 B. Ortega-Contreras1
B. Ortega-Contreras1 A. Armella1
A. Armella1 J. Appel1
J. Appel1 D. Mennickent1,2J. Araya2
D. Mennickent1,2J. Araya2 M. González3E. Castro4A. M. Obregón5
M. González3E. Castro4A. M. Obregón5 L. Lamperti1
L. Lamperti1 J. Gutiérrez6
J. Gutiérrez6 E. Guzmán-Gutiérrez1*
E. Guzmán-Gutiérrez1*- 1Pregnancy Diseases Laboratory, Department of Clinical Biochemistry and Immunology, Faculty of Pharmacy, Universidad de Concepción, Concepción, Chile
- 2Department of Instrumental Analysis, Faculty of Pharmacy, Universidad de Concepción, Concepción, Chile
- 3Department of Obstetrics and Gynecology, Faculty of Medicine, Universidad de Concepción, Concepción, Chile
- 4Departamento de Obstetricia y Puericultura, Facultad de Ciencias de la Salud, Universidad de Atacama, Copiapó, Chile
- 5Faculty of Health Care, Universidad San Sebastián, Concepción, Chile
- 6Faculty of Health Sciences, Universidad San Sebastián, Santiago,Chile
Gestational Diabetes Mellitus (GDM) is a highly prevalent maternal pathology characterized by maternal glucose intolerance during pregnancy that is, associated with severe complications for both mother and offspring. Several risk factors have been related to GDM; one of the most important among them is genetic predisposition. Numerous single nucleotide polymorphisms (SNPs) in genes that act at different levels on various tissues, could cause changes in the expression levels and activity of proteins, which result in glucose and insulin metabolism dysfunction. In this review, we describe various SNPs; which according to literature, increase the risk of developing GDM. These SNPs include: (1) those associated with transcription factors that regulate insulin production and excretion, such as rs7903146 (TCF7L2) and rs5015480 (HHEX); (2) others that cause a decrease in protective hormones against insulin resistance such as rs2241766 (ADIPOQ) and rs6257 (SHBG); (3) SNPs that cause modifications in membrane proteins, generating dysfunction in insulin signaling or cell transport in the case of rs5443 (GNB3) and rs2237892 (KCNQ1); (4) those associated with enzymes such as rs225014 (DIO2) and rs9939609 (FTO) which cause an impaired metabolism, resulting in an insulin resistance state; and (5) other polymorphisms, those are associated with growth factors such as rs2146323 (VEGFA) and rs755622 (MIF) which could cause changes in the expression levels of these proteins, producing endothelial dysfunction and an increase of pro-inflammatory cytokines, characteristic on GDM. While the pathophysiological mechanism is unclear, this review describes various potential effects of these polymorphisms on the predisposition to develop GDM.
Highlights
1) Several SNPs cause predisposition to GDM.
2) SNPs associated with GDM mainly affect endocrine pancreas function and adipose tissue response to insulin.
3) Physiopathology induced by these SNPs could explain GDM development.
1 Introduction
The American Diabetes Association (ADA) defines gestational diabetes mellitus (GDM) as a type of diabetes diagnosed at the second or third trimester of pregnancy in a mother not diagnosed with pregestational diabetes (ADA, 2020). In recent years, it has been observed that the incidence of this pathology is increasing along with obesity and type 2 diabetes mellitus (T2DM) (Coustan, 2013; ADA, 2019). It is estimated that the worldwide prevalence of GDM varies between 1.7 and 11.7% (Griffin et al., 2000; Schneider et al., 2012). This considerable variation is due to differences among the populations and diagnostic criteria used in each country. It has been estimated that countries with higher incidence of GDM are those of Middle East and North Africa with 12.9%, followed by Southeast Asia (11.7%) (Zhu and Zhang, 2016). In developed countries such as the United States, Australia, Canada and the United Kingdom, its prevalence is less than 6% (Zhao et al., 2016). South American countries show high GDM prevalence: about 15% of pregnant women from Peru and Chile have been diagnosed with GDM in the last 20 years (Belmar et al., 2004; Huidobro et al., 2004; Larrabure-Torrealva et al., 2018; Garmendia et al., 2019).
One of the proposed risk factors for GDM is obesity. In fact, GDM women usually have a body mass index (BMI) higher or equal to 25 kg/m2 (Shah et al., 2011). An increase in proinflammatory cytokines have been reported in obese pregnant women affected with GDM (Kinalski et al., 2005; Kuzmicki et al., 2008; Pantham et al., 2015). This pro-inflammatory state stimulates the synthesis of xanthurenic acid, which has been associated with the development of T2DM, prediabetes, and GDM (Bennink and Schreurs, 1975; Oxenkrug, 2015; Law and Zhang, 2017). Moreover, in GDM pregnant women there is a supraphysiological insulin resistance state induced in part by pro-inflammatory cytokines (Sonagra et al., 2014). For that reason, GDM pregnancies are associated with high HOMA-IR index values (Wang et al., 2018).
The main maternal outcomes of GDM are hyperglycemia, GDM in future pregnancies, future development of T2DM (Griffin et al., 2000; Ben-Haroush et al., 2004), obesity, and preeclampsia (Coustan, 2013; Kampmann et al., 2015). A study by the HAPO Study Cooperative Research Group demonstrated that in GDM pregnancies, the main fetal outcomes are macrosomia, neonatal hyperinsulinemia, caesarean section, and neonatal hypoglycemia (HAPO Study Cooperative Research Group, 2002; HAPO Study Cooperative Research Group, 2008). Furthermore, newborns from GDM pregnancies have a high risk of developing T2DM and obesity in the long-term (Dabelea et al., 2000).
Maternal and fetal outcomes are associated with modifiable and non-modifiable factors (Shaat et al., 2007). One of these non-modifiable factors is genetics. In this line, diverse Genome-Wide Association Studies (GWAS) have shown that genetic variables of the single nucleotide polymorphisms (SNPs) type associated with T2DM have also been related with high predisposition to GDM, which has been studied in diverse populations (Kwak et al., 2012; Huerta-Chagoya et al., 2015; Lowe et al., 2016). This association has been proposed because both GDM and T2DM are associated with similar pathophysiology mechanisms, including insulin resistance and a chronic inflammatory state (Zajdenverg and Negrato, 2017). Furthermore, GDM increases the risk of progression to T2DM; nevertheless, the magnitude of this effect is variable in different populations (Plows et al., 2018; Vounzoulaki et al., 2020). Unlike T2DM, GDM is triggered by placental and maternal hormones that cause a transitory insulin resistance that in most cases disappears after pregnancy, and only affects pregnant women (Mack and Tomich, 2017).
The positive associations of GDM and genetic variations is not clear in all cases (Anghebem-Oliveira et al., 2017), and the mechanisms by which these could contribute to GDM development have not been fully described. Therefore, this review summarizes the main genetic variants that have been described for GDM, emphasizing their potential pathophysiological mechanisms on the generation of GDM.
2 Genetic Factors Associated With Gestational Diabetes Mellitus and Its Pathophysiological Mechanism Over the GDM Etiology
GDM etiology could be associated with genetic variations that are related to T2DM development (Buchanan and Xiang, 2005; Gong et al., 2016). GWAS have identified numerous loci associated with the risk of GDM (Dalfrà et al., 2020). In this review, we classify different SNPs that have been associated with T2DM and GDM, according to the protein they encode (Table 1).
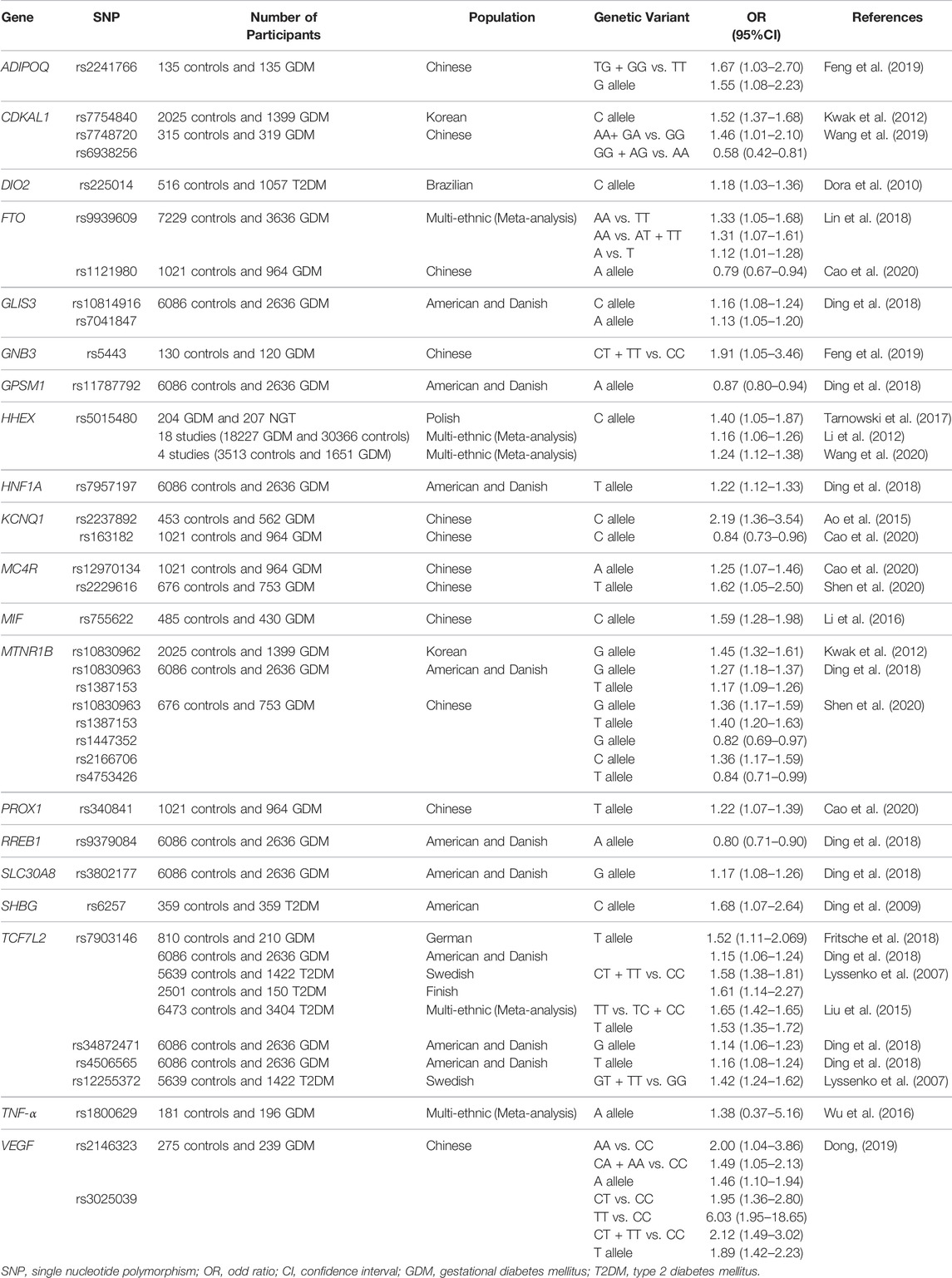
TABLE 1. Single nucleotide polymorphisms associated with gestational diabetes mellitus.
The presence of genetic variants can result in changes in the expression and function of the encoded protein, affecting diverse physiological actions. For this reason, polymorphisms can be clinically relevant for various pathologies (Ramírez-Bello and Jiménez-Morales, 2017). In the following section, we focus on explaining the pathophysiological mechanisms caused by the main aforementioned genetic factors.
