. faecalis AHG0090 produce heat and proteinase K labile bioactives. The effect of the heat and proteinase K treatments was assessed using the LS174T-NF-κBluc reporter cell line. The suppressive effects of the supernatants were assessed against the appropriate medium control and significant differences are indicated. ns, not significant, *p < 0.05, **p < 0.01, ***p < 0.001 as determined by one-way ANOVA with Dunnett’s multiple comparison test.
Enterococcus faecalis AHG0090 Inhibits NF-κB-p65 Subunit Nuclear Translocation and Cytokine Expression
Cytokine mediate activation of the NF-κB pathway results in nuclear translocation of the NF-κB-p65 subunit. We examined the ability of culture supernatant harvested from MX phase cultures of E. faecalis AHG0090 to suppress NF-κB-p65 subunit nuclear translocation. NF-κB-p65 subunit nuclear translocation induced by IL-1β in Caco-2 cells was unaffected by treatment with BHI medium. In contrast, E. faecalis AHG0090 MX phase culture supernatant treatment significantly reduced nuclear translocation in a similar fashion to the pharmacological inhibitor I3C (Figure 5A). As expected, I3C and E. faecalis AHG0090 culture supernatant suppressed expression of the NF-κB-dependent genes IL-6 (p < 0.05), IL-8 (p < 0.01) and CXCL10 (p < 0.001), as determined by qRT-PCR (Figure 5B).
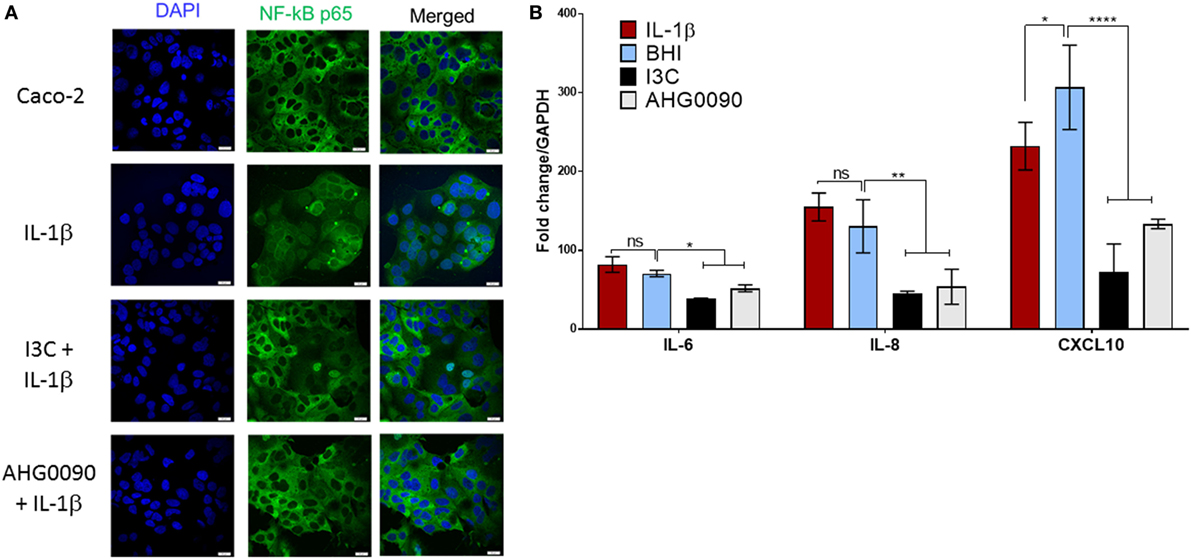
Figure 5. (A) Enterococcus faecalis AHG0090 mid-exponential (MX) phase culture supernatant suppresses nuclear factor kappa B (NF-κB)-p65 subunit nuclear translocation. The cell nuclei and NF-κB-p65 subunit are shown in blue and green, respectively. The nuclei for the Brain Heart Infusion (BHI) treated Caco-2 cells in the central panel are largely black revealing that treatment does not result in nuclear translocation. As expected, BHI treatment does not prevent NF-κB-p65 nuclear translocation following IL-1β treatment as indicated by green staining of the nuclei. By contrast, treatment with I3C and E. faecalis AHG0090 MX phase culture supernatant suppressed IL-1β-induced NF-κB-p65 nuclear translocation. The scale bars represent 10 µm. (B) E. faecalis AHG0090 MX phase culture supernatants suppress expression of NF-κB-p65-dependent cytokines. The expression of IL-6, IL-8 and CXCL10 was assessed by quantitative reverse transcriptase PCR. The data are normalized to GAPDH gene expression and presented as the fold-change relative to unstimulated cells. E. faecalis AHG0090 MX phase culture supernatants suppress expression of IL-6 (*p < 0.05), IL-8 (**p < 0.01) and CXCL10 (****p < 0.0001) as determined using one-way ANOVA with Dunnett’s multiple comparison test.
