 Vanessa Brinkmann
Vanessa Brinkmann Niloofar Ale-Agha
Niloofar Ale-Agha Judith Haendeler
Judith Haendeler Natascia Ventura
Natascia Ventura- 1IUF – Leibniz Research Institute for Environmental Medicine, Düsseldorf, Germany
- 2Institute of Clinical Chemistry and Laboratory Diagnostic, Medical Faculty, Heinrich Heine University of Düsseldorf, Düsseldorf, Germany
Aging is the most important risk factor for the development of major life-threatening diseases such as cardiovascular disorders, cancer, and neurodegenerative disorders. The aging process is characterized by the accumulation of damage to intracellular macromolecules and it is concurrently shaped by genetic, environmental and nutritional factors. These factors influence the functionality of mitochondria, which play a central role in the aging process. Mitochondrial dysfunction is one of the hallmarks of aging and is associated with increased fluxes of ROS leading to damage of mitochondrial components, impaired metabolism of fatty acids, dysregulated glucose metabolism, and damage of adjacent organelles. Interestingly, many of the environmental (e.g., pollutants and other toxicants) and nutritional (e.g., flavonoids, carotenoids) factors influencing aging and mitochondrial function also directly or indirectly affect the activity of a highly conserved transcription factor, the Aryl hydrocarbon Receptor (AhR). Therefore, it is not surprising that many studies have already indicated a role of this versatile transcription factor in the aging process. We also recently found that the AhR promotes aging phenotypes across species. In this manuscript, we systematically review the existing literature on the contradictory studies indicating either pro- or anti-aging effects of the AhR and try to reconcile the seemingly conflicting data considering a possible dependency on the animal model, tissue, as well as level of AhR expression and activation. Moreover, given the crucial role of mitochondria in the aging process, we summarize the growing body of evidence pointing toward the influence of AhR on mitochondria, which can be of potential relevance for aging.
1. Aging Hallmarks and Associated Changes
Aging is defined as the time-dependent biological deterioration of structural, cellular and tissue components as well as physiological functions (e.g., stress resistance, immune system, ability to sense and move, decline of organ functionality). It is accompanied by an increased risk for the development of major life-threatening diseases such as cardiovascular disorders, cancer and neurodegenerative disorders (Lopez-Otin et al., 2013; Kaeberlein, 2016). The interest in aging research showed an increase in the past 60 years with a number of almost 7000 new PubMed entries in 2018. 60 years of research have advanced our knowledge on the molecular mechanisms of aging and a number of theories of aging have been proposed (Harman, 1956; Hamilton, 1966; Kirkwood, 1977; Villeponteau, 1997). These theories primarily fall into two categories (i) programmed and (ii) non-programmed based theories. While programmed theories of aging propose that mainly changes in the activity of specific genes, hormones, or the immune system are accountable for the aging process, non-programmed theories consider cellular damage resulting from the interaction with toxicants from the environment or by-products of metabolism the primary factor contributing to aging (Jin, 2010). It seems however most likely that the interaction between genetic and environmental factors shapes the aging process (Kenyon, 2010; Dato et al., 2017). Although aging is a very complex phenomenon and a variety of factors can affect different cells/tissue/organs and their interconnectivity at the same time, different evolutionarily conserved aging hallmarks have been described: (1) genomic instability, (2) telomere attrition, (3) epigenetic alterations, (4) loss of proteostasis, (5) deregulated nutrient sensing, (6) mitochondrial dysfunction, (7) cellular senescence, (8) stem cell exhaustion, and (9) altered intercellular communication (Lopez-Otin et al., 2013; Tigges et al., 2014). Indeed, apart from telomere attrition, all mentioned hallmarks can be observed in vertebrate as well as invertebrate model organisms (Kaeberlein, 2013). Mitochondria clearly play a central role in the aging process and it is interesting to note that while mitochondrial dysfunction itself is one of the hallmarks of aging, severe mitochondrial dysfunction can also promote most if not all other hallmarks of aging.
1.1 Mitochondria and Aging
Mitochondria are highly interconnected organelles and are composed of two specialized membranes, an intermembrane space and a matrix containing a circular DNA, which reminds us of their bacterial origin. Mitochondria play a central role in cell and organismal homeostasis and beside their major role in energy metabolism, they also control additional crucial cellular functions ranging from iron and calcium homeostasis to cell death and survival pathways. Given the central importance of mitochondria, cells developed a variety of protective mechanisms to cope with, prevent and repair their damage and alterations thus ensuring cells with the appropriate amount of functional mitochondria in physiological as well as stressful conditions. The “Mitochondrial Free Radical Theory of Aging,” MFRTA, which has taken central stage for several decades (Harman, 1956) in the aging field, states that during life reactive oxygen species (ROS) produced during mitochondria respiration gradually induce irreversible molecular and cellular damages with consequent functional decline ultimately playing a causal role in the aging process. It is therefore not surprising that failure of mitochondrial quality control pathways or severe, non-repairable mitochondrial damage, lead to a plethora of disorders and accelerate the aging process. More surprisingly, yet interestingly, is instead that the MFRTA theory has been recently questioned by the growing body of evidence showing that mild (as opposed to severe) increase in ROS and mild mitochondrial stress can actually promote healthy aging in an evolutionarily conserved manner (Ristow and Zarse, 2010; Ristow and Schmeisser, 2011; Munkacsy and Rea, 2014; Schiavi and Ventura, 2014). This provocative finding (in the field referred to as threshold effect or mitohormesis) has completely changed our classical view of the role of mitochondria in the aging process and stimulated the investigation of novel strategies to promote healthy aging. Taken MFRTA and mitohormesis together it is envisioned that mitochondria play a pivotal role in cell homeostasis and therefore in the aging process.
Dato et al. (2017) estimated that genetic factors only account for one quarter, while environmental and epigenetic factors account for three-quarters of age-associated changes. Considering the high influence of environmental factors on aging, we have recently investigated the role of a central environmental sensor, the highly conserved transcription factor Aryl-hydrocarbon Receptor (AhR) in the aging process (Eckers et al., 2016). Here, we will first review the conflicting evidence pointing to both pro- and anti-aging roles for AhR in aging. We will then try to reconcile these findings based on possible age-, tissue- or dose-dependent activation, and finally discuss pieces of evidence indicating a possible interaction between the AhR and mitochondria, which could be of relevance for the aging process.
2. AhR and Aging
The AhR was discovered in 1976 by Poland et al. (1976) as a dioxin-binding protein. The activity of this highly conserved transcription factor is historically dependent on the binding of ligands to its ligand binding domain (LBD). The functionality of this transcription factor is shaped by its functional domains: a basic helix-loop-helix domain (bHLH), two Per-ARNT-Sim (PAS) domains and a transcriptional activation domain (TAD). The N-terminal bHLH domain is involved in DNA binding, binding of heat shock protein 90 (Hsp90), and dimerization with AhR nuclear translocator (Arnt) (Ashida et al., 2008; Abel and Haarmann-Stemmann, 2010). The PAS A domain is required for binding to the Arnt, while the PAS B domain carries the LBD and thus is relevant for ligand binding but also interaction with the AhR-interacting protein [Aip (also XAP2)] and Hsp90. Carboxy-terminal of the AhR is a TAD (Ashida et al., 2008; Abel and Haarmann-Stemmann, 2010). In the absence of ligands, the AhR resides in the cytoplasm bound to Hsp90, Aip, and p23 (Ikuta et al., 1998; Ashida et al., 2008). These co-factors stabilize the ligand-affine state of the AhR and prevent its degradation (Ma and Whitlock, 1997; Meyer and Perdew, 1999). The functions of these co-factors are crucial and AhR is degraded in the absence of Aip or Hsp90 (Hwang et al., 2016). Binding of a ligand causes conformational changes resulting in the exposure of the nuclear localization signal (NLS) and the dissociation from Hsp90, Aip, and p23. In this state, AhR can shuttle to the nucleus, where it dimerizes with Arnt. The AhR-Arnt heterodimer then binds to the xenobiotic responsive elements (XREs) (core sequence 5′-GCGTG-3′) of AhR target genes. These target genes include phase-I detoxification genes like cytochrome P450 (cyp) monooxygenase genes (e.g., cyp1A1 or cyp1B1), phase-II detoxification genes like UDP glycosyltransferases (ugts) (e.g., ugt1A1 or ugt1A6), and glutathione S-transferases (gsts) (e.g., gstA1 or gstA2) (Yueh et al., 2003; Ashida et al., 2008; Xue et al., 2017). To avoid the constant activation of the AhR, negative feedback loops regulate the AhR cascade pathway (Mulero-Navarro and Fernandez-Salguero, 2016; Xue et al., 2017). It is interesting to note that different AhR target genes (e.g., enzymes involved in glutathione synthesis and modulation) as well as genes regulating or regulated by AhR (e.g., Sirt1, p53, Hif1, p300, and HSP90) are involved in the aging process (Henry and Gasiewicz, 1993; Marlowe et al., 2004; Koizumi et al., 2014; Li et al., 2014; Ming et al., 2015; Panchanathan et al., 2015; Ajami et al., 2017; Janssens et al., 2019; Sutter et al., 2019).
