 Zahra Ghahremani
Zahra Ghahremani Nuria Escudero
Nuria Escudero Ester Saus
Ester Saus Toni Gabaldón
Toni Gabaldón F. Javier Sorribas
F. Javier Sorribas- 1Departament d’Enginyeria Agroalimentària i Biotecnologia, Universitat Politècnica de Catalunya, Barcelona, Spain
- 2Bioinformatics and Genomics Programs, Centre for Genomic Regulation (CRG), Barcelona Institute of Science and Technology, Barcelona, Spain
- 3Department of Experimental and Health Sciences, Universitat Pompeu Fabra (UPF), Barcelona, Spain
- 4ICREA, Barcelona, Spain
Meloidogyne spp. are the most damaging plant parasitic nematodes for horticultural crops worldwide. Pochonia chlamydosporia is a fungal egg parasite of root-knot and cyst nematodes able to colonize the roots of several plant species and shown to induce plant defense mechanisms in fungal-plant interaction studies, and local resistance in fungal-nematode-plant interactions. This work demonstrates the differential ability of two out of five P. chlamydosporia isolates, M10.43.21 and M10.55.6, to induce systemic resistance against M. incognita in tomato but not in cucumber in split-root experiments. The M10.43.21 isolate reduced infection (32–43%), reproduction (44–59%), and female fecundity (14.7–27.6%), while the isolate M10.55.6 only reduced consistently nematode reproduction (35–47.5%) in the two experiments carried out. The isolate M10.43.21 induced the expression of the salicylic acid pathway (PR-1 gene) in tomato roots 7 days after being inoculated with the fungal isolate and just after nematode inoculation, and at 7 and 42 days after nematode inoculation too. The jasmonate signaling pathway (Lox D gene) was also upregulated at 7 days after nematode inoculation. Thus, some isolates of P. chlamydosporia can induce systemic resistance against root-knot nematodes but this is plant species dependent.
Introduction
The root-knot nematodes (RKN), Meloidogyne spp., are obligate parasites of plants. The genus comprises more than 100 species, but only four of them are considered the most damaging plant parasitic nematodes due to its wide range of plant hosts, worldwide distribution, and high reproductive capacity (Jones et al., 2013). The RKN infective juveniles (J2) enter the root near the elongation zone and migrate intercellularly to establish a permanent feeding site into the vascular cylinder, inducing the formation of giant cells and root galls by affecting cell wall architecture, plant development, defenses, and metabolism (Shukla et al., 2018). Once the infection occurs, J2 become sedentary, and molt three times to achieve the mature adult female stage. The most frequent and damaging tropical species, M. arenaria, M. incognita, and M. javanica, reproduce parthenogenetically. The female lays a large number of eggs in a gelatinous matrix, the egg mass, located on the surface or within the galled roots.
The damage potential of some Meloidogyne species has been summarized (Greco and Di Vito, 2009; Wesemael et al., 2011). M. arenaria, M. incognita and M. javanica are responsible for the majority of vegetable yield losses caused by plant parasitic nematodes (Sikora and Fernández, 2005). Among vegetables, those belonging to the solanaceae and cucurbitaceae families are commonly included in rotation schemes because they are economically important for growers. The estimation of maximum crop yield losses caused by the nematode in field and plastic greenhouse cultivation varies according to the plant germplasm, environmental conditions and agronomic practices. For instances, maximum yield losses from 62 to 100% have been reported in susceptible tomato cultivars (Seid et al., 2015; Giné and Sorribas, 2017), 30 to 60% in aubergine (Sikora and Fernández, 2005), 50% in cantaloupe (Sikora and Fernández, 2005), 37 to 50% in watermelon (Sikora and Fernández, 2005; López-Gómez et al., 2014), and 88% in non-grafted or grafted cucumber on Cucurbita hybrid rootstocks (Giné et al., 2014, 2017). Control of RKN is conducted mainly with fumigant and non-fumigant nematicides (Djian-Caporalino, 2012; Talavera et al., 2012). However, due to environmental and toxicological concerns, some legislative regulations, such as the European Directive 2009/128/EC, aim to reduce the use of pesticides by promoting alternative methods such as biological control and plant resistance.
Several nematode antagonists belonging to different taxonomic groups have been described (Stirling, 2014). They can act in three ways: (1) directly parasitizing several or specific RKN development stages, such as Pasteuria penetrans (Davies et al., 2011); (2) indirectly by repelling, immobilizing and/or killing them by means of metabolites, and/or inducing plant response, such as Fusarium oxysporum strain Fo162 (Dababat and Sikora, 2007a,b); or (3) both directly and indirectly, such as Trichoderma atroviride strain T11 or Trichoderma harzianum strain T-78 (de Medeiros et al., 2017; Martínez-Medina et al., 2017). Pochonia chlamydosporia (syn. Metacordyceps chlamydosporia) is a fungal antagonist of RKN and cyst nematodes that acts directly by parasitizing eggs, and could also acts indirectly. This fungal species colonizes endophytically the root of several plants, including barley (Maciá-Vicente et al., 2009), tomato (Bordallo et al., 2002), potato (Manzanilla-López et al., 2011), or Arabidopsis (Zavala-Gonzalez et al., 2017), inducing plant defense mechanisms, such as the formation of papillae (Bordallo et al., 2002) and the modulation of miRNA in tomato (Pentimone et al., 2018) or plant defense genes related to salicylic acid and jasmonic acid pathways in barley and Arabidopsis (Larriba et al., 2015; Zavala-Gonzalez et al., 2017). It is assumed that some of these defense mechanisms could suppress root infection, development and/or reproduction of RKN (Escudero and Lopez-Llorca, 2012), but as far we know, only one study has proven the induction of local resistance (de Medeiros et al., 2015), and none to elucidate the capability of this fungal species to induce systemic resistance. As P. chlamydosporia parasitizes RKN eggs, a split-root system is required to determine the capability of this nematophagous fungus to induce plant resistance avoiding the direct interaction with the nematode. Therefore, in the present study, the capability of five P. chlamydosporia isolates to induce plant resistance against M. incognita was assessed in a split-root system. To assess whether the response was plant dependent, tomato and cucumber were used as representatives of solanaceous and cucurbit crops frequently including in rotation schemes.