2.1 Polymorphisms Associated With Transcription Factors
2.1.1 TCF7L2 Genetic Variants Associated to GDM
The transcription factor 7-like 2 (TCF7L2) gene is located on chromosome 10 and encodes for a nuclear protein that participates in gene expression regulation involved in the fusion of insulin secreting granules in pancreatic beta cells (da Silva et al., 2009). In a multiracial study, carriers of the T allele of SNP rs7903146 (risk-allele) showed an increased risk of T2DM (OR: 1.58) and a faster deterioration of insulin secretion (Lyssenko et al., 2007; Zhang et al., 2013; Liu et al., 2015). Interestingly, Asian pregnant women homozygous for the TT genotype, had a higher risk of developing GDM (OR = 2.08), followed by Hispanic/Latin (OR = 1.80), and white (OR = 1.51) (Lin et al., 2016). Other ethnic groups have also been studied (Shaat et al., 2007; Zhang et al., 2013; Wu et al., 2016). Therefore, the presence of the T allele in the rs7903146 variant could be a genetic risk factor for GDM. Other SNPs of the same gene have been related with a higher risk to develop GDM in a meta-analysis (Chang et al., 2017), i.e., carriers of the T allele for the rs12255372 variant (OR: 1.46) (Zhang et al., 2013).
TCF7L2 is expressed in several tissues, including the islets of Langerhans, and liver (Zhou et al., 2014). This factor participates as a transcriptional effector in the Wnt signaling pathway where it regulates the transcription of diverse genes, including some of those involved in the production and function of incretin hormones and in blood glucose homoeostasis (Schinner et al., 2008; Ip et al., 2012).
Reduced mRNA levels of this gene in the pancreatic islets are related to a significant increase in the apoptosis of beta cells and a decrease in their proliferation, which causes a reduction in insulin secretion (Shu et al., 2008; Savic et al., 2011). Moreover, evidence in the murine model Tcf7l2 (null), shown an alteration in glucose metabolism causing hypoglycemia. In contrast, the overexpression of this gene in the same animal model using Cre recombinase resulted in glucose intolerance (Savic et al., 2011). This results are related to the high expression levels of TCF7L2 mRNA in T2DM patients (Lyssenko et al., 2007), and could be an interesting idea for study in GDM models.
The SNP rs7903146 of the TCF7L2 gene is located in its intronic region upstream exon 5 in a regulatory site, specifically in an islet-selective open chromatin site. This means that in human islet cells, the chromatin state at rs7903146 is more open in chromosomes carrying the T allele, and have a greater enhancer activity compared to the C allele (Gaulton et al., 2010). This could explain the association of the T-allele with an impaired glucose-stimulated insulin secretion (Dahlgren et al., 2007). Moreover, carriers of this risk allele exhibit a significantly higher expression of TCF7L2 mRNA in the pancreatic islets and an increased hepatic glucose production (Lyssenko et al., 2007). Carriers of the TT genotype for this polymorphism have higher concentrations of blood glucose, proinsulin, and incretin hormones, compared to the normal genotype group (Gjesing et al., 2011). Likewise, the insulinogenic index, derived from an oral glucose tolerance test, is diminished in carriers of the T-allele, which would be associated with an impaired insulin secretion and not with resistance to this hormone (Saxena et al., 2006). However, there are not work in GDM models, or human that demonstrated this association in pregnancies with glucose intolerance.
2.1.2 HHEX Genetic Variant Associated to GDM
GWAS have associated several polymorphisms with GDM, and the Hematopoietically-expressed homeobox (HHEX) gene rs5015480 stands out among them. This gene codifies for a transcription factor that is part of a homeobox gene family involved in developmental and hematopoietic differentiation processes (Bedford et al., 1993). In a study conducted in Poland, the risk C-allele was associated with a genetic predisposition to develop GDM (OR: 1.40) and also to an increased BMI in pregnant women (Tarnowski et al., 2017). In addition, a meta-analysis showed a strong association of the CC genotype in contrast to the TT genotype (OR: 1.65) in different populations (Wang et al., 2020).
HHEX is a transcription factor that is also linked to the Wnt signaling pathway. It is essential for cell growth and for the development of organs such as the thyroid, pancreas, liver, and brain (Tarnowski et al., 2017). In the adult endocrine pancreas, this factor is selectively expressed in the somatostatin-secreting delta cells, where it has been observed to regulate the differentiation of this cell type (Zhang J. et al., 2014). It is described that a decrease in somatostatin levels causes a paracrine inhibition of the insulin release from β cells (Moldovan et al., 1995). In this context, it has been suggested that this transcription factor directly activates the transcription of the somatostatin gene (Zhang J. et al., 2014).
In the HHEX gene, the risk C-allele of the rs5015480 variation is associated with altered β cells secretion. Other variations of this gene are also involved in reduced fasting insulin secretion, insulin sensitivity, and glucose-stimulated insulin secretion (Pivovarova et al., 2009). Some authors have proposed that the rs5015480 variation could affect early stages of insulin secretion due to the reduced insulinogenic index, described in risk allele carriers (Dimas et al., 2014), which could be related to insulin secretion regulation exerted by delta cells (Zhang J. et al., 2014).
Genetic modifications in these transcription factors could cause alterations in the protein expression involved in the development of GDM. In fact, it is proposed that both TCF7L2 and HHEX have an important role in the regulation of insulin secretion in GDM (Saxena et al., 2006; Dimas et al., 2014). In fact, the polymorphisms TCF7L2 rs7903146 and HHEX rs5015480 could decrease insulin secretion during pregnancy, favoring GDM development due to an alteration at the level of pancreatic cells; and consequently, in the production and secretion of insulin.
2.2 Polymorphisms Associated With Hormones
2.2.1 ADIPOQ Genetic Variant Associated to GDM
A relevant hormone in GDM is adiponectin, due to its protective role against insulin resistance. The risk G-allele in rs2241766 (T > G) produces a silent mutation in nucleotide 45 in the adiponectin gene (ADIPOQ). This genetic variation is associated with obesity in several studies. Also, carriers of the G/G genotype for this polymorphism had a higher risk of T2DM (OR: 1.70) compared to the T/T genotype (Hara et al., 2002). Moreover, a meta-analysis (Bai et al., 2020) demonstrated an increased risk of GDM in Asian (OR: 2.08) and European (OR: 1.52), but a diminished risk in the American population (OR: 0.642) carriers of the G allele of this SNP.
The adiponectin hormone is synthesized in adipose tissue, where it modulates diverse metabolic processes, among them: metabolism of lipids and fatty acids, reduction of plasmatic triglycerides and improvement of glucose metabolism by an increase in insulin sensitivity (Karbowska and Kochan, 2006). Moreover, adiponectin reduces the expression of adhesion molecules in endothelial cells, the transformation of macrophages into foamy cells, the expression of tumor necrosis factor α (TNF-α) and the proliferation of smooth muscle tissue cells (Mierzyński et al., 2018).
There are genetic variations in the adiponectin gene (ADIPOQ) such as rs2241766, where the risk G-allele produces a silent mutation in nucleotide 45, which does not cause an amino acid change. However, being very close to the exon-intron limit, it could affect the splicing machinery (Salazar et al., 2010). The presence of this allele can completely inactivate the activity of the adiponectin promoter and the expression of the ADIPOQ gene, and thus can decrease adiponectin levels (Chu et al., 2013).
During pregnancy, the plasmatic levels of this hormone are normally decreased; however, such levels are even lower in the presence of this variation (Huang et al., 2019). In fact, lower levels of adiponectin could be related to an increase in the formation of dense LDL particles (Lara-Castro et al., 2007), and to an increase in the expression of pro-inflammatory cytokines such as TNF-α, IL-6 and IL-8 (Hussain et al., 2018).
All this evidence supports the idea that the adiponectin gene could be a susceptibility factor for developing GDM (Takhshid et al., 2015; Xu et al., 2016).
2.2.2 SHBG Genetic Variant Associated to GDM
The SHBG gene codifies for the sex hormone-binding globulin, which could contribute to the pathophysiology of GDM. This gene has been associated with the risk of diabetes mellitus (Ding et al., 2009; Hedderson et al., 2014). In fact, low levels of pre-gestational SHBG increase the risk of developing GDM (OR: 4.06) (Hedderson et al., 2014).
In addition, the rs6257 (T > C) polymorphism of this gene has been associated with plasmatic concentrations of SHBG. It has been shown that carriers of the CC or CT risk genotypes have lower plasmatic concentrations of this hormone compared to the carriers of the normal TT genotype (Ding et al., 2009; Hedderson et al., 2014). Although there are still not studies that demonstrate association between GDM and these genetics variants, changes in SHBG concentration due to these variants, could increase this risk.
SHBG is synthesized in the liver and is responsible for the transport of androgens in circulation and for the regulation of the bioavailability of these hormones. It is also involved in receptor-mediated processes where it regulates the effects of dihydrotestosterone (DHT) and estradiol (Fortunati, 1999).
An in-silico study postulates that rs6257 favors the union of the forkhead box protein A2 (FOXA2) element to the SHBG gene, repressing its transcription by a splicing defect (Ding et al., 2009). This idea was tested in HepG2 cells (Wu, 2015). Furthermore, it is described that in GDM there is a decrease in the plasmatic concentrations of SHBG compared to pregnant women without this pathology (Bartha et al., 2000; Tawfeek et al., 2017; Faal et al., 2019). This phenomenon was also studied in a trophoblast cell model (HTR8 Sv-neo) exposed to high levels of insulin, showing a decrease in SHBG mRNA and protein levels (Zhang et al., 2016; Feng et al., 2018). This could be explained by a reduction in the signaling of the phosphatidylinositol 3-kinase (PI3K/Akt) pathway, which mediates the transduction of insulin signals (Feng et al., 2018). Although literature is scarce, the reduction of GLUT-4, GLUT-3 and IRS-1 expression in GDM patients could be correlated with a lower SHBG activity, which could favor insulin resistance (Zhang et al., 2016).
2.3 Polymorphisms Associated With Membrane Proteins
2.3.1 GNB3 Genetic Variant Associated to GDM
Heterotrimeric G-proteins are relevant components of transmembrane receptors and are involved in the regulation of different intracellular signaling pathways. The 825C > T SNP in the gene of the G-protein β3 subunit (GNB3) has been linked to metabolic features such as hypertension, atherosclerosis and immunological response. The T-allele has also been associated with obesity risk in German, Chinese and South African populations (Siffert et al., 1999). In pregnant women, carriers of the risk allele have a higher weight gain during gestation (Dishy et al., 2003). Interestingly, it has also been described that the CT and TT genotypes are significantly related to a higher risk of GDM (OR: 1.91; 95%CI: 1.053–3.463) (Feng et al., 2019); however, there is still a lack of studies to confirm this relationship.
The G protein is involved in the regulation of glucose levels through the metabolic pathway of insulin signaling. Also, it is involved in the stimulation of second messengers such as adenylate cyclase, epinephrine signaling pathways and glucagon receptors in the liver, muscular and fatty tissue cells (Rizvi et al., 2016).
The most studied polymorphism of the GNB3 gene is C825T, which is produced in exon 10 and it generates an alternative splicing causing the loss of 41 amino acids, structurally modifying this protein. The risk T-allele has been associated with a higher production of the G protein beta three subunit, causing an increased activation of this protein. With that increased activation comes an enhanced activity of the potassium channels at the cardiac level and vasoconstriction mediated by α-adrenoreceptors, which are directly related with arterial hypertension (Andersen et al., 2006). Another study demonstrated that the CC genotype of this SNP might be associated with higher obesity-related metabolic traits, such as triglyceride and total cholesterol in non-obese subjects (Hsiao et al., 2013).
Unfortunately, there is no direct mechanism that explains the relationship of this polymorphism and insulin resistance. However, obesity-related metabolic traits are tightly linked to GDM pathophysiology.