Various compounds influence the activity of the AhR, but not all of them are direct ligands. In fact, for some of the compounds modulating the activity of the AhR the direct mechanism is not known. Other compounds modulate AhR activity through an indirect mechanism. For this reason, in this review, we will refer to AhR modulators rather than ligands unless their direct binding to the LBD has been verified. A very well studied group of AhR modulators are xenobiotics. Particularly 2,3,7,8-tetrachlorodibenzo-p-dioxin (dioxin) is a planar molecule that has been shown to bind to the binding pocket of the LBD of mammalian AhR (Poland et al., 1976). Other modulators of the AhR are polyphenols, which can be found in a variety of different fruits and vegetables (Amakura et al., 2003), but whether they really bind to the AhR or affect its activity through other mechanisms is largely elusive (see Xue et al., 2017 for a detailed review). Epigallocatechin gallate, for example, binds to HSP90 and by doing so inhibits AhR signaling (Palermo et al., 2005). Curcumin instead can directly bind the AhR (Ciolino et al., 1998) and inhibits its downstream signaling through inhibition of AhR phosphorylation by the protein kinase C (PKC) (Nishiumi et al., 2007). An in silico analysis of the interactions between quercetin and the LBD of human AhR showed that quercetin can bind to specific residues in the AhR binding pocket (Jin et al., 2018). Interestingly, many of the polyphenols modulating AhR activity also affect mitochondria (Sandoval-Acuna et al., 2014). Besides these exogenous modulators, endogenous compounds affect AhR activity as well. These endogenous modulators are mainly, but not solely tryptophan derivatives, like the high-affinity ligand 6-formylindolo[3,2-b]carbazole (FICZ), a photoproduct of tryptophan, which is produced in response to UVB light (Rannug et al., 1987; Fritsche et al., 2007). Another endogenous but low-affinity AhR modulator is kynurenine (Opitz et al., 2011). More recently, compounds produced by the microbiota have been identified as AhR modulators. These are, similarly to the endogenous AhR modulators, mostly derivatives of tryptophan such as indole, indoxyl-3-sulfate, indole-3-propionic acid, indole-3-aldehyde, indole-3-acetate, and tryptamine (Jin et al., 2014; Rothhammer et al., 2016).
It is interesting to note that many of these AhR modulators may affect aging or age-associated diseases. Dioxin exposure, for example, can cause cancer, and cardiovascular diseases (Mandal, 2005; Marinkovic et al., 2010). Exposure to the xenobiotic and AhR ligand benzo[a]pyrene, which directly binds to the AhR (Okey et al., 1984) and causes its nuclear localization, shortens the lifespan in mice (Sakakibara et al., 2005), and promotes neurodegeneration as well as Alzheimer’s disease and Parkinson’s disease-like phenotypes in zebrafish (Gao et al., 2015). However, the actual involvement of the AhR in these processes was not investigated in these studies. Instead, in another study, benzo[a]pyrene was shown to cause cancer in an AhR-dependent manner (Shimizu et al., 2000) and similarly, the endogenous AhR modulator kynurenine promotes tumor formation through the AhR (Opitz et al., 2011). Moreover, another work showed that mice carrying a low-affinity AhR allele are more susceptible to benzo[a]pyrene-induced lethality than mice with a high-affinity AhR allele, suggesting the importance of the degree of AhR activation (Kerley-Hamilton et al., 2012). Many of the plant-derived dietary AhR modulators, on the other hand, have life- and health-extending effects across species. Curcumin, for example, shows protective effects on age-related neurodegenerative diseases in different species (Lim et al., 2001; Alavez et al., 2011; Caesar et al., 2012). Also, the AhR modulator quercetin extends lifespan in Caenorhabditis elegans (C. elegans) (Kampkotter et al., 2008; Pietsch et al., 2009), Drosophila melanogaster (Drosophila) (Proshkina et al., 2016) and mice (Xu et al., 2018). Interestingly, indole produced by commensal Escherichia coli was found to extend the lifespan of C. elegans, Drosophila and mice in an AhR-dependent manner (Sonowal et al., 2017) but in all other studies it has not been investigated whether the compounds mediate healthspan in an AhR dependent manner. While all of these studies focus on the effect of specific compounds on aging, there are only a few studies directly linking AhR and aging. In fact, a search on the MEDLINE/PubMed database with the Medical Subject Headings (MeSH) terms “ah receptor” and “aging” gave only 29 results. Here, we want to review the current state of research on the role of the AhR in aging in different model organisms as well as humans. We have however deliberately decided not to describe the large body of association studies correlating AhR activity/expression to age-associated diseases in human, which, although very interesting, would require a separate review.
2.1 AhR and Aging in Invertebrates
When studying aging, invertebrate model organisms offer some advantages over vertebrates: they are small, easy to cultivate, cheap in maintenance and most importantly have a short lifespan. These characteristics allow the performance of aging studies with a large number of individuals in a short time. Moreover, the conservation of the major aging pathways, as well as aging features, have made invertebrates like the nematode C. elegans, and the fruit fly Drosophila melanogaster elected model organisms of aging research (Kenyon, 2010; Kaeberlein, 2013). The homologs of the AhR are ahr-1 (Powell-Coffman et al., 1998) and spineless (Duncan et al., 1998) in C. elegans and Drosophila, respectively. AhR in C. elegans and Drosophila differ from mammalian AhR in their structure and their ligand binding ability. In fact, the classical AhR ligand dioxin does not bind AHR-1 (Powell-Coffman et al., 1998) and most likely neither Spineless (Duncan et al., 1998). Additionally, no direct binding of a ligand has ever been shown in C. elegans or Drosophila. In vitro studies showed that the Drosophila AhR is constitutively active which might result in the inability of ligand-dependent activation (Kudo et al., 2009). Recent studies show however that indoles from commensal bacteria extend the lifespan of C. elegans and Drosophila in an AhR-dependent manner (Sonowal et al., 2017). This might suggest that microbiota-derived small molecules could act as evolutionarily conserved AhR modulators. For a long time, the main focus of AhR research has been on the response to xenobiotics and the inability of invertebrate AhR to bind dioxin might be one of the reasons why few is known about the function of invertebrate AhRs. However, a conserved function of the AhR in the regulation of developmental processes has been shown in C. elegans and Drosophila: C. elegans ahr-1 mutants develop slightly slower than wild type (Aarnio et al., 2010) and have defects in neuronal development (Huang et al., 2004; Qin and Powell-Coffman, 2004; Qin et al., 2006; Smith et al., 2013). Similarly, Drosophila spineless mutants have defects in the development of antenna (Burgess and Duncan, 1990) and sensory neurons (Kim et al., 2006). Another conserved function of the AhR might be the regulation of fertility. C. elegans ahr-1 mutants have a slightly reduced egg number and an increased embryonic lethality (Aarnio et al., 2010). Although no such function is described in Drosophila, reproduction is affected by AhR in mice (Baba et al., 2008). These studies suggest that the physiological functions of the AhR might be conserved during evolution. Most importantly, AhR has as well an evolutionarily conserved role in the regulation of aging (Eckers et al., 2016). In contrast to the detrimental effects of loss of AhR function in early life, during aging a decreased AhR expression is beneficial: in a cross-species study, we showed that the AhR promotes aging phenotypes in human, mice and C. elegans (Eckers et al., 2016) and thus present an evolutionarily conserved role of the AhR in the aging process. More specifically, C. elegans carrying a loss of function allele of ahr-1 had a longer lifespan and an increase in physiological functions (e.g., motility and pharyngeal pumping) and stress resistance during aging (Eckers et al., 2016). Moreover, a higher spontaneous movement activity of C. elegans, D. melanogaster and humanized mice with reduced AhR expression or activity is reported (Williams et al., 2014). Although this is not a direct phenomenon in aging, a decreased movement can be considered a parameter for aging. A potential over-activation of the AhR during the aging process is further supported in C. elegans by the observation of increased ahr-1 mRNA expression during aging (Sonowal et al., 2017).