E. faecalis AHG0090 Is Adapted for Gut Colonization
We sequenced the E. faecalis AHG0090 genome to provide insights into the factors supporting colonization and persistence in the infant gut. We produced 2,925,542 bp of DNA sequence at 107× coverage. The sequenced data were assembled into 116 contigs providing a contig N50 of 144,336 bp and L50 of 8. Critically, the genome was assessed by CheckM as being essentially complete (99.63%) and free from contamination. The genome has a G+C content of 37.3% and is predicted to contain 2,929 protein-coding genes and 60 structural RNAs. Analysis of E. faecalis phylogeny using the GTDB revealed that E. faecalis AHG0090 clusters closely with three strains termed E. faecalis TX0630, TX0635 and TX0645, and distally from the E. faecalis type strains (E. faecalis ATCC19433 and 29200), and other gut commensal (e.g., E. faecalis PC1.1, 62 and Symbioflor1) and pathogenic (E. faecalis V583) strains (Figure 6A). Although E. faecalis is characterized by extensive horizontal gene transfer there is a high degree of synteny between E. faecalis AHG0090 and the closed commensal (E. faecalis 62) and pathogenic (E. faecalis V583) strains (Figure 6B).
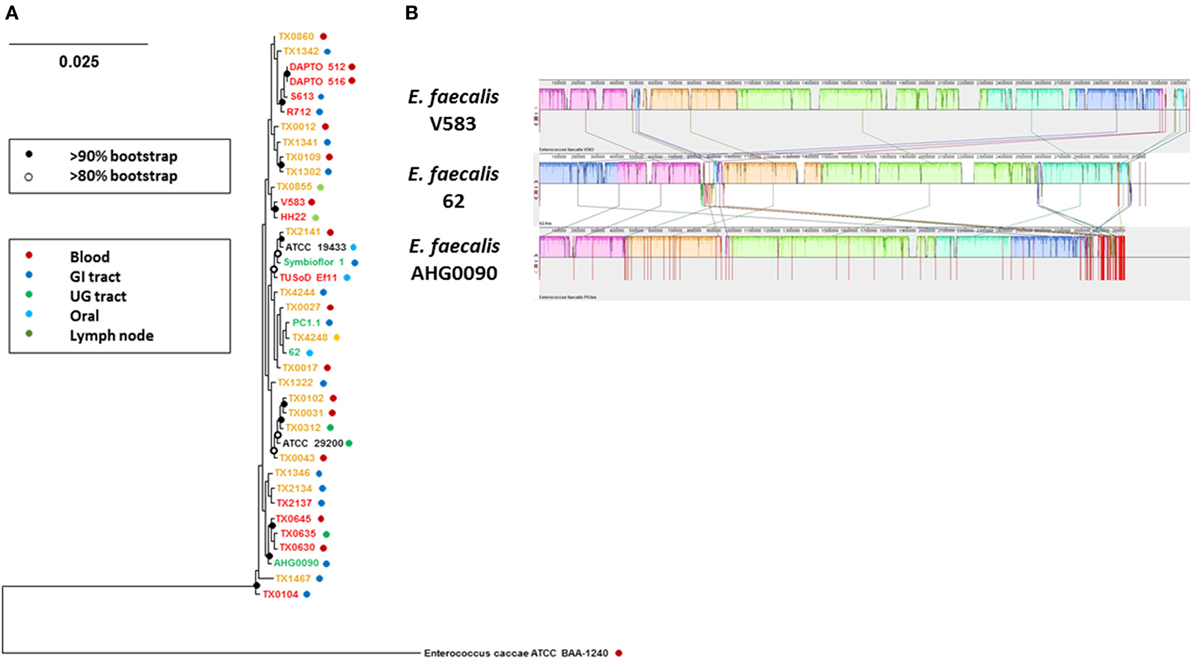
Figure 6. (A) GTDB-based phylogeny of Enterococcus faecalis as determined from the concatenation of 120 universal bacterial-specific marker genes. The source of individual commensal (green), uncharacterized (orange) and pathogenic (red) strains is indicated. The bootstrap values are indicated using a cut-off of >80 or >90%. (B) The extent of genome synteny between E. faecalis AHG0090 and a representative pathogenic strain (E. faecalis V583) and commensal (E. faecalis 62) that have closed genome sequences. The red lines for E. faecalis V583 and 62 indicated boundaries of the chromosomes and plasmids whereas the red lines for E. faecalis AHG0090 indicate individual contig sequences.
We identified several plasmids in the E. faecalis AHG0090 genome sequence. pAHG0090c is a predicted to be 76,529 bp and is comprised of 11 contigs. It is predicted to encode 80 genes and displays sequence similarity and synteny to another plasmid from E. faecalis NKH15 [pMG2200 (106,527 bp), 99% identity and 72% coverage]. Enterococcal plasmids are widely shared through horizontal transfer, and we identified an aggregation substance encoding regulon that mediates efficient contact between donor and recipient bacteria to facilitate plasmid transfer (38) and adhesion to host cells (39, 40). We also identified two closed endogenous plasmid sequences in the genome sequence data. pAHG0090b is a 5,121 bp plasmid that exhibits sequence similarity to plasmids from E. faecalis 62 [EF62pA (5,143 bp), 99% identity and 100% coverage] (41) and E. faecalis S-86 [pS86 (5,149 bp), 99% identity and 99% coverage] (42). pAHG0090b has a G+C content of 37% and is predicted to encode 6 proteins. pAHG0090a is a 1,925 bp plasmid that exhibits extensive sequence similarity to a cryptic plasmid of similar size from Enterococcus faecium 226 [pMBB1 (1,932 bp), 96% identity and 67% coverage] (43), and larger cryptic plasmids from Lactococcus fermentum KC5b [pKC5b (4,392 bp), 99% identity and 75% coverge] (44) and Lactococcus lactis [pCRL291.1 (4,640 bp), 89% identity and 70% coverage]. pAHG0090a has a G+C content of 33% and is predicted to encode a single protein predicted to function in plasmid replication.