Materials and Methods
Plant Material, Nematode, and Fungi
Tomato cv. Durinta and cucumber cv. Dasher II were used in this study. For all the experiments, seeds were surface-sterilized in a 50% sterilized bleach solution (35 g L−1 active chlorine) for 2 min, washed three times in sterilized distilled water for 10 s each, sown in a tray containing sterile vermiculite, and maintained in a growth chamber at 25°C ± 2°C with a 16 h:8 h (light:dark) photoperiod.
Five P. chlamydosporia isolates M10.41.42, M10.43.21, M10.51.3, M10.55.6, and M10.62.2 were used. The fungal isolates were obtained from horticultural commercial growing sites in northeastern Spain from RKN eggs (Giné et al., 2012) and maintained as single-spore isolates at the Universitat Politècnica de Catalunya. Fungal chlamydospores were produced in barley seeds following the procedure of Becerra and collaborators (Becerra Lopez‐Lavalle et al., 2012) with some modifications. Briefly, for each isolate, three 200 g batches of barley seeds were soaked for 18 h and each batch sterilized in an Erlenmeyer flask at 121°C for 22 min over two consecutive days, then were incubated at 25°C ± 2°C in the dark. Afterward, 10 5-mm plugs from the edge of each P. chlamydosporia isolate grown in CMA were added to each Erlenmeyer flask and they were shaken at 5-day intervals to homogenize fungal growth. After a month, the number of chlamydospores produced on barley was determined following the procedure of Kerry and Bourne (2002). Three 10-seed subsamples per Erlenmeyer were plated onto CMA and incubated at 25 ± 2°C in the dark for 2 weeks to assess putative contaminations prior to being used. The viability of the chlamydospores was assessed as in Escudero et al. (2017).
J2 of the isolate Agropolis of Meloidogyne incognita were used as inoculum. Eggs were extracted from tomato roots by blender maceration in a 5% commercial bleach (40 g L−1 NaOCl) solution for 5 min (Hussey and Barker, 1973). The egg suspension was passed through a 74-μm aperture sieve to remove root debris, and eggs were collected on a 25-μm sieve and placed on Baermann trays (Whitehead and Hemming, 1965) at 25 ± 2°C. Nematodes were collected daily using a 25-μm sieve for 7 days and stored at 9°C until their use.
Induction of Systemic Plant Resistance by P. chlamydosporia Isolates Against Root-Knot Nematodes
Tomato and cucumber were grown in a split-root system as described in previous studies (de Medeiros et al., 2017; Martínez-Medina et al., 2017). In this system, the root is divided into two halves transplanted in two adjacent pots: the inducer, inoculated with the antagonist, and the responder, inoculated with the nematode. Briefly, the main root of 5-day-old seedlings was excised and plantlets were individually transplanted in seedling trays containing sterile vermiculite and maintained under the same conditions for 2 weeks for cucumber, and 3 weeks for tomato plants. Afterward, plantlets were transferred to the split-root system by splitting roots into two halves planted in two adjacent 200 cm3 pots filled with sterilized sand. Four treatments were assessed for each fungal isolate: Fungi-RKN and None-RKN, to assess the capability of each fungal isolate to induce plant response against RKN, and Fungi-None and None-None, to assess the effect of each fungal isolate on plant growth. Each treatment was replicated 10 times, and the experiment was conducted two times. The inducer part of the root of the treatments containing Fungi was inoculated with 105 viable chlamydospores of P. chlamydosporia just before transplanting. One week later, the responder part of the root of the treatments containing RKN was inoculated at a rate of 1 J2 per cm3 of soil. The treatment None-None, with no inoculation with either fungi or RKN received the same volume of water. The plants were maintained in a growth chamber in the same conditions described previously in a completely randomized design for 40 days. The plants were irrigated as needed and fertilized with Hoagland solution twice per week. Soil temperatures were recorded daily at 30-min intervals with a PT100 probe (Campbell Scientific Ltd) placed in the pots at a depth of 4 cm. At the end of the experiments, both inducer and responder root fresh weight and the shoot dry weight of each single plant were measured. Roots from the RKN-inoculated responder were immersed in a 0.01% erioglaucine solution for 45 min to stain the egg masses (Omwega et al., 1988) before counting them. Afterward, the eggs were extracted from the roots as in Hussey and Barker (1973)’s method and counted. The number of egg masses was considered as the infective capability of the nematode because it indicates the number of J2 able to penetrate, to infect the root, and to develop into egg-laying females. The number of eggs was considered the reproductive capability of the nematode, and the female fecundity was calculated as the number of eggs per egg mass.
The tomato and cucumber root colonization by each fungal isolates was estimated by quantifying the fungal DNA by qPCR at the end of the second experiment. The inducer part of the root was washed three times in sterilized distilled water for 10 s each and then blotted onto sterile paper. Per each fungal isolate three biological replicates were assessed. Each biological replicate consisted of the inducer part of the roots from three plants pooled together. The DNA was extracted from each biological replicate following the Lopez-Llorca et al. (2010)’s procedure. qPCR reactions were performed using the FastStart Universal SYBR Green Master (Roche) mix in a final volume of 25 μl containing 50 ng of total DNA and 0.3 μM of each primer (5′ to 3′ direction) VCP1-1F (CGCTGGCTCTCTCACTAAGG) and VCP1-2R (TGCCAGTGTCAAGGACGTAG) (Escudero and Lopez-Llorca, 2012). Negative controls containing sterile water instead of DNA were included. Reactions were performed in duplicate in a Stratagene Mx3005P thermocycler (Agilent Technologies) using the following thermal cycling conditions: initial denaturation step at 95°C for 2 min, then 40 cycles at 95°C for 30 s, and 62°C for 30 s. Genomic DNA dilutions of the fungal isolate M10.43.21 were used to define a calibration curve from 5 pg to 50 ng. After each run, the specificity of the PCR amplicons was verified by melting curve analysis and agarose gel electrophoresis. The fungal DNA biomass of each isolate was referred to the total DNA biomass (50 ng) and expressed as a proportion.