2.2 AhR and Aging in Mice
Currently, four different strains of mice with a complete AhR deficiency (AhR−/−) exist, which have been generated by different laboratories. In two of these strains, the coding part of exon 1 of the AhR gene was replaced with either a neomycin resistance cassette (Fernandez-Salguero et al., 1995) or the bacterial β-galactosidase gene fused to a NLS (Mimura et al., 1997) (thereby deleting the translation start codon as well as a stretch of basic amino acids that may play a role in DNA binding. The third line was generated by deletion of exon 2, which encodes the bHLH domain. Deletion of this exon leads to out of frame splicing from exon 1 into exon 3 and translation termination in codon 24, such that no functional AhR is produced (Schmidt et al., 1996). The AhR-deficiency does not result in lethality during in utero development, as the pups in all lines show a Mendelian distribution of the different genotypes (AhR+/+, AhR+/−, AhR−/−). Recently, a fourth AhR knockout mouse model (C57BL/6-AhrTM 1.2Arte) was created by Taconic1. These mice carry a deletion in exon 3, resulting in an out of frame splicing of exons 2 to exon 4. For simplicity, in this review the four different mice strains will be designated as AhRΔ1neo/Δ1neo (Fernandez-Salguero et al., 1995) AhRΔ1gal/Δ1gal (Mimura et al., 1997), AhRΔ2/Δ2 (Schmidt et al., 1996) and AhRΔ3/Δ3 (Taconic), respectively. These mouse strains show different phenotypes. On the one hand they exhibit common features like alterations in hepatic development, reproductive health, and retarded growth during the first 4 weeks compared to wild type mice. On the other hand, they show differences in immune system and reaction and susceptibility to infection, which is possibly due, at least in part, to differences in the genetic background (Fernandez-Salguero et al., 1995; Schmidt et al., 1996; Fernandez-Salguero et al., 1997; Lahvis et al., 2005; Baba et al., 2008; Esser, 2009; Butler et al., 2012).
Given that the AhR is heavily involved in detoxification it is not surprising that AhR deficiency has a profound effect on the hepatic system. All four AhR-deficient mice have reduced liver size, portal fibrosis, and a persistent intrahepatic porto-systemic shunt. Furthermore AhR−/− mice display an increased susceptibility to hepatocarcinogenesis and developed larger tumors (Moreno-Marin et al., 2017). In contrast, the AhRΔ1neo/Δ1neo improved the regenerative potential of the lung in response to the deleterious effects of acute toxin exposure (Morales-Hernandez et al., 2017).
AhR is not only involved in xenobiotic metabolism, but also in regulation of inflammation like macrophage M1/M2 polarization and cytokine secretion. It is discussed that AhR activation induces oxidative stress as a result of excessive generation of ROS. Recent studies indicate that AhR also affects several age-associated processes, such as vascular function or cellular senescence and age-associated macular degeneration (Hu et al., 2013; Singh et al., 2014; Eckers et al., 2016; Bravo-Ferrer et al., 2019). The recently generated AhRΔ3/Δ3 mice showed a premature aging phenotype resulting in a reduced life span. Those mice display functionality decline in several organs (Fernandez-Salguero et al., 1995; Bravo-Ferrer et al., 2019). In contrast, the AhRΔ2/Δ2 mice show a similar survival rate as wild type mice until 15 months of age and do not display a premature aging phenotype (Singh et al., 2014).
Atherosclerosis is assumed as an age-related, chronic inflammatory disease. Several studies have demonstrated that activation of AhR by dioxin or benzo[a]pyrene promotes atherosclerosis (Schmidt et al., 1996; Curfs et al., 2005; Wu et al., 2011). AhR overexpressing mice, which display a 10-fold higher affinity to benzo[a]pyrene, were crossed to apolipoprotein E deficient mice. Those mice display larger hearts under basal conditions. Moreover, mice showed increased numbers of atherosclerotic plaques in response to benzo[a]pyrene compared to a congenic mouse strain expressing an AhR with lower affinity (Kerley-Hamilton et al., 2012). We demonstrated the impact of AhR in the vascular system. By using the AhRΔ2/Δ2 mice, which display no phenotype in adulthood, we showed a decrease in vascular stiffness, which was accompanied by increased eNOS-activity and NO-bioavailability (Eckers et al., 2016). On the other hand, AhRΔ1neo/Δ1neo mice demonstrated cardiac hypertrophy, thickening of the arterial media and increased numbers of vascular smooth muscle cells in the arterial wall (Sauzeau et al., 2011).
In summary, the four AhR-deficient mice show different phenotypes with respect to aging (Table 1). However, since the AhR is needed during development and as a response to environmental, one has to consider that an early embryonic defect could result in a different outcome in adulthood. Therefore, it is maybe difficult to use mice, which are total AhR knockouts. One should rather use AhR conditional knockout mice to induce AhR deficiency in adulthood.
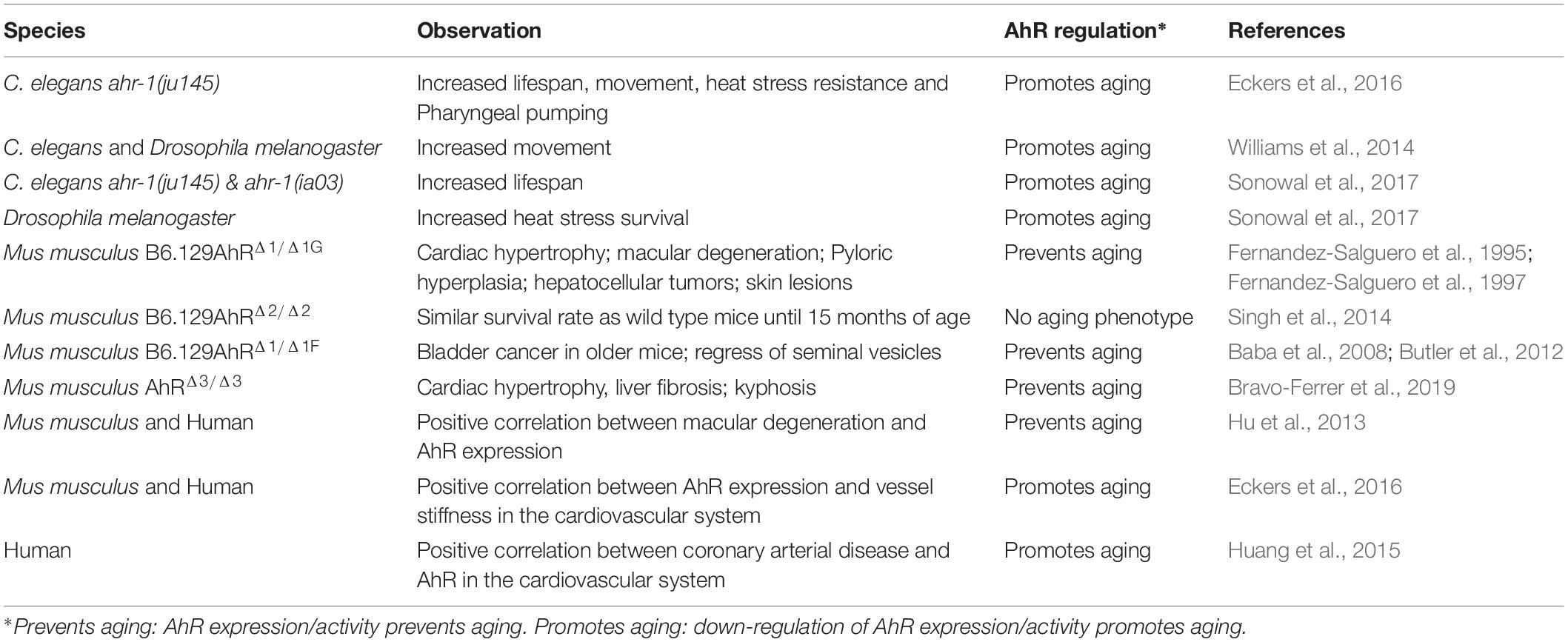
Table 1. Age-related phenotypes in different species and tissues.