We also identified pEHR513112 in the genome sequence. There were four differences between the compiled pEHR513112 sequence and the vector carried by E. faecalis AHG0090. We identified a G86T transversion in the cphy_3290 promoter and a C2407T transition in oriT vector modules. We also identified a 1 bp deletion at the 5′ (AscI) end and a separate 11 bp deletion at the 3′ (PmeI) end of the pMB1 module. Notably, these deletions occurred within the primer sequences used to PCR amplify the pMB1 module.
The E. faecalis AHG0090 chromosome is predicted to encode a range of niche factors that likely support colonization and persistence in the infant gut. Many enterococci are non-motile and the ability to adhere to the host epithelium likely plays a role in preventing washout. We identified an Ebp-like pilus, the microbial surface component recognizing adhesive matrix molecules protein Ace and several adhesins including EfbA that mediate adhesion to host structural factors (e.g., collagen, fibrinogen, and laminin) and support biofilm formation (45–47). E. faecalis AHG0090 also encodes several proteins that enable foraging of host glycans (48, 49) and it also encodes both the GelE and SprE proteases that have been proposed to support the nutrient requirements of the bacterium by digesting host proteins and cells (50). We also identified several proteins that likely modulate interactions with the host immune system including an internalin like protein that may support intracellular persistence (51) and a capsule that may contribute to immune evasion (52). Notably, we also identified a TIR domain protein previously shown to suppress MyD88 signaling and NF-κB activation by E. faecalis V583 (13). As expected, E. faecalis AHG0090 does not encode Mam like sequences and we did not identify any candidate genes and/or regulons likely to encode a low molecular weight NF-κB suppressive peptidic bioactive [e.g., bacteriocin CBT-SL5 (53), the E. faecalis SL-5 bacteriocin CBT-SL5 is likely same as bacteriocin ESL5 which is produced by the same strain (54)] such as the bioactive we describe.
Discussion
The early microbial colonizers of the gut help establish a homeostatic relationship between the host and its microbiota (4, 55). E. faecalis comprises part of the vaginal (12, 56) and breastmilk (57–59) microbiota, and is widely shared between mothers and their infants. It is increasingly recognized that early life events (e.g., method of birth, feeding) modify risks for several chronic diseases (60, 61) and this may be related at least in part to early differences in gut colonization and immune modulation. Much remains to be discovered about the bacteria and bioactive factors that underpin these events, and whether they could be exploited to optimize health and appropriate establishment of gut mucosal immunity.
In this study, we describe the isolation of a genetically tractable E. faecalis strain from infant stool and demonstrate that it produces a potent NF-κB suppressive bioactive. The suppressive activity of E. faecalis AHG0090 was clearly apparent in early EX phase culture supernatants and did not increase significantly from MX phase onward. This suggests the bioactive is produced in early growth and persists in the culture supernatant through the proceeding phases of growth. The closely related bacterium L. plantarum WCFS1 also exerts NF-κB and IFNγ suppressive effects and produces bioactives in a growth phase-dependent manner (37, 62), possibly as a response of the bacterium to increased nutrient limitation during the transition from mid-log to stationary phase. We determined that the ability of E. faecalis AHG0090 to suppress NF-κB in our reporter cell lines was comparable to that of F. prausnitzii A2-165. F. prausnitzii is widely regarded as a model anti-inflammatory fastidious gut bacterium and produces a 15 kDa protein termed Mam that underpins NF-κB suppression (16, 17). Mam-derived peptides are detectable in F. prausnitzii culture supernatants, however, it has not yet been reported whether the NF-κB suppressive activity is mediated by Mam and/or its peptide derivatives. While this remains to be further explored, we showed that NF-κB activation was suppressed by the <3 kDa culture supernatant fraction, and the suppressive effect was abrogated by heat or proteinase K treatment. Taken together, this suggests suppression of NF-κB by F. prausnitzii in our assays was mediated by <3 kDa Mam-derived peptides. Our data also suggests the NF-κB suppressive activity of E. faecalis AHG0090 is mediated by a low molecular weight peptidic bioactive although it is more resilient to heat and proteinase K treatment than Mam-derived peptides.