2.3.2 KCNQ1 Genetic Variant Associated to GDM
It is suggested that the gene of the potassium voltage-gated channel subfamily Q member 1 (KCNQ1), participates on the regulation of insulin secretion in the pancreas, and it has been described as one of the candidate genes for GDM. Evidence shows that the rs2237892 variation of this gene is significantly associated with increased glucose levels, impaired insulin secretion, and higher GDM risk (OR: 1.99) in Asian population (Yasuda et al., 2008; Shin et al., 2010; Ao et al., 2015). Correspondingly, a study including 637 Pakistani women associated the presence of the risk allele A of this polymorphism, with an enhanced GDM risk (OR: 2.07) (Fatima et al., 2016). On the other hand, the risk genotype of rs163182 has been associated with lower GDM risk (OR: 0.84) (Cao et al., 2020).
The KCNQ1 gene encodes for the alpha subunit of the pore-forming potassium channel (KvLQT1) performing an important role in the control of the vascular repolarization process (Yasuda et al., 2008). This gene is expressed in epithelial cells, including those of the endocrine and exocrine pancreas. In addition, KCNQ1 channels are expressed in insulin-secretor INS-1 cells, where they depolarize the membrane potential of the pancreatic beta cells allowing insulin secretion (Kwak et al., 2010).
Genetic variations in the sequence of KCNQ1 such as rs2237892 cause changes in the translation and/or in post-translational modifications, reducing β cells function (Jonsson et al., 2009). Thus, a decreased secretory capacity of the β cells could increase GDM risk by reducing the secretory capacity of insulin and limiting its compensation (Wang et al., 2013). In fact, GDM patients carrying the risk allele have higher fasting plasma glucose levels and lower insulin secretion (Wang et al., 2013).
2.4 Polymorphisms Associated to Enzymes
2.4.1 DIO2 Genetic Variant Associated to GDM
The DIO2 gene codifies to an enzyme that is ubiquitous in the human body, and that regulates the conversion of tetraiodothyronine (T4) to triiodothyronine (T3). It is proposed that the rs225014 (T > C) polymorphism causes a change in the amino acid sequence from a threonine (Thr) to an alanine (Ala) on codon 92 (Thr92Ala) (Estivalet et al., 2011). This variation has been associated with higher GDM risk (OR: 1.29) (Asadi et al., 2016).
Deiodinase 2 (DIO2) beside the essential role in thyroid hormones homeostasis, is also involved in brain growth and maturation, glucose uptake in the muscle, among other effects (Loubiere et al., 2010; Galton et al., 2014; Yang et al., 2016). This enzyme is mainly found in the central nervous system, hypophysis, skeletal muscle, thyroid, heart, bones, and adipose tissue (Coppotelli et al., 2006).
The rs225014 genotype is associated with DIO2 expression (Bomer et al., 2015), and the presence of the risk C allele, which causes the substitution of a Threonine for an Alanine, reducing the DIO2 activity (Estivalet et al., 2011). In fact, it has been observed that homozygous patients for this polymorphism have a lower enzymatic activity, evidenced by a 37% decrease in DIO2 speed in skeletal muscle, and a reduction around 90% in its maximal velocity (Vmax). These results may explain the association of this variation with insulin resistance. As the skeletal muscle is the main site of insulin-dependent glucose uptake, a lower DIO2 activity would decrease the amount of T3 generated in the skeletal muscle, and with this, the expression of genes involved in energy use, such as GLUT4, leading to insulin resistance (Canani et al., 2005).
2.4.2 FTO Genetic Variant Associated to GDM
The fat mass and obesity-associated gene (FTO) codifies for a dioxygenase enzyme that is found within the cell nucleus. It is reported that FTO participates in mRNA processing and splicing processes. Regarding the FTO gene, the polymorphisms rs9939609, rs8050136, and rs1421085 are associated with BMI increase, risk of obesity, and T2DM. However, controversy still exists about the association between certain polymorphisms and GDM (Hotta et al., 2008; He et al., 2018). Indeed, rs8050136 and rs14211085 SNPs were not associated with GDM risk (de Melo et al., 2015; Anghebem-Oliveira et al., 2017; Lin et al., 2018; Tarnowski et al., 2018), but with proinflammatory state and weight gain during pregnancy (Saucedo et al., 2017).
The FTO rs9939609 (T > A) polymorphism is located in the first intron of this gene (Frayling et al., 2007). Recent evidence suggests that this polymorphism is strongly correlated with GDM risk (OR: 1.31) in Caucasian subjects (Lin et al., 2018). Also, the A-allele is associated with rapid weight gain during pregnancy (Lawlor et al., 2011; Martins et al., 2016).
These genetic variants could alter the expression or enzymatic activity of FTO, leading to changes in the metabolism that could impair glucose metabolism and generate insulin resistance (Saber-Ayad et al., 2019). Subsequently, the GDM risk could be increased.
The FTO protein acts within the nucleus demethylating the N6-methyladenosines in mRNA and regulating the splicing of genes involved in adipogenesis such as FABP4, PPARγ, C/EBPα and PLIN1 (Zhao et al., 2014a; Ben-Haim et al., 2015; Merkestein et al., 2015; Zhang et al., 2015; Bartosovic et al., 2017).
In murine models, the Fto gene is widely expressed in the brain, including the hypothalamic nucleus, which is related to energy intake regulation (Speakman, 2015). Studies in primary culture of Mouse Embryonic Fibroblasts (MEFs) have demonstrated that FTO regulates the splicing of the RUNX1T1 (Runt-related transcription factor 1) gene (Zhao et al., 2014b), which is involved in the early stages of adipogenesis (Rochford et al., 2004). FTO exerts its action during the splicing of the mRNA transcript of this gene, causing the exclusion of exon 6 and generating a short pro-adipogenic isoform of RUNX1T1, which increases adipocytes proliferation. The latter is a condition that is favored when FTO is overexpressed (Merkestein et al., 2015). Moreover, FTO overexpression in C57B/6J mice is associated with weight gain and increased adipogenic activity (Merkestein et al., 2015).
Variations in the first intron of the FTO gene have been associated with higher BMI and T2DM, and it has been reported that there is a 47 kb region that comprehends several SNPs associated with these pathologies. The rs9939609 variant has been extensively studied (Frayling et al., 2007). The mechanism by which this variant causes obesity is still unclear, however, heterozygous subjects for this polymorphism have higher levels of primary FTO transcripts of the risk A-allele, than of the T-allele (Berulava and Horsthemke, 2010), and this could cause higher levels of FTO expression favoring adipogenesis. However, the latter relationship has not yet been reported in literature.
Another SNP close to rs9939609 and associated with an increased BMI is rs8050136. The DNA sequence that included A-allele of this polymorphism preferentially binds with CUTL1, a transcription factor that increases the FTO expression. It has been proposed that, given the proximity of these variants, the rs9939609 SNP could also increase FTO expression by the same mechanism (Stratigopoulos et al., 2008).
2.5 Polymorphisms Associated With Growth Factors
2.5.1 VEGFA Genetic Variant Associated to GDM
Another group of genes associated with GDM are growth factors. One of them is the vascular endothelial growth factor A (VEGFA), which has an essential role in angiogenesis by inducing the migration of endothelial precursor cells from the bone marrow, and causing differentiation and proliferation of endothelial progenitor cells in angiogenesis sites (Saavedra et al., 2017). A study involving the VEGFA gene showed that the A allele of the rs2146323 polymorphism and the T allele of the rs3025039 variant, increase the risk of developing GDM (OR: 1.456 and 1.894, respectively) (Dong, 2019). Additionally, the frequency of the risk haplotypes of rs2010963, rs833069, rs2146323 and rs3025010 is higher in GDM patients compared to normal pregnancies (Dong, 2019).
VEGFA has the function of promoting endothelial cell proliferation and increasing vascular permeability to induce angiogenesis. In fact, the placentas of GDM patients show hypervascularization, which is explained by a greater demand of oxygen by the fetus due to an increase in fetal aerobic metabolism stimulated by insulin (Troncoso et al., 2017). A study conducted in a murine model with GDM induced by a high fat diet, showed that those who had GDM had a high placental inflammatory response evidenced by increased IL-1β and TNFα, and a higher degree of placental hypoxia denoted by an enhanced expression of inducible hypoxia factor-1α (HIF-1α) and VEGF-A. In this sample, altered placental vascular development due to hypervascularization was also present (Li, 2013). Therefore, in GDM, VEGF would be involved in angiogenesis processes in the placenta, but it is not clear whether genetic alterations could explain this mechanism. In fact, variations of the VEGF gene (rs2146323 and rs3025039) have only been associated with an increased risk of GDM, and therefore could be caused by an abnormal expression of VEGF, increasing its levels in these patients (Dong, 2019).
However, molecular mechanisms explaining this dysfunction have not been described for these polymorphisms. Evidence for rs735286 indicates that it is located in the second VEGF intron, and this variant involves a region that has putative binding sites for transcription factors such as the myeloid zinc finger protein (MZF-1), that regulates VEGFA expression. Therefore, rs735286 could affect the transcription process and cause changes in splicing (Morris et al., 1994; Churchill et al., 2008).
Although the rs2146323 polymorphism is also found in intron 2, it is located relatively distant from these binding sites, so the mechanism would be apparently different (Churchill et al., 2008). On the other hand, it has been suggested that rs3025039, being in a 3′-UTR region, could alter the conformation of the mRNA, which would decrease the transcript degradation, causing an increase in VEGF expression (Dibbens et al., 1999; Tahara et al., 2009).
2.5.2 MIF Genetic Variant Associated to GDM
The macrophage migration inhibitory factor (MIF), is a pro-inflammatory cytokine secreted by T-lymphocytes in response to delayed-type hypersensitivity, which exerts an inhibitory effect on macrophage migration. The rs755622 polymorphism of this gene (MIF-173G/C) is associated with higher GDM risk (OR: 1.59) (Li et al., 2016), and the rs1007888 polymorphism is related to high levels of blood glucose and insulin (Zhan et al., 2015).
MIF is a pro-inflammatory cytokine that is secreted in response to delayed-type hypersensitivity and exerts an inhibitory effect on macrophage migration (Hamidi et al., 2019). It has been described that MIF controls underlying metabolic and inflammatory processes during periods of stress, regulating glucose homeostasis and macrophage infiltration into adipose tissue. Also, MIF is expressed and secreted by adipose tissue, evidencing higher levels in obesity (Mejia-Montilla et al., 2015).
The rs755622 variation is located in the promoter region of MIF and the C allele is related with a higher transcriptional activity of the MIF gene. In fact, patients with T2DM have shown increased MIF levels (Hamidi et al., 2019). Also, it has a direct association with insulin resistance through the production of some pro-inflammatory cytokines and adipokines, including resistin and IL-6. Additionally, this variation has been related to an increased risk of GDM in Chinese women; hence, more studies should be carried out in other populations to determine the role of this polymorphism in this pregnancy disease (Li et al., 2016). However, more studies are needed to confirm the association of these variants and GDM.
2.6 Other SNPs Associated With GDM
The IRS-1 is an intracellular adaptor protein that plays a key role in insulin signaling. The IRS1 rs1801278 (C > T) variant has been related to GDM in meta-analyses (Zhang Y. et al., 2014; Wu et al., 2016), similarly to the rs7578326 (A > G) polymorphism (Voight et al., 2010; Zheng et al., 2013; Zhao et al., 2017). IRS1 rs1801278 is a missense polymorphism that decreases the phosphorylation of IRS-1 in vitro (McGettrick et al., 2005), reducing the binding of p85 to IRS-1 and the activity of PI3K in different cell lines (Hribal et al., 2000; Sentinelli et al., 2006), and leading to insulin resistance. On the other hand, rs7578326 is an intron variant positioned in a distal regulatory element that targets IRS1 (Lu et al., 2013). The risk allele A removes a CpG site and avoids its methylation (Dayeh et al., 2013). This genetic variant has been linked to higher levels of transcript in skeletal muscle, with no changes in the levels of IRS1 mRNA (Soyal et al., 2015). Further studies are needed to elucidate the exact role of this SNP on the pathophysiology of T2DM and GDM.
