 Jönike Dreyer1
Jönike Dreyer1 Leon M. T. Dicks
Leon M. T. Dicks- 1Department of Microbiology, Stellenbosch University, Stellenbosch, South Africa
- 2Department of Conservation Ecology and Entomology, Stellenbosch University, Stellenbosch, South Africa
The genus Xenorhabdus of the family Enterobacteriaceae, are mutualistically associated with entomopathogenic nematodes of the genus Steinernema. Although most of the associations are species-specific, a specific Xenorhabdus sp. may infect more than one Steinernema sp. During the Xenorhabdus–Steinernema life cycle, insect larvae are infected and killed, while both mutualists produce bioactive compounds. These compounds act synergistically to ensure reproduction and proliferation of the nematodes and bacteria. A single strain of Xenorhabdus may produce a variety of antibacterial and antifungal compounds, some of which are also active against insects, nematodes, protozoa, and cancer cells. Antimicrobial compounds produced by Xenorhabdus spp. have not been researched to the same extent as other soil bacteria and they may hold the answer to novel antibacterial and antifungal compounds. This review summarizes the bioactive secondary metabolites produced by Xenorhabdus spp. and their application in disease control. Gene regulation and increasing the production of a few of these antimicrobial compounds are discussed. Aspects limiting future development of these novel bioactive compounds are also pointed out.
Introduction
Since the discovery of penicillin in 1928 and the introduction of sulphonamides in 1935, more than 20 classes of antibiotics entered the market (Coates et al., 2002; Powers, 2004). The majority of these antibiotics were developed between 1940 and 1962 (Coates et al., 2011). No novel classes of antibiotics were developed between 1968 and 1998. Antibiotics developed up to 1960 protected humans from infections for approximately 50 years (Coates et al., 2011). Within 2 years of marketing, resistance is usually observed, even to new classes of compounds (Bax et al., 1998). With the current rate at which bacteria develop resistance, we may need more than 20 new classes of antibiotics to last us for the next 50 years (thus up to 2060).
Infections caused by Gram-negative bacteria are difficult to treat and many produce metallo-β-lactamase that neutralize carbapenems (Kumarasamy et al., 2010). Strains of Staphylococcus aureus developed resistance to penicillin in the early 1940s, shortly after its introduction into the market (Chambers and Deleo, 2009). Resistance to methicillin was recorded just 1 year after its introduction. Nowadays, most of the deaths caused by S. aureus are due to MRSA (Klein et al., 2007). Other pathogens in the so-called ESKAPE group for which new antibiotics are urgently needed are Enterococcus faecium, Acinetobacter baumanii, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Enterobacter species (Rice, 2010).
Another emerging pathogen is Clostridium difficile. Infections caused by C. difficile increased dramatically over the last decade, especially in patients with irritable bowel disease (IBD) (Nguyen et al., 2008). Severe cases of CDI are treated with oral metronidazole (250–500 mg four times a day for 10–14 days), or oral vancomycin (125–500 mg four times a day for 10–14 days). Metronidazole is often administered intravenously, in doses of 500 mg four times daily (Persky and Brandt, 2000). Although metronidazole is the antibiotic of choice, failure rates of 22 to 38% have been reported and many strains have developed resistance (Miller et al., 2010, 2011).
The rate at which bacteria develop resistance to antibiotics depends on the genetic characteristics of the pathogen and the mode of action of the antibiotic. For instance, resistance to rifampicin, which inhibits DNA-dependent RNA polymerase, develops much faster than resistance to antibiotics that target cell membranes (Lambert, 2005; Zhanel et al., 2008). Bacteria have several mechanisms to protect themselves against antibiotics, e.g., (i) inactivation of the antibiotic (e.g., production of β-lactamase degrading the β-lactam ring in penicillins and cephalosporins), (ii) changing membrane permeability to reduce the uptake of an antibiotic, (iii) changing efflux pumps to increase the excretion of an antibiotic from the cell, (iv) increasing production of the target enzyme, (v) finding alternative mechanisms to bypass damaged cell components, and (vi) altering the target site to render the antibiotic ineffective. Some antibiotics, such as fluoroquinolones, induce the SOS response in cells, which increases the level of errors in DNA replication and by doing so, increases resistance (Da Re et al., 2009).
It is evident that we need novel antibiotics to treat bacterial infections. Oxazolidinone (linezolid by Pfizer) and cyclic lipopeptide (daptomycin by Cubist), with activity against Gram-positive bacteria, including MRSA, are two of the most recent antibiotics released into the market (Coates et al., 2011). There may be a number of yet to be published antibiotics that are currently in preclinical development, but the overall conclusion is that we are heading for a disaster if antibiotics with broader antimicrobial activity are not developed in the next few years. The rate at which novel antibiotics are being developed is just not sufficient to control bacterial infections. We need to focus our efforts in developing antibiotics that target complex bacterial systems, such as cell membranes.
Xenorhabdus spp. produce various bioactive compounds throughout their life cycle and the genus is an underestimated and neglected source of novel bioactive compounds. All strains live in close association with entomopathogenic nematodes (EPNs) of the family Steinernematidae. Biologically active compounds produced by Xenorhabdus spp. have a broad-spectrum of antimicrobial activity, inhibiting the growth of bacteria, fungi and protozoa, the development of insects and nematodes, and the formation of cancerous cells (Webster et al., 2002). The variety of bioactive compounds produced by Xenorhabdus spp. differ, even between strains of the same species. Polyketide synthetases (PKS) and non-ribosomal peptide synthetases (NRPS) are responsible for the production of a diverse group of peptides, e.g., depsipeptides (Lang et al., 2008; Zhou et al., 2013; Kronenwerth et al., 2014), xenocoumacins (Reimer, 2013), and PAX (peptide-antimicrobial-Xenorhabdus) peptides (Fuchs et al., 2011). Other Xenorhabdus antimicrobial compounds include benzylideneacetone (Ji et al., 2004), indole derivatives (Sundar and Chang, 1993), and ribosomal-encoded bacteriocins (Thaler et al., 1995; Singh and Banerjee, 2008).
The Steinernema life cycle is shown in Figure 1. Phase I cells of Xenorhabdus are associated with reproducing nematodes, but changes to phase II when nematodes infect the insect cadaver (Akhurst, 1980). Phase I cells are larger than phase II cells, have crystalline inclusion bodies, are mobile (by swarming), and produce proteases, lipases, and bioactive compounds (Akhurst, 1982; Boemare and Akhurst, 1988). The two phases are clearly distinguished by staining with bromothymol blue and triphenyltetrazolium chloride (Kaya and Stock, 1997). Phase I cells streaked onto growth media supplemented with a combination of the two dyes form dark blue colonies with a red core and are surrounded by a clear zone. Exceptions to the rule have been reported, i.e., phase I cells not absorbing bromothymol blue (Koppenhöfer, 2007). Nematodes infected with Xenorhabdus enter the insect host through natural openings, such as the mouth, anus and respiratory spiracles, and migrates to the hemocoel. Once in the hemocoel, the nematodes enter a feeding phase, whereupon the bacteria are released (by defecation) and start to produce compounds to repress the insect’s immune system (Webster et al., 2002). The protein UnA, produced by some strains of Xenorhabdus nematophila, prevents the hemocytes of the insect to aggregate and form capsules or nodules that would surround the nematodes and bacteria (Ribeiro et al., 1999). Outer membrane proteins and lipopolysaccharides produced by X. nematophila prevents adhesion to hemocytes of Galleria mellonella Linnaeus (Dunphy and Webster, 1991). This inhibits the activation of phenoloxidase, an important enzyme in the insects’ immune response artillery (Forst et al., 1997). Some strains of X. nematophila inhibits the activity of phospholipase A2 (PLA2), which is partly responsible for the biosynthesis of eicosanoids (Dunphy and Webster, 1991; Park and Kim, 2000, 2003). The absence of eicosanoids results in severe immune depression and causes septicemia. Xenorhabdus budapestensis D43 produces a 57 kDa insecticidal protein that activates the phenoloxidase cascade and elicits an intense immune response in G. mellonella larvae (Yang et al., 2012). This leads to an excessive production of quinones, which are toxic to the larvae. X. nematophila, Xenorhabdus japonica, Xenorhabdus kozodoii, and Xenorhabdus beddingii cause apoptosis of insect hemocytes (Dunphy and Webster, 1984; Brillard et al., 2001; Cho and Kim, 2004). In the case of X. nematophila, the compound responsible for cytotoxicity has been identified as protein CyA. Production of these exoenzymes and toxins occur hours, killing the insect within 24 to 48 h (Khush and Lemaitre, 2000). With the depletion of nutrients, the juvenile nematodes switch to the infective juvenile (IJ) stage and re-associates with the bacteria. The nematodes then leave the cadaver in search of a new host (Poinar, 1990; Goodrich-Blair and Clarke, 2007; Stock and Goodrich-Blair, 2008). The genomes of Xenorhabdus spp. are large (e.g., 4.43 Mb in the case of X. nematophila) and the cells often harbor megaplasmids (e.g., 155 kba). Recent studies have shown that the genomic DNA of Xenorhabdus spp. contain many genes involved in the synthesis of insecticidal and antimicrobial compounds (Ogier et al., 2010; Chaston et al., 2011). Recent findings of gene rearrangements suggests that Xenorhabdus spp. are highly adaptable to environmental changes (Ogier et al., 2010).