The E. faecalis AHG0090 genome sequence allowed us to readily predict “known” functionalities. Both commensal and pathogenic strains of E. faecalis have previously been shown to produce NF-κB suppressive factors (12, 13). E. faecalis V583 encodes a TIR domain containing protein, TcpF, that suppresses NF-κB by interfering with MyD88 signaling (13). NF-κB suppression by TcpF is dependent on contact between the bacterium and host cells and this protein is also encoded by E. faecalis AHG0090 and other non-pathogenic isolates [e.g., E. faecalis PC1.1 (63), E. faecalis 62 (41)]. Separately, the human vaginal isolate E. faecalis MN1 produces an NF-κB suppressive tetramic acid termed reutericyclin (12). The reutericyclin regulon has been described in L. reuteri and includes a non-ribosomal peptide synthetase (NRPS) and polyketide synthetase (PKS) enzymes that function in its biosynthesis (64). E. faecalis AHG0090 does not encode the reutericyclin regulon and nor does it encode any NRPS or PKS genes. We did not identify any Mam like sequences which is consistent with its narrow phylogenetic distribution (16) nor did we identify any genes that might encode the candidate <3 kDa peptidic bioactive. It is increasingly facile to produce microbial genomic sequence data but the ability to link genes with function remains challenging. It is estimated that the human gut microbiome is comprised of as much as 9.8 million non-redundant genes (65). However, despite the wealth of microbial (meta)genomic data that is now publicly available the vast majority of genes remain functionally uncharacterized (66, 67). For instance, it is widely acknowledged the gut microbiota exerts a broad range of immunomodulatory activities [e.g., see Ref. (68)], however, with some notable exceptions (16, 69), the genes underpinning these capacities remain largely cryptic.
Microbial culturing is a time consuming and labor-intensive process although this provides the best opportunity to link genes with function. We believe focusing culturing efforts on genetically tractable strains will ultimately expedite the functional dissection of the microbiome. We previously observed the pEHR plasmids are stably maintained in their recipient hosts (15). We have now demonstrated that they are maintained in E. faecalis AHG0090 and that this strain can be manipulated using standard molecular microbiology approaches for transformation and plasmid curing. We did identify some minor differences between the compiled pEHR vector sequences and those produced from the genome sequence data and we believe that these likely occurred during the vector construction process. We are continuing to extend the functionalities of the pEHR vector system and we anticipate that this will enable us to apply forward and/or reverse genetic approaches to functionally dissect E. faecalis AHG0090 and other gut microbes. For instance, NF-κB is a master regulator of inflammation and gut barrier integrity, and is central to the pathogenesis of several chronic (gut) diseases (70–72). The gut microbiota produces a plethora of immunomodulatory bioactives and these could be used as lead molecules to catalyze the development of new biotechnologies and therapeutics. Genetic methods offer new opportunities to identify these bioactives and they complement existing -omic-based methods for gene and protein function discovery (73, 74).
In conclusion, we demonstrated that MPM can be used to isolate genetically tractable bacteria from the human gut that possess potent anti-inflammatory activities. Although E. faecalis is among the best characterized Firmicutes affiliated gut bacteria our data suggests this taxon possesses novel anti-inflammatory capacities. Several fastidious anaerobic gut bacteria have been suggested as next generation probiotics for chronic gut diseases but E. faecalis may be a superior candidate due to its ease of propagation. We anticipate the genetic dissection of E. faecalis AHG0090 will provide new insights into the immunomodulatory capacity of this taxon, and a deeper understanding of the early life events that help establish a tolerogenic immune response.
Ethics Statement
The raw stool sample used in this study was collected from a healthy 2-year-old female child. The donor had not taken antibiotics during the 3-month period prior to collection. The child was recruited as part of a study into the link between the gut microbiota and type 1 diabetes susceptibility. All study samples were collected in accordance with the recommendations of the Mater Health Services Human Research Ethics Committee (HREC/13/MHS/21/AM02). All subjects gave written informed consent in accordance with the Declaration of Helsinki, with written consent provided from parents or legal guardians for all subjects <13 years. The protocol was approved by the Mater Health Services Human Research Ethics Committee.
Author Contributions
PÓC conceived the study with MMcG, JB, and MM; PÓC isolated Enterococcus faecalis AHG0090 and performed the genetic characterization; RG and PÓC prepared samples for analysis and performed the immunomodulatory characterizations; RG performed the immunofluorescence and gene expression experiments; EH and PÓC performed the genome analyses; PÓC, RG, EH, MMcG, JB, and MM analyzed the data; and PÓC wrote the manuscript with RG, EH, MMcG, JB, and MM.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We gratefully acknowledge the Australian Centre for Ecogenomics for the library construction and genome sequencing, and Emma Hamilton-Williams for collecting the infant fecal sample.
Funding
This research was supported via funds provided by the University of Queensland Faculty of Medicine (PÓC, JB, and MM), the University of Queensland Diamantina Institute (MM), and the University of Queensland Microbiome Challenge Grant (MM and PÓC), as part of its contribution to the International Human Microbiome Consortium. PÓC is supported by The University of Queensland’s Reginald Ferguson Fellowship in Gastroenterology. MMcG is supported by an NHMRC Senior Research Fellowship. The Translational Research Institute is supported by a grant from the Australian Government.
Footnotes
- ^http://imagej.nih.gov/ij/ (Accessed: March, 2016).
- ^https://github.com/Ecogenomics/GTDBLite (Accessed: August, 2017).
- ^http://ecogenomics.github.io/BamM/ (Accessed: August, 2017).