Dynamic Regulation of the Jasmonic and Salicylic Acid Pathways by P. chlamydospora and M. incognita
Tomato seeds were sterilized as previously described. Three-week-old tomato seedlings were transferred to 200 cm3 pots with sterilized sand and maintained in a growth chamber as previously described. The fungal isolate M10.43.21 was selected for this experiment because it reduced nematode infectivity and reproduction in the split-root system experiments. The experiment consisted of two treatments: non-inoculated and co-inoculated to determine the expression of genes related to the salicylic acid and jasmonic acid pathways. In the co-inoculated treatment, the soil was inoculated with 105 viable chlamydospores just before transplanting and with 1 J2 of M. incognita per cm3 1 week after transplanting. Each treatment was replicated 40 times. An additional treatment only inoculated with the nematode was included to determine the effect of the fungal isolate on nematode reproduction. The fungus and nematode inoculation procedure was as previously stated.
The expression of the pathogenesis-related protein 1 (PR-1) gene and the lipoxygenase (Lox D) gene the from the salicylic acid and jasmonic acid pathways, respectively, was evaluated at three time points: just after nematode inoculation, that is, at 0 days after nematode inoculation (dani), at 7 dani, and at 42 dani. At each assessment time, roots were washed three times with sterile distilled water, placed onto sterilized filter paper, frozen in liquid nitrogen and stored at −80°C until being used. At 7 dani, the J2 were stained inside roots with acid Fuchsin following the Byrd et al. (1983) procedure to confirm that nematode had penetrated and infected. At the end of the experiment (42 dani), the number of eggs per plant from three plants for each treatment was determined by extracting them as described previously.
Total RNA from roots was isolated using the PureLink RNA Mini Kit (Invitrogen), according to the manufacturer’s instructions. Afterward, the DNA-free kit (Invitrogen) was used to remove the remaining DNA from the sample. Total RNA integrity and quantity of the samples were assessed by means of agarose gel, NanoDrop 1000 Spectrophotometer (Thermo Scientific) and Qubit RNA BR assay kit (Thermo Fisher Scientific). To assure that the sample was DNA free, a PCR was carried out. Then, the RNA was retro-transcribed with the SuperScript II (Invitrogen) according to manufacturer’s instructions. The relative gene expression was estimated with the ∆∆Ct methodology (Livak and Schmittgen, 2001), using the ubiquitin (UBI) gene as a reference gene (Song et al., 2015). Primers used in the RT-qPCR were (5′ to 3′ direction): LeUbi3-F (TCCATCTCGTGCTCCGTCT), LeUbi3-R (GAACCTTTCCAGTGTCATCAACC) Song et al. (2015), LoxD-F (GACTGGTCCAAGTTCACGATCC), LoxD-R (ATGTGCTGCCAATATAAATGGTTCC) Fujimoto et al. (2011), LEPR1F (GCAACACTCTGGTGGACCTT), and LEPR1R (ATGGACGTTGTCCTCTCCAG) Gayoso et al. (2007). qPCR reactions were performed in a final volume of 20 μl with 1 μl of cDNA, 0.3 mM of the corresponding primers, and 1X Fast SYBR Green Master Mix (Applied Biosystems). The qPCR was performed in a 7900HT Fast Real Time PCR System thermocycler (Applied Biosystems) using: 20 s at 95°C followed by 40 cycles of 30 s at 95°C and 60 s at 60°C (Gayoso et al., 2007). The specificity of PCR amplicons was verified as described previously. Reactions were performed with three biological replicates per treatment. Each biological replicate consisted of the roots from three plants pooled together. Two technical replicates per biological replicate were assessed.
Statistical Analysis
Statistical analyses were performed using the JMP software v8 (SAS institute Inc., Cary, NC, USA). Both data normality and homogeneity of variances were assessed. When confirmed, a paired comparison using the Student’s t-test was done. Otherwise, paired comparison was done using the non-parametric Wilcoxon test, or multiple comparison using the Kruskal-Wallis test and groups separated by Dunn’s test (p ≤ 0.05). The repetitions of the split-root experiments for each crop were compared using the non-parametric Wilcoxon test, and considered as one experiment when no differences (p ≤ 0.05) were found.
Results
Induction of Systemic Plant Resistance by P. chlamydosporia Isolates Against M. incognita
The split-root system experiments with tomato differed (p < 0.05) between them and were treated separately. But no differences were found between the two split-root experiments with cucumber and thus we considered them as replicates of a single experiment. Both tomato and cucumber fresh root weight of the two halves of the split-root system of the None-None treatment did not differ (p < 0.05) (data not shown), showing that the split-root system did not influence root development. Shoot dry biomass did not differ in any fungal isolate-plant species combination (p < 0.05) (data not shown).
Two of the five P. chlamyodosporia isolates induced resistance in tomato plants in both experiments, but none of them did in cucumber (Table 1). The fungal isolate M10.43.21 reduced both the number of egg masses per plant (32–43%), the number of eggs per plant (44–59%), and the female fecundity (14.7–27.6%), while the isolate M10.55.6 reduced the number of eggs per plant in both experiments (35–47.5%) but the number of egg masses or the female fecundity in only one.