The SNP 43, SNP 44, SNP 63, and Indel 19 variants of the CAPN10 gene were not associated with GDM risk (Shaat et al., 2005; Neuhaus et al., 2013; Khan et al., 2014; Zhang et al., 2019; Ustianowski et al., 2021), except in some genetic association models of SNP 63 and SNP 44 (Cui et al., 2016), and in CAPN10 SNP-43/19/63 haplotypes (Hou et al., 2017). Nevertheless, the Indel-19 and SNP-19 variants were associated with higher glucose levels in GDM women (Castro-Martínez et al., 2018). In addition, GWAS have discovered several genetic variants associated with both T2DM and GDM. Among them, the rs7754840 polymorphism of the CDKAL1 gene (C-allele) and the rs10830962 polymorphism of the MTNR1B gene (G-allele) have been associated to a higher GDM risk (OR: 1.518; OR: 1.454, respectively). The risk alleles of these polymorphisms have also been associated with a decreased fasting insulinemia in GDM women (Kwak et al., 2012).
In a meta-analysis, other polymorphisms were associated with the risk of developing GDM, such as the A allele of the rs1800629 polymorphism in the TNF-α gene (OR 2.69), the T-allele of the rs4402960 SNP in the IGF2BP2 gene (OR: 1.22), and the G-allele of the rs10830963 variant in the MTNR1B gene (OR: 1.28). This last variant is also associated with GDM risk, in Asian (OR 1.23) and Caucasian (OR: 1.49) populations, and women with pre-gestational BMI ≥25 kg/m2 (OR 1.24) (Wu et al., 2016).
3 Concluding Remarks
Scientific literature has evidenced various polymorphisms associated with GDM, which can affect one or several functions. However, the specific mechanism by how these polymorphisms can affect the body physiology has not been addressed. Figure 1 summarizes the main tissues that would be affected by the described polymorphisms during pregnancy. We emphasize that the main organs associated with energy metabolism such as skeletal muscle, adipose tissue and pancreas could be involved, but clearly the placenta is an organ that plays an important role in the context of a pregnancy-related pathology. Although the placenta is a fetal organ that is genetically different from the mother, it is difficult to know whether maternal genetic alterations could be linked to placental defects.
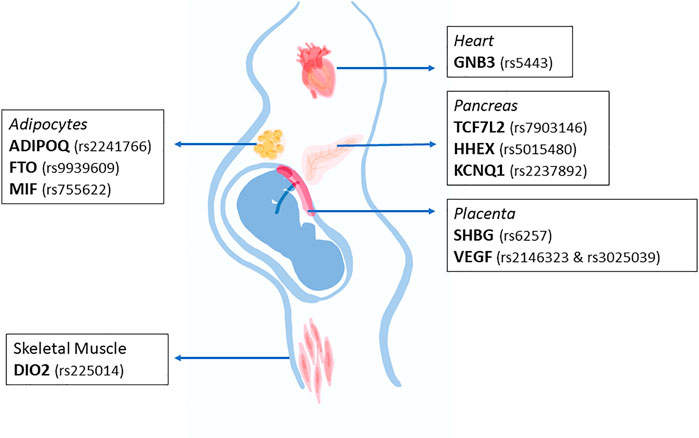
FIGURE 1. Major organs affected by the main SNPs associated with gestational diabetes mellitus. The main organs affected by the SNPs described in this review are heart, adipose tissue, pancreas, and skeletal muscle. Interestingly, the SNPs described would affect some functions in the indicated organ, favoring the onset of gestational diabetes mellitus.
The endocrine pancreas is a key organ in the synthesis of several hormones such as somatostatin, glucagon, and insulin. The latter is relevant to understand the pathophysiology of GDM. In fact, as mentioned earlier, GDM courses with supraphysiological insulin resistance. Accordingly, and as shown in Figure 2, the polymorphisms KCNQ1 rs2237892, TCF7L2 rs7903146, and HHEX rs5015480 are closely linked in the reduction of insulin secretion by beta cells, which could explain GDM development. One cause of insulin resistance is obesity. That explains why polymorphisms that have a consequence on metabolism, such as adipose tissue hyperplasia, are associated with GDM, as shown in Figure 3. Indeed, the polymorphisms FTO rs9939609, ADIPOQ rs2241766, and MIF rs755622 are strongly linked to the presence of insulin resistance.
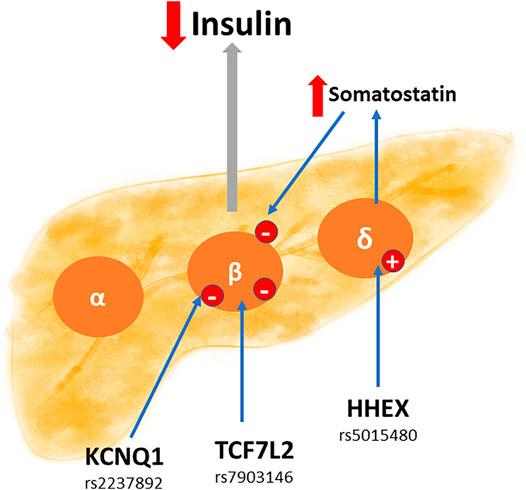
FIGURE 2. Potential SNPs capable of reducing the production of maternal insulin at a pancreatic level. Some SNPs can affect the secreting function of pancreatic insulin. In this sense, the SNPs KCNQ1 rs2237892, TCF7L2 rs7903146 and HHEX rs5015480, reduce (down red arrow) direct or indirectly the production and secretion of insulin at the level of the beta cells (β). Moreover, the same HHEX SNP can stimulate (up red arrow) somatostatin secretion in delta cells (δ), and this hormone is an inhibitor of insulin secretion. Impaired insulin production favors insulin resistance, and therefore the appearance of gestational diabetes mellitus.
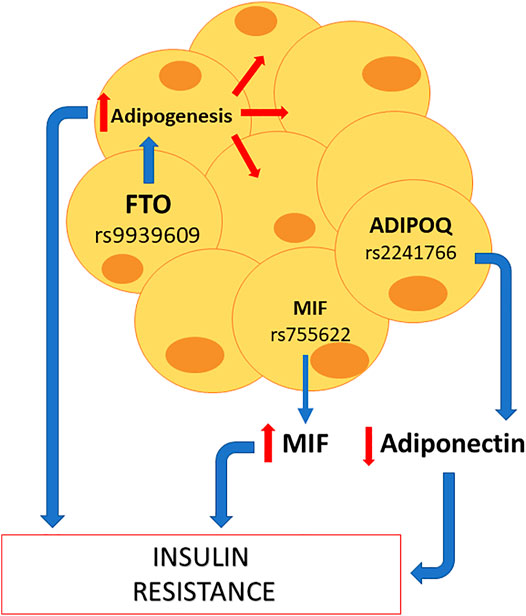
FIGURE 3. Potential SNPs capable of promoting maternal insulin resistance at adipose level. Some SNPs that can affect the adipocyte function are FTO rs9939609, which increases adipogenesis; MIF rs755622, which increases MIF levels; and ADIPOQ rs2241766, which reduces adiponectin levels. All these three effects favor the appearance of insulin resistance, and subsequently, gestational diabetes mellitus.
Unfortunately, evidence is insufficient to understand all the pathophysiological changes observed in GDM at the genetic level. In fact, GDM, as well as T2DM, are polygenic pathologies. However, current scientific evidence leads us to believe that certain polymorphisms could favor alterations in key organs, such as the pancreas and the adipose tissue, promoting insulin resistance during pregnancy and GDM.
Author Contributions
BO-C and AA tabulated literature information; BO-C, AA, JAP, MG, EC, AO, JG, LL and JAR prepared the figures and improved the manuscript; BO-C, DM and EG-G wrote the manuscript.
Funding
This work was supported by ANID, FOVI210057 for EG-G, the National Grant for Scientific and Technological Development, FONDECYT 11170710, and 1180935 for EG-G and JG; ANID (PhD scholarship 21190736) and Universidad de Concepción/MINEDUC (Project UCO fund 1866) for DM; ANID (MSc scholarship 22201750) for BO-C.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
American Diabetes Association (2020). 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes. Diabetes Care 43 (Suppl. 1), S14–S31. doi:10.2337/dc20-S002
American Diabetes Association (2019). Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2019. Diabetes Care 42 (Suppl. 1), S13–S28. doi:10.2337/dc19-S002
Andersen G., Overgaard J., Albrechtsen A., Glümer C., Borch-Johnsen K., Jørgensen T., et al. (2006). Studies of the Association of the GNB3 825C> T Polymorphism with Components of the Metabolic Syndrome in white Danes. Diabetologia 49 (1), 75–82. doi:10.1007/s00125-005-0049-7
Anghebem-Oliveira M. I., Martins B. R., Alberton D., Ramos E., Picheth G., Rego F. (2017). Type 2 Diabetes-Associated Genetic Variants of FTO, LEPR, PPARg, and TCF7L2 in Gestational Diabetes in a Brazilian Population. Arch. Endocrinol. Metab. 61 (3), 238–248. doi:10.1590/2359-3997000000258
Ao D., Wang H. J., Wang L. F., Song J. Y., Yang H. X., Wang Y. (2015). The Rs2237892 Polymorphism in KCNQ1 Influences Gestational Diabetes Mellitus and Glucose Levels: A Case-Control Study and Meta-Analysis. PLoS One 10 (6), e0128901. doi:10.1371/journal.pone.0128901
Asadi M., Sadeghi A., Rezvanfar M. R., Talaie A., Rafiei F. (2016). The Genetics Study of Gestational Diabetes in Iranian Women and DIO2 Gene. Mol. Med. J. 2 (1), 43–47.