References
1. Chassaing B, Kumar M, Baker MT, Singh V, Vijay-Kumar M. Mammalian gut immunity. Biomed J (2014) 37:246–58. doi:10.4103/2319-4170.130922
2. Dominguez-Bello MG, Costello EK, Contreras M, Magris M, Hidalgo G, Fierer N, et al. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci U S A (2010) 107:11971–5. doi:10.1073/pnas.1002601107
3. Chung H, Kasper DL. Microbiota-stimulated immune mechanisms to maintain gut homeostasis. Curr Opin Immunol (2010) 22:455–60. doi:10.1016/j.coi.2010.06.008
4. Fulde M, Hornef MW. Maturation of the enteric mucosal innate immune system during the postnatal period. Immunol Rev (2014) 260:21–34. doi:10.1111/imr.12190
5. Hopkins MJ, Macfarlane GT, Furrie E, Fite A, Macfarlane S. Characterisation of intestinal bacteria in infant stools using real-time PCR and northern hybridisation analyses. FEMS Microbiol Ecol (2005) 54:77–85. doi:10.1016/j.femsec.2005.03.001
6. Kirtzalidou EI, Mitsou EK, Pramateftaki P, Kyriacou A. Screening fecal enterococci from Greek healthy infants for susceptibility to antimicrobial agents. Microb Drug Resist (2012) 18(6):578–85. doi:10.1089/mdr.2012.0028
7. Adlerberth I. Factors influencing the establishment of the intestinal microbiota in infancy. Nestle Nutr Workshop Ser Pediatr Program (2008) 62:13–29; discussion 29–33. doi:10.1159/000146245
8. Wopereis H, Oozeer R, Knipping K, Belzer C, Knol J. The first thousand days – intestinal microbiology of early life: establishing a symbiosis. Pediatr Allergy Immunol (2014) 25:428–38. doi:10.1111/pai.12232
9. Wang S, Ng LH, Chow WL, Lee YK. Infant intestinal Enterococcus faecalis down-regulates inflammatory responses in human intestinal cell lines. World J Gastroenterol (2008) 14:1067–76. doi:10.3748/wjg.14.1067
10. Are A, Aronsson L, Wang S, Greicius G, Lee YK, Gustafsson JA, et al. Enterococcus faecalis from newborn babies regulate endogenous PPARγ activity and IL-10 levels in colonic epithelial cells. Proc Natl Acad Sci U S A (2008) 105:1943–8. doi:10.1073/pnas.0711734105
11. Wang S, Hibberd ML, Pettersson S, Lee YK. Enterococcus faecalis from healthy infants modulates inflammation through MAPK signaling pathways. PLoS One (2014) 9:e97523. doi:10.1371/journal.pone.0097523
12. Brosnahan AJ, Merriman JA, Salgado-Pabón W, Ford B, Schlievert PM. Enterococcus faecalis inhibits superantigen toxic shock syndrome toxin-1-induced interleukin-8 from human vaginal epithelial cells through tetramic acids. PLoS One (2013) 8:e61255. doi:10.1371/journal.pone.0061255
13. Zou J, Baghdayan AS, Payne SJ, Shankar N. A TIR domain protein from E. faecalis attenuates MyD88-mediated signaling and NF-kappaB activation. PLoS One (2014) 9:e112010. doi:10.1371/journal.pone.0112010
14. Im J, Baik JE, Kim KW, Kang SS, Jeon JH, Park OJ, et al. Enterococcus faecalis lipoteichoic acid suppresses Aggregatibacter actinomycetemcomitans lipopolysaccharide-induced IL-8 expression in human periodontal ligament cells. Int Immunol (2015) 27:381–91. doi:10.1093/intimm/dxv016
15. Ó Cuív P, Smith WJ, Pottenger S, Burman S, Shanahan ER, Morrison M. Isolation of genetically tractable most-wanted bacteria by metaparental mating. Sci Rep (2015) 5:13282. doi:10.1038/srep13282
16. Quévrain E, Maubert MA, Michon C, Chain F, Marquant R, Tailhades J, et al. Identification of an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn’s disease. Gut (2016) 65:415–25. doi:10.1136/gutjnl-2014-307649
17. Breyner NM, Michon C, de Sousa CS, Vilas Boas PB, Chain F, Azevedo VA, et al. Microbial anti-inflammatory molecule (MAM) from Faecalibacterium prausnitzii shows a protective effect on DNBS and DSS-induced colitis model in mice through inhibition of NF-κB pathway. Front Microbiol (2017) 8:114. doi:10.3389/fmicb.2017.00114
18. Mcsweeney CS, Denman SE, Mackie RI. Rumen bacteria. In: Makkar HPS, Mcsweeney CS, editors. Methods in Gut Microbial Ecology for Ruminants. Dordrecht: Springer (2005). p. 23–37.
19. Hanahan D. Techniques for transformation of E. coli. In: Glover DM, editor. DNA Cloning: A Practical Approach. Oxford, United Kingdom: IRL Press (1985). p. 109–35.