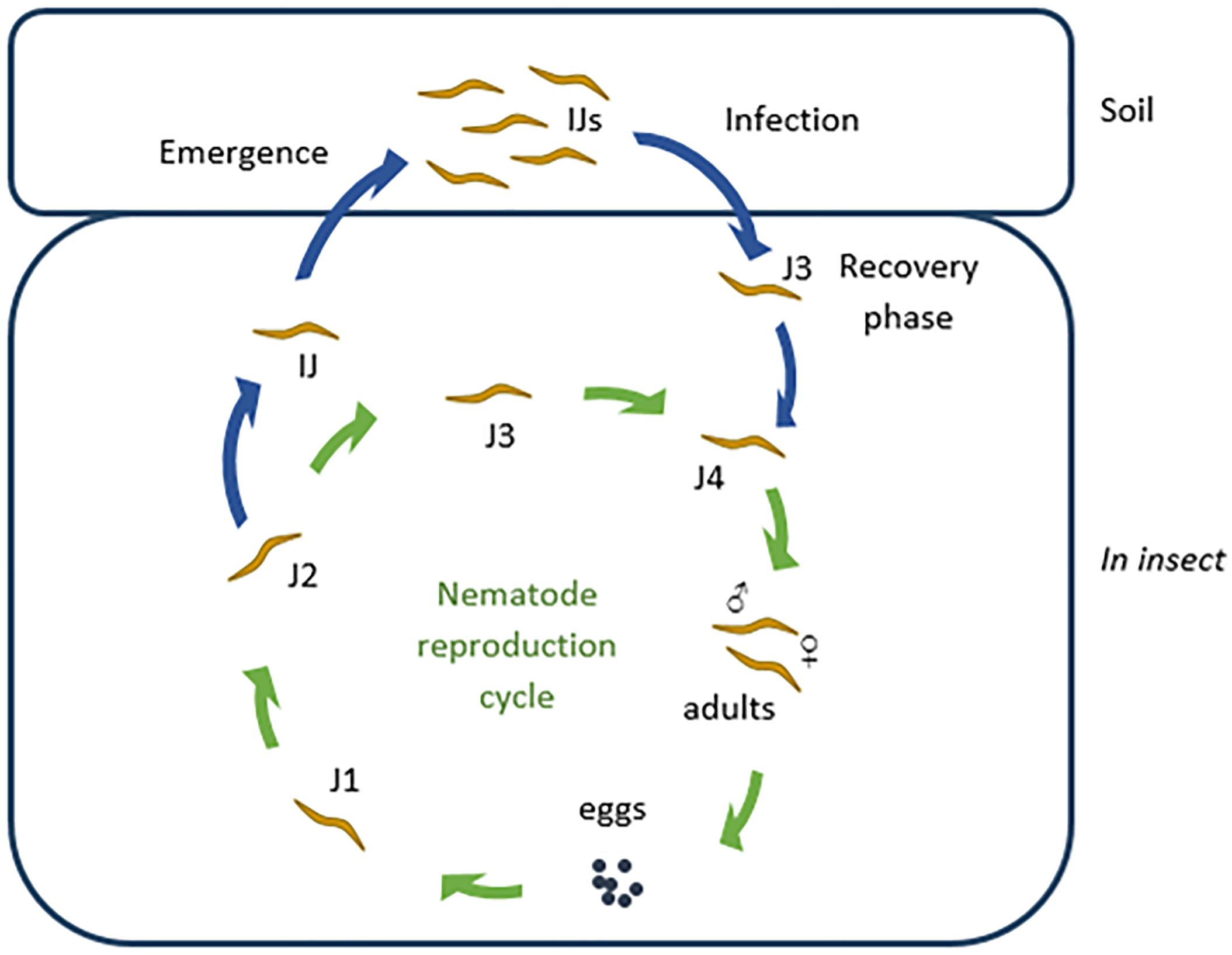
Figure 1. The Steinernema life cycle. The infective juvenile (IJ) nematodes infect an insect host and recover to the feeding phase (J3). J3 nematodes molt into fourth phase (J4) juveniles, which in turn develop into male and female adults. These adults reproduce and lay eggs. The eggs hatch as first phase juveniles (J1) which feed and molt to second, third, and fourth juvenile phases (J2–J4), and ultimately into adults. After one to three generations, when nutrients are depleted, second phase juveniles develop into IJs (special third phase juveniles). Each of the IJs host Xenorhabdus bacteria in their receptacle. These IJs then leave the cadaver and await a new prey.
Until recently, the general assumption was that a specific Xenorhabdus sp. can only infect one Steinernema species. Sicard et al. (2004) has shown that the fitness of Steinernema carpocapsae improved when associated with X. nematophila, but not when associated with non-native Xenorhabdus spp. Murfin et al. (2015) reported an increase in the fitness of Steinernema nematodes when infected with a strain of Xenorhabdus bovienii native to the nematode, or when associated with a strain from another Steinernema sp. closely related to the original nematode. Some authors hypothesized that the association of a specific Xenorhabdus sp. with more than one Steinernema sp. is an indication that the respective nematodes are phylogenetically related. The findings of Lee and Stock (2010) provided the final answer to this hypothesis by showing that host switching of Xenorhabdus spp. may occur within clades and between clades, up to 17 times. Steinernema beitlechemie from the Cameroonense-clade (Çimen et al., 2016) is associated with Xenorhabdus khoisanae. However, X. khoisanae was first isolated from Steinernema khoisanae of the Glaseri-clade (Nguyen et al., 2006; Ferreira et al., 2013). In a more recent paper, further evidence of X. khoisanae switching between clades was reported when the species was isolated from Steinernema sacchari of the Cameroonense-clade (Dreyer et al., 2017).
Steinernema nematodes infected with Xenorhabdus spp. is a highly effective natural way of controlling insect pests. Steinernema yirgalemense infected with a few as 50 infected juvenile nematodes (IJs) caused 100% mortality of false codling moth (Thaumatotibia leucotreta, Meyrick) larvae (Malan et al., 2011). Other studies have shown similar results against mealy bugs (Planococcus ficus, Signoret) (Le Vieux and Malan, 2013), sugarcane stalk borer (Eldana saccharina, Walker) (Pillay et al., 2009), fruit flies Ceratitis capitate (Wiedemann) and Ceratitis rosa (Karsch) (Malan and Manrakhan, 2009; James et al., 2018).
Secondary Metabolites Produced by Xenorhabdus Species
Xenorhabdus bacteria are known to produce broad-spectrum compounds with activity against bacteria, fungi, insects, nematodes, protozoa, and cancer cells (Webster et al., 2002). These activities each play a unique role in the protection and bioconversion of the host cadaver, and promote reproduction and growth of the nematodes. Paul et al. (1981) identified several novel antibacterial compounds produced by Xenorhabdus spp. Since this discovery, various additional bioactive Xenorhabdus compounds have been reported. Fodor et al. (2012) summarized novel antimicrobial peptides produced by Xenorhabdus szentirmaii and X. budapestensis, and focused mostly on the effect these peptides have on the plant pathogens Agrobacterium, Burkholderia, Clavibacter, Curtobacterium, Dyckeya, Erwinia, Pectobacterium, Ralstonia, Pseudomonas, Xanthomonas, Phytophthora, Pythium, Botrytis, Alternaria, and Fusarium species.
Only a few papers have been published on the regulation of antimicrobial compounds produced by Xenorhabdus spp. In X. nematophila, the leucine responsive protein (lrp) plays a role in regulating symbiosis with nematodes and pathogenicity to insects (Cowles et al., 2007; Hussa et al., 2015). Lrp may also be involved in regulating antibiotic production, as strains without the lrp gene had no antimicrobial activity toward Micrococcus luteus and Bacillus subtilis (Cowles et al., 2007). In the case of Photorhabdus luminescens, lrp led to the overproduction of desmethylphurealipid A. Injection of desmethylphurealipid A into G. mellonella and Manduca sexta larvae reduced the mRNA levels for antimicrobial-peptide-encoding genes, suggesting that these molecules may play a role in insect pathogenicity (Nollmann et al., 2015). Based on results obtained with microarray analyses (Engel et al., 2017), lrp regulates genes encoding the biosynthesis of xenematides, xenortides, rhabdopeptides, xenocoumacins and PAX-peptides, as well as six other genes with an unknown biosynthetic function. In Salmonella enterica, the LysR-type transcriptional regulator protein (LeuO) acts as a repressor for SPI-1 (Salmonella pathogenicity island) and is often described as an antagonist of heat-stable nucleoid-structuring protein (H-NS) (Chen and Wu, 2005; Espinosa and Casadesús, 2014). In Vibrio cholerae, LeuO is part of the ToxR (a membrane-spanning transcription factor) regulon and down-regulates important virulence factors. In X. nematophila and P. luminescens LeuO is described as a regulator for virulence factors and plays a role in the regulation of natural products. LeuO attenuates the production of most examined natural products, including nematophin, an antifungal and antibacterial peptide (Li et al., 1997a). Xenocoumacins, xenematides, and xenortides display antimicrobial activity against a broad spectrum of bacteria (Lang et al., 2008) and help eliminate microorganisms from the insect cadaver. Xenematides have insecticidal activity, rhabdopeptides are active against insect hemocytes (Lang et al., 2008; Reimer et al., 2013), and xenortides are cytotoxic against mammalian L6 cells (Reimer et al., 2014). These antimicrobial compounds are often produced at very low concentrations. Production levels may be scaled up by exchanging the promoters in front of biosynthesis genes with stronger or constitutive promoters (Bode et al., 2015), clone the genes into a replicative overexpression plasmid, or express the biosynthesis genes in different hosts (Schimming et al., 2014). Engel et al. (2017) reported that expression of the LeuO regulator of X. nematophila in X. szentirmaii led to overproduction of the GameXPeptide. The authors have also shown that this may lead to the production of antimicrobial compounds normally not produced by the natural host.
Several classes of structurally diverse secondary metabolites with a broad spectrum of bioactivity, including insecticidal, antifungal, antibacterial, nematicidal, and cytotoxicity, have been isolated from different Xenorhabdus strains (Brachmann and Bode, 2013). These include depsipeptides such as xenematides, xenocoumacins, fabclavines, pristinamycin, xenortides, rhabdopeptides, bicornitun, PAX peptides, cabanillasin, nemaucin, dithiolopyrrolone derivatives, indole-containing compounds, benzylideneacetone, rhabduscin, bacteriocins and a few unnamed peptides (Table 1). The diverse profile of bioactive compounds produced by Xenorhabdus spp. is supported by the large variation in gene clusters (as many as 23; Reimer et al., 2014). The secondary metabolic compounds produced by the Xenorhabdus spp. included in this review, and the accession numbers of the respective genome and gene sequences listed in GenBank, are included in Table 1. The molecular structures of the bioactive compounds are shown in Figure 2. Despite the large variation in bioactive compounds, none have been commercialized in chemical form. However, several applications of nematodes associated with Xenorhabdus symbionts have been used as biological control agents (Ehlers, 2001; Chavarría-Hernández et al., 2010).