Bai Y., Tang L., Li L. (2020). The Roles of ADIPOQ Rs266729 and MTNR1B Rs10830963 Polymorphisms in Patients with Gestational Diabetes Mellitus: a Meta-Analysis. Gene 730, 144302. doi:10.1016/j.gene.2019.144302
Bartha J. L., Comino‐Delgado R., Romero-Carmona R., Gomez‐Jaen M. C. (2000). Sex Hormone‐binding Globulin in Gestational Diabetes. Acta Obstet. Gyn. Scan. 79 (10), 839–845. doi:10.1080/0001634000916921210.1034/j.1600-0412.2000.079010839.x
Bartosovic M., Molares H. C., Gregorova P., Hrossova D., Kudla G., Vanacova S. (2017). N6-methyladenosine Demethylase FTO Targets Pre-mRNAs and Regulates Alternative Splicing and 3′-end Processing. Nucleic Acids Res. 45 (19), 11356–11370. doi:10.1093/nar/gkx778
Bedford F. K., Ashworth A., Enver T., Wiedemann L. M. (1993). HEX: a Novel Homeobox Gene Expressed during Haematopoiesis and Conserved between Mouse and Human. Nucleic Acids Res. 21 (5), 1245–1249. doi:10.1093/nar/21.5.1245
Belmar C., Salinas P., Becker J., Abarzúa F., Olmos P., González P., et al. (2004). Incidencia de diabetes gestacional según distintos métodos diagnósticos y sus implicancias clínicas. Rev. Chil. Obstet. Ginecol. 69 (1), 2–7. doi:10.4067/S0717-75262004000100002
Ben-Haim M. S., Moshitch-Moshkovitz S., Rechavi G. (2015). FTO: Linking m6A Demethylation to Adipogenesis. Cell. Res. 25 (1), 3–4. doi:10.1038/cr.2014.162
Ben-Haroush A., Yogev Y., Hod M. (2004). Epidemiology of Gestational Diabetes Mellitus and its Association with Type 2 Diabetes. Diabet. Med. 21, 103–113. doi:10.1046/j.1464–5491.2003.00985.x10.1046/j.1464-5491.2003.00985.x
Bennink H. J., Schreurs W. H. (1975). Improvement of Oral Glucose Tolerance in Gestational Diabetes by Pyridoxine. Br. Med. J. 3 (5974), 13–15. doi:10.1136/bmj.3.5974.13
Berulava T., Horsthemke B. (2010). The Obesity-Associated SNPs in Intron 1 of the FTO Gene Affect Primary Transcript Levels. Eur. J. Hum. Genet. 18 (9), 1054–1056. doi:10.1038/ejhg.2010.71
Bomer N., den Hollander W., Ramos Y. F., Bos S. D., van der Breggen R., Lakenberg N., et al. (2015). Underlying Molecular Mechanisms of DIO2 Susceptibility in Symptomatic Osteoarthritis. Ann. Rheum. Dis. 74 (8), 1571–1579. doi:10.1136/annrheumdis-2013-204739
Buchanan T. A., Xiang A. H. (2005). Gestational Diabetes Mellitus. J. Clin. Invest. 115 (3), 485–491. doi:10.1172/JCI2453110.1172/jci200524531
Canani L. H., Capp C., Dora J. M., Meyer E. L. S., Wagner M. S., Harney J. W., et al. (2005). The Type 2 Deiodinase A/G (Thr92Ala) Polymorphism Is Associated with Decreased Enzyme Velocity and Increased Insulin Resistance in Patients with Type 2 Diabetes Mellitus. J. Clin. Endocrinol. Metab. 90 (6), 3472–3478. doi:10.1210/jc.2004-1977
Cao M., Zhang L., Chen T., Shi A., Xie K., Li Z., et al. (2020). Genetic Susceptibility to Gestational Diabetes Mellitus in a Chinese Population. Front. Endocrinol. 11, 247. doi:10.3389/fendo.2020.00247
Castro-Martínez A. G., Sánchez-Corona J., Vázquez-Vargas A. P., García-Zapién A. G., López-Quintero A., Villalpando-Velazco H. J., et al. (2018). Association Analysis of Calpain 10 Gene Variants/haplotypes with Gestational Diabetes Mellitus Among Mexican Women. Cell. Mol. Biol. 64 (3), 81–86. doi:10.14715/cmb/2018.64.3.13
Chang S., Wang Z., Wu L., Lu X., Shangguan S., Xin Y., et al. (2017). Association between TCF7L2 Polymorphisms and Gestational Diabetes Mellitus: A Meta‐analysis. J. Diabetes Investig. 8 (4), 560–570. doi:10.1111/jdi.12612
Chu H., Wang M., Zhong D., Shi D., Ma L., Tong N., et al. (2013). AdipoQ Polymorphisms Are Associated with Type 2 Diabetes Mellitus: a Meta‐analysis Study. Diabetes Metab. Res. Rev. 29 (7), 532–545. doi:10.1002/dmrr.2424
Churchill A. J., Carter J. G., Ramsden C., Turner S. J., Yeung A., Brenchley P. E., et al. (2008). VEGF Polymorphisms Are Associated with Severity of Diabetic Retinopathy. Invest. Ophthalmol. Vis. Sci. 49 (8), 3611–3616. doi:10.1167/iovs.07-1383
Coppotelli G., Summers A., Chidakel A., Ross J. M., Celi F. S. (2006). Functional Characterization of the 258 A/G (D2-ORFa-Gly3Asp) Human Type-2 Deiodinase Polymorphism: a Naturally Occurring Variant Increases the Enzymatic Activity by Removing a Putative Repressor Site in the 5′ UTR of the Gene. Thyroid 16 (7), 625–632. doi:10.1089/thy.2006.16.625
Coustan D. R. (2013). Gestational Diabetes Mellitus. Clin. Chem. 59 (9), 1310–1321. doi:10.1373/clinchem.2013.203331
Cui J., Xu X., Yin S., Chen F., Li P., Song C. (2016). Meta-analysis of the Association between Four CAPN10 Gene Variants and Gestational Diabetes Mellitus. Arch. Gynecol. Obstet. 294 (3), 447–453. doi:10.1007/s00404-016-4140-8
da Silva X. G., Loder M. K., McDonald A., Tarasov A. I., Carzaniga R., Kronenberger K., et al. (2009). TCF7L2 Regulates Late Events in Insulin Secretion from Pancreatic Islet Beta-Cells. Diabetes 58 (4), 894–905. doi:10.2337/db08-1187
Dabelea D., Hanson R. L., Lindsay R. S., Pettitt D. J., Imperatore G., Gabir M. M., et al. (2000). Intrauterine Exposure to Diabetes Conveys Risks for Type 2 Diabetes and Obesity: a Study of Discordant Sibships. Diabetes 49 (12), 2208–2211. doi:10.2337/diabetes.49.12.2208
Dahlgren A., Zethelius B., Jensevik K., Syvänen A. C., Berne C. (2007). Variants of the TCF7L2 Gene Are Associated with Beta Cell Dysfunction and Confer an Increased Risk of Type 2 Diabetes Mellitus in the ULSAM Cohort of Swedish Elderly Men. Diabetologia 50 (9), 1852. doi:10.1007/s00125-007-0746-5
Dalfrà M. G., Burlina S., Del Vescovo G. G., Lapolla A. (2020). Genetics and Epigenetics: New Insight on Gestational Diabetes Mellitus. Front. Endocrinol. 11, 936. doi:10.3389/fendo.2020.602477
Dayeh T. A., Olsson A. H., Volkov P., Almgren P., Rönn T., Ling C. (2013). Identification of CpG-SNPs Associated with Type 2 Diabetes and Differential DNA Methylation in Human Pancreatic Islets. Diabetologia 56, 1036–1046. doi:10.1007/s00125-012-2815-7
de Melo S. F., Frigeri H. R., dos Santos-Weiss I. C. R., Rea R. R., de Souza E. M., Alberton D., et al. (2015). Polymorphisms in FTO and TCF7L2 Genes of Euro-Brazilian Women with Gestational Diabetes. Clin. Biochem. 48 (16-17), 1064–1067. doi:10.1016/j.clinbiochem.2015.06.013
Dibbens J. A., Miller D. L., Damert A., Risau W., Vadas M. A., Goodall G. J. (1999). Hypoxic Regulation of Vascular Endothelial Growth Factor mRNA Stability Requires the Cooperation of Multiple RNA Elements. Mol. Biol. Cel. 10 (4), 907–919. doi:10.1091/mbc.10.4.907
Dimas A. S., Lagou V., Barker A., Knowles J. W., Mägi R., Hivert M. F., et al. (2014). Impact of Type 2 Diabetes Susceptibility Variants on Quantitative Glycemic Traits Reveals Mechanistic Heterogeneity. Diabetes 63 (6), 2158–2171. doi:10.2337/db13-0949
Ding E. L., Song Y., Manson J. E., Hunter D. J., Lee C. C., Rifai N., et al. (2009). Sex Hormone–Binding Globulin and Risk of Type 2 Diabetes in Women and Men. N. Engl. J. Med. 361 (12), 1152–1163. doi:10.1056/NEJMoa0804381
Ding M., Chavarro J., Olsen S., Lin Y., Ley S. H., Bao W., et al. (2018). Genetic Variants of Gestational Diabetes Mellitus: a Study of 112 SNPs Among 8722 Women in Two Independent Populations. Diabetologia 61 (8), 1758–1768. doi:10.1007/s00125-018-4637-8
Dishy V., Gupta S., Landau R., Xie H. G., Kim R. B., Smiley R. M., et al. (2003). G-protein β3 Subunit 825 C/T Polymorphism Is Associated with Weight Gain during Pregnancy. Pharmacogenet 13 (4), 241–242. doi:10.1097/00008571-200304000-00009
Dong P. P. (2019). Association of Vascular Endothelial Growth Factor Expression and Polymorphisms with the Risk of Gestational Diabetes Mellitus. J. Clin. Lab. Anal. 33 (2), e22686. doi:10.1002/jcla.22686
Dora J. M., Machado W. E., Rheinheimer J., Crispim D., Maia A. L. (2010). Association of the Type 2 Deiodinase Thr92Ala Polymorphism with Type 2 Diabetes: Case-Control Study and Meta-Analysis. Eur. J. Endocrinol. 163 (3), 427. doi:10.1530/EJE-10-0419
Estivalet A. A., Leiria L. B., Dora J. M., Rheinheimer J., Bouças A. P., Maia A. L., et al. (2011). D2 Thr92Ala and PPARγ2 Pro12Ala Polymorphisms Interact in the Modulation of Insulin Resistance in Type 2 Diabetic Patients. Obesity 19 (4), 825–832. doi:10.1038/oby.2010.231
Faal S., Abedi P., Jahanfar S., Ndeke J. M., Mohaghegh Z., Sharifipour F., et al. (2019). Sex Hormone Binding Globulin for Prediction of Gestational Diabetes Mellitus in Pre-conception and Pregnancy: A Systematic Review. Diabetes Res. Clin. Pract. 152, 39–52. doi:10.1016/j.diabres.2019.04.028
Fatima S. S., Chaudhry B., Khan T. A., Farooq S. (2016). KCNQ1 Rs2237895 Polymorphism Is Associated with Gestational Diabetes in Pakistani Women. Pak. J. Med. Sci. 32 (6), 1380–1385. doi:10.12669/pjms.326.11052
Feng C., Jin Z., Chi X., Zhang B., Wang X., Sun L., et al. (2018). SHBG Expression Is Correlated with PI3K/AKT Pathway Activity in a Cellular Model of Human Insulin Resistance. Gynecol. Endocrinol. 34 (7), 567–573. doi:10.1080/09513590.2017.1411474