20. Lyras D, Rood JI. Conjugative transfer of RP4-oriT shuttle vectors from Escherichia coli to Clostridium perfringens. Plasmid (1998) 39:160–4. doi:10.1006/plas.1997.1325
21. Green MR, Sambrook J. Preparation of plasmid DNA by alkaline lysis with sodium dodecyl sulfate: minipreps. Cold Spring Harb Protoc (2016) 2016:pdb.prot093344. doi:10.1101/pdb.prot093344
22. Lane DS. 16S and 23S rRNA sequencing. In: Stackebrandt E, Goodfellow M, editors. Nucleic Acid Techniques in Bacterial Systematics. New York, NY: John Wiley (1990). p. 115–48.
23. Ó Cuív P, Aguirre de Cárcer D, Jones M, Klaassens ES, Worthley DL, Whitehall VL, et al. The effects from DNA extraction methods on the evaluation of microbial diversity associated with human colonic tissue. Microb Ecol (2011) 61:353–62. doi:10.1007/s00248-010-9771-x
24. Van Der Helm E, Geertz-Hansen HM, Genee HJ, Malla S, Sommer MO. deFUME: dynamic exploration of functional metagenomic sequencing data. BMC Res Notes (2015) 8:328. doi:10.1186/s13104-015-1281-y
25. Cole JR, Wang Q, Fish JA, Chai B, McGarrell DM, Sun Y, et al. Ribosomal database project: data and tools for high throughput rRNA analysis. Nucleic Acids Res (2014) 42:D633–42. doi:10.1093/nar/gkt1244
26. Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol (2012) 19:455–77. doi:10.1089/cmb.2012.0021
27. Parks DH, Imelfort M, Skennerton CT, Hugenholtz P, Tyson GW. CheckM: assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res (2015) 25(7):1043–55. doi:10.1101/gr.186072.114
28. Darling AE, Mau B, Perna NT. progressiveMauve: multiple genome alignment with gene gain, loss and rearrangement. PLoS One (2010) 5:e11147. doi:10.1371/journal.pone.0011147
29. Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA, et al. The RAST server: rapid annotations using subsystems technology. BMC Genomics (2008) 9:75. doi:10.1186/1471-2164-9-75
30. Parks DH, Rinke C, Chuvochina M, Chaumeil PA, Woodcroft BJ, Evans PN, et al. Recovery of nearly 8,000 metagenome-assembled genomes substantially expands the tree of life. Nat Microbiol (2017) 2:1533–42. doi:10.1038/s41564-017-0012-7
31. Price MN, Dehal PS, Arkin AP. FastTree 2 – approximately maximum-likelihood trees for large alignments. PLoS One (2010) 5:e9490. doi:10.1371/journal.pone.0009490
32. Markowitz VM, Chen IM, Palaniappan K, Chu K, Szeto E, Grechkin Y, et al. IMG: the integrated microbial genomes database and comparative analysis system. Nucleic Acids Res (2012) 40:D115–22. doi:10.1093/nar/gkr1044
33. Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar, et al. ARB: a software environment for sequence data. Nucleic Acids Res (2004) 32:1363–71. doi:10.1093/nar/gkh293
34. Antipov D, Hartwick N, Shen M, Raiko M, Lapidus A, Pevzner PA. plasmidSPAdes: assembling plasmids from whole genome sequencing data. Bioinformatics (2016) 32:3380–7. doi:10.1093/bioinformatics/btw493
35. Ó Cuív P, de Wouters T, Giri R, Mondot S, Smith WJ, Blottière HM, et al. The gut bacterium and pathobiont Bacteroides vulgatus activates NF-kappaB in a human gut epithelial cell line in a strain and growth phase dependent manner. Anaerobe (2017) 47:209–17. doi:10.1016/j.anaerobe.2017.06.002
36. Noss EH, Nguyen HN, Chang SK, Watts GFM, Brenner MB. Genetic polymorphism directs IL-6 expression in fibroblasts but not selected other cell types. Proc Natl Acad Sci U S A (2015) 112:14948–53. doi:10.1073/pnas.1520861112
37. Zvanych R, Lukenda N, Kim JJ, Li X, Petrof EO, Khan WI, et al. Small molecule immunomodulins from cultures of the human microbiome member Lactobacillus plantarum. J Antibiot (2014) 67:85–8. doi:10.1038/ja.2013.126
38. Bhatty M, Cruz MR, Frank KL, Gomez JA, Andrade F, Garsin DA, et al. Enterococcus faecalis pCF10-encoded surface proteins PrgA, PrgB (aggregation substance) and PrgC contribute to plasmid transfer, biofilm formation and virulence. Mol Microbiol (2015) 95:660–77. doi:10.1111/mmi.12893
39. Olmsted SB, Dunny GM, Erlandsen SL, Wells CL. A plasmid-encoded surface protein on Enterococcus faecalis augments its internalization by cultured intestinal epithelial cells. J Infect Dis (1994) 170:1549–56. doi:10.1093/infdis/170.6.1549
40. Vanek NN, Simon SI, Jacques-Palaz K, Mariscalco MM, Dunny GM, Rakita RM. Enterococcus faecalis aggregation substance promotes opsonin-independent binding to human neutrophils via a complement receptor type 3-mediated mechanism. FEMS Immunol Med Microbiol (1999) 26:49–60. doi:10.1016/S0928-8244(99)00120-0
41. Brede DA, Snipen LG, Ussery DW, Nederbragt AJ, Nes IF. Complete genome sequence of the commensal Enterococcus faecalis 62, isolated from a healthy Norwegian infant. J Bacteriol (2011) 193:2377–8. doi:10.1128/JB.00183-11
42. Martinez-Bueno M, Valdivia E, Galvez A, Maqueda M. pS86, a new theta-replicating plasmid from Enterococcus faecalis. Curr Microbiol (2000) 41:257–61. doi:10.1007/s002840010130
43. Wyckoff HA, Barnes M, Gillies KO, Sandine WE. Characterization and sequence analysis of a stable cryptic plasmid from Enterococcus faecium 226 and development of a stable cloning vector. Appl Environ Microbiol (1996) 62:1481–6.