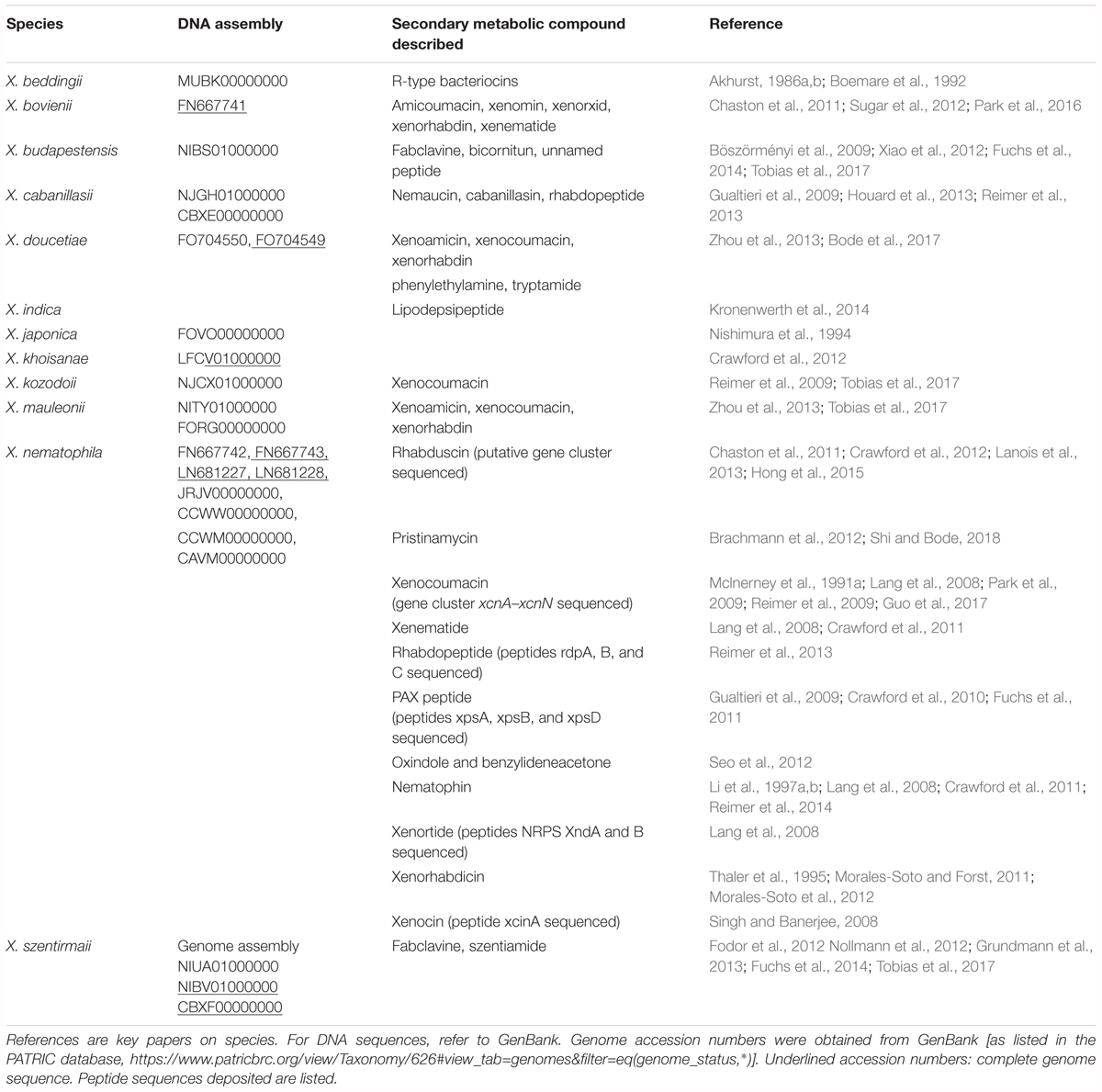
Table 1. Genome and gene assemblies of Xenorhabdus spp. and production of secondary antimicrobial compounds.
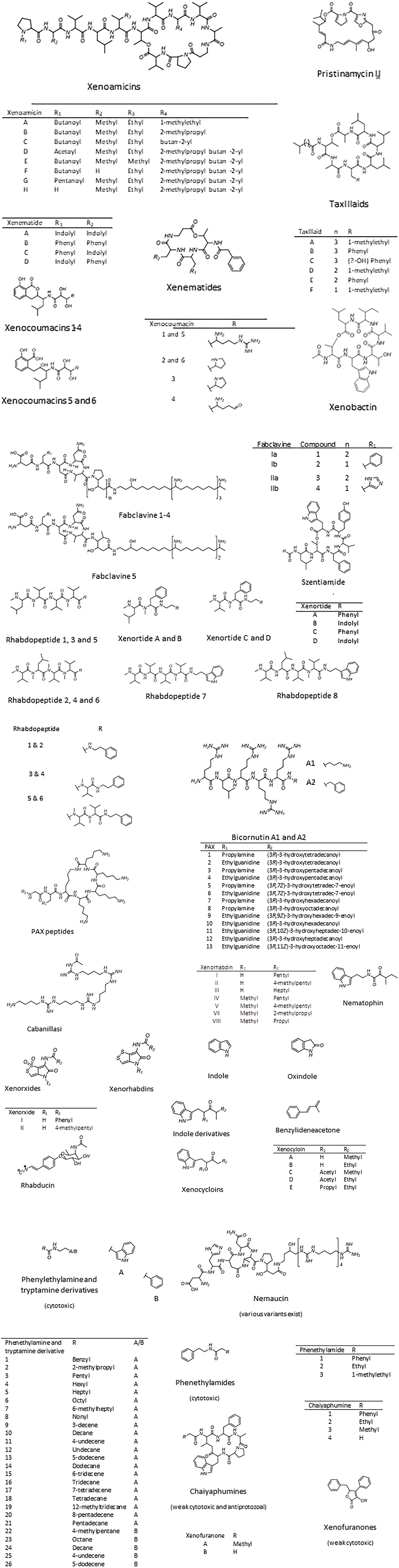
Figure 2. Xenorhabdus bioactive compounds. Bioactive compounds with unknown structures include the antibacterial xenoprotec, bicornituns C and D, and the two bacteriocins, xenorhabdicin, and xenocin. Compiled from Boemare et al. (1992), Paik et al. (2001), Hwang et al. (2003), El-Hag and El-Sadawy (2008), Gaultieri et al. (2012).
Depsipeptides
Depsipeptides are peptides with one or more amide group replaced by a hydroxy acid, leading to the formation of an ester bond. These peptides generally contain alternating peptide and ester bonds. Thus far, five classes of depsipeptides have been described. The first class, classified as xenoamicin, are tridecadepsipeptides with hydrophobic amino acids and are produced by Xenorhabdus doucetiae and Xenorhabdus mauleonii (Zhou et al., 2013). Concluded from the genome sequence of X. doucetiae DSM 17909, xenoamicins are encoded by five non-ribosomal peptide synthetases (NRPSs), XabABCD and an aspartic acid decarboxylase (XabE). Thirteen modules within XabABCD have been linked to the synthesis of xenoamicin. XabE is involved in the formation of β-alanine. Based on the hydrophobic characteristics, xenoamicin interacts with the cytoplasmic membrane. However, no antibacterial or antifungal activity has been recorded for xenoamicin A, which implies a different mode of activity. Anti-protozoal and weak cytotoxic activities have been reported for xenoamicin A, but the target sites have not been identified.
The second class of depsipeptides, the lipodepsipeptides, has an additional fatty acid chain attached to one of the amino acids and is produced by Xenorhabdus indica (Kronenwerth et al., 2014). The peptides are named after their amino acid sequence, T-A-X-L-L-L-A (X = L, F, or Y), and are referred to as taxlllaids (A-G). Seven variants were described, each classified based on the length of the fatty acid chain, the third amino acid and the overall structure of the molecule, i.e., an open chain or ring structure. The synthesis of taxlllaids are encoded by a gene cluster consisting of two NRPSs, and TxlA and TxlB, with four and three modules, respectively. Natural taxlllaid A and synthetic taxlllaids B-G have antiprotozoal activity. Taxlllaid A is also cytotoxic to human carcinoma cells (HeLa) (Kronenwerth et al., 2014).
The third class of depsipeptides are classified as indole-containing xenematides. Xenematide A was the first example, isolated from X. nematophila (Lang et al., 2008). The molecule is cyclic, antibacterial and weakly insecticidal. Three years later, Crawford et al. (2011) isolated three more xenematides (B–D) from X. nematophila and showed with gene knockout studies that XNC1_2713 encodes the production of xenematide A. Xenematides are not restricted to X. nematophila or the genus Xenorhabdus, as protein homologs have been identified in X. bovienii and Photorhabdus asymbiotica.
The final two depsipeptide classes consist of xenobactin and szentiamide (Nollmann et al., 2012; Grundmann et al., 2013). Xenobactin was isolated from an unknown Xenorhabdus sp., strain PB30.3, and szentiamide from X. szentirmaii. Both peptides are active against Plasmodium falciparum and have some activity against Trypanosoma brucei rhodesiense and Trypanosoma cruzi. Szentiamide does not have any antibacterial or antifungal activity, but has a weak cytotoxic activity against G. mellonella hemocytes. Contrary to szentiamide, xenobactin has no cytotoxic activity, but is active against M. luteus. The antibacterial activity is likely due to the hydrophobic nature of the peptide and the target in M. luteus is most probably the cell membrane.
Xenocoumacins
Xenocoumacins, first described by Mclnerney et al. (1991a), have benzopyran in the amino acid chain and are mostly produced by X. nematophila. Xcn1 is active against Gram-positive and Gram-negative bacteria, and has antifungal and anti-ulcer activity. Xcn2 has less antibacterial activity and no antifungal activity, but has anti-ulcer activity. More recently, Reimer (2013) discovered that Xcn2 is produced from Xcn1 through reactions encoded by genes xcnM and xcnN. In a study conducted by Park et al. (2009), the xcnM gene was inactivated, which led to an increased Xcn1 level, as expected, but also decreased cell viability 20-fold. The conversion of Xcn1 to Xcn2 was therefore suggested to be a mechanism used by the bacteria to avoid self-toxicity. Xcn1 is modified by various reactions to produce Xcn2, Xcn3, Xcn4, Xcn5, and Xcn6. The latter four were isolated from X. nematophila and X. kozodoii (Reimer et al., 2009).