2.3 AhR and Aging in Humans
Evidence for the role of AhR in aging in human subjects or human cell culture systems is rare. Most of these studies focus on the effect of certain AhR modulators on aging or associated parameters. A recent study showed for example that activation of the AhR by airborne polycyclic aromatic hydrocarbons induced cell aging and the expression of aging-related genes in human skin cells (Qiao et al., 2017). Interestingly, the expression of aging-related genes was inhibited by the presence of an AhR antagonist (Qiao et al., 2017). While studies using AhR ligands/modulators are very valuable for finding treatments for the prevention of pollution-induced aging or disease phenotypes, the direct effect of AhR in these processes is elusive since even high-affinity ligands like dioxin also affect cells in an AhR-independent manner (Hossain et al., 1998). There are only very few studies on the role of AhR on aging in the absence of modulators. A study from 2013 shows that AhR activity decreases during aging in human retinal pigment epithelial cells (Hu et al., 2013). Moreover, AhR protein levels were lower in cells from old donors compared to young donors, while AhR mRNA levels remained unaltered (Hu et al., 2013). They verified their findings in a mouse model and associated decreased activity of the AhR to age-related macular degeneration-like pathology (Hu et al., 2013). In 2016 we found that AhR expression is positively correlated to cardiovascular aging in humans (Eckers et al., 2016). Pulse wave velocity as the up to now best marker for vascular aging is increased with age and is positively correlated with AhR mRNA levels. Thus, increased AhR expression seems to be associated with old age in humans, thus we propose that AhR expression level is an indicator for vessel functionality (Eckers et al., 2016). Along the same lines, AhR expression has been linked to the incidence of coronary arterial disease in an epidemiological study on a Chinese population (Huang et al., 2015). They found increased AhR mRNA expression in coronary arterial disease patients compared to controls and suggested AhR as a diagnostic biomarker for coronary arterial disease (Huang et al., 2015).
In summary, the few studies on the role of AhR in human aging, similar to mice studies, display a complex role of the AhR. It has to be noted that one should clearly separate the effects of AhR in the development and in the aging process from invertebrates to vertebrates. However, it could be possible that the effect of the AhR on aging is tissue-dependent (Table 1) as well as environment dependent. Thus, more research in adult and aged invertebrates, vertebrates and humans is needed to understand the pathophysiological role of AhR in aging in different tissues, organs, as well as in the whole organism.
3. AhR-Mitochondria Crosstalk
Mitochondria play a central role in the aging process, are targeted by environmental pollutants and represent a central hub in nutrient metabolism. Interestingly, both environmental pollutants and dietary factors, such as polyphenols, can influence the transcriptional activity of the AhR. We, therefore, envisioned a possible crosstalk between AhR and mitochondria. Surprisingly, while mitochondria are extensively studied (more than 150,000 publications on PubMed) and, there are many studies investigating the influence of different AhR modulators on mitochondria, there are only 32 publications directly linking mitochondria to the AhR.
The effects of polyphenols on AhR and mitochondria are complex (reviewed in Sandoval-Acuna et al., 2014; Xue et al., 2017). On the one hand natural polyphenols are ROS scavengers and thereby influence mitochondria, which are targeted by ROS, but also mitochondrial down-stream signaling through the scavenging of mitochondrial ROS. This ROS scavenging function might be as well important for the influence of polyphenols on AhR activity. ROS can activate AhR through the conversion of tryptophan to FICZ (Smirnova et al., 2016) and thus the ROS-scavenging properties of polyphenols might prevent this activation and connect AhR with mitochondria. On the other hand, additional ROS-scavenging independent functions of polyphenols are reported on mitochondria like regulation of mitochondrial biogenesis, mitochondrial membrane potential, and mitochondrial electron transport chain activity (Sandoval-Acuna et al., 2014). Given the central role of Sirt1 in the aging process, a very interesting ROS-scavenging independent mode of action of polyphenols on mitochondria is the induction of mitochondrial biogenesis through Sirt1. Several polyphenols indeed were shown to activate Sirt1 (reviewed in Ajami et al., 2017). Considering the interaction between AhR and Sirt1 (Koizumi et al., 2014; Ming et al., 2015; Sutter et al., 2019), the notion that ROS activates AhR (Smirnova et al., 2016), and the multiple roles polyphenols may exert on mitochondria, these findings provide indirect evidence for a possible crosstalk between AhR, Sirt1 and mitochondria of relevance for the aging process.
The first direct evidence of a link between mitochondria and the AhR was published in 2002 (Senft et al., 2002). In this study, Senft and co-workers investigated the role of AhR signaling in the increase of mitochondrial ROS upon dioxin treatment in the liver of mice. Specifically, they found that dioxin treatment induced mitochondrial ROS in wild type but not in AhR−/− mice (Senft et al., 2002). Interestingly, they noticed that the basal mitochondrial ROS levels were lower in AhR−/− mice (Senft et al., 2002), which might suggest that AhR has an impact on the mitochondria not only in the presence of ligands but also under normal conditions. Dioxin exposure was also shown to decrease mitochondrial membrane potential in spermatozoa of mice in an AhR-dependent manner (Fisher et al., 2005). Similarly, embryonic stem cells and beating cardiomyocytes from AhR−/− mice are protected against the dioxin-induced increase in markers of mitochondrial stress and of mtDNA damage (Wang et al., 2016). Together, these studies suggest that AhR mediates mitochondrial dysfunction in response to dioxin. Moreover, benzo[a]pyrene was shown to increase mitochondrial dysfunction and decrease the mitochondrial membrane potential, resulting in the depletion of ATP levels along with inhibition of the oxygen consumption rate in the human keratinocyte cell line (HACAT). In this study, it was shown that the removal of damaged mitochondria by mitophagy is reduced in AhR and CYP1B1 (an AhR target gene) knockdown but a direct link between AhR and mitophagy was not established (Das et al., 2017). In another study genetic ablation of the AhR resulted in reduced expression of Superoxide Dismutases (SODs) in fibroblasts. Thus, those fibroblasts are more sensitive to cigarette smoke resulting in increased cell death and reduced proliferation, which is accompanied by decreased mitochondrial membrane potential (Rico De Souza et al., 2011). The detrimental effect of loss of AhR function is further supported by studies in embryonic hearts of mice, where the disruption of AhR signaling leads to mitochondrial dysfunction (Carreira et al., 2015b). Similarly, female, but not male mice exposed to dioxin as embryos showed altered expression in genes of the canonical mitochondrial pathway and a higher number of mitochondria in the heart (Carreira et al., 2015a). In adult AhR−/− mice those changes were not observed (Carreira et al., 2015a). In both studies the effect of dioxin treatment was not investigated in AhR−/− mice.
AhR does not only influence mitochondrial function but two studies have recently suggested that the AhR is also localized within the mitochondria. Tappenden et al. (2011) were the first to identify an interaction between the AhR and the ATP5α1 subunit of the ATP synthase complex in different cell lines. Further analysis of the exact localization of the mitochondrial AhR in murine hepatoma cells showed that it localizes inside the intermembrane space (Hwang et al., 2016). Interestingly, when treated with dioxin, mitochondrial localization of the AhR and interaction with ATP5α1 were lost (Tappenden et al., 2011; Hwang et al., 2016), suggesting that AhR only localizes inside the mitochondria in the absence of ligands. Considering that AhR is bound by AIP in the absence of ligands and that AIP has been found to interact with the Mitochondrial import receptor subunit TOMM20 and to mediate preprotein transport in mitochondria (Yano et al., 2003), AIP could be the critical mediator of AhR localization into mitochondria. Indeed, siRNA against TOMM20 reduces mitochondrial AhR, but not cytoplasmatic or nuclear AhR by 70% (Hwang et al., 2016). Thus, Hwang et al. proposed that AIP and HSP90 contribute to the mitochondrial localization of AhR by interacting with TOMM20, which imports AhR into the intermembrane space (Hwang et al., 2016).