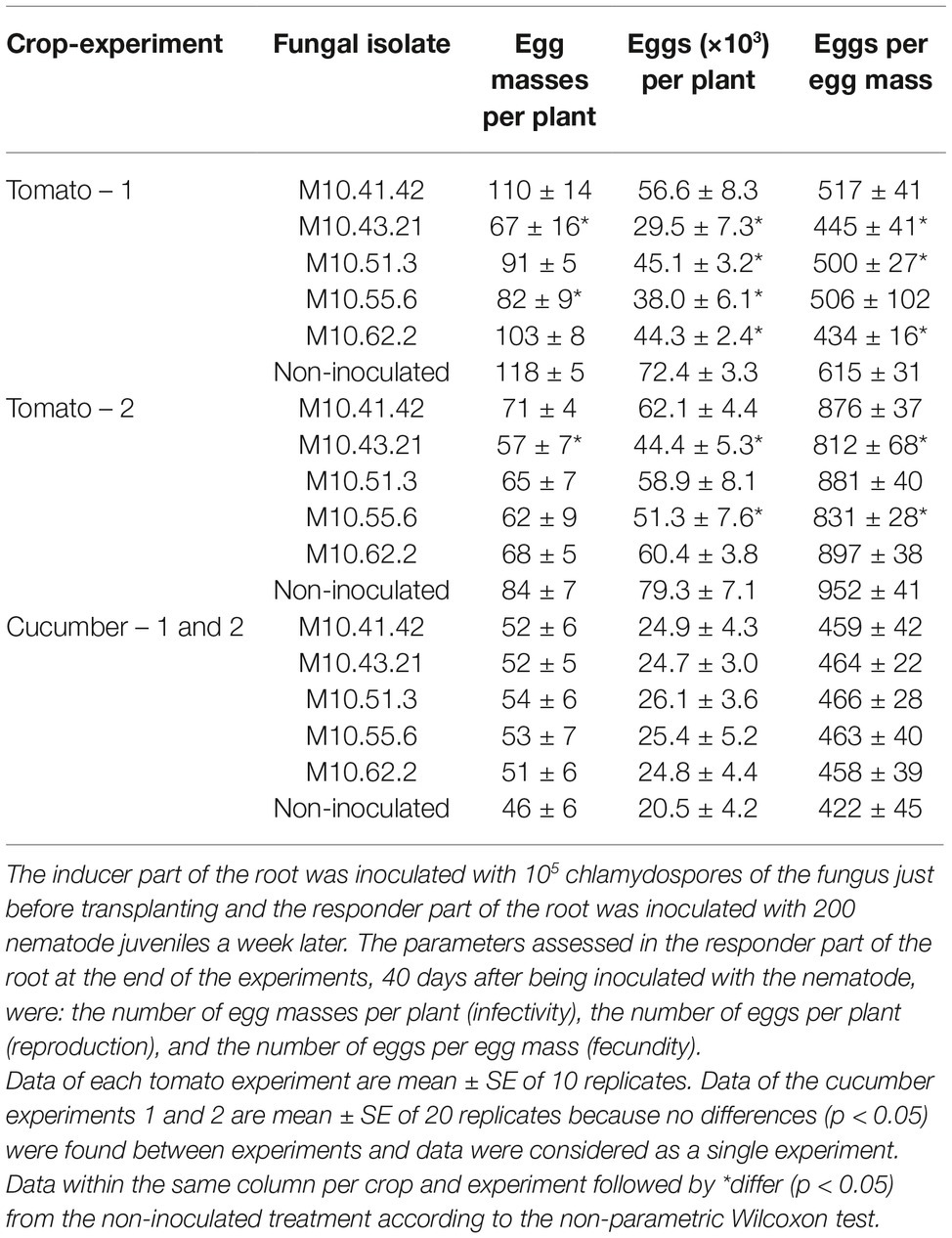
Table 1. Capability of P. chlamydosporia isolates to induce systemic resistance in tomato cv. Durinta or cucumber cv. Dasher II against Meloidogyne incognita in two split root experiments.
P. chlamydosporia isolates differed in the level of root colonization estimated by qPCR irrespective of the plant species (Figure 1). The standard curves for qPCR obtained by representing the cycle thresholds (Ct) against the log of 10-fold serial dilution of DNA from isolate M10.43.21 were accurate and reproducible to estimate the DNA concentration of the different treatments (tomato = −3.66x + 19.36; R2 = 0.9736 and cucumber y = −3.4937x + 21.29; R2 = 0.9947). Regarding tomato, isolate M10.43.21 was the best root colonizer followed by M10.55.6 (Figure 1A). In relation to cucumber, isolate M10.55.6 was the best root colonizer followed by M10.43.21 (Figure 1B). The fungus was not detected in the inducer part of the root from treatments None-None and None-RKN.
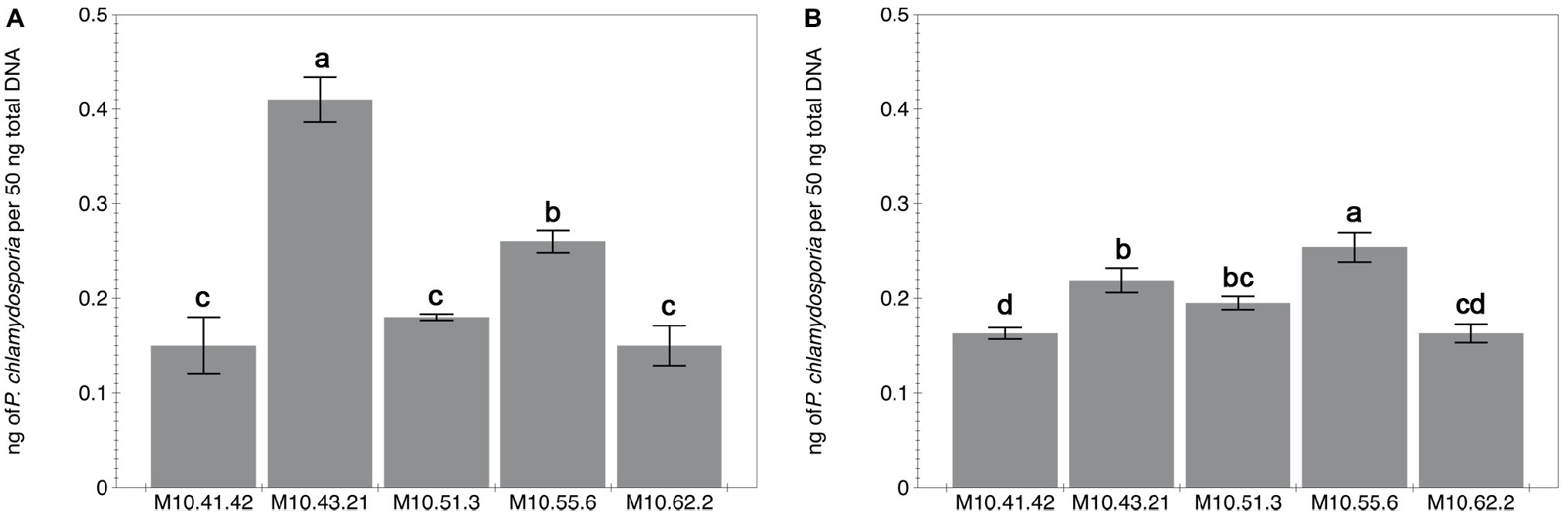
Figure 1. Capability of five P. chlamydosporia isolates to colonize roots of tomato cv. Durinta (A) and cucumber cv. Dasher II (B). The root colonization is expressed as the proportion of fungal DNA biomass (isolates) per 50 ng of the total DNA biomass extracted from the inducer part of the root of the split-root experiment 2. Each value is mean ± SE of three biological samples with two technical replicates each. Different letters indicate statistical differences between isolates (p < 0.05) according to the Dunn’s test.