Feng Y., Jiang C. D., Chang A. M., Shi Y., Gao J., Zhu L., et al. (2019). Interactions Among Insulin Resistance, Inflammation Factors, Obesity-Related Gene Polymorphisms, Environmental Risk Factors, and Diet in the Development of Gestational Diabetes Mellitus. J. Matern. Fetal Neonatal. Med. 32 (2), 339–347. doi:10.1080/14767058.2018.1446207
Fortunati N. (1999). Sex Hormone-Binding Globulin: Not Only a Transport Protein. What News Is Around the Corner? J. Endocrinol. Invest. 22 (3), 223–234. doi:10.1007/BF03343547
Frayling T. M., Timpson N. J., Weedon M. N., Zeggini E., Freathy R. M., Lindgren C. M., et al. (2007). A Common Variant in the FTO Gene Is Associated with Body Mass index and Predisposes to Childhood and Adult Obesity. Science 316 (5826), 889–894. doi:10.1126/science.1141634
Fritsche L., Sarief M., Wagner R., Stefan N., Lehmann R., Häring H. U., et al. (2018). Western Diet Triggers NLRP3-dependent Innate Immune Reprogramming. Diabetes Res. Clin. Pract. 146, 251–257. doi:10.1016/j.diabres.2018.11.003
Galton V. A., de Waard E., Parlow A. F., St Germain D. L., Hernandez A. (2014). Life without the Iodothyronine Deiodinases. Endocrinology 155 (10), 4081–4087. doi:10.1210/en.2014-1184
Garmendia M. L., Mondschein S., Montiel B., Kusanovic J. P. (2019). Trends and Predictors of Gestational Diabetes Mellitus in Chile. Int. J. Gynecol. Obstet. 148 (2), 210–218. In press. doi:10.1002/ijgo.13023
Gaulton K. J., Nammo T., Pasquali L., Simon J. M., Giresi P. G., Fogarty M. P., et al. (2010). A Map of Open Chromatin in Human Pancreatic Islets. Nat. Genet. 42 (3), 255–259. doi:10.1038/ng.530
Gjesing A. P., Kjems L. L., Vestmar M. A., Grarup N., Linneberg A., Deacon C. F., et al. (2011). Carriers of the TCF7L2 Rs7903146 TT Genotype Have Elevated Levels of Plasma Glucose, Serum Proinsulin and Plasma Gastric Inhibitory Polypeptide (GIP) during a Meal Test. Diabetologia 54 (1), 103–110. doi:10.1007/s00125-010-1940-4
Gong L. L., Liu H., Liu L. H. (2016). Relationship between Hypothyroidism and the Incidence of Gestational Diabetes: A Meta-Analysis. Taiwan. J. Obstet. Gyne. 55 (2), 171–175. doi:10.1016/j.tjog.2016.02.004
Griffin M. E., Coffey M., Johnson H., Scanlon P., Foley M., Stronge J., et al. (2000). Universal vs. Risk Factor‐based Screening for Gestational Diabetes Mellitus: Detection Rates, Gestation at Diagnosis and Outcome. Diabetic Med. 17 (1), 26–32. doi:10.1046/j.1464-5491.2000.00214.x
Hamidi A. K., Arzaghi S. M., Qorbani M., Khatami F., Ebrahimi M., Bandarian F., et al. (2019). MIF 173 G>C Variation Was Associated with Depressive Disorder in Type 2 Diabetes in an Iranian Population. Psychoneuroendocrino 104, 243–248. doi:10.1016/j.psyneuen.2019.03.011
HAPO Study Cooperative Research Group (2008). Hyperglycemia and Adverse Pregnancy Outcomes. N. Engl. J. Med. 358 (19), 1991–2002. doi:10.1056/NEJMoa0707943
HAPO Study Cooperative Research Group (2002). The Hyperglycemia and Adverse Pregnancy Outcome (HAPO) Study. Int. J. Gynecol. Obstet. 78 (1), 69–77. doi:10.1016/s0020-7292(02)00092-9
Hara K., Boutin P., Mori Y., Tobe K., Dina C., Yasuda K., et al. (2002). Genetic Variation in the Gene Encoding Adiponectin Is Associated with an Increased Risk of Type 2 Diabetes in the Japanese Population. Diabetes 51 (2), 536–540. doi:10.2337/diabetes.51.2.536
He H., Cao W., Zeng Y., Huang Z., Du W., Guan N., et al. (2018). Lack of Associations between the FTO Polymorphisms and Gestational Diabetes: A Meta-Analysis and Trial Sequential Analysis. Gene 677, 169–175. doi:10.1016/j.gene.2018.07.064
Hedderson M. M., Xu F., Darbinian J. A., Quesenberry C. P., Sridhar S., Kim C., et al. (2014). Prepregnancy SHBG Concentrations and Risk for Subsequently Developing Gestational Diabetes Mellitus. Diabetes Care. 37 (5), 1296–1303. doi:10.2337/dc13-1965
Hotta K., Nakata Y., Matsuo T., Kamohara S., Kotani K., Komatsu R., et al. (2008). Variations in the FTO Gene Are Associated with Severe Obesity in the Japanese. J. Hum. Genet. 53 (6), 546–553. doi:10.1007/s10038-008-0283-1
Hou Z., Li M., Cao Y. (2017). TCF7L2, CAPN10 Polymorphisms Are Associated with Gestational Diabetes Mellitus (GDM) Risks: a Meta-Analysis. Gynecol. Endocrinol. 33 (5), 399–404. doi:10.1080/09513590.2017.1290066
Hribal M. L., Federici M., Porzio O., Lauro D., Borboni P., Accili D., et al. (2000). The Gly→Arg972 Amino Acid Polymorphism in Insulin Receptor Substrate-1 Affects Glucose Metabolism in Skeletal Muscle Cells. J. Clin. Endocrinol. Metab. 85, 2004–2013. doi:10.1210/jc.85.5.200410.1210/jcem.85.5.6608
Hsiao T. J., Hwang Y., Liu C. H., Chang H. M., Lin E. (2013). Association of the C825T Polymorphism in the GNB3 Gene with Obesity and Metabolic Phenotypes in a Taiwanese Population. Genes Nutr. 8 (1), 137–144. doi:10.1007/s12263-012-0304-8
Huang L. T., Wu S. L., Liao X., Ma S. J., Tan H. Z. (2019). Adiponectin Gene Polymorphisms and Risk of Gestational Diabetes Mellitus: A Meta-Analysis. World J. Clin. Cases 7 (5), 572. doi:10.12998/wjcc.v7.i5.572
Huerta-Chagoya A., Vázquez-Cárdenas P., Moreno-Macías H., Tapia-Maruri L., Rodríguez-Guillén R., López-Vite E., et al. (2015). Genetic Determinants for Gestational Diabetes Mellitus and Related Metabolic Traits in Mexican Women. PLoS One 10 (5), e0126408. doi:10.1371/journal.pone.0126408
Huidobro A., Fulford A., Carrasco E. (2004). Incidencia de diabetes gestacional y su relación con obesidad en embarazadas chilenas. Rev. Med. Chil. 132 (8), 931–938. doi:10.4067/S0034-98872004000800004
Hussain M. K., Deli F. A., Algenabi A. H. A., Abdul-Rudha K. H. (2018). Adiponectin Gene Polymorphisms as a Predictor for Development of Type 2 Diabetes Mellitus in Iraqi Population. Gene 662, 118–122. doi:10.1016/j.gene.2018.03.087
Ip W., Chiang Y. A., Jin T. (2012). The Involvement of the Wnt Signaling Pathway and TCF7L2 in Diabetes Mellitus: The Current Understanding, Dispute, and Perspective. Cell. Biosci. 2, 28. doi:10.1186/2045-3701-2-28
Jonsson A., Isomaa B., Tuomi T., Taneera J., Salehi A., Nilsson P., et al. (2009). A Variant in the KCNQ1 Gene Predicts Future Type 2 Diabetes and Mediates Impaired Insulin Secretion. Diabetes 58 (10), 2409–2413. doi:10.2337/db09-0246
Kampmann U., Madsen L. R., Skajaa G. O., Iversen D. S., Moeller N., Ovesen P. (2015). Gestational Diabetes: a Clinical Update. World J. Diabetes 6 (8), 1065–1072. doi:10.4239/wjd.v6.i8.1065
Karbowska J., Kochan Z. (2006). Role of Adiponectin in the Regulation of Carbohydrate and Lipid Metabolism. J. Physiol. Pharmacol. 57 (Suppl. 6), 103–113.
Khan I. A., Movva S., Shaik N. A., Chava S., Jahan P., Mukkavali K. K., et al. (2014). Investigation of Calpain 10 (Rs2975760) Gene Polymorphism in Asian Indians with Gestational Diabetes Mellitus. Meta. Gene 2, 299–306. doi:10.1016/j.mgene.2014.03.001
Kinalski M., Telejko B., Kuźmicki M., Krętowski A., Kinalska I. (2005). Tumor Necrosis Factor Alpha System and Plasma Adiponectin Concentration in Women with Gestational Diabetes. Horm. Metab. Res. 37 (7), 450–454. doi:10.1055/s-2005-870238
Kuzmicki M., Telejko B., Zonenberg A., Szamatowicz J., Kretowski A., Nikolajuk A., et al. (2008). Circulating Pro-and Anti-inflammatory Cytokines in Polish Women with Gestational Diabetes. Horm. Metab. Res. 40 (8), 556–560. doi:10.1055/s-2008-1073166
Kwak S. H., Kim S. H., Cho Y. M., Go M. J., Cho Y. S., Choi S. H., et al. (2012). A Genome-wide Association Study of Gestational Diabetes Mellitus in Korean Women. Diabetes 61 (2), 531–541. doi:10.2337/db11-1034
Kwak S. H., Kim T. H., Cho Y. M., Choi S. H., Jang H. C., Park K. S. (2010). Polymorphisms in KCNQ1 Are Associated with Gestational Diabetes in a Korean Population. Horm. Res. Paediatr. 74 (5), 333–338. doi:10.1159/000313918
Lara-Castro C., Fu Y., Chung B. H., Garvey W. T. (2007). Adiponectin and the Metabolic Syndrome: Mechanisms Mediating Risk for Metabolic and Cardiovascular Disease. Curr. Opin. Lipidol. 18 (3), 263–270. doi:10.1097/MOL.0b013e32814a645f
Larrabure-Torrealva G. T., Martinez S., Luque-Fernandez M. A., Sanchez S. E., Mascaro P. A., Ingar H., et al. (2018). Prevalence and Risk Factors of Gestational Diabetes Mellitus: Findings from a Universal Screening Feasibility Program in Lima, Peru. BMC Pregnancy Childb 18 (1), 303. doi:10.1186/s12884-018-1904-0
Law K. P., Zhang H. (2017). The Pathogenesis and Pathophysiology of Gestational Diabetes Mellitus: Deductions from a Three-Part Longitudinal Metabolomics Study in China. Clin. Chim. Acta 468, 60–70. doi:10.1016/j.cca.2017.02.008
Lawlor D. A., Fraser A., Macdonald-Wallis C., Nelson S. M., Palmer T. M., Davey Smith G., et al. (2011). Maternal and Offspring Adiposity-Related Genetic Variants and Gestational Weight Gain. Am. J. Clin. Nutr. 94 (1), 149–155. doi:10.3945/ajcn.110.010751
Li C., Qiao B., Qi W., Zhan Y., Ma C., Zhao L., et al. (2016). Association of Macrophage Migration Inhibitory Factor Polymorphisms with Gestational Diabetes Mellitus in Han Chinese Women. Gynecol. Obstet. Invest. 81 (1), 84–89. doi:10.1159/000398796
Li H. P. (2013). Gestational Diabetes Induces Chronic Hypoxia Stress and Excessive Inflammatory Response in Murine Placenta. Int. J. Clin. Exp. Pathol. 6 (4), 650–659.