Taken together, these studies strongly suggest that the effects of AhR on mitochondrial function are likely tissue-, age-, and maybe even sex-dependent (Table 2). Additionally, the presence of dioxin seems to have a strong impact on the outcome of the study. Nonetheless, these are mainly in vitro studies and causal-effect, as well as mechanistic studies in primary cells and model organisms in more physiological conditions, are required to clearly establish a possible crosstalk between AhR and mitochondria. Interestingly, in C. elegans, animals with reduced mitochondrial function and ahr-1 mutants share some phenotypic features like slower larval development, alterations in fat metabolism and sensory neurons and most importantly lifespan extension (Rea et al., 2007; Aarnio et al., 2010; Schiavi et al., 2013; Smith et al., 2013; Maglioni et al., 2014; Eckers et al., 2016). Moreover, long-lived mitochondrial mutants have increased levels of cyps, ugts, and gsts (Cristina et al., 2009; Liu et al., 2014; Mao et al., 2019), which, at least in mammals are known target genes of the AhR. Investigating the potential AhR-mitochondrial crosstalk in appropriate in vivo model systems will certainly help revealing its potential causal role in different pathophysiological contexts including aging and associated pathologies.

Table 2. Tissue- and age-specific effects of AhR expression on mitochondrial function in mice.
Author Contributions
NV and JH contributed to the conception of the review. VB and NA-A wrote the sections of the manuscript. All authors contributed to the manuscript revision, editing read, and approved the submitted version.
Funding
This work was supported by funding from the Deutsche Forschungsgemeinschaft to NV (DFG grant VE663/8-1) and to JH (SFB 1116, Project A04).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to acknowledge Dr. Joachim Altschmied for critical reading of the manuscript. We would like to apologize for all the papers from estimate scientists and colleagues, which we could not cite.
Footnotes
References
Aarnio, V., Storvik, M., Lehtonen, M., Asikainen, S., Reisner, K., Callaway, J., et al. (2010). Fatty acid composition and gene expression profiles are altered in aryl hydrocarbon receptor-1 mutant Caenorhabditis elegans. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 151, 318–324. doi: 10.1016/j.cbpc.2009.12.006
Abel, J., and Haarmann-Stemmann, T. (2010). An introduction to the molecular basics of aryl hydrocarbon receptor biology. Biol. Chem. 391, 1235–1248. doi: 10.1515/BC.2010.128
Ajami, M., Pazoki-Toroudi, H., Amani, H., Nabavi, S. F., Braidy, N., Vacca, R. A., et al. (2017). Therapeutic role of sirtuins in neurodegenerative disease and their modulation by polyphenols. Neurosci. Biobehav. Rev. 73, 39–47. doi: 10.1016/j.neubiorev.2016.11.022
Alavez, S., Vantipalli, M. C., Zucker, D. J., Klang, I. M., and Lithgow, G. J. (2011). Amyloid-binding compounds maintain protein homeostasis during ageing and extend lifespan. Nature 472, 226–229. doi: 10.1038/nature09873
Amakura, Y., Tsutsumi, T., Sasaki, K., Yoshida, T., and Maitani, T. (2003). Screening of the inhibitory effect of vegetable constituents on the aryl hydrocarbon receptor-mediated activity induced by 2,3,7,8-tetrachlorodibenzo-p-dioxin. Biol. Pharm. Bull. 26, 1754–1760. doi: 10.1248/bpb.26.1754
Ashida, H., Nishiumi, S., and Fukuda, I. (2008). An update on the dietary ligands of the AhR. Expert Opin. Drug Metab. Toxicol. 4, 1429–1447. doi: 10.1517/17425255.4.11.1429
Baba, T., Shima, Y., Owaki, A., Mimura, J., Oshima, M., Fujii-Kuriyama, Y., et al. (2008). Disruption of aryl hydrocarbon receptor (AhR) induces regression of the seminal vesicle in aged male mice. Sex Dev. 2, 1–11. doi: 10.1159/000117714
Bravo-Ferrer, I., Cuartero, M. I., Medina, V., Ahedo-Quero, D., Pena-Martinez, C., Perez-Ruiz, A., et al. (2019). Lack of the aryl hydrocarbon receptor accelerates aging in mice. FASEB J. 33, 12644–12654. doi: 10.1096/fj.201901333R
Burgess, E. A., and Duncan, I. (1990). Direct control of antennal identity by the spineless-aristapedia gene of Drosophila. Mol. Gen. Genet. 221, 347–357.
Butler, R., Inzunza, J., Suzuki, H., Fujii-Kuriyama, Y., Warner, M., and Gustafsson, J. A. (2012). Uric acid stones in the urinary bladder of aryl hydrocarbon receptor (AhR) knockout mice. Proc. Natl. Acad. Sci. U.S.A. 109, 1122–1126. doi: 10.1073/pnas.1120581109
Caesar, I., Jonson, M., Nilsson, K. P., Thor, S., and Hammarstrom, P. (2012). Curcumin promotes A-beta fibrillation and reduces neurotoxicity in transgenic Drosophila. PLoS One 7:e31424. doi: 10.1371/journal.pone.0031424
Carreira, V. S., Fan, Y., Kurita, H., Wang, Q., Ko, C. I., Naticchioni, M., et al. (2015a). Disruption of Ah receptor signaling during mouse development leads to abnormal cardiac structure and function in the adult. PLoS One 10:e0142440. doi: 10.1371/journal.pone.0142440
Carreira, V. S., Fan, Y., Wang, Q., Zhang, X., Kurita, H., Ko, C. I., et al. (2015b). Ah receptor signaling controls the expression of cardiac development and homeostasis genes. Toxicol. Sci. 147, 425–435. doi: 10.1093/toxsci/kfv138
Ciolino, H. P., Daschner, P. J., Wang, T. T., and Yeh, G. C. (1998). Effect of curcumin on the aryl hydrocarbon receptor and cytochrome P450 1A1 in MCF-7 human breast carcinoma cells. Biochem. Pharmacol. 56, 197–206. doi: 10.1016/s0006-2952(98)00143-9
Cristina, D., Cary, M., Lunceford, A., Clarke, C., and Kenyon, C. (2009). A regulated response to impaired respiration slows behavioral rates and increases lifespan in Caenorhabditis elegans. PLoS Genet. 5:e1000450. doi: 10.1371/journal.pgen.1000450
Curfs, D. M., Knaapen, A. M., Pachen, D. M., Gijbels, M. J., Lutgens, E., Smook, M. L., et al. (2005). Polycyclic aromatic hydrocarbons induce an inflammatory atherosclerotic plaque phenotype irrespective of their DNA binding properties. FASEB J. 19, 1290–1292. doi: 10.1096/fj.04-2269fje
Das, D. N., Naik, P. P., Mukhopadhyay, S., Panda, P. K., Sinha, N., Meher, B. R., et al. (2017). Elimination of dysfunctional mitochondria through mitophagy suppresses benzo[a]pyrene-induced apoptosis. Free Radic. Biol. Med. 112, 452–463. doi: 10.1016/j.freeradbiomed.2017.08.020
Dato, S., Rose, G., Crocco, P., Monti, D., Garagnani, P., Franceschi, C., et al. (2017). The genetics of human longevity: an intricacy of genes, environment, culture and microbiome. Mech. Ageing Dev. 165, 147–155. doi: 10.1016/j.mad.2017.03.011
Duncan, D. M., Burgess, E. A., and Duncan, I. (1998). Control of distal antennal identity and tarsal development in Drosophila by spineless-aristapedia, a homolog of the mammalian dioxin receptor. Genes Dev. 12, 1290–1303. doi: 10.1101/gad.12.9.1290