Fabclavines
Fabclavines, or peptide-polyketide-polyamino peptides, are produced by X. budapestensis and X. szentirmaii and are encoded by a combined PKS-NRPS gene cluster (Fuchs et al., 2014). The peptide moiety is synthesized by the FclI and FclJ NRPSs, while in the PKS, FclK is responsible for catalyzing the elongation of the peptide moiety’s proline residue. These peptides are active against Gram-positive and Gram-negative bacteria, Saccharomyces cerevisiae, Plasmodium falciparum, Trypanosoma brucei, and Trypanosoma cruzi. Fabclavines and cationic antimicrobial peptides are structurally very similar. The latter displayed increased activity when combined with antibiotics (Hancock, 2001).
Pristinamycin
Pristinamycin forms part of the streptogramin A family of antibiotics and was until recently known to be produced by streptomycetes only. Pristinamycin consists of approximately 30% pristinamycin I and 70% pristinamycin II. Component II occurs in two forms, pristinamycin IIA and IIB, of which IIA is the most abundant (Blanc et al., 1995). Pristinamycin IIA is also produced by X. nematophila and is encoded by PKS/NRPS (Brachmann et al., 2012). The biosynthetic gene clusters in X. nematophila and Streptomyces pristinaspiralis are very similar. Interestingly, further analysis of X. nematophila showed that it does not contain a gene cluster for the biosynthesis of pristinamycin IA. The pxn (pristinamycin IIA, X. nematophila) gene cluster, however, is associated with transposases, suggesting that the genes were obtained through horizontal gene transfer. This might explain the absence of the pristinamycin IA gene cluster in X. nematophila.
Xenortides
To date, four xenortides, namely xenortides A–D, have been identified from X. nematophila (Lang et al., 2008; Crawford et al., 2011; Reimer et al., 2014). These peptides are biosynthesized by a gene cluster consisting of two NRPS genes (xndA and xndB). Xenortides have weak antiprotozoal activity, with the tryptamides (xenortides B and D) being more active than the phenylethylamides (xenortides A and C), and xenortide B being the most active (Reimer et al., 2014).
Rhabdopeptides
Rhabdopeptides are linear, non-ribosomally produced, and structurally similar to xenortides. A total of eight rhabdopeptides have been identified, rhabdopeptides 1, 2, 3, and 4 are from X. nematophila, and 7 and 8 from Xenorhabdus cabanillasii (Reimer et al., 2013). Rhabdopeptide 2 has weak cytotoxic activity against myoblasts. Rhabdopeptides 2, 7, and 8 have antiprotozoal activity, and 7 and 8 are weakly hemotoxic. These peptides are produced at high concentrations 4 days after infection, but levels stagnate after 10 days, suggesting that they are important during the insect bioconversion and nematode reproduction stages. The gene cluster responsible for the biosynthesis of these peptides consists of a three-module NRPS gene, RdpABC.
Bicornitun
Xenorhabdus budapestensis produce the arginine rich, bioactive compounds bicornitun A1, A2, B, and C (Böszörményi et al., 2009). The NRPS responsible for the production of bicornitun A1 was identified as BicA. This was determined by cloning the bicA gene, which encodes BicA, into an expression vector and expressing bicornitun A1 in Escherichia coli (Fuchs et al., 2012). The bicornitun complex (a combination of bicornitun A–C) is cytotoxic toward Phytophthora nicotianae by inhibiting colony formation, as well as mycelial growth. Erwinia amylovora and Bacillus subtilis is also susceptible to the bicornitun complex.
PAX Peptides
PAX peptides 1 to 5 were first identified by Gualtieri et al. (2009) and described as lysine-rich cyclolipopeptides. These peptides, produced by X. nematophila, have antifungal and antibacterial activity. However, they do not show cytotoxic activity and did not lead to increased mortality when injected into insects. An additional eight PAX peptides were identified and their structures elucidated by Fuchs et al. (2011). Three NRPS genes (paxABC) are responsible for the biosynthesis of the PAX compounds. The three NRPSs (PaxA, PaxB, and PaxC) contains 3, 9, and 10 domains, respectively.
Cabanillasin and Nemaucin
The peptides cabanillasin and nemaucin were isolated from X. cabanillasii. Cabanillasin was efficient at inhibiting the growth of human pathogenic filamentous fungi and yeasts (Houard et al., 2013). Nemaucin was, active against methicillin resistant S. aureus (MRSA). Common genes may be involved in the production of these two peptides, as both have four units of the amino-1 guanidino-butane moiety and are produced by the same organism. Nemaucin is, however, structurally more similar to fabclavine 1a from X. budapestensis than cabanillasin, and differs only by having a shorter C-terminal (Fuchs et al., 2014).
Dithiolopyrrolone Derivatives
These derivatives include the two metabolites, xenorhabdins and xenorxides. Xenorhabdins have a typical heterobicyclic pyrrolinonodithiole core, which is characteristic of dithiolopyrrolone compounds (Challinor and Bode, 2015). Xenorxides, in turn, are structurally similar to xenorhabdins and are produced when the sulfur moiety of xenorhabdins is oxidized (Webster et al., 1996). Xenorhabdins and xenorxides have antibacterial, antifungal and insecticidal activities (Mclnerney et al., 1991a,b; Li et al., 1995). Some of these dithiopyrrolone derivatives have anticancer properties. The suggested mode of action is inhibition of RNA synthesis (Joshi et al., 1982; Oliva et al., 2001).
Indole-Containing Compounds
Indole is an aromatic heterocyclic compound, consisting of a fused pyrrole- and benzene ring (Sainsbury, 2001). Various bacterial species produce indole and indole derivatives that play a role in the regulation of bacterial physiology (Lee and Lee, 2010). Indole derivatives isolated form X. nematophila and X. bovienii are active against Gram-positive and Gram-negative bacteria, as well as fungi. Sundar and Chang (1993) studied these compounds and found the mode of action the inhibition of RNA synthesis. Growing bacteria have a relatively narrow range of ppGpp concentrations and indole derivatives increase this concentration, leading to a reduction in RNA synthesis and, ultimately, a reduction in growth rate. Seo et al. (2012) identified the indole-containing compound, oxindole, as well as indole, also produced by X. nematophila. These compounds have weak phospholipase A2 inhibitory effects. As mentioned elsewhere, phospolipase A2 is an enzyme required for the production of eicosanoids. Eicosanoids, in turn, are crucial for activating an immune response in the insect by modulating and mediating hemocyte behavior (Shrestha and Kim, 2007). It is thus safe to assume that indole-containing compounds inhibit the immune response of the insect, making it more susceptible to microbial infection. Proschak et al. (2014) identified additional indole derivatives, xenocycloins (A-E), also produced by X. bovienii. These compounds have no antibacterial activities, but xenocycloins B and D are active against G. mellonella hemocytes. Xenocyloins therefore, also contribute to the insecticidal activity of these bacteria. Xenematides, discussed under depsipeptides, are also known to contain the indole structure.
Another indole containing compound, nematophin, is highly active against MRSA strains (Li et al., 1997a). In a study conducted by Li et al. (1997b), minimal inhibitory concentrations of nematophin and its derivatives against S. aureus strains were determined and it was proven that compounds with an α-carbonyl acyl group inhibited the growth of S. aureus. However, compounds where the α-carbonyl acyl group was reduced or transferred to a corresponding α-methoximino acyl group, bioactivity decreased or disappeared. It was therefore suggested, that this α-carbonyl acyl group is essential for the bioactivity of these compounds.
Unnamed Peptides
Two antimicrobial peptides, GP-19 and EP-20, have been isolated from X. budapestensis (Xiao et al., 2012). These peptides show broad-spectrum antimicrobial activity against fungi and bacteria, but the mode of action is yet to be unraveled. GP-19 has a neutral charge and is proposed to cause a disruptive effect on the membrane by mobilizing to the cell surface and possibly penetrating the membrane. As EP-20 has a negative charge it most likely does not have the same mode of action. This peptide is proposed to have an intracellular effect, by inhibiting cell wall, nucleic acid, and protein synthesis.
Benzylideneacetone
The moderately hydrophobic compound, benzylideneacetone, isolated from X. nematophila, is active against Gram-negative plant pathogenic bacteria. This compound has been used in the industry for various applications, including as a flavoring additive in soaps, cosmetics, detergents and cigarettes, as well as a food additive in candy, gelatin, and puddings. Even though it has been used for some time, it was only discovered in 2004 to have antibacterial activity (Ji et al., 2004). Benzylideneacetone also inhibits phospholipases A2, which, as described, results in the inhibition of the immune response of the insect (Seo et al., 2012).
Rhabduscin
Rhabduscin is an insecticidal tyrosine derivative, produced by X. nematophila. The insecticidal activity of this compound is achieved by inhibiting the enzyme phenoloxidase to a low nanomolar-level with an IC50 measurement of approximately 64.1 nM. Phenoloxidase is important in the melanization pathway of the insect’s immune system. Inhibition thereof leads to inhibition of one of the primary innate immune responses (Crawford et al., 2012).
Bacteriocins
Xenorhabdus bacteria also produce bacteriocins, for example, xenocin, which is produced by X. nematophila. Interestingly, the antibacterial activity of xenocin was only observed when bacterial strains were grown in minimal medium and not in enrichment medium such as Luria or nutrient broth. Xenocin production is triggered by a low iron concentration. The role of iron depletion has been proposed to be linked to an iron repressed protein, which may act as a toxin receptor on sensitive bacterial strains. This bacteriocin is therefore, only produced in the host larva when nutrient concentrations are low and competition intensifies (Singh and Banerjee, 2008). Another bacteriocin, produced by X. nematophila as well as X. bovienii, the phage tail-like xenorhabdicin, is bactericidal (Thaler et al., 1995; Morales-Soto et al., 2012). Xenorhabdus owes its activity against closely related bacteria to these bacteriocins, which are essential for keeping the environment free of other Xenorhabdus spp. and its sister genus, Photorhabdus spp. X. beddingii is also able to produce bacteriocins, however these bacteriocins have not been characterized.
Upregulating the Production of Xenorhabdus Antimicrobials
When producing antibiotics, it is of the utmost importance that the fermentation conditions are optimal to avoid the squandering of time and money. Antibiotic production in Xenorhabdus has been optimized at various time periods, mostly by one research group from the Northwest University of Agriculture and Forestry, China. This group focused on antibiotic production by X. bovienii YL002 (Fang et al., 2012) and, X. nematophila TB (Fang et al., 2010) and YL001 (Wang et al., 2008), while another group focused on a specific X. nematophila strain isolated from S. carpocapsae BJ (Xiufen et al., 2001). Factors taken into consideration for these studies were the environmental parameters; initial medium pH, temperature, rotary speed, inoculation volume, medium volume in flask, fermentation time, dissolved oxygen levels, and growth media.