Li X., Li Y., Song B., Guo S., Chu S., Jia N., et al. (2012). Hematopoietically-expressed Homeobox Gene Three Widely-Evaluated Polymorphisms and Risk for Diabetes: a Meta-Analysis. PLoS One 7 (11), e49917. doi:10.1371/journal.pone.0049917
Lin P. C., Lin W. T., Yeh Y. H., Wung S. F. (2016). Transcription Factor 7-like 2 (TCF7L2) Rs7903146 Polymorphism as a Risk Factor for Gestational Diabetes Mellitus: a Meta-Analysis. PLoS One 11 (4), e0153044. doi:10.1371/journal.pone.0153044
Lin Z., Wang Y., Zhang B., Jin Z. (2018). Association of Type 2 Diabetes Susceptible Genes GCKR, SLC30A8, and FTO Polymorphisms with Gestational Diabetes Mellitus Risk: a Meta-Analysis. Endocrine 62 (1), 34–45. doi:10.1007/s12020-018-1651-z
Liu X. H., Xie C. G., An Y., Zhang X. X., Wu W. B. (2015). Meta-Analysis of the Association between the Rs7903146 Polymorphism at the TCF7l2 Locus and Type 2 Diabetes Mellitus Susceptibility. Genet. Mol. Res. 14 (4), 16856–16862. doi:10.4238/2015.December.14.12
Loubiere L. S., Vasilopoulou E., Bulmer J. N., Taylor P. M., Stieger B., Verrey F., et al. (2010). Expression of Thyroid Hormone Transporters in the Human Placenta and Changes Associated with Intrauterine Growth Restriction. Placenta 31 (4), 295–304. doi:10.1016/j.placenta.2010.01.013
Lowe W. L., Scholtens D. M., Sandler V., Hayes M. G. (2016). Genetics of Gestational Diabetes Mellitus and Maternal Metabolism. Curr. Diab. Rep. 16 (2), 15. doi:10.1007/s11892-015-0709-z
Lu Y., Zhou Y., Tian W. (2013). Combining Hi-C Data with Phylogenetic Correlation to Predict the Target Genes of Distal Regulatory Elements in Human Genome. Nucleic Acids Res. 41, 10391–10402. doi:10.1093/nar/gkt785
Lyssenko V., Lupi R., Marchetti P., Del Guerra S., Orho-Melander M., Almgren P., et al. (2007). Mechanisms by Which Common Variants in the TCF7L2 Gene Increase Risk of Type 2 Diabetes. J. Clin. Invest. 117 (8), 2155–2163. doi:10.1172/JCI30706
Mack L. R., Tomich P. G. (2017). Gestational Diabetes Obstet. Gynecol. Clin. North. Am. 44 (2), 207–217. doi:10.1016/j.ogc.2017.02.002
Martins M. C., Trujillo J., Farias D. R., Struchiner C. J., Kac G. (2016). Association of the FTO (Rs9939609) and MC4R (Rs17782313) Gene Polymorphisms with Maternal Body Weight during Pregnancy. Nutrition 32 (11-12), 1223–1230. doi:10.1016/j.nut.2016.04.009
McGettrick A. J., Feener E. P., Kahn R. R. (2005). Human Insulin Receptor Substrate-1 (IRS-1) Polymorphism G972R Causes IRS-1 to Associate with the Insulin Receptor and Inhibit Receptor Autophosphorylation. J. Biol. Chem. 280, 6441–6446. doi:10.1074/jbc.M412300200
Mejia-Montilla J., Álvarez-Mon M., Reyna-Villasmil E., Torres-Cepeda D., Santos-Bolívar J., Reyna-Villasmil N., et al. (2015). Macrophage Migration Inhibitory Factor in Obese and Non Obese Women with Polycystic Ovary Syndrome. Endocrinol. Nutr. 62 (1), 31–37. doi:10.1016/j.endonu.2014.09.00510.1016/j.endoen.2014.09.009
Merkestein M., Laber S., McMurray F., Andrew D., Sachse G., Sanderson J., et al. (2015). FTO Influences Adipogenesis by Regulating Mitotic Clonal Expansion. Nat. Commun. 6, 6792. doi:10.1038/ncomms7792
Mierzyński R., Dłuski D., Nowakowski Ł., Poniedziałek-Czajkowska E., Leszczyńska-Gorzelak B. (2018). Adiponectin and Omentin Levels as Predictive Biomarkers of Preterm Birth in Patients with Gestational Diabetes Mellitus. Biomed. Res. Int. 2018, 1–9. doi:10.1155/2018/7154216
Moldovan S., Atiya A., Adrian T. E., Kleinman R. M., Lloyd K., Olthoff K., et al. (1995). Somatostatin Inhibits B-Cell Secretion via a Subtype-2 Somatostatin Receptor in the Isolated Perfused Human Pancreas. J. Surg. Res. 59 (1), 85–90. doi:10.1006/jsre.1995.1136
Morris J. F., Hromas R., Rauscher F. J. (1994). Characterization of the DNA-Binding Properties of the Myeloid Zinc-finger Protein Mzf1–2 Independent DNA-Binding Domains Recognize 2 DNA Consensus Sequences with a Common G-Rich Core. Mol. Cel. Biol. 14 (3), 1786–1795. doi:10.1128/mcb.14.3.178610.1128/mcb.14.3.1786-1795.1994
Neuhaus T., Graf C., Stier S., Knapp M., Grunewald E., Ko Y.-D., et al. (2013). Variation in the Calpain-10 Gene Is Not Associated with Gestational Diabetes Mellitus. Scand. J. Clin. Lab. Invest. 74 (1), 59–66. doi:10.3109/00365513.2013.857427
Oxenkrug G. F. (2015). Increased Plasma Levels of Xanthurenic and Kynurenic Acids in Type 2 Diabetes. Mol. Neurobiol. 52 (2), 805–810. doi:10.1007/s12035-015-9232-0
Pantham P., Aye I. L. H., Powell T. L. (2015). Inflammation in Maternal Obesity and Gestational Diabetes Mellitus. Placenta 36 (7), 709–715. doi:10.1016/j.placenta.2015.04.006
Pivovarova O., Nikiforova V. J., Pfeiffer A. F., Rudovich N. (2009). The Influence of Genetic Variations in HHEX Gene on Insulin Metabolism in the German MESYBEPO Cohort. Diabetes Metab. Res. Rev. 25 (2), 156–162. doi:10.1002/dmrr.926
Plows J. F., Stanley J. L., Baker P. N., Reynolds C. M., Vickers M. H. (2018). The Pathophysiology of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 19 (11), 3342. doi:10.3390/ijms19113342
Ramírez-Bello J., Jiménez-Morales M. (2017). Implicaciones funcionales de los polimorfismos de un solo nucleótido (SNP) en genes codificantes de proteínas y no codificantes en enfermedades multifactoriales [Functional implications of single nucleotide polymorphisms (SNPs) in protein-coding and non-coding RNA genes in multifactorial diseases]. Gaceta Med. de Mexico 153 (2), 238–250.
Rizvi S., Raza S. T., Rahman Q., Mahdi F. (2016). Role of GNB3, NET, KCNJ11, TCF7L2 and GRL Genes Single Nucleotide Polymorphism in the Risk Prediction of Type 2 Diabetes Mellitus. 3 Biotech. 6 (2), 255. doi:10.1007/s13205-016-0572-x
Rochford J. J., Semple R. K., Laudes M., Boyle K. B., Christodoulides C., Mulligan C., et al. (2004). ETO/MTG8 Is an Inhibitor of C/EBPβ Activity and a Regulator of Early Adipogenesis. Mol. Cel. Biol. 24 (22), 9863–9872. doi:10.1128/MCB.24.22.9863-9872.2004
Saavedra J. S., Zúñiga L. F., Freyre S. I., Muñoz G. W., Salguero C. (2017). El rol de VEGF en la angiogénesis fisiológica y tumoral. Med 39 (3), 190–209.
Saber-Ayad M., Manzoor S., El Serafi A., Mahmoud I., Hammoudeh S., Rani A., et al. (2019). The FTO Rs9939609 “A” Allele Is Associated with Impaired Fasting Glucose and Insulin Resistance in Emirati Population. Gene 681, 93–98. doi:10.1016/j.gene.2018.09.053
Salazar L. A., Colivoro K., Díaz A., Sepúlveda S., Cuevas A., Saavedra N., et al. (2010). Asociación del polimorfismo rs2241766 del gen de la adiponectina y enfermedad arterial coronaria en individuos del sur de Chile. Rev. Chil. Cardiol. 29 (2), 214–220. doi:10.4067/S0718-85602010000200007
Saucedo R., Valencia J., Gutierrez C., Basurto L., Hernandez M., Puello E., et al. (2017). Gene Variants in the FTO Gene Are Associated with Adiponectin and TNF-Alpha Levels in Gestational Diabetes Mellitus. Diabetol. Metab. Syndr. 9 (1), 1–7. doi:10.1186/s13098-017-0234-0
Savic D., Ye H., Aneas I., Park S. Y., Bell G. I., Nobrega M. A. (2011). Alterations in TCF7L2 Expression Define its Role as a Key Regulator of Glucose Metabolism. Genome Res. 21 (9), 1417–1425. doi:10.1101/gr.123745.111
Saxena R., Gianniny L., Burtt N. P., Lyssenko V., Giuducci C., Sjögren M., et al. (2006). Common Single Nucleotide Polymorphisms in TCF7L2 Are Reproducibly Associated with Type 2 Diabetes and Reduce the Insulin Response to Glucose in Nondiabetic Individuals. Diabetes 55 (10), 2890–2895. doi:10.2337/db06-0381
Schinner S., Ülgen F., Papewalis C., Schott M., Woelk A., Vidal-Puig A., et al. (2008). Regulation of Insulin Secretion, Glucokinase Gene Transcription and Beta Cell Proliferation by Adipocyte-Derived Wnt Signalling Molecules. Diabetologia 51 (1), 147–154. doi:10.1007/s00125-007-0848-0
Schneider S., Bock C., Wetzel M., Maul H., Loerbroks A. (2012). The Prevalence of Gestational Diabetes in Advanced Economies. J. Perinat. Med. 40 (5), 511–552. doi:10.1515/jpm-2012-0015
Sentinelli F., Filippi E., Cavallo M. G., Romeo S., Fanelli M., Baroni M. G. (2006). The G972R Variant of the Insulin Receptor Substrate-1 Gene Impairs Insulin Signaling and Cell Differentiation in 3T3L1 Adipocytes; Treatment with a PPARγ Agonist Restores normal Cell Signalling and Differentiation. J. Endocrinol. 188, 271–285. doi:10.1677/joe.1.06290
Shaat N., Ekelund M., Lernmark Å., Ivarsson S., Almgren P., Berntorp K., et al. (2005). Association of the E23K Polymorphism in the KCNJ11 Gene with Gestational Diabetes Mellitus. Diabetologia 48 (12), 2544–2551. doi:10.1007/s00125-005-0035-0
Shaat N., Lernmark Å., Karlsson E., Ivarsson S., Parikh H., Berntorp K., et al. (2007). A Variant in the Transcription Factor 7-like 2 (TCF7L2) Gene Is Associated with an Increased Risk of Gestational Diabetes Mellitus. Diabetologia 50 (5), 972–979. doi:10.1007/s00125-007-0623-2
Shah A., Stotland N. E., Cheng Y. W., Ramos G. A., Caughey A. B. (2011). The Association between Body Mass index and Gestational Diabetes Mellitus Varies by Race/ethnicity. Am. J. Perinat. 28 (7), 515–520. doi:10.1055/s-0031-1272968
Shen Y., Jia Y., Li Y., Gu X., Wan G., Zhang P., et al. (2020). Genetic Determinants of Gestational Diabetes Mellitus: a Case–Control Study in Two Independent Populations. Acta Diabetol. 57 (7), 843–852. doi:10.1007/s00592-020-01485-w
Shin H. D., Park B. L., Shin H. J., Kim J. Y., Park S., Kim B., et al. (2010). Association of KCNQ1 Polymorphisms with the Gestational Diabetes Mellitus in Korean Women. J. Clin. Endocrinol. Metab. 95 (1), 445–449. doi:10.1210/jc.2009-1393
Shu L., Sauter N. S., Schulthess F. T., Matveyenko A. V., Oberholzer J., Maedler K. (2008). Transcription Factor 7-like 2 Regulates β-cell Survival and Function in Human Pancreatic Islets. Diabetes 57 (3), 645–653. doi:10.2337/db07-0847
Siffert W., Forster P., Jöckel K. H., Mvere D. A., Brinkmann B., Naber C., et al. (1999). Worldwide Ethnic Distribution of the G Protein β3 Subunit 825T Allele and its Association with Obesity in Caucasian, Chinese, and Black African Individuals. J. Am. Soc. Nephrol. 10 (9), 1921–1930. doi:10.1681/asn.v1091921
Sonagra A. D., Biradar S. M., Dattatreya K., Murthy D. S. J. (2014). Normal Pregnancy - a State of Insulin Resistance. J. Clin. Diagn. Res. 8 (11), CC01–3. doi:10.7860/JCDR/2014/10068.5081
Soyal S. M., Felder T., Auer S., Oberkofler H., Iglseder B., Paulweber B., et al. (2015). Associations of Haplotypes Upstream of IRS1 with Insulin Resistance, Type 2 Diabetes, Dyslipidemia, Preclinical Atherosclerosis, and Skeletal Muscle LOC646736 mRNA Levels. J. Diabetes Res. 2015, 405371. doi:10.1155/2015/405371
Speakman J. R. (2015). The ‘Fat Mass and Obesity Related’ (FTO) Gene: Mechanisms of Impact on Obesity and Energy Balance. Curr. Obes. Rep. 4 (1), 73–91. doi:10.1007/s13679-015-0143-1
Stratigopoulos G., Padilla S. L., LeDuc C. A., Watson E., Hattersley A. T., McCarthy M. I., et al. (2008). Regulation of Fto/Ftm Gene Expression in Mice and Humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 294 (4), R1185–R1196. doi:10.1152/ajpregu.00839.2007
Tahara T., Arisawa T., Shibata T., Nakamura M., Yamashita H., Yoshioka D., et al. (2009). Effect of Polymorphisms in the 3′-untranslated Region (3′-UTR) of VEGF Gene on Gastric Pre-malignant Condition. Anticancer. Res. 29 (2), 485–489.