Eckers, A., Jakob, S., Heiss, C., Haarmann-Stemmann, T., Goy, C., Brinkmann, V., et al. (2016). The aryl hydrocarbon receptor promotes aging phenotypes across species. Sci. Rep. 6:19618. doi: 10.1038/srep19618
Esser, C. (2009). The immune phenotype of AhR null mouse mutants: not a simple mirror of xenobiotic receptor over-activation. Biochem. Pharmacol. 77, 597–607. doi: 10.1016/j.bcp.2008.10.002
Fernandez-Salguero, P., Pineau, T., Hilbert, D. M., Mcphail, T., Lee, S. S., Kimura, S., et al. (1995). Immune system impairment and hepatic fibrosis in mice lacking the dioxin-binding Ah receptor. Science 268, 722–726. doi: 10.1126/science.7732381
Fernandez-Salguero, P. M., Ward, J. M., Sundberg, J. P., and Gonzalez, F. J. (1997). Lesions of aryl-hydrocarbon receptor-deficient mice. Vet. Pathol. 34, 605–614. doi: 10.1177/030098589703400609
Fisher, M. T., Nagarkatti, M., and Nagarkatti, P. S. (2005). Aryl hydrocarbon receptor-dependent induction of loss of mitochondrial membrane potential in epididydimal spermatozoa by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Toxicol. Lett. 157, 99–107. doi: 10.1016/j.toxlet.2005.01.008
Fritsche, E., Schafer, C., Calles, C., Bernsmann, T., Bernshausen, T., Wurm, M., et al. (2007). Lightening up the UV response by identification of the arylhydrocarbon receptor as a cytoplasmatic target for ultraviolet B radiation. Proc. Natl. Acad. Sci. U.S.A. 104, 8851–8856. doi: 10.1073/pnas.0701764104
Gao, D., Wu, M., Wang, C., Wang, Y., and Zuo, Z. (2015). Chronic exposure to low benzo[a]pyrene level causes neurodegenerative disease-like syndromes in Zebrafish (Danio rerio). Aquat. Toxicol. 167, 200–208. doi: 10.1016/j.aquatox.2015.08.013
Hamilton, W. D. (1966). The moulding of senescence by natural selection. J. Theor. Biol. 12, 12–45. doi: 10.1016/0022-5193(66)90184-6
Harman, D. (1956). Aging: a theory based on free radical and radiation chemistry. J. Gerontol. 11, 298–300. doi: 10.1093/geronj/11.3.298
Henry, E. C., and Gasiewicz, T. A. (1993). Transformation of the aryl hydrocarbon receptor to a DNA-binding form is accompanied by release of the 90 kDa heat-shock protein and increased affinity for 2,3,7,8-tetrachlorodibenzo-p-dioxin. Biochem. J. 294(Pt 1), 95–101. doi: 10.1042/bj2940095
Hossain, A., Tsuchiya, S., Minegishi, M., Osada, M., Ikawa, S., Tezuka, F. A., et al. (1998). The Ah receptor is not involved in 2,3,7,8-tetrachlorodibenzo- p-dioxin-mediated apoptosis in human leukemic T cell lines. J. Biol. Chem. 273, 19853–19858. doi: 10.1074/jbc.273.31.19853
Hu, P., Herrmann, R., Bednar, A., Saloupis, P., Dwyer, M. A., Yang, P., et al. (2013). Aryl hydrocarbon receptor deficiency causes dysregulated cellular matrix metabolism and age-related macular degeneration-like pathology. Proc. Natl. Acad. Sci. U.S.A. 110, E4069–E4078. doi: 10.1073/pnas.1307574110
Huang, S., Shui, X., He, Y., Xue, Y., Li, J., Li, G., et al. (2015). AhR expression and polymorphisms are associated with risk of coronary arterial disease in chinese population. Sci. Rep. 5:8022. doi: 10.1038/srep08022
Huang, X., Powell-Coffman, J. A., and Jin, Y. (2004). The AHR-1 aryl hydrocarbon receptor and its co-factor the AHA-1 aryl hydrocarbon receptor nuclear translocator specify GABAergic neuron cell fate in C. elegans. Development 131, 819–828. doi: 10.1242/dev.00959
Hwang, H. J., Dornbos, P., Steidemann, M., Dunivin, T. K., Rizzo, M., and Lapres, J. J. (2016). Mitochondrial-targeted aryl hydrocarbon receptor and the impact of 2,3,7,8-tetrachlorodibenzo-p-dioxin on cellular respiration and the mitochondrial proteome. Toxicol. Appl. Pharmacol. 304, 121–132. doi: 10.1016/j.taap.2016.04.005
Ikuta, T., Eguchi, H., Tachibana, T., Yoneda, Y., and Kawajiri, K. (1998). Nuclear localization and export signals of the human aryl hydrocarbon receptor. J. Biol. Chem. 273, 2895–2904. doi: 10.1074/jbc.273.5.2895
Janssens, G. E., Lin, X. X., Millan-Arino, L., Kavsek, A., Sen, I., Seinstra, R. I., et al. (2019). Transcriptomics-based screening identifies pharmacological inhibition of Hsp90 as a means to defer aging. Cell Rep. 27, 467.e6–480.e6. doi: 10.1016/j.celrep.2019.03.044
Jin, U. H., Lee, S. O., Sridharan, G., Lee, K., Davidson, L. A., Jayaraman, A., et al. (2014). Microbiome-derived tryptophan metabolites and their aryl hydrocarbon receptor-dependent agonist and antagonist activities. Mol. Pharmacol. 85, 777–788. doi: 10.1124/mol.113.091165
Jin, U. H., Park, H., Li, X., Davidson, L. A., Allred, C., Patil, B., et al. (2018). Structure-dependent modulation of aryl hydrocarbon receptor-mediated activities by flavonoids. Toxicol. Sci. 164, 205–217. doi: 10.1093/toxsci/kfy075
Kaeberlein, M. (2016). The biology of aging: citizen scientists and their pets as a bridge between research on model organisms and human subjects. Vet. Pathol. 53, 291–298. doi: 10.1177/0300985815591082
Kampkotter, A., Timpel, C., Zurawski, R. F., Ruhl, S., Chovolou, Y., Proksch, P., et al. (2008). Increase of stress resistance and lifespan of Caenorhabditis elegans by quercetin. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 149, 314–323. doi: 10.1016/j.cbpb.2007.10.004
Kerley-Hamilton, J. S., Trask, H. W., Ridley, C. J., Dufour, E., Lesseur, C., Ringelberg, C. S., et al. (2012). Inherent and benzo[a]pyrene-induced differential aryl hydrocarbon receptor signaling greatly affects life span, atherosclerosis, cardiac gene expression, and body and heart growth in mice. Toxicol. Sci. 126, 391–404. doi: 10.1093/toxsci/kfs002
Kim, M. D., Jan, L. Y., and Jan, Y. N. (2006). The bHLH-PAS protein Spineless is necessary for the diversification of dendrite morphology of Drosophila dendritic arborization neurons. Genes Dev. 20, 2806–2819. doi: 10.1101/gad.1459706
Koizumi, M., Tatebe, J., Watanabe, I., Yamazaki, J., Ikeda, T., and Morita, T. (2014). Aryl hydrocarbon receptor mediates indoxyl sulfate-induced cellular senescence in human umbilical vein endothelial cells. J. Atheroscler. Thromb. 21, 904–916. doi: 10.5551/jat.23663
Kudo, K., Takeuchi, T., Murakami, Y., Ebina, M., and Kikuchi, H. (2009). Characterization of the region of the aryl hydrocarbon receptor required for ligand dependency of transactivation using chimeric receptor between Drosophila and Mus musculus. Biochim. Biophys. Acta 1789, 477–486. doi: 10.1016/j.bbagrm.2009.06.003
Lahvis, G. P., Pyzalski, R. W., Glover, E., Pitot, H. C., Mcelwee, M. K., and Bradfield, C. A. (2005). The aryl hydrocarbon receptor is required for developmental closure of the ductus venosus in the neonatal mouse. Mol. Pharmacol. 67, 714–720. doi: 10.1124/mol.104.008888
Li, Z. D., Wang, K., Yang, X. W., Zhuang, Z. G., Wang, J. J., and Tong, X. W. (2014). Expression of aryl hydrocarbon receptor in relation to p53 status and clinicopathological parameters in breast cancer. Int. J. Clin. Exp. Pathol. 7, 7931–7937.