As expected, the optimization for specific strains varies. There are, however a few trends in the results of these studies. The optimal fermentation conditions are a pH from 6.0 to 8.24, temperature of 25–32°C, rotary speed of 150–220 rpm, inoculation volume of 4–15%, medium volume of 54–100 ml/250 ml flask and a fermentation time of 54–72 h. The dissolved oxygen level was tested for only X. nematophila YL001 and was optimal when it was shifted during fermentation from 70% after the first 18 h to 50% for the remaining 54 h. The optimal growth media was tested for X. nematophila TB and X. bovienii YL002, however, the ingredients and amount of each ingredient differs for the respective recipes.
Crawford et al. (2010) identified one of the main compounds that leads to increased small metabolite production in X. nematophila. Xenorhabdus bacteria are known to produce higher concentrations of bioactive compounds when in G. mellonella hemolymph than grown in vitro (Maxwell et al., 1994). Therefore, it was hypothesized that one or more compounds present in insect hemolymph are responsible for activating the production of bioactive compounds. This led to the selective purification of G. mellonella hemolymph, which led to the discovery of proline as the activating signal. Supplementing bacterial cultures with D-proline did not increase the production of bioactive compounds, however, L-proline did. L-proline is thought to be a generic activating signal as it is present in various insect larvae.
The addition of L-proline to bacterial cultures led to an increase in xenematide, three indole derivatives and rhabduscin biosynthesis. Another indole-containing compound that was affected by an increase in L-proline is nematophin. This L-proline increase led to a decrease in the production of nematophin but an increase in its reduced derivative. L-proline therefore, regulates a metabolic shift in this case, rather than an increase in nematophin production.
It is evident that production of bioactive compounds requires optimization of the production protocol. This is necessary both for use in industry, as well as in research. The optimization process is however not an easy task and extended research is needed for this process, especially since the protocol will be specific for each bacterial strain and product desired.
Conclusion
Even though Xenorhabdus is not one of the generally known antimicrobial metabolite sources, it is clear to see why Pidot et al. (2014) refer to it as a neglected antibiotic source. It is evident that Xenorhabdus bacteria are an excellent source for novel antimicrobial metabolites. Various studies (Barkai-Golan, 2001; Böszörményi et al., 2009; Shapiro-Ilan et al., 2009; Fang et al., 2011), have revealed the significant potential of these bioactive secondary metabolites not only in vitro, but also in vivo. These studies investigated the use of these compounds in only the agricultural industry. However, these compounds may also be exploited in various other industries, including the healthcare and food industries.
A number of papers have been published on Xenorhabdus bacteria and their bioactive compounds. However, this is only the tip of the iceberg. A study done by Crawford et al. (2010), stated that the X. nematophila DSM 3370T genome contains various gene clusters encoding small molecule antimicrobial metabolites. The number of potential metabolites estimated to be produced by this bacterium vastly exceeds the amount of known antibiotic metabolites. Furthermore, it is generally known that different Xenorhabdus species, and even strains, produce different bioactive compounds. Therefore, it is clear that the possibilities regarding novel bioactive compounds produced by Xenorhabdus bacteria are virtually endless. Furthermore, taking into consideration the current antibiotic resistance crisis, novel antibiotic discovery is of the essence and Xenorhabdus bacteria might hold the key to human survival in the 21st century.
Although the possibilities of discovering novel antimicrobial compounds from Xenorhabdus spp. is promising, methods need to be developed to produce these compounds at much higher concentrations. This may be difficult, as most of these antimicrobial compounds are produced non-ribosomally and are thus not a single gene product. However, increased production will follow as we gain more insight on the control of the metabolic pathways. Further research is also required on the effect different growth media and nutrients may have on the production of these bioactive compounds.
Author Contributions
JD and LD wrote the manuscript. AM revised the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank the National Research Foundation of South Africa for a scholarship to JD.
References
Akhurst, R. (1980). Morphological and functional dimorphism in Xenorhabdus spp., bacteria symbiotically associated with the insect pathogenic nematodes Neoaplectana and Heterorhabditis. J. Gen. Microbiol. 121, 303–309. doi: 10.1099/00221287-121-2-303
Akhurst, R. J. (1982). Antibiotic activity of Xenorhabdus spp., bacteria symbiotically associated with insect pathogenic nematodes of the families Heterorhabditidae and Steinernematidae. J. Gen. Microbiol. 128, 3061–3065. doi: 10.1099/00221287-128-12-3061
Akhurst, R. J. (1986a). Xenorhabdus nematophilus subsp. beddingii (Enterobacteriaceae): a new subspecies of bacteria mutualisticallly associated with entomopathogenic nematodes. Int. J. Syst. Bacteriol. 36, 454–457. doi: 10.1099/00207713-36-3-454
Akhurst, R. J. (1986b). Xenorhabdus nematophilus subsp. poinarii: its interaction with insect pathogenic nematodes. Syst. Appl. Microbiol. 8, 142–147. doi: 10.1016/S0723-2020(86)80162-X
Barkai-Golan, R. (2001). Postharvest Diseases of Fruits and Vegetables: Development and Control, 1st Edn. Amsterdam: Elsevier.
Bax, R. P., Anderson, R., Crew, J., Fletcher, P., Johnson, T., Kaplan, E., et al. (1998). Antibiotic resistance – what can we do? Nat. Med. 4, 545–546. doi: 10.1038/nm0598-545
Blanc, V., Lagneaux, D., Didier, P., Gil, P., Lacroix, P., and Crouzet, J. (1995). Cloning and analysis of structural genes from Streptomyces pristinaespiralis encoding enzymes involved in the conversion of pristinamycin IIB to pristinamycin IIA (PIIA): PIIA synthase and NADH:riboflavin 5’-phosphate oxidoreductase. J. Bacteriol. 177, 5206–5214. doi: 10.1128/jb.177.18.5206-5214.1995
Bode, E., Brachmann, A. O., Kegler, C., Simsek, R., Dauth, C., Zhou, Q., et al. (2015). Simple “on-demand” production of bioactive natural products. Chembiochem 16, 1115–1119. doi: 10.1002/cbic.201500094
Bode, E., He, Y., Vo, T. D., Schultz, R., Kaiser, M., and Bode, H. B. (2017). Biosynthesis and function of simple amides in Xenorhabdus doucetiae. Environ. Microbiol. 19, 4564–4575. doi: 10.1111/1462-2920.13919
Boemare, N. E., and Akhurst, R. J. (1988). Biochemical and physiological characterization of colony form variants in Xenorhabdus spp. (Enterobacteriaceae). J. Gen. Microbiol. 134, 751–761. doi: 10.1099/00221287-134-3-751
Boemare, N. E., Boyer-Giglio, M. H., Thaler, J. O., Akhurst, R. J., and Brehelin, M. (1992). Lysogeny and bacteriocinogeny in Xenorhabdus nematophilus and other Xenorhabdus spp. Appl. Environ. Microbiol. 58, 3032–3037.
Böszörményi, E., Érsek, T., Fodor, A. M., Fodor, A. M., Földes, L. S., Hevesi, M., et al. (2009). Isolation and activity of Xenorhabdus antimicrobial compounds against the plant pathogens Erwinia amylovora and Phytophthora nicotianae. J. Appl. Microbiol. 107, 746–759. doi: 10.1111/j.1365-2672.2009.04249.x
Brachmann, A. O., and Bode, H. B. (2013). Identification and bioanalysis of natural products from insect symbionts and pathogens. Adv. Biochem. Eng. Biotechnol. 135, 123–155. doi: 10.1007/10_2013_192
Brachmann, A. O., Reimer, D., Lorenzen, W., Alonso, E. A., Kopp, Y., Piel, J., et al. (2012). Reciprocal cross talk between fatty acid and antibiotic biosynthesis in a nematode symbiont. Angew. Chem. Int. Ed. Engl. 51, 12086–12089. doi: 10.1002/anie.201205384
Brillard, J., Ribeiro, C., Boemare, N., Brehélin, M., and Givaudan, A. (2001). Two distinct hemolytic activities in Xenorhabdus nematophila are active against immunocompetent insect cells. Appl. Environ. Microbiol. 67, 2515–2525. doi: 10.1128/AEM.67.6.2515-2525.2001
Challinor, V. L., and Bode, H. B. (2015). Bioactive natural products from novel microbial sources. Ann. N. Y. Acad. Sci. 1354, 82–97. doi: 10.1111/nyas.12954
Chambers, H. F., and Deleo, F. R. (2009). Resistance Staphylococcus aureus to penicillin and methicillin. Nat. Rev. Microbiol. 7, 629–641. doi: 10.1038/nrmicro2200
Chaston, J. M., Suen, G., Tucker, S. L., Andersen, A. W., Bhasin, A., Bode, E., et al. (2011). The entomopathogenic bacterial endosymbionts Xenorhabdus and Photorhabdus: convergent lifestyles from divergent genomes. PLoS One 6:e27909. doi: 10.1371/journal.pone.0027909
Chavarría-Hernández, N., Ortega-Morales, E., Vargas-Torres, A., Chavarría-Hernández, J.-C., and Rodríguez-Hernández, A.-I. (2010). Submerged monoxenic culture of the entomopathogenic nematode, Steinernema carpocapsae CABA01, in a mechanically agitated bioreactor: evolution of the hydrodynamic and mass transfer conditions. Biotechnol. Bioprocess Eng. 15, 580–589. doi: 10.1007/s12257-009-3107-z
Chen, C.-C., and Wu, H.-Y. (2005). LeuO protein delimits the transcriptionally active and repressive domains on the bacterial chromosome. J. Biol. Chem. 280, 15111–15121. doi: 10.1074/jbc.M414544200
Cho, S., and Kim, Y. (2004). Hemocyte apoptosis induced by entomopathogenic bacteria, Xenorhabdus and Photorhabdus, in Bombyx mori. J. Asia Pac. Entomol. 7, 195–200. doi: 10.1016/j.cbpb.2014.04.003
Çimen, H., Pùža, V., Nermu’, J., Hatting, J., Ramakuwela, T., Faktorová, L., et al. (2016). Steinernema beitlechemi n. sp., a new entomopathogenic nematode (Nematoda: Steinernematidae) from South Africa. Nematogy 18, 439–453. doi: 10.1163/15685411-00002968
Coates, A., Hu, Y., Bax, R., and Page, C. (2002). The future challenges facing the development of new antimicrobial drugs. Nat. Rev. Drug Discov. 1, 895–910. doi: 10.1038/nrd940