Takhshid M. A., Haem Z., Aboualizadeh F. (2015). The Association of Circulating Adiponectin and + 45 T/G Polymorphism of Adiponectin Gene with Gestational Diabetes Mellitus in Iranian Population. J. Diabetes Metab. Disord. 14 (1), 30. doi:10.1186/s40200-015-0156-z
Tarnowski M., Bujak J., Kopytko P., Majcher S., Ustianowski P., Dziedziejko V., et al. (2018). Effect of FTO and IGF2BP2 Gene Polymorphisms on Duration of Pregnancy and Apgar Scores in Women with Gestational Diabetes. J. Obstet. Gynaecol. 39 (2), 1–6. doi:10.1080/01443615.2018.1502263
Tarnowski M., Malinowski D., Safranow K., Dziedziejko V., Czerewaty M., Pawlik A. (2017). Hematopoietically Expressed Homeobox (HHEX) Gene Polymorphism (Rs5015480) Is Associated with Increased Risk of Gestational Diabetes Mellitus. Clin. Genet. 91 (6), 843–848. doi:10.1111/cge.12875
Tawfeek M. A., Alfadhli E. M., Alayoubi A. M., El-Beshbishy H. A., Habib F. A. (2017). Sex Hormone Binding Globulin as a Valuable Biochemical Marker in Predicting Gestational Diabetes Mellitus. BMC. Womens Health 17 (1), 18. doi:10.1186/s12905-017-0373-3
Troncoso F., Acurio J., Herlitz K., Aguayo C., Bertoglia P., Guzman-Gutierrez E., et al. (2017). Gestational Diabetes Mellitus Is Associated with Increased Pro-migratory Activation of Vascular Endothelial Growth Factor Receptor 2 and Reduced Expression of Vascular Endothelial Growth Factor Receptor 1. PLoS One 12 (8), e0182509. doi:10.1371/journal.pone.0182509
Ustianowski P., Malinowski D., Kopytko P., Czerewaty M., Tarnowski M., Dziedziejko V., et al. (2021). ADCY5, CAPN10 and JAZF1 Gene Polymorphisms and Placental Expression in Women with Gestational Diabetes. Life 11 (8), 806. doi:10.3390/life11080806
Voight B. F., Scott L. J., Steinthorsdottir V., Morris A. P., Dina C., Welch R. P., et al. (2010). Twelve Type 2 Diabetes Susceptibility Loci Identified through Large-Scale Association Analysis. Nat. Genet. 42, 579–589. doi:10.1038/ng.609
Vounzoulaki E., Khunti K., Abner S. C., Tan B. K., Davies M. J., Gillies C. L. (2020). Progression to Type 2 Diabetes in Women with a Known History of Gestational Diabetes: Systematic Review and Meta-Analysis. BMJ (Clinical research ed.) 369, m1361. doi:10.1136/bmj.m1361
Wang H., Miao K., Zhao J., Liu L., Cui G., Chen C., et al. (2013). Common Variants in KCNQ1 Confer Increased Risk of Type 2 Diabetes and Contribute to the Diabetic Epidemic in East Asians: A Replication and Meta‐Analysis. Ann. Hum. Genet. 77 (5), 380–391. doi:10.1111/ahg.12029
Wang K., Chen Q., Feng Y., Yang H., Wu W., Zhang P., et al. (2019). Single Nucleotide Polymorphisms in CDKAL1 Gene Are Associated with Risk of Gestational Diabetes Mellitus in Chinese Population. J. Diabetes Res. 2019, 3618103. doi:10.1155/2019/3618103
Wang X., Ding Y., Zhang X., Rao J., Yu H., Pan H. (2020). The Association between HHEX Single-Nucleotide Polymorphism Rs5015480 and Gestational Diabetes Mellitus: A Meta-Analysis. Medicine 99 (12). doi:10.1097/MD.0000000000019478
Wang X., Yang T., Miao J., Liu H., Wu K., Guo J., et al. (2018). Correlation between Maternal and Fetal Insulin Resistance in Pregnant Women with Gestational Diabetes Mellitus. Clin. Lab. 64 (6), 945–953. doi:10.7754/Clin.Lab.2018.171214
Wu L., Cui L., Tam W. H., Ma R. C., Wang C. C. (2016). Genetic Variants Associated with Gestational Diabetes Mellitus: a Meta-Analysis and Subgroup Analysis. Sci. Rep. 6, 30539. doi:10.1038/srep30539
Wu T. S. (2015). Functional Characterization of Sex Hormone-Binding Globulin Genetic Polymorphism. Vancouver: University of British Columbia. [Doctoral dissertation].
Xu F., Zhang H., Qi H. (2016). No Association of Adiponectin + 45 T/G Polymorphism with the Risk of Gestational Diabetes Mellitus: Evidence from a Meta-Analysis. J. Renin. Angiotensin. Aldosterone. Syst. 17 (2), 1–6. doi:10.1177/1470320316653283
Yang S., Shi F. T., Leung P. C., Huang H. F., Fan J. (2016). Low Thyroid Hormone in Early Pregnancy∫ Is Associated with an Increased Risk of Gestational Diabetes Mellitus. J. Clin. Endocrinol. Metab. 101 (11), 4237–4243. doi:10.1210/jc.2016-1506
Yasuda K., Miyake K., Horikawa Y., Hara K., Osawa H., Furuta H., et al. (2008). Variants in KCNQ1 Are Associated with Susceptibility to Type 2 Diabetes Mellitus. Nat. Genet. 40 (9), 1092. doi:10.1038/ng.207
Zajdenverg L., Negrato C. A. (2017). Gestational Diabetes Mellitus and Type 2 Diabetes: Same Disease in a Different Moment of Life? Maybe Not. Arch. Endocrinol. Metab. 61 (3), 208–210. doi:10.1590/2359-3997000000276
Zhan Y., Li C., Chen J., Yu S., Gao Q., Wang Y. P., et al. (2015). Association between Macrophage Migration Inhibitory Factor Rs1007888 and GDM. Genet. Mol. Res. 14 (1), 797–804. doi:10.4238/2015.February.2.4
Zhang B., Jin Z., Sun L., Zheng Y., Jiang J., Feng C., et al. (2016). Expression and Correlation of Sex Hormone-Binding Globulin and Insulin Signal Transduction and Glucose Transporter Proteins in Gestational Diabetes Mellitus Placental Tissue. Diabetes Res. Clin. Pract. 119, 106–117. doi:10.1016/j.diabres.2016.07.003
Zhang C., Bao W., Rong Y., Yang H., Bowers K., Yeung E., et al. (2013). Genetic Variants and the Risk of Gestational Diabetes Mellitus: A Systematic Review. Hum. Reprod. Update 19 (4), 376–390. doi:10.1093/humupd/dmt013
Zhang J., McKenna L. B., Bogue C. W., Kaestner K. H. (2014b). The Diabetes Gene Hhex Maintains δ-cell Differentiation and Islet Function. Genes. Dev. 28 (8), 829–834. doi:10.1101/gad.235499.113
Zhang M., Zhang Y., Ma J., Guo F., Cao Q., Zhang Y., et al. (2015). The Demethylase Activity of FTO (Fat Mass and Obesity Associated Protein) Is Required for Preadipocyte Differentiation. PLoS One 10 (7), e0133788. doi:10.1371/journal.pone.0133788
Zhang X., Shi C., Wei L., Sun F., Ji L. (2019). The Association between the Rs2975760 and Rs3792267 Single Nucleotide Polymorphisms of Calpain 10 (CAPN10) and Gestational Diabetes Mellitus. Med. Sci. Monit. 25, 5137. doi:10.12659/MSM.914930
Zhang Y., Sun C. M., Hu X. Q., Zhao Y. (2014a). Relationship between Melatonin Receptor 1B and Insulin Receptor Substrate 1 Polymorphisms with Gestational Diabetes Mellitus: A Systematic Review and Meta-Analysis. Sci. Rep. 4, 1–7. doi:10.1038/srep06113
Zhao P., Liu E., Qiao Y., Katzmarzyk P. T., Chaput J. P., Fogelholm M., et al. (2016). Maternal Gestational Diabetes and Childhood Obesity at Age 9–11: Results of a Multinational Study. Diabetologia 59 (11), 2339–2348. doi:10.1007/s00125-016-4062-9
Zhao W., Rasheed A., Tikkanen E., Lee J. J., Butterworth A. S., Howson J. M. M., et al. (2017). Identification of New Susceptibility Loci for Type 2 Diabetes and Shared Etiological Pathways with Coronary Heart Disease. Nat. Genet. 49, 1450–1457. doi:10.1038/ng.3943
Zhao X., Yang Y., Sun B. F., Shi Y., Yang X., Xiao W., et al. (2014a). FTO-dependent Demethylation of N6-Methyladenosine Regulates MRNA Splicing and Is Required for Adipogenesis. Cel. Res. 24 (12), 1403–1419. doi:10.1038/cr.2014.151
Zhao X., Yang Y., Sun B. F., Zhao Y. L., Yang Y. G. (2014b). FTO and Obesity: Mechanisms of Association. Curr. Diab. Rep. 14 (5), 486. doi:10.1007/s11892-014-0486-0
Zheng J. S., Arnett D. K., Parnell L. D., Smith C. E., Li D., Borecki I. B., et al. (2013). Modulation by Dietary Fat and Carbohydrate of IRS1 Association with Type 2 Diabetes Traits in Two Populations of Different Ancestries. Diabetes Care 36, 2621–2627. doi:10.2337/dc12-2607
Zhou Y., Park S. Y., Su J., Bailey K., Ottosson-Laakso E., Shcherbina L., et al. (2014). TCF7L2 Is a Master Regulator of Insulin Production and Processing. Hum. Mol. Genet. 23 (24), 6419–6431. doi:10.1093/hmg/ddu359
Keywords: gestational diabetes mellitus, single nucleotide polymorphism, insulin resistance, genetic risk factors, insulin signaling dysfunction
Citation: Ortega-Contreras B, Armella A, Appel J, Mennickent D, Araya J, González M, Castro E, Obregón AM, Lamperti L, Gutiérrez J and Guzmán-Gutiérrez E (2022) Pathophysiological Role of Genetic Factors Associated With Gestational Diabetes Mellitus. Front. Physiol. 13:769924. doi: 10.3389/fphys.2022.769924
Received: 03 September 2021; Accepted: 15 March 2022;
Published: 04 April 2022.
Edited by:
Carlos Galaviz-Hernandez, Instituto Politécnico Nacional (IPN), MexicoReviewed by:
Kuanfeng Xu, Nanjing Medical University, ChinaColm J. McElwain, University College Cork, Ireland
Copyright © 2022 Ortega-Contreras, Armella, Appel, Mennickent, Araya, González, Castro, Obregón, Lamperti, Gutiérrez and Guzmán-Gutiérrez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: E. Guzmán-Gutiérrez, eguzman@udec.cl