Dynamic Regulation of the Jasmonic and Salicylic Acid Pathways by P. chlamydosporia and M. incognita in Tomato
Changes in the expression of genes PR-1 and Lox D from the salicylic acid and jasmonic acid pathways at 0, 7, and 42 dani are shown in Figure 2. The expression of the PR-1 gene in roots inoculated with the fungal isolate M10.43.21 was upregulated at 0, 7, and 42 dani compared to the non-inoculated plants (Figures 2A–C). Regarding the jasmonic acid pathway (Figures 2D–F), the gene Lox D was only upregulated at 7 dani.
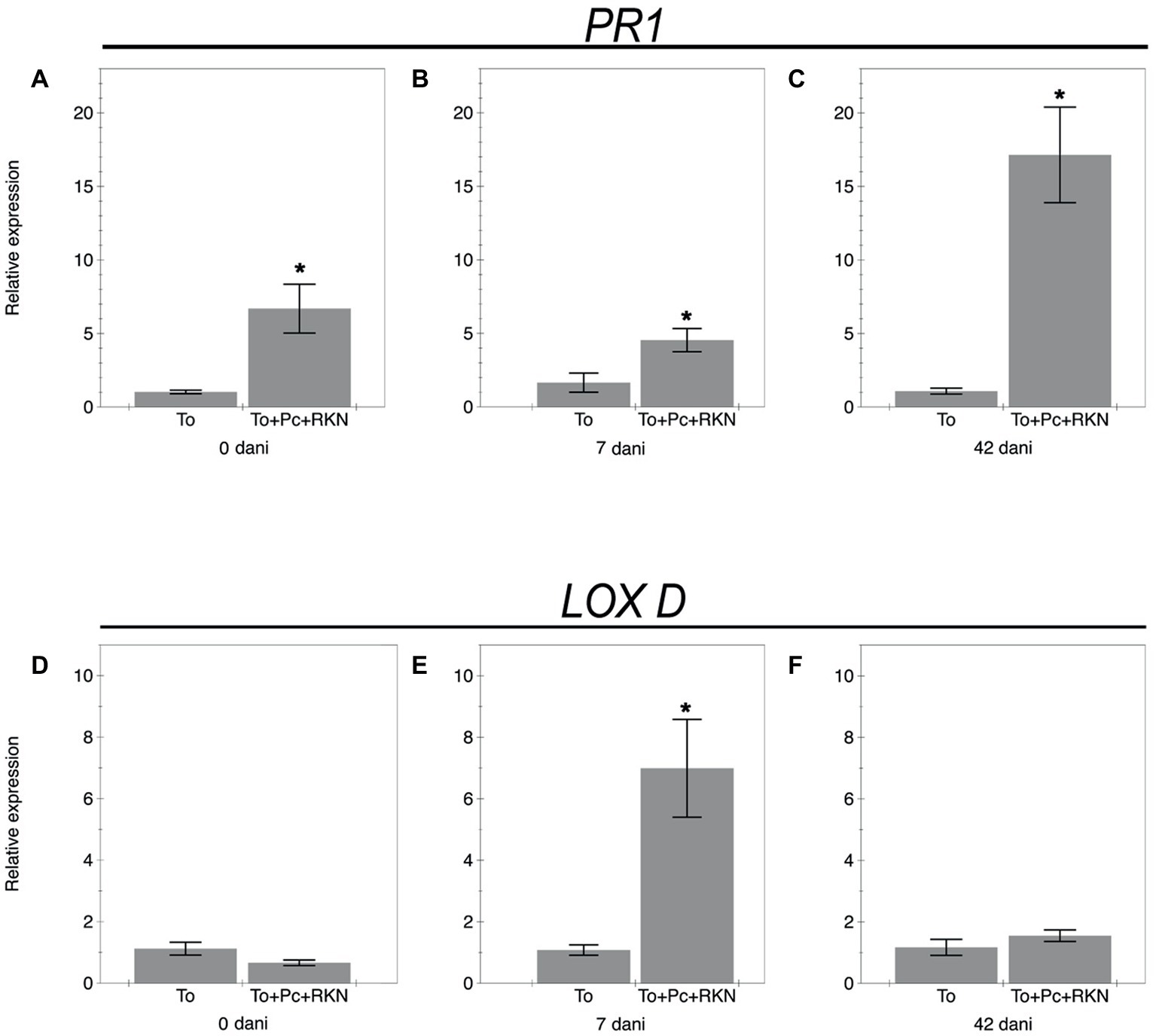
Figure 2. Relative expression of genes related to the salicylic acid and jasmonic acid pathways. The expression of genes Pr1 (A–C) and Lox D (D–F) in roots of the tomato cv. Durinta non inoculated (To) or inoculated with 105 chlamydospores of P. chlamydosporia isolate M10.43.21 just before transplanting and with 200 J2 of M. incognita per plant a week after transplanting (To+Pc + RKN) at three time points: just after nematode inoculation (0 dani), at 7 dani, and at 42 dani. Each value is mean ± SE of three biological samples with two technical replicates each. Asterisks indicate significant differences (p < 0.05) according to the non-parametric Wilcoxon test.
The nematode reproduction in plants co-inoculated with the fungal isolate was suppressed by 60% compared to plants only inoculated with the nematode (Figure 3).