Coates, A. R. M., Halls, G., and Hu, Y. (2011). Novel classes of antibiotics or more of the same? Br. J. Pharmacol. 163, 184–194. doi: 10.1111/j.1476-5381.2011.01250.x
Cowles, K. N., Cowles, C. E., Richards, G. R., Martens, E. C., and Goodrich-Blair, H. (2007). The global regulator Lrp contributes to mutualism, pathogenesis and phenotypic variation in the bacterium Xenorhabdus nematophila. Cell. Microbiol. 9, 1311–1323. doi: 10.1111/j.1462-5822.2006.00873.x
Crawford, J. M., Kontnik, R., and Clardy, J. (2010). Regulating alternative lifestyles in entomopathogenic bacteria. Curr. Biol. 20, 69–74. doi: 10.1016/j.cub.2009.10.059
Crawford, J. M., Portmann, C., Kontnik, R., Walsh, C. T., and Clardy, J. (2011). NRPS substrate promiscuity diversifies the xenematides. Org. Lett. 13, 5144–5147. doi: 10.1021/ol2020237
Crawford, J. M., Portmann, C., Zhang, X., Roeffaers, M. B. J., and Clardy, J. (2012). Small molecule perimeter defense in entomopathogenic bacteria. Proc. Natl. Acad. Sci. U.S.A. 109, 10821–10826. doi: 10.1073/pnas.1201160109
Da Re, S., Garnier, F., Guerin, E., Campoy, S., Denis, F., and Ploy, M. C. (2009). The SOS response promotes qnrB quinolone-resistance determinant expression. EMBO Rep. 10, 929–933. doi: 10.1038/embor.2009.99
Dreyer, J., Malan, A. P., and Dicks, L. M. T. (2017). Three novel Xenorhabdus–Steinernema associations and evidence of strains of X. khoisanae switching between different clades. Curr. Microbiol. 74, 938–942. doi: 10.1007/s00284-017-1266-2
Dunphy, G. B., and Webster, J. M. (1984). Interaction of Xenorhabdus nematophilus subsp. nematophilus with the haemolymph of Galleria mellonella. J. Insect Physiol. 30, 883–889. doi: 10.1016/0022-1910(84)90063-5
Dunphy, G. B., and Webster, J. M. (1991). Antihemocytic surface components of Xenorhabdus nematophilus var. dutki and their modification by serum nonimmune larvae of Galleria mellonella. J. Invertebr. Pathol. 58, 40–51. doi: 10.1016/0022-2011(91)90160-R
Ehlers, R.-U. (2001). Mass production of entomopathogenic nematodes for plant protection. Appl. Microbiol. Biotechnol. 56, 623–633. doi: 10.1007/s002530100711
El-Hag, H. A. A., and El-Sadawy, H. A. (2008). Xenoprotec: antimicrobial agent derived from Xenorhabdus indica. Am. Eurasian J. Agric. Environ. Sci. 3, 568–576.
Engel, Y., Windhorst, C., Lu, X., Goodrich-Blair, H., and Bode, H. B. (2017). The global regulators Lrp, LeuO, and HexA control secondary metabolism in entomopathogenic bacteria. Front. Microbiol. 8:209. doi: 10.3389/fmicb.2017.00209
Espinosa, E., and Casadesús, J. (2014). Regulation of Salmonella enterica pathogenicity island 1 (SPI-1) by the LysR-type regulator LeuO. Mol. Microbiol. 91, 1057–1069. doi: 10.1111/mmi.12500
Fang, X. L., Feng, J. T., Zhang, W. G., Wang, Y. H., and Zhang, X. (2010). Optimization of growth medium and fermentation conditions for improved antibiotic activity of Xenorhabdus nematophila TB using a statistical approach. Afr. J. Biotechnol. 9, 8068–8077. doi: 10.5897/AJB09.1939
Fang, X. L., Han, L.-R., Cao, X.-Q., Zhu, M.-X., Zhang, X., and Wang, Y.-H. (2012). Statistical optimization of process variables for antibiotic activity of Xenorhabdus bovienii. PLoS One 7:e38421. doi: 10.1371/journal.pone.0038421
Fang, X. L., Li, Z. Z., Wang, Y. H., and Zhang, X. (2011). In vitro and in vivo antimicrobial activity of Xenorhabdus bovienii YL002 against Phytophthora capsici and Botrytis cinerea. J. Appl. Microbiol. 111, 145–154. doi: 10.1111/j.1365-2672.2011.05033.x
Ferreira, T., van Reenen, C. A., Endo, A., Sproër, C., Malan, A. P., and Dicks, L. M. T. (2013). Description of Xenorhabdus khoisanae sp. nov., the symbiont of the entomopathogenic nematode Steinernema khoisanae. Int. J. Syst. Evol. Microbiol. 63, 3220–3224. doi: 10.1099/ijs.0.049049-0
Fodor, A., Varga, I., Havesi, M., Máthé-Fodor, A., Racsko, J., and Hogan, J. A. (2012). “Novel anti-microbial peptides of Xenorhabdus origin against multidrug resistant plant pathogens,” in A Search for Antibacterial Agents, ed. V. Bobbarala (Rijeka: InTech Press), 147–195.
Forst, S., Dowds, B., Boemare, N., and Stackebrandt, E. (1997). Xenorhabdus and Photorhabdus spp.: bugs that kill bugs. Annu. Rev. Microbiol. 51, 47–72. doi: 10.1146/annurev.micro.51.1.47
Fuchs, S. W., Grundmann, F., Kurz, M., Kaiser, M., and Bode, H. B. (2014). Fabclavines: bioactive peptide-polyketide-polyamino hybrids from Xenorhabdus. ChembioChem 15, 512–516. doi: 10.1002/cbic.201300802
Fuchs, S. W., Proschak, A., Jaskolla, T. W., Karas, M., and Bode, H. B. (2011). Structure elucidation and biosynthesis of lysine-rich cyclic peptides in Xenorhabdus nematophila. Org. Biomol. Chem. 9, 3130–3132. doi: 10.1039/c1ob05097d
Fuchs, S. W., Sachs, C. C., Kegler, C., Nollmann, F. I., Karas, M., and Bode, H. B. (2012). Neutral loss fragmentation pattern based screening for arginine-rich natural products in Xenorhabdus and Photorhabdus. Anal. Chem. 84, 6948–6955. doi: 10.1021/ac300372p
Gaultieri, M., Villain-Guillot, P., Givaudan, A., and Pages, S. (2012). Nemaucin, an antibiotic produced by entomopathogenic Xenorhabdus cabanillasii. Patent WO2012085177 A1. Geneva: World Intellectual Property Organization.
Goodrich-Blair, H., and Clarke, D. J. (2007). Mutualism and pathogenesis in Xenorhabdus and Photorhabdus: two roads to the same destination. Mol. Microbiol. 64, 260–268. doi: 10.1111/j.1365-2958.2007.05671.x
Grundmann, F., Kaiser, M., Kurz, M., Schiell, M., Batzer, A., and Bode, H. B. (2013). Structure determination of the bioactive depsipeptide xenobactin from Xenorhabdus sp. PB30.3. RSC Adv. 3, 22072–22077. doi: 10.1039/c3ra44721a
Gualtieri, M., Aumelas, A., and Thaler, J.-O. (2009). Identification of a new antimicrobial lysine-rich cyclolipopeptide family from Xenorhabdus nematophila. J. Antibiot. 62, 295–302. doi: 10.1038/ja.2009.31
Guo, S., Zhang, S., Fang, X., Liu, Q., Gao, J., Bilal, M., et al. (2017). Regulation of antimicrobial activity and xenocoumacins biosynthesis by pH in Xenorhabdus nematophila. Microb. Cell Fact. 16:203. doi: 10.1186/s12934-017-0813-7
Hancock, R. E. (2001). Cationic peptides: effectors in innate immunity and novel antimicrobials. Lancet Infect. Dis. 1, 156–164. doi: 10.1016/S1473-3099(01)00092-5
Hong, S. J., Ullah, I., Park, G. S., Jung, B. K., Choi, J., Khan, A. R., et al. (2015). Draft genome sequence and annotation of the insect pathogenic bacterium Xenorhabdus nematophila strain C2-3, isolated from nematode Steinernema carpocapsae in the Republic of Korea. Genome Announc. 3:e01521-14. doi: 10.1128/genomeA.01521-14
Houard, J., Aumelas, A., Noël, T., Pages, S., Givaudan, A., Fitton-Ouhabi, V., et al. (2013). Cabanillasin, a new antifungal metabolite, produced by entomopathogenic Xenorhabdus cabanillasii JM26. J. Antibiot. 66, 617–620. doi: 10.1038/ja.2013.58
Hussa, E. A., Casanova-Torres,ÁM., and Goodrich-Blair, H. (2015). The global transcription factor Lrp controls virulence modulation in Xenorhabdus nematophila. J. Bacteriol. 197, 3015–3025. doi: 10.1128/JB.00272-15
Hwang, S.-Y., Paik, S., Park, S.-H., Kim, H.-S., Lee, I.-S., Kim, S.-P., et al. (2003). N-phenethyl-2-phenylacetamide isolated from Xenorhabdus nematophilus induces apoptosis through caspase activation and calpain-mediated Bax cleavage in U937 cells. Int. J. Oncol. 22, 151–157. doi: 10.3892/ijo.22.1.151
James, M., Addison, P., and Malan, A. P. (2018). Surveying and screening South African entomopathogenic nematodes for the control of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann). Crop Protect. 105, 41–48. doi: 10.1016/j.cropro.2017.11.008
Ji, D., Yi, Y., Kang, G.-H., Choi, Y.-H., Kim, P., Baek, N.-I., et al. (2004). Identification of an antibacterial compound, benzylideneacetone, from Xenorhabdus nematophila against major plant-pathogenic bacteria. FEMS Microbiol. Lett. 239, 241–248. doi: 10.1016/j.femsle.2004.08.041
Joshi, A., Verma, M., and Chakravorty, M. (1982). Thiolutin-resistant mutants of Salmonella typhimurium. Antimicrob. Agents Chemother. 22, 541–547. doi: 10.1128/AAC.22.4.541
Kaya, H. K., and Stock, S. P. (1997). “Techniques in insect nematology,” in Manual of Techniques in Insect Pathology, ed. L. A. Lacey (London: Academic Press), 281–324. doi: 10.1016/B978-012432555-5/50016-6
Khush, R. J., and Lemaitre, B. (2000). Genes that fight infection: what the Drosophila genome says about animal immunity. Trends Genet. 16, 442–449. doi: 10.1016/S0168-9525(00)02095-3
Klein, E., Smith, D. L., and Laxminarayan, R. (2007). Hospitalizations and deaths caused by methicillin-resistant Staphylococcus aureus, United States, 1999–2005. Emerg. Infect. Dis. 13, 1840–1846. doi: 10.3201/eid1312.070629
Koppenhöfer, H. S. (2007). “Bacterial symbionts of Steinernema and Heterorhabditis,” in Entomopathogenic Nematodes: Systematics, Phylogeny and Bacterial Symbionts, 5th Edn, ed. K. B. Nguyen (Leiden: Brill), 735–808. doi: 10.1163/ej.9789004152939.i-816.43
Kronenwerth, M., Bozhüyük, K. A. J., Kahnt, A. S., Steinhilber, D., Gaudriault, S., Kaiser, M., et al. (2014). Characterisation of taxlllaids A-G; natural products from Xenorhabdus indica. Chem. Eur. J. 20, 17478–17487. doi: 10.1002/chem.201403979
Kumarasamy, K. K., Toleman, M. A., Walsh, T. R., Bagaria, J., Butt, F., Balakrishnan, R., et al. (2010). Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological and epidemiological study. Lancet Infect. Dis. 10, 597–602. doi: 10.1016/S1473-3099(10)70143-2
Lambert, P. A. (2005). Bacterial resistance to antibiotics: modified target sites. Adv. Drug Deliv. Rev. 57, 1471–1485. doi: 10.1016/j.addr.2005.04.003
Lang, G., Kalvelage, T., Peters, A., Wiese, J., and Imhoff, J. F. (2008). Linear and cyclic peptides from the entomopathogenic bacterium Xenorhabdus nematophilus. J. Nat. Prod. 71, 1074–1077. doi: 10.1021/np800053n
Lanois, A., Ogier, J. C., Gouzy, J., Laroui, C., Rouy, Z., Givaudan, A., et al. (2013). Draft genome sequence and annotation of the entomopathogenic bacterium Xenorhabdus nematophila strain F1. Genome Announc. 1:e00342-13. doi: 10.1128/genomeA.00342-13
Le Vieux, P. D., and Malan, A. P. (2013). The potential use of entomopathogenic nematodes to control Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae). S. Afr. J. Enol. Vitic. 34, 296–306.