Lim, G. P., Chu, T., Yang, F., Beech, W., Frautschy, S. A., and Cole, G. M. (2001). The curry spice curcumin reduces oxidative damage and amyloid pathology in an Alzheimer transgenic mouse. J. Neurosci. 21, 8370–8377. doi: 10.1523/jneurosci.21-21-08370.2001
Liu, Y., Samuel, B. S., Breen, P. C., and Ruvkun, G. (2014). Caenorhabditis elegans pathways that surveil and defend mitochondria. Nature 508, 406–410. doi: 10.1038/nature13204
Lopez-Otin, C., Blasco, M. A., Partridge, L., Serrano, M., and Kroemer, G. (2013). The hallmarks of aging. Cell 153, 1194–1217. doi: 10.1016/j.cell.2013.05.039
Ma, Q., and Whitlock, J. P. (1997). A novel cytoplasmic protein that interacts with the Ah receptor, contains tetratricopeptide repeat motifs, and augments the transcriptional response to 2,3,7,8-tetrachlorodibenzo-p-dioxin. J. Biol. Chem. 272, 8878–8884. doi: 10.1074/jbc.272.14.8878
Maglioni, S., Schiavi, A., Runci, A., Shaik, A., and Ventura, N. (2014). Mitochondrial stress extends lifespan in C. elegans through neuronal hormesis. Exp. Gerontol. 56, 89–98. doi: 10.1016/j.exger.2014.03.026
Mandal, P. K. (2005). Dioxin: a review of its environmental effects and its aryl hydrocarbon receptor biology. J. Comp. Physiol. B 175, 221–230. doi: 10.1007/s00360-005-0483-3
Mao, K., Ji, F., Breen, P., Sewell, A., Han, M., Sadreyev, R., et al. (2019). Mitochondrial dysfunction in C. elegans activates mitochondrial relocalization and nuclear hormone receptor-dependent detoxification genes. Cell Metab. 29, 1182.e4–1191.e4. doi: 10.1016/j.cmet.2019.01.022
Marinkovic, N., Pasalic, D., Ferencak, G., Grskovic, B., and Stavljenic Rukavina, A. (2010). Dioxins and human toxicity. Arh. Hig. Rada. Toksikol. 61, 445–453. doi: 10.2478/10004-1254-61-2010-2024
Marlowe, J. L., Knudsen, E. S., Schwemberger, S., and Puga, A. (2004). The aryl hydrocarbon receptor displaces p300 from E2F-dependent promoters and represses S phase-specific gene expression. J. Biol. Chem. 279, 29013–29022. doi: 10.1074/jbc.m404315200
Meyer, B. K., and Perdew, G. H. (1999). Characterization of the AhR-hsp90-XAP2 core complex and the role of theimmunophilin-related protein XAP2 in AhR stabilization. Biochemistry 38, 8907–8917. doi: 10.1021/bi982223w
Mimura, J., Yamashita, K., Nakamura, K., Morita, M., Takagi, T. N., Nakao, K., et al. (1997). Loss of teratogenic response to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in mice lacking the Ah (dioxin) receptor. Genes Cells 2, 645–654. doi: 10.1046/j.1365-2443.1997.1490345.x
Ming, M., Zhao, B., Shea, C. R., Shah, P., Qiang, L., White, S. R., et al. (2015). Loss of sirtuin 1 (SIRT1) disrupts skin barrier integrity and sensitizes mice to epicutaneous allergen challenge. J. Allergy Clin. Immunol. 135, 936.e4–945.e4. doi: 10.1016/j.jaci.2014.09.035
Morales-Hernandez, A., Nacarino-Palma, A., Moreno-Marin, N., Barrasa, E., Paniagua-Quinones, B., Catalina-Fernandez, I., et al. (2017). Lung regeneration after toxic injury is improved in absence of dioxin receptor. Stem Cell Res. 25, 61–71. doi: 10.1016/j.scr.2017.10.009
Moreno-Marin, N., Barrasa, E., Morales-Hernandez, A., Paniagua, B., Blanco-Fernandez, G., Merino, J. M., et al. (2017). Dioxin receptor adjusts liver regeneration after acute toxic injury and protects against liver carcinogenesis. Sci. Rep. 7:10420. doi: 10.1038/s41598-017-10984-w
Mulero-Navarro, S., and Fernandez-Salguero, P. M. (2016). New trends in aryl hydrocarbon receptor biology. Front. Cell Dev. Biol. 4:45. doi: 10.3389/fcell.2016.00045
Munkacsy, E., and Rea, S. L. (2014). The paradox of mitochondrial dysfunction and extended longevity. Exp. Gerontol. 56, 221–233. doi: 10.1016/j.exger.2014.03.016
Nishiumi, S., Yoshida, K., and Ashida, H. (2007). Curcumin suppresses the transformation of an aryl hydrocarbon receptor through its phosphorylation. Arch. Biochem. Biophys. 466, 267–273. doi: 10.1016/j.abb.2007.08.007
Okey, A. B., Dube, A. W., and Vella, L. M. (1984). Binding of benzo(a)pyrene and dibenz(a,h)anthracene to the Ah receptor in mouse and rat hepatic cytosols. Cancer Res. 44, 1426–1432.
Opitz, C. A., Litzenburger, U. M., Sahm, F., Ott, M., Tritschler, I., Trump, S., et al. (2011). An endogenous tumour-promoting ligand of the human aryl hydrocarbon receptor. Nature 478, 197–203. doi: 10.1038/nature10491
Palermo, C. M., Westlake, C. A., and Gasiewicz, T. A. (2005). Epigallocatechin gallate inhibits aryl hydrocarbon receptor gene transcription through an indirect mechanism involving binding to a 90 kDa heat shock protein. Biochemistry 44, 5041–5052. doi: 10.1021/bi047433p
Panchanathan, R., Liu, H., and Choubey, D. (2015). Activation of p53 in human and murine cells by DNA-damaging agents differentially regulates aryl hydrocarbon receptor levels. Int. J. Toxicol. 34, 242–249. doi: 10.1177/1091581815578013
Pietsch, K., Saul, N., Menzel, R., Sturzenbaum, S. R., and Steinberg, C. E. (2009). Quercetin mediated lifespan extension in Caenorhabditis elegans is modulated by age-1, daf-2, sek-1 and unc-43. Biogerontology 10, 565–578. doi: 10.1007/s10522-008-9199-6
Poland, A., Glover, E., and Kende, A. S. (1976). Stereospecific, high affinity binding of 2,3,7,8-tetrachlorodibenzo-p-dioxin by hepatic cytosol. Evidence that the binding species is receptor for induction of aryl hydrocarbon hydroxylase. J. Biol. Chem. 251, 4936–4946.
Powell-Coffman, J. A., Bradfield, C. A., and Wood, W. B. (1998). Caenorhabditis elegans orthologs of the aryl hydrocarbon receptor and its heterodimerization partner the aryl hydrocarbon receptor nuclear translocator. Proc. Natl. Acad. Sci. U.S.A. 95, 2844–2849. doi: 10.1073/pnas.95.6.2844
Proshkina, E., Lashmanova, E., Dobrovolskaya, E., Zemskaya, N., Kudryavtseva, A., Shaposhnikov, M., et al. (2016). Geroprotective and radioprotective activity of quercetin, (-)-epicatechin, and ibuprofen in Drosophila melanogaster. Front. Pharmacol. 7:505. doi: 10.3389/fphar.2016.00505
Qiao, Y., Li, Q., Du, H. Y., Wang, Q. W., Huang, Y., and Liu, W. (2017). Airborne polycyclic aromatic hydrocarbons trigger human skin cells aging through aryl hydrocarbon receptor. Biochem. Biophys. Res. Commun. 488, 445–452. doi: 10.1016/j.bbrc.2017.04.160
Qin, H., and Powell-Coffman, J. A. (2004). The Caenorhabditis elegans aryl hydrocarbon receptor, AHR-1, regulates neuronal development. Dev. Biol. 270, 64–75. doi: 10.1016/s0012-1606(04)00119-8
Qin, H., Zhai, Z., and Powell-Coffman, J. A. (2006). The Caenorhabditis elegans AHR-1 transcription complex controls expression of soluble guanylate cyclase genes in the URX neurons and regulates aggregation behavior. Dev. Biol. 298, 606–615. doi: 10.1016/j.ydbio.2006.07.017
Rannug, A., Rannug, U., Rosenkranz, H. S., Winqvist, L., Westerholm, R., Agurell, E., et al. (1987). Certain photooxidized derivatives of tryptophan bind with very high affinity to the Ah receptor and are likely to be endogenous signal substances. J. Biol. Chem. 262, 15422–15427.