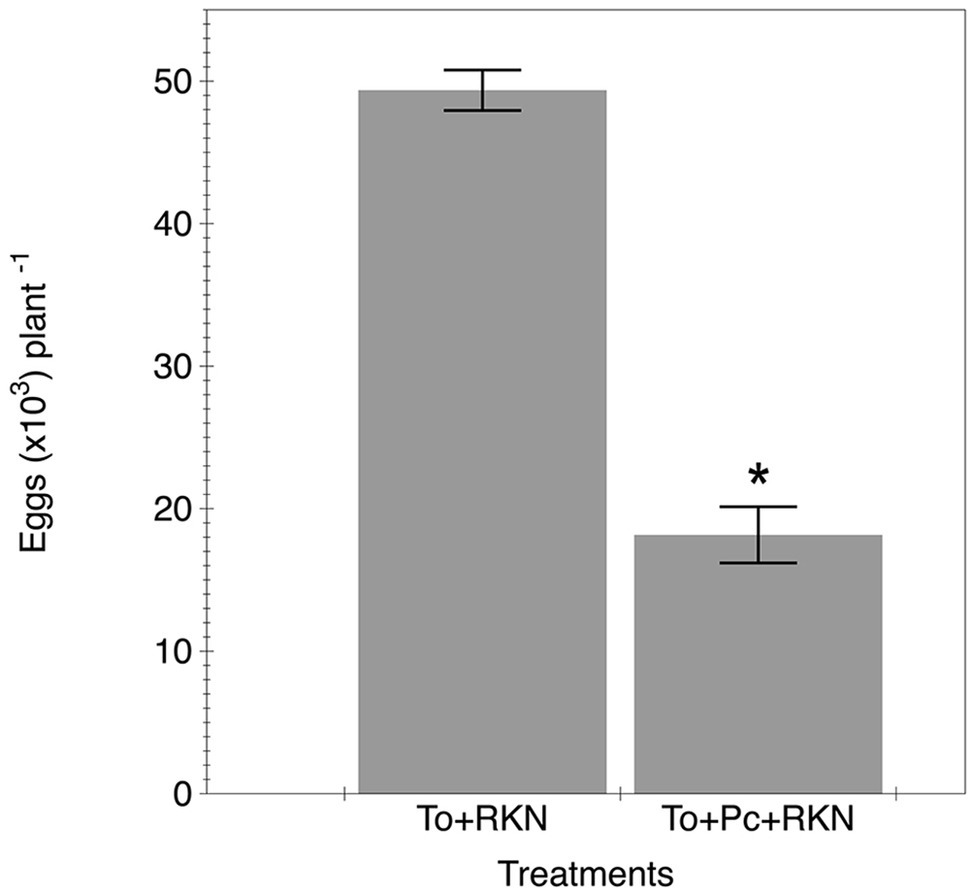
Figure 3. Effect of primed tomato plants by P. chamydosporia on M. incognita reproduction. Number of eggs produced in the tomato cv. Durinta after 42 days of being inoculated with 200 J2 of M. incognita per plant a week after transplanting (To + RKN) or inoculated with 105 chlamydospores of P. chlamydosporia isolate M10.43.21 just before transplanting and with the nematode at the same rate and time mentioned before (To + Pc + RKN). Each value is mean ± SE of three replications. Asterisk indicates significant differences (p < 0.05) according to the non-parametric Wilcoxon test.
Discussion
The results of this study provide evidence for the ability of some P. chlamydosporia isolates to induce systemically resistance against M. incognita, and that this induction is dependent on the plant species. The isolate M10.43.21 showed the most consistent response in both split-root experiments with tomato, and was the reason for selecting it to determine the hormone modulation in this plant species. The mechanisms responsible for the endophyte-induced resistance are unclear (Schouten, 2016). Both salicylic acid- and jasmonic acid-dependent signaling pathways have been proposed as responsible for the systemically induced resistance to Meloidogyne spp. in tomato in split-root experiments (Selim, 2010; de Medeiros et al., 2017; Martínez-Medina et al., 2017). Martínez-Medina et al. (2017) reported that Trichoderma harzianum T-78 induced the upregulation of genes related to salicylic acid at early stage of nematode infection, whereas those related to jasmonic acid were upregulated from 3 to 21 days after nematode inoculation. In our study, the P. chlamydosporia isolate primed salicylic acid from the first assessment time (7 days after fungal inoculation and just after nematode inoculation) until the end of the experiment (42 dani). This effect could be responsible for the suppression of nematode infection at early stages, as well as the infection by the J2 produced by the primary inoculum that were able to overcome the plant defense mechanisms. In addition, the upregulation of the Lox D gene, related to jasmonic acid, at 7 dani could affect nematode reproduction and fecundity. Thus, the induction of the salicylic acid and jasmonic acid signaling pathways by the fungal isolate M10.43.21 in tomato counteract the suppression of these phytohormones by the nematode described in the susceptible tomato-nematode interaction (Shukla et al., 2018). The three-phase model proposed to explain the induced protection to RKN by Trichoderma harzianum T-78 in tomato consisting of salicylic acid induction suppressing RKN infection followed by jasmonic acid induction suppressing RKN reproduction and fecundity and finally salicylic acid induction affecting root infection by the next J2 generation (Martínez-Medina et al., 2017) is valid for P. chlamydosporia. Other local plant defense mechanisms induced by P. chlamydosporia against RKN have been reported, including the increase of the peroxidases (POX) and poliphenoloxidases (PPO) enzymes activity at root nematode invasion stage (24–96 h after nematode inoculation) (de Medeiros et al., 2015). However, considering that P. chlamydosporia does not extensively colonize the root (Maciá-Vicente et al., 2009; Escudero and Lopez-Llorca, 2012), even being improved by chitosan irrigation (Escudero et al., 2017), the effect of local defense mechanisms alone may be insufficient to achieve significant nematode suppression.
Not all P. chlamydosporia isolates induced systemic resistance in tomato. The variability of this attribute among fungal isolates can be added to other observations previously reported, such as the production of chlamydospores, root colonization, plant growth promotion, or egg parasitism (Kerry and Bourne, 2002; Zavala-Gonzalez et al., 2015). In fact, egg parasitism can even differ between single-fungal spore isolates originating from the same field population (Giné et al., 2016). The frequency of occurrence of P. chlamydosporia in horticultural production sites under integrated and organic standards has increased since the 1990s in northeastern Spain (Giné et al., 2012), showing that this fungal species is adapted to environmental characteristics and agronomic practices. Field populations of P. chlamydosporia can contain individuals representing a diversity of functions that are highly beneficial to plants, such as plant growth promoters which enhance plant tolerance; inducers of plant defense mechanisms suppressing infection, development and reproduction of RKN; efficient egg parasites suppressing the RKN inoculum; or saprophytic behavior contributing to the organic matter cycle and plant nutrition. A given proportion of P. chlamydosporia representing some or all of these functions could be present in a given soil and adapted to the plant species involved in the rotation scheme and contributing to their health status. It seems that most of these functions are not interlinked. In fact, none of the five fungal isolates assessed in our study was a plant growth promoter. Thus, molecular tools must be developed to facilitate knowledge of the functional composition of the fungal field population in a given soil.