44. Pavlova SI, Kilic AO, Topisirovic L, Miladinov N, Hatzos C, Tao L. Characterization of a cryptic plasmid from Lactobacillus fermentum KC5b and its use for constructing a stable Lactobacillus cloning vector. Plasmid (2002) 47:182–92. doi:10.1016/S0147-619X(02)00005-7
45. Nallapareddy SR, Qin X, Weinstock GM, Hook M, Murray BE. Enterococcus faecalis adhesin, ace, mediates attachment to extracellular matrix proteins collagen type IV and laminin as well as collagen type I. Infect Immun (2000) 68:5218–24. doi:10.1128/IAI.68.9.5218-5224.2000
46. Montealegre MC, La Rosa SL, Roh JH, Harvey BR, Murray BE. The Enterococcus faecalis EbpA pilus protein: attenuation of expression, biofilm formation, and adherence to fibrinogen start with the rare initiation codon ATT. MBio (2015) 6:e00467–15. doi:10.1128/mBio.00467-15
47. Singh KV, La Rosa SL, Somarajan SR, Roh JH, Murray BE. The fibronectin-binding protein EfbA contributes to pathogenesis and protects against infective endocarditis caused by Enterococcus faecalis. Infect Immun (2015) 83:4487–94. doi:10.1128/IAI.00884-15
48. Bohle LA, Mathiesen G, Vaaje-Kolstad G, Eijsink VG. An endo-beta-N-acetylglucosaminidase from Enterococcus faecalis V583 responsible for the hydrolysis of high-mannose and hybrid-type N-linked glycans. FEMS Microbiol Lett (2011) 325:123–9. doi:10.1111/j.1574-6968.2011.02419.x
49. Garbe J, Sjögren J, Cosgrave EF, Struwe WB, Bober M, Olin AI, et al. EndoE from Enterococcus faecalis hydrolyzes the glycans of the biofilm inhibiting protein lactoferrin and mediates growth. PLoS One (2014) 9:e91035. doi:10.1371/journal.pone.0091035
50. Fisher K, Phillips C. The ecology, epidemiology and virulence of Enterococcus. Microbiology (2009) 155:1749–57. doi:10.1099/mic.0.026385-0
51. Brinster S, Posteraro B, Bierne H, Alberti A, Makhzami S, Sanguinetti M, et al. Enterococcal leucine-rich repeat-containing protein involved in virulence and host inflammatory response. Infect Immun (2007) 75:4463–71. doi:10.1128/IAI.00279-07
52. Thurlow LR, Thomas VC, Fleming SD, Hancock LE. Enterococcus faecalis capsular polysaccharide serotypes C and D and their contributions to host innate immune evasion. Infect Immun (2009) 77:5551–7. doi:10.1128/IAI.00576-09
53. Lee YJ, Choi HJ, Kang TW, Kim HO, Chung MJ, Park YM. CBT-SL5, a bacteriocin from Enterococcus faecalis, suppresses the expression of interleukin-8 induced by Propionibacterium acnes in cultured human keratinocytes. J Microbiol Biotechnol (2008) 18:1308–16.