Rea, S. L., Ventura, N., and Johnson, T. E. (2007). Relationship between mitochondrial electron transport chain dysfunction, development, and life extension in Caenorhabditis elegans. PLoS Biol. 5:e259. doi: 10.1371/journal.pbio.0050259
Rico De Souza, A., Zago, M., Pollock, S. J., Sime, P. J., Phipps, R. P., and Baglole, C. J. (2011). Genetic ablation of the aryl hydrocarbon receptor causes cigarette smoke-induced mitochondrial dysfunction and apoptosis. J. Biol. Chem. 286, 43214–43228. doi: 10.1074/jbc.M111.258764
Ristow, M., and Schmeisser, S. (2011). Extending life span by increasing oxidative stress. Free Radic. Biol. Med. 51, 327–336. doi: 10.1016/j.freeradbiomed.2011.05.010
Ristow, M., and Zarse, K. (2010). How increased oxidative stress promotes longevity and metabolic health: the concept of mitochondrial hormesis (mitohormesis). Exp. Gerontol. 45, 410–418. doi: 10.1016/j.exger.2010.03.014
Rothhammer, V., Mascanfroni, I. D., Bunse, L., Takenaka, M. C., Kenison, J. E., Mayo, L., et al. (2016). Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and central nervous system inflammation via the aryl hydrocarbon receptor. Nat. Med. 22, 586–597. doi: 10.1038/nm.4106
Sakakibara, H., Nakagawa, S., Wakameda, H., Nakagiri, Y., Kamata, K., Das, S. K., et al. (2005). Effects of Japanese kelp (kombu) on life span of benzo[a]pyrene-fed mice. J. Nutr. Sci. Vitaminol. 51, 369–373. doi: 10.3177/jnsv.51.369
Sandoval-Acuna, C., Ferreira, J., and Speisky, H. (2014). Polyphenols and mitochondria: an update on their increasingly emerging ROS-scavenging independent actions. Arch. Biochem. Biophys. 559, 75–90. doi: 10.1016/j.abb.2014.05.017
Sauzeau, V., Carvajal-Gonzalez, J. M., Riolobos, A. S., Sevilla, M. A., Menacho-Marquez, M., Roman, A. C., et al. (2011). Transcriptional factor aryl hydrocarbon receptor (Ahr) controls cardiovascular and respiratory functions by regulating the expression of the Vav3 proto-oncogene. J. Biol. Chem. 286, 2896–2909. doi: 10.1074/jbc.M110.187534
Schiavi, A., Torgovnick, A., Kell, A., Megalou, E., Castelein, N., Guccini, I., et al. (2013). Autophagy induction extends lifespan and reduces lipid content in response to frataxin silencing in C. elegans. Exp. Gerontol. 48, 191–201. doi: 10.1016/j.exger.2012.12.002
Schiavi, A., and Ventura, N. (2014). The interplay between mitochondria and autophagy and its role in the aging process. Exp. Gerontol. 56, 147–153. doi: 10.1016/j.exger.2014.02.015
Schmidt, J. V., Su, G. H., Reddy, J. K., Simon, M. C., and Bradfield, C. A. (1996). Characterization of a murine Ahr null allele: involvement of the Ah receptor in hepatic growth and development. Proc. Natl. Acad. Sci. U.S.A. 93, 6731–6736. doi: 10.1073/pnas.93.13.6731
Senft, A. P., Dalton, T. P., Nebert, D. W., Genter, M. B., Puga, A., Hutchinson, R. J., et al. (2002). Mitochondrial reactive oxygen production is dependent on the aromatic hydrocarbon receptor. Free Radic. Biol. Med. 33, 1268–1278. doi: 10.1016/s0891-5849(02)01014-6
Shimizu, Y., Nakatsuru, Y., Ichinose, M., Takahashi, Y., Kume, H., Mimura, J., et al. (2000). Benzo[a]pyrene carcinogenicity is lost in mice lacking the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. U.S.A. 97, 779–782. doi: 10.1073/pnas.97.2.779
Singh, K. P., Bennett, J. A., Casado, F. L., Walrath, J. L., Welle, S. L., and Gasiewicz, T. A. (2014). Loss of aryl hydrocarbon receptor promotes gene changes associated with premature hematopoietic stem cell exhaustion and development of a myeloproliferative disorder in aging mice. Stem Cells Dev. 23, 95–106. doi: 10.1089/scd.2013.0346
Smirnova, A., Wincent, E., Vikstrom Bergander, L., Alsberg, T., Bergman, J., Rannug, A., et al. (2016). Evidence for new light-independent pathways for generation of the endogenous aryl hydrocarbon receptor agonist FICZ. Chem. Res. Toxicol. 29, 75–86. doi: 10.1021/acs.chemrestox.5b00416
Smith, C. J., O’brien, T., Chatzigeorgiou, M., Spencer, W. C., Feingold-Link, E., Husson, S. J., et al. (2013). Sensory neuron fates are distinguished by a transcriptional switch that regulates dendrite branch stabilization. Neuron 79, 266–280. doi: 10.1016/j.neuron.2013.05.009
Sonowal, R., Swimm, A., Sahoo, A., Luo, L., Matsunaga, Y., Wu, Z., et al. (2017). Indoles from commensal bacteria extend healthspan. Proc. Natl. Acad. Sci. U.S.A. 114, E7506–E7515. doi: 10.1073/pnas.1706464114
Sutter, C. H., Olesen, K. M., Bhuju, J., Guo, Z., and Sutter, T. R. (2019). AHR regulates metabolic reprogramming to promote SIRT1-dependent keratinocyte differentiation. J. Invest. Dermatol. 139, 818–826. doi: 10.1016/j.jid.2018.10.019
Tappenden, D. M., Lynn, S. G., Crawford, R. B., Lee, K., Vengellur, A., Kaminski, N. E., et al. (2011). The aryl hydrocarbon receptor interacts with ATP5alpha1, a subunit of the ATP synthase complex, and modulates mitochondrial function. Toxicol. Appl. Pharmacol. 254, 299–310. doi: 10.1016/j.taap.2011.05.004
Tigges, J., Krutmann, J., Fritsche, E., Haendeler, J., Schaal, H., Fischer, J. W., et al. (2014). The hallmarks of fibroblast ageing. Mech. Ageing Dev. 138, 26–44. doi: 10.1016/j.mad.2014.03.004
Villeponteau, B. (1997). The heterochromatin loss model of aging. Exp. Gerontol. 32, 383–394. doi: 10.1016/s0531-5565(96)00155-6
Wang, Q., Kurita, H., Carreira, V., Ko, C. I., Fan, Y., Zhang, X., et al. (2016). Ah receptor activation by dioxin disrupts activin, BMP, and WNT signals during the early differentiation of mouse embryonic stem cells and inhibits cardiomyocyte functions. Toxicol. Sci. 149, 346–357. doi: 10.1093/toxsci/kfv246
Williams, E. G., Mouchiroud, L., Frochaux, M., Pandey, A., Andreux, P. A., Deplancke, B., et al. (2014). An evolutionarily conserved role for the aryl hydrocarbon receptor in the regulation of movement. PLoS Genet. 10:e1004673. doi: 10.1371/journal.pgen.1004673
Wu, D., Nishimura, N., Kuo, V., Fiehn, O., Shahbaz, S., Van Winkle, L., et al. (2011). Activation of aryl hydrocarbon receptor induces vascular inflammation and promotes atherosclerosis in apolipoprotein E−/− mice. Arterioscler. Thromb. Vasc. Biol. 31, 1260–1267. doi: 10.1161/ATVBAHA.110.220202
Xu, M., Pirtskhalava, T., Farr, J. N., Weigand, B. M., Palmer, A. K., Weivoda, M. M., et al. (2018). Senolytics improve physical function and increase lifespan in old age. Nat. Med. 24, 1246–1256. doi: 10.1038/s41591-018-0092-9
Xue, Z., Li, D., Yu, W., Zhang, Q., Hou, X., He, Y., et al. (2017). Mechanisms and therapeutic prospects of polyphenols as modulators of the aryl hydrocarbon receptor. Food Funct. 8, 1414–1437. doi: 10.1039/c6fo01810f
Yano, M., Terada, K., and Mori, M. (2003). AIP is a mitochondrial import mediator that binds to both import receptor Tom20 and preproteins. J. Cell Biol. 163, 45–56. doi: 10.1083/jcb.200305051
Keywords: aryl hydrocarbon receptor, aging, mitochondria, C. elegans, mice, human
Citation: Brinkmann V, Ale-Agha N, Haendeler J and Ventura N (2020) The Aryl Hydrocarbon Receptor (AhR) in the Aging Process: Another Puzzling Role for This Highly Conserved Transcription Factor. Front. Physiol. 10:1561. doi: 10.3389/fphys.2019.01561
Received: 01 November 2019; Accepted: 11 December 2019;
Published: 14 January 2020.
Edited by:
Ovidiu Constantin Baltatu, Khalifa University, United Arab EmiratesReviewed by:
Ian James Martins, Edith Cowan University, AustraliaMordhwaj S. Parihar, Vikram University, India
Copyright © 2020 Brinkmann, Ale-Agha, Haendeler and Ventura. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Judith Haendeler, judith.haendeler@hhu.de; Natascia Ventura, natascia.ventura@hhu.de