P. chlamydosporia has been reported as the main biotic factor responsible for soil suppressiveness to RKN in horticultural crops (Giné et al., 2016). In soils with low antagonistic potential, the use of fungal isolates with both direct and indirect action mechanisms could suppress RKN. Indeed, primed plants along with egg parasitism will protect against infection and reproduction of RKN and decrease the inoculum viability. The combination of the two modes of action will result in a decrease of the nematode population growth rate, and consequently lower crop yield losses. Alternatively, combining the use of P. chlamydosporia with plant defense activators can produce a similar effect. Vieira Dos Santos et al. (2014) found a reduction in RKN reproduction when P. chlamydosporia was combined with the application of cis-jasmone, as well as an increase in fungal egg parasitism.
In conclusion, this study proves that some P. chlamydosporia isolates induce systemic resistance to M. incognita in tomato but none of them in cucumber. Thus, this response is plant species dependent. In future studies, the interaction between P. chlamydosporia isolates and selected economically important crops should be characterized to elucidate the mechanisms and genes involved in inducing plant resistance in order to maximize the efficacy of control.
Data Availability
All datasets generated for this study are included in the manuscript and/or the supplementary files.
Author Contributions
FS and NE conceived, designed, supervised the experiments, the data collection, and analyses. ZG performed the experiments, analyzed the data, and wrote the draft of the manuscript. NE and ES performed the gene expression analyses. TG provided reagents, materials, and advice. NE, ES, TG, and FS reviewed and wrote the final draft of the manuscript.
Funding
This study was supported by projects AGL2013-49040-C2-1-R and AGL2017-89785-R financed by the Spanish Ministry of Economy and Competitiveness (MINECO) and the European Regional Development Fund (FEDER).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Thanks are given to Ms. Sheila Alcala and Ms. Maria Julià for their technical support.
References
Becerra Lopez‐Lavalle, L. A., Pottera, N., and Brubakerb, C. L. (2012). Development of a rapid, accurate glasshouse bioassay for assessing fusarium wilt disease responses in cultivated Gossypium species. Plant Pathol. 61, 1112–1120. doi: 10.1111/j.1365-3059.2012.02603.x
Bordallo, J. J., Lopez-Llorca, L. V., Jansson, H. B., Salinas, J., Persmark, L., and Asensio, L. (2002). Colonization of plant roots by egg-parasitic and nematode-trapping fungi. New Phytol. 154, 491–499. doi: 10.1046/j.1469-8137.2002.00399.x
Byrd, D. W. Jr., Kirkpatrick, T., and Barker, K. R. (1983). An improved technique for clearing and staining tissues for detection of nematodes. J. Nematol. 15, 142–143.
Dababat, A. E. F. A., and Sikora, R. A. (2007a). Influence of the mutualistic endophyte Fusarium oxysporum 162 on Meloidogyne incognita attraction and invasion. Nematology 9, 771–776. doi: 10.1163/156854107782331225
Dababat, A. A., and Sikora, R. A. (2007b). Induced resistance by the mutualistic endophyte, Fusarium oxysporum strain 162, toward Meloidogyne incognita on tomato. Biocont. Sci. Technol. 17, 969–975. doi: 10.1080/09583150701582057
Davies, K. G., Rowe, J., Manzanilla-López, R., and Opperman, C. H. (2011). Re-evaluation of the life-cycle of the nematode-parasitic bacterium Pasteuria penetrans in root-knot nematodes, Meloidogyne spp. Nematology 13, 825–835. doi: 10.1163/138855410x552670
de Medeiros, H. A., de Araújo Filho, J. V., de Freitas, L. G., Castillo, P., Rubio, M. B., Hermosa, R., et al. (2017). Tomato progeny inherit resistance to the nematode Meloidogyne javanica linked to plant growth induced by the biocontrol fungus Trichoderma atroviride. Sci. Rep. 7:40216. doi: 10.1038/srep40216
de Medeiros, H. A., Resende, R. S., Ferreira, F. C., Freitas, L. G., and Rodrigues, F. A. (2015). Induction of resistance in tomato against Meloidogyne javanica by Pochonia chlamydosporia. Nematoda 2:e10015. doi: 10.4322/nematode.10015
Djian-Caporalino, C. (2012). Root-knot nematode (Meloidogyne spp.), a growing problem in French vegetable crops. EPPO Bull. 42, 127–137. doi: 10.1111/j.1365-2338.2012.02530.x
Escudero, N., and Lopez-Llorca, L. V. (2012). Effects on plant growth and root-knot nematode infection of an endophytic GFP transformant of the nematophagous fungus Pochonia chlamydosporia. Symbiosis 57, 33–42. doi: 10.1007/s13199-012-0173-3
Escudero, N., Lopez-Moya, F., Ghahremani, Z., Zavala-Gonzalez, E. A., Alaguero-Cordovilla, A., Ros-Ibañez, C., et al. (2017). Chitosan increases tomato root colonization by Pochonia chlamydosporia and their combination reduces root-knot nematode damage. Front. Plant Sci. 8:1415. doi: 10.3389/fpls.2017.01419
Fujimoto, T., Tomitaka, Y., Abe, H., Tsuda, S., Futai, K., and Mizuhubo, T. (2011). Expression profile of jasmonic acid-induced genes and the induced resistance against the root-knot nematode (Meloidogyne incognita) in tomato plants (Solanum lycopersicum) after foliar treatment with methyl jasmonate. J. Plant Physiol. 168, 1084–1097. doi: 10.1016/j.jplph.2010.12.002
Gayoso, C., Ilárduya, O. M., Pomar, F., and Merino de Caceres, F. (2007). Assessment of real-time PCR as a method for determining the presence of Verticillium dahliae in different solanaceae cultivars. Eur. J. Plant Pathol. 118, 199–209. doi: 10.1007/s10658-007-9134-8
Giné, A., Bonmatí, M., Sarro, A., Stchiegel, A., Valero, J., Ornat, C., et al. (2012). Natural occurrence of fungal egg parasites of root-knot nematodes, Meloidogyne spp. in organic and integrated vegetable production systems in Spain. BioControl 58, 407–416. doi: 10.1007/s10526-012-9495-6
Giné, A., Carrasquilla, M., Martínez-Alonso, M., Gaju, N., and Sorribas, F. J. (2016). Characterization of soil suppressiveness to root-knot nematodes in organic horticulture in plastic greenhouse. Front. Plant Sci. 7:164. doi: 10.3389/fpls.2016.00164
Giné, A., González, C., Serrano, L., and Sorribas, F. J. (2017). Population dynamics of Meloidogyne incognita on cucumber grafted onto the Cucurbita hybrid RS841 or ungrafted and yield losses under protected cultivation. Eur. J. Plant Pathol. 148, 795–805. doi: 10.1007/s10658-016-1135-z
Giné, A., López-Gómez, M., Vela, M. D., Ornat, C., Talavera, M., Verdejo-Lucas, S., et al. (2014). Thermal requirements and population dynamics of root-knot nematodes on cucumber and yield losses under protected cultivation. Plant Pathol. 63, 1446–1453. doi: 10.1111/ppa.12217
Giné, A., and Sorribas, F. J. (2017). Quantitative approach for the early detection of selection for virulence of Meloidogyne incognita on resistant tomato in plastic greenhouses. Plant Pathol. 66, 1338–1344. doi: 10.1111/ppa.12679
Greco, N., and Di Vito, M. (2009). “Population dynamics and damage levels” in Root-knot nematodes. eds. R. N. Perry, M. Moens, and J. L. Starr (Wallingford, UK: CABI International), 246–274.