54. Kang BS, Seo JG, Lee GS, Kim JH, Kim SY, Han YW, et al. Antimicrobial activity of enterocins from Enterococcus faecalis SL-5 against Propionibacterium acnes, the causative agent in acne vulgaris, and its therapeutic effect. J Microbiol (2009) 47:101–9. doi:10.1007/s12275-008-0179-y
55. Battersby AJ, Gibbons DL. The gut mucosal immune system in the neonatal period. Pediatr Allergy Immunol (2013) 24:414–21. doi:10.1111/pai.12079
56. Nami Y, Abdullah N, Haghshenas B, Radiah D, Rosli R, Yari Khosroushahi A. A newly isolated probiotic Enterococcus faecalis strain from vagina microbiota enhances apoptosis of human cancer cells. J Appl Microbiol (2014) 117:498–508. doi:10.1111/jam.12531
57. Jiménez E, Delgado S, Fernández L, García N, Albújar M, Gómez A, et al. Assessment of the bacterial diversity of human colostrum and screening of staphylococcal and enterococcal populations for potential virulence factors. Res Microbiol (2008) 159:595–601. doi:10.1016/j.resmic.2008.09.001
58. Albesharat R, Ehrmann MA, Korakli M, Yazaji S, Vogel RF. Phenotypic and genotypic analyses of lactic acid bacteria in local fermented food, breast milk and faeces of mothers and their babies. Syst Appl Microbiol (2011) 34:148–55. doi:10.1016/j.syapm.2010.12.001
59. Kozak K, Charbonneau D, Sanozky-Dawes R, Klaenhammer T. Character-ization of bacterial isolates from the microbiota of mothers’ breast milk and their infants. Gut Microbes (2015) 6:341–51. doi:10.1080/19490976.2015.1103425
60. Renz-Polster H, David MR, Buist AS, Vollmer WM, O’Connor EA, Frazier EA, et al. Caesarean section delivery and the risk of allergic disorders in childhood. Clin Exp Allergy (2005) 35:1466–72. doi:10.1111/j.1365-2222.2005.02356.x
61. Ng SC, Tang W, Leong RW, Chen M, Ko Y, Studd C, et al. Environmental risk factors in inflammatory bowel disease: a population-based case-control study in Asia-Pacific. Gut (2015) 64:1063–71. doi:10.1136/gutjnl-2014-307410
62. van Baarlen P, Troost FJ, van Hemert S, van der Meer C, de Vos WM, de Groot PJ, et al. Differential NF-kappaB pathways induction by Lactobacillus plantarum in the duodenum of healthy humans correlating with immune tolerance. Proc Natl Acad Sci U S A (2009) 106:2371–6. doi:10.1073/pnas.0809919106
63. Ó Cuív P, Klaassens ES, Smith WJ, Mondot S, Durkin AS, Harkins DM, et al. Draft genome sequence of Enterococcus faecalis PC1.1, a candidate probiotic strain isolated from human feces. Genome Announc (2013) 1:e00160–12. doi:10.1128/genomeA.00160-12
64. Lin XB, Lohans CT, Duar R, Zheng J, Vederas JC, Walter J, et al. Genetic determinants of reutericyclin biosynthesis in Lactobacillus reuteri. Appl Environ Microbiol (2015) 81:2032–41. doi:10.1128/AEM.03691-14
65. Li J, Jia H, Cai X, Zhong H, Feng Q, Sunagawa S, et al. An integrated catalog of reference genes in the human gut microbiome. Nat Biotechnol (2014) 32:834–41. doi:10.1038/nbt.2942
66. Anton BP, Chang YC, Brown P, Choi HP, Faller LL, Guleria J, et al. The COMBREX project: design, methodology, and initial results. PLoS Biol (2013) 11:e1001638. doi:10.1371/journal.pbio.1001638
67. Dantas G, Sommer MO, Degnan PH, Goodman AL. Experimental approaches for defining functional roles of microbes in the human gut. Annu Rev Microbiol (2013) 67:459–75. doi:10.1146/annurev-micro-092412-155642
68. Geva-Zatorsky N, Sefik E, Kua L, Pasman L, Tan TG, Ortiz-Lopez A, et al. Mining the human gut microbiota for immunomodulatory organisms. Cell (2017) 168:928–43.e11. doi:10.1016/j.cell.2017.01.022
69. Mazmanian SK, Liu CH, Tzianabos AO, Kasper DL. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell (2005) 122:107–18. doi:10.1016/j.cell.2005.05.007
70. Atreya I, Atreya R, Neurath MF. NF-κB in inflammatory bowel disease. J Intern Med (2008) 263:591–6. doi:10.1111/j.1365-2796.2008.01953.x
71. Sakamoto K, Maeda S. Targeting NF-kappaB for colorectal cancer. Expert Opin Ther Targets (2010) 14:593–601. doi:10.1517/14728221003769903
72. Esser N, Paquot N, Scheen AJ. Anti-inflammatory agents to treat or prevent type 2 diabetes, metabolic syndrome and cardiovascular disease. Expert Opin Investig Drugs (2015) 24:283–307. doi:10.1517/13543784.2015.974804
73. Clarke P, Ó Cuív P, O’Connell M. Novel mobilizable prokaryotic two-hybrid system vectors for high-throughput protein interaction mapping in Escherichia coli by bacterial conjugation. Nucleic Acids Res (2005) 33:e18. doi:10.1093/nar/gni011
Keywords: gut microbiota, Enterococcus faecalis, metaparental mating, genetic, Faecalibacterium prausnitzii, nuclear factors kappa B, peptide, bioactive
Citation: Ó Cuív P, Giri R, Hoedt EC, McGuckin MA, Begun J and Morrison M (2018) Enterococcus faecalis AHG0090 is a Genetically Tractable Bacterium and Produces a Secreted Peptidic Bioactive that Suppresses Nuclear Factor Kappa B Activation in Human Gut Epithelial Cells. Front. Immunol. 9:790. doi: 10.3389/fimmu.2018.00790
Received: 26 October 2017; Accepted: 29 March 2018;
Published: 18 April 2018
Edited by:
Ian Marriott, University of North Carolina at Charlotte, United StatesReviewed by:
Mario M. D’Elios, Università degli Studi di Firenze, ItalyMaryam Dadar, Razi Vaccine and Serum Research Institute, Iran
Copyright: © 2018 Ó Cuív, Giri, Hoedt, McGuckin, Begun and Morrison. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Páraic Ó Cuív, p.ocuiv@uq.edu.au
†Present address: Michael A. McGuckin, Faculty of Medicine, Dentistry and Health Sciences, The University of Melbourne, Parkville, VIC, Australia