Lee, J.-H., and Lee, J. (2010). Indole as an intercellular signal in microbial communities. FEMS Microbiol. Rev. 34, 426–444. doi: 10.1111/j.1574-6976.2009.00204.x
Lee, M.-M., and Stock, S. P. (2010). A multigene approach for assessing evolutionary relationships of Xenorhabdus spp. (γ-Proteobacteria), the bacterial symbionts of entomopathogenic Steinernema nematodes. J. Invertebr. Pathol. 104, 67–74. doi: 10.1016/j.jip.2010.01.005
Li, J., Chen, G., and Webster, J. M. (1997a). Nematophin, a novel antimicrobial substance produced by Xenorhabdus nematophilus (Enterobactereaceae). Can. J. Microbiol. 43, 770–773.
Li, J., Chen, G., and Webster, J. M. (1997b). Synthesis and antistaphylococcal activity of nematophin and its analogs. Bioorg. Med. Chem. Lett. 7, 1349–1352. doi: 10.1016/S0960-894X(97)00223-0
Li, J., Chen, G., Webster, J. M., and Czyzewska, E. (1995). Antimicrobial metabolites from a bacterial symbiont. J. Nat. Prod. 58, 1081–1086. doi: 10.1021/np50121a016
Malan, A. P., Knoetze, R., and Moore, S. D. (2011). Isolation and identification of entomopathogenic nematodes from citrus orchards in South Africa and their biocontrol potential against false codling moth. J. Invertebr. Pathol. 108, 115–125. doi: 10.1016/j.jip.2011.07.006
Malan, A. P., and Manrakhan, A. (2009). Susceptibility of the Mediterranean fruit fly (Ceratitis capitata) and the Natal fruit fly (Ceratitis rosa) to entomopathogenic nematodes. J. Invertebr. Pathol. 100, 47–49. doi: 10.1016/j.jip.2008.09.007
Maxwell, P. W., Chen, G., Webster, J. M, and Dunphy, G. B. (1994). Stability and activities of antibiotics produced during infection of the insect Galleria mellonella by two isolates of Xenorhabdus nematophilus. Appl. Environ. Microbiol. 60, 715–721.
Mclnerney, B. V., Gregson, R. P., Lacey, M. J., Akhurst, R. J., Lyons, G. R., Rhodes, S. H., et al. (1991a). Biologically active metabolites from Xenorhabdus spp., part 1. Dthiolopyrrolone derivatives with antibiotic activity. J. Nat. Prod. 54, 774–784.
McInerney, B. V., Taylor, W. C., Lacey, M. J., Akhurst, R. J., and Gregson, R. P. (1991b). Biologically active metabolites from Xenorhabdus spp., part 2. Benzopyran-1-one derivatives with gastroprotective activity. J. Nat. Prod. 54, 785–795.
Miller, C. B., Chen, L., Sexton, D., and Anderson, D. (2011). Comparison of the burdens of hospital-onset, healthcare facility-associated Clostridium difficile infection and of healthcare-associated infection due to methicillin-resistant Staphylococcus aureus in community hospitals. Infect. Control Hosp. Epidemiol. 32, 387–390. doi: 10.1086/659156
Miller, C. B., Dellon, E., Isaacs, K., and Gangarosa, L. (2010). Fecal bacteriotherapy via colonoscopy as rescue therapy for refractory and recurrent Clostridium difficile-associated diarrhea. Am. J. Gastroenterol. 105:S323.
Morales-Soto, N., and Forst, S. A. (2011). The xnp1 P2-like tail synthesis gene cluster encodes xenorhabdicin and is required for interspecies competition. J. Bacteriol. 193, 3624–3632. doi: 10.1128/JB.00092-11
Morales-Soto, N., Gaudriault, S., Ogier, J. C., Thappeta, K. R. V., and Forst, S. (2012). Comparative analysis of P2-type remnant prophage loci in Xenorhabdus bovienii and Xenorhabdus nematophila required for xenorhabdicin production. FEMS Microbiol. Lett. 333, 69–76. doi: 10.1111/j.1574-6968.2012.02600.x
Murfin, K. E., Lee, M.-M., Klassen, J. L., McDonald, B. R., Larget, B., Forst, S., et al. (2015). Xenorhabdus bovienii strain diversity impacts coevolution and symbiotic maintenance with Steinernema spp. nematode hosts. mBio 6:e00076. doi: 10.1128/mBio.00076-15
Nguyen, G. C., Kaplan, G. G., Harris, M. L., and Brant, S. R. (2008). A national survey of the prevalence and impact of Clostridium difficile infection among hospitalized inflammatory bowel disease patients. Am. J. Gastroenterol. 103, 1443–1450. doi: 10.1111/j.1572-0241.2007.01780.x
Nguyen, K. B., Malan, A., and Gozel, U. (2006). Steinernema khoisanae n. sp. (Rhabditida: Steinernematidae), a new entomopathogenic nematode from South Africa. Nematology 8, 157–175. doi: 10.1017/S0022149X15000097
Nishimura, Y., Hagiwara, A., Suzuki, T., and Yamanaka, S. (1994). Xenorhabdus japonicus sp-nov associated with the nematode Steinernema kushidai. World J. Microbiol. Biotechnol. 10, 207–210. doi: 10.1007/BF00360889
Nollmann, F. I., Dowling, A., Kaiser, M., Deckmann, K., Grösch, S., French-Constant, R., et al. (2012). Synthesis of szentiamide, a depsipeptide from entomopathogenic Xenorhabdus szentirmaii with activity against Plasmodium falciparum. Beilstein J. Org. Chem. 8, 528–533. doi: 10.3762/bjoc.8.60
Nollmann, F. I., Heinrich, A. K., Brachmann, A. O., Morisseau, C., Mukherjee, K., Casanova-Torres, ÁM., et al. (2015). A Photorhabdus natural product inhibits insect juvenile hormone epoxide hydrolase. Chembiochem 16, 766–771. doi: 10.1002/cbic.201402650
Ogier, J.-C., Calteau, A., Forst, S., Goodrich-Blair, H., Roche, D., Rouy, Z., et al. (2010). Units of plasticity in bacterial genomes: new insight from the comparative genomics of two bacteria interacting with invertebrates, Photorhabdus and Xenorhabdus. BMC Genomics 11:568. doi: 10.1186/1471-2164-11-568
Oliva, B., O’Neill, A., Wilson, J. M., O’Hanlon, P. J., and Chopra, I. (2001). Antimicrobial properties and mode of action of the pyrrothine holomycin. Antimicrob. Agents Chemother. 45, 532–539. doi: 10.1128/AAC.45.2.532-539.2001
Paik, S., Park, Y. H., Suh, S. II, Kim, H. S., Lee, I. S., Park, M. K., et al. (2001). Unusual cytotoxic phenethylamides from Xenorhabdus nematophilus. Bull. Korean Chem. Soc. 22, 372–374.