Hussey, R. S., and Barker, K. R. (1973). A comparison of methods of collecting inoculate of Meloidogyne spp. including a new technique. Plant Dis. Rep. 57, 1025–1028.
Jones, J. T., Haegeman, A., Danchin, E. G., Gaur, H. S., Helder, J., Jones, M. G., et al. (2013). Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 14, 946–961. doi: 10.1111/mpp.12057
Kerry, B. R., and Bourne, J. M. (2002). A manual for research on Verticillium chlamydosporium, a potential biological control agent for root-knot nematodes. Gent: IOBC/WPRS.
Larriba, E., Jaime, M. D. L. A., Nislow, C., Martín-Nieto, J., and Lopez-Llorca, L. V. (2015). Endophytic colonization of barley (Hordeum vulgare) roots by the nematophagous fungus Pochonia chlamydosporia reveals plant growth promotion and a general defense and stress transcriptomic response. J. Plant Res. 128, 665–678. doi: 10.1007/s10265-015-0731-x
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods 25, 502–508. doi: 10.1006/meth.2001.1262
López-Gómez, M., Giné, A., Vela, M. D., Ornat, C., Sorribas, F. J., Talavera, M., et al. (2014). Damage function and thermal requirements of Meloidogyne javanica and Meloidogyne incognita on watermelon. Ann. Appl. Biol. 165, 466–473. doi: 10.1111/aab.12154
Lopez-Llorca, L. V., Gómez-Vidal, S., Monfort, E., Larriba, E., Casado-Vela, J., Elortza, F., et al. (2010). Expression of serine proteases in egg-parasitic nematophagous fungi during barley root colonization. Fungal Genet. Biol. 47, 342–351. doi: 10.1016/j.fgb.2010.01.004
Maciá-Vicente, J. G., Rosso, L. C., Ciancio, A., Jansson, H. B., and Lopez-Llorca, L. V. (2009). Colonisation of barley roots by endophytic Fusarium equiseti and Pochonia chlamydosporia: effects on plant growth and disease. Ann. Appl. Biol. 155, 391–401. doi: 10.1111/j.1744-7348.2009.00352.x
Manzanilla-López, R. H., Esteves, I., Powers, S. J., and Kerry, B. R. (2011). Effects of crop plants on abundance of Pochonia chlamydosporia and other fungal parasites of root-knot and potato cyst nematode. Ann. Appl. Biol. 159, 118–129. doi: 10.1111/j.1744-7348.2011.00479.x
Martínez-Medina, A., Fernandez, I., Lok, G. B., Pozo, M. J., Pieterse, C. M., and Van Wees, S. C. (2017). Shifting from priming of salicylic acid- to jasmonic acid-regulated defences by Trichoderma protects tomato against the root knot nematode Meloidogyne incognita. New Phytol. 213, 1363–1377. doi: 10.1111/nph.14251
Omwega, C., Thomason, I. J., and Roberts, P. A. (1988). A non-destructive technique for screening bean germ plasm for resistance to Meloidogyne incognita. Plant Dis. 72, 970–972. doi: 10.1094/PD-72-0970
Pentimone, I., Lebrón, R., Hackenberg, M., Rosso, L. C., Colagiero, M., Nigro, F., et al. (2018). Identification of tomato miRNAs responsive to root colonization by endophytic Pochonia chlamydosporia. Appl. Microbiol. Biotechnol. 102, 907–919. doi: 10.1007/s00253-017-8608-7
Schouten, A. (2016). Mechanisms involved in nematode control by endophytic fungi. Annu. Rev. Phytopathol. 54, 121–142. doi: 10.1146/annurev-phyto-080615-100114
Seid, A., Fininsa, C., Mekete, T., Decraemer, W., and Wesemael, W. M. L. (2015). Tomato (Solanum lycopersicum) and root-knot nematodes (Meloidogyne spp.) – a century-old battle. Nematology 17, 995–1009. doi: 10.1163/15685411-00002935
Selim, M. L. M. (2010). Biological, chemical and molecular studies on the systemic induced resistance in tomato against Meloidogyne incognita caused by the endophytic Fusarium oxysporum, Fo162. PhD thesis. Bonn, Germany: University of Bonn.
Shukla, N., Yadav, R., Kaur, P., Rasmussen, S., Goel, S., Agarwal, M., et al. (2018). Transcriptome analysis of root-knot nematode (Meloidogyne incognita)-