Park, D., Ciezki, K., van der Hoeven, R., Singh, S., Reimer, D., Bode, H. B., et al. (2009). Genetic analysis of xenocoumacin antibiotic production in the mutualistic bacterium Xenorhabdus nematophila. Mol. Microbiol. 73, 938–949. doi: 10.1111/j.1365-2958.2009.06817.x
Park, H. B., Perez, C. E., Perry, E. K., and Crawford, J. M. (2016). Activating and attenuating the amicoumacin antibiotics. Molecules 21:E824. doi: 10.3390/molecules21070824
Park, Y., and Kim, Y. (2000). Eicosanoids rescue Spodoptera exigua infected with Xenorhabdus nematophilus, the symbiotic bacteria to the entomopathogenic nematode Steinernema carpocapsae. J. Insect Physiol. 46, 1469–1476. doi: 10.1016/S0022-1910(00)00071-8
Park, Y., and Kim, Y. (2003). Xenorhabdus nematophilus inhibits p-bromophenacyl bromide (BPB)-sensitive PLA2 of Spodoptera exigua. Arch. Insect Biochem. Physiol. 54, 134–142. doi: 10.1002/arch.10108
Paul, V. J., Frautschy, S., Fenical, W., and Nealson, K. H. (1981). Antibiotics in microbial ecology. J. Chem. Ecol. 7, 589–597. doi: 10.1007/BF00987707
Persky, S. E., and Brandt, L. J. (2000). Treatment of recurrent Clostridium difficile-associated diarrhea by administration of donated stool directly through a colonoscope. Am. J. Gastroenterol. 95, 3283–3285.
Pidot, S. J., Coyne, S., Kloss, F., and Hertweck, C. (2014). Antibiotics from neglected bacterial sources. Int. J. Med. Microbiol. 304, 14–22. doi: 10.1016/j.ijmm.2013.08.011
Pillay, U., Martin, L. A., Rutherford, R. S., and Berry, S. D. (2009). Entomopathogenic nematodes in sugarcane in South Africa. Proc. S. Afr. Technol. Ass. 82, 538–541.
Poinar, G. O. (1990). “Biology and taxonomy of Steinernematidae and Heterorhabditidae,” in Entomopathogenic Nematodes in Biological Control, eds R. Gaugler and H. K. Kaya (Boca Raton, FL: CRC Press), 23–62.
Powers, J. H. (2004). Antimicrobial drug development – the past, the present, and the future. Clin. Microbiol. Infect. 4, 23–31. doi: 10.1111/j.1465-0691.2004.1007.x
Proschak, A., Zhou, Q., Schöner, T., Thanwisai, A., Kresovic, D., Dowling, A., et al. (2014). Biosynthesis of the insecticidal xenocyloins in Xenorhabdus bovienii. Chembiochem 15, 369–372. doi: 10.1002/cbic.201300694
Reimer, D. (2013). Identification and Characterization of Selected Secondary Metabolite Biosynthetic Pathways from Xenorhabdus nematophila. Ph.D. thesis, Johann Wolfgang Groethe-Universität, Frankfurt.
Reimer, D., Cowles, K. N., Proschak, A., Nollmann, F. I., Dowling, A. J., Kaiser, M., et al. (2013). Rhabdopeptides as insect-specific virulence factors from entomopathogenic bacteria. Chembiochem 14, 1991–1997. doi: 10.1002/cbic.201300205
Reimer, D., Luxenburger, E., Brachmann, A. O., and Bode, H. B. (2009). A new type of pyrrolidine biosynthesis is involved in the late steps of xenocoumacin production in Xenorhabdus nematophila. Chembiochem 10, 1997–2001. doi: 10.1002/cbic.200900187
Reimer, D., Nollmann, F. I., Schultz, K., Kaiser, M., and Bode, H. B. (2014). Xenortide biosynthesis by entomopathogenic Xenorhabdus nematophila. J. Nat. Prod. 77, 1976–1980. doi: 10.1021/np500390b
Ribeiro, C., Duvic, B., Oliveira, P., Givaudan, A., Palha, F., Simoes, N., et al. (1999). Insect immunity-effects of factors produced by a nematobacterial complex on immunocompetent cells. J. Insect Pathol. 45, 677–685. doi: 10.1016/S0022-1910(99)00043-8
Rice, L. B. (2010). Progress and challenges in implementing the research on ESKAPE pathogens. Infect. Control Hosp. Epidemiol. 31(Suppl. 1), S7–S10. doi: 10.1086/655995
Sainsbury, M. (2001). “Benzo[b]pyrrole, benzo[b]furan and benzo[b]thiophene,” in Heterocyclic Chemistry, Vol. 8, ed. M. Sainsbury (Cambridge: Royal Society of Chemistry), 79–114.
Schimming, O., Fleischhacker, F., Nollmann, F. I., and Bode, H. B. (2014). Yeast homologous recombination cloning leading to the novel peptides ambactin and xenolindicin. Chembiochem 15, 1290–1294. doi: 10.1002/cbic.201402065
Seo, S., Lee, S., Hong, Y., and Kim, Y. (2012). Phospholipase A2 inhibitors synthesized by two entomopathogenic bacteria, Xenorhabdus nematophila and Photorhabdus temperate subsp. temperata. Appl. Environ. Microbiol. 78, 3816–3823. doi: 10.1128/AEM.00301-12
Shapiro-Ilan, D. I., Reilly, C. C., and Hotchkiss, M. W. (2009). Suppressive effects of metabolites from Photorhabdus and Xenorhabdus spp. on phytopathogens of peach and pecan. Arch. Phytopathol. Plant Prot. 42, 715–728. doi: 10.1080/03235400701390539
Shi, Y.-M., and Bode, H. B. (2018). Chemical language and warfare of bacterial natural products in bacteria–nematode–insect interactions. Nat. Prod. Rep. 35, 309–335. doi: 10.1039/C7NP00054E
Shrestha, S., and Kim, Y. (2007). An entomopathogenic bacterium, Xenorhabdus nematophila, inhibits hemocyte phagocytosis of Spodoptera exigua by inhibiting phospholipase A2. J. Invertebr. Pathol. 96, 64–70. doi: 10.1016/j.jip.2007.02.009
Sicard, M., Ferdy, J.-B., Pagès, S., Le Brun, N., Godelle, B., Boemare, N., et al. (2004). When mutualists are pathogens: an experimental study of the symbioses between Steinernema (entomopathogenic nematodes) and Xenorhabdus (bacteria). J. Evol. Biol. 17, 985–993. doi: 10.1111/j.1420-9101.2004.00748.x
Singh, J., and Banerjee, N. (2008). Transcriptional analysis and functional characterization of a gene pair encoding iron-regulated xenocin and immunity proteins of Xenorhabdus nematophila. J. Bacteriol. 190, 3877–3885. doi: 10.1128/JB.00209-08
Stock, S. P., and Goodrich-Blair, H. (2008). Entomopathogenic nematodes and their bacterial symbionts: the inside out of a mutualistic association. Symbiosus 46, 65–75.
Sugar, D. R., Murfin, K. E., Chaston, J. M., Andersen, A. W., Richards, G. R., deLéon, L., et al. (2012). Phenotypic variation and host interactions of Xenorhabdus bovienii SS-2004, the entomopathogenic symbiont of Steinernema jollieti nematodes. Environ. Microbiol. 14, 924–939. doi: 10.1111/j.1462-2920.2011.02663.x
Sundar, L., and Chang, F. N. (1993). Antimicrobial activity and biosynthesis of indole antibiotics produced by Xenorhabdus nematophilus. J. Gen. Microbiol. 139, 3139–3148. doi: 10.1099/00221287-139-12-3139
Thaler, J. O., Baghdiguian, S., and Boemare, N. (1995). Purification and characterization of xenorhabdicin, a phage tail-like bacteriocin, from the lysogenic strain F1 of Xenorhabdus nematophilus. Appl. Environ. Microbiol. 61, 2049–2052.
Tobias, N. J., Wolff, H., Djahanschiri, B., Grundmann, F., Kronenwerth, M., Shi, Y. M., et al. (2017). Natural product diversity associated with the nematode symbionts Photorhabdus and Xenorhabdus. Nat. Microbiol. 2, 1676–1685. doi: 10.1038/s41564-017-0039-9
Wang, Y.-H., Feng, J.-T., Zhang, Q., and Zhang, X. (2008). Optimization of fermentation condition for antibiotic production by Xenorhabdus nematophila with response surface methodology. J. Appl. Microbiol. 104, 735–744. doi: 10.1111/j.1365-2672.2007.03599.x
Webster, J. M., Chen, G., Hu, K., and Li, J. (2002). “Bacterial metabolites,” in Entomopathogenic Nematology, ed. R. Gaugler (London: CAB International), 99–114. doi: 10.1079/9780851995670.0099
Webster, J. M., Li, J., and Chen, G. (1996). Xenorxides with antibacterial and antimycotic properties. Patent WO1996032396 A1. Geneva: World Intellectual Property Organization.
Xiao, Y., Meng, F., Qiu, D., and Yang, X. (2012). Two novel antimicrobial peptides purified from the symbiotic bacteria Xenorhabdus budapestensis NMC-10. Peptides 35, 253–260. doi: 10.1016/j.peptides.2012.03.027
Xiufen, Y., Huaiwen, Y., Heng, J., and Zheng, L. (2001). Effect of fermentation conditions on antibiotic production of Xenorhabdus nematophilus. Wei sheng wu xue tong bao 28, 12–16.
Yang, J., Zeng, H.-M., Lin, H.-F., Yang, X.-F., Liu, Z., Guo, L.-H., et al. (2012). An insecticidal protein from Xenorhabdus budapestensis that results in prophenoloxidase activation in the wax moth, Galleria mellonella. J. Invertebr. Pathol. 110, 60–67. doi: 10.1016/j.jip.2012.02.006
Zhanel, G. G., Trapp, S., Gin, A. S., DeCorby, M., Lagace-Wiens, P. R., Rubinstein, E., et al. (2008). Dalbavancin and telavancin: novel lipoglycopeptides for the treatment of gram-positive infections. Expert Rev. Anti Infect. Ther. 6, 67–81. doi: 10.1586/14787210.6.1.67
Keywords: Xenorhabdus, bioactive compounds, secondary metabolites, antimicrobial properties, antimicrobial peptides
Citation: Dreyer J, Malan AP and Dicks LMT (2018) Bacteria of the Genus Xenorhabdus, a Novel Source of Bioactive Compounds. Front. Microbiol. 9:3177. doi: 10.3389/fmicb.2018.03177
Received: 18 June 2018; Accepted: 07 December 2018;
Published: 19 December 2018.
Edited by:
Santi M. Mandal, Indian Institute of Technology Kharagpur, IndiaReviewed by:
Prabuddha Dey, Rutgers University – The State University of New Jersey, United StatesEkramul Islam, University of Kalyani, India
Copyright © 2018 Dreyer, Malan and Dicks. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Leon M. T. Dicks, LMTD@sun.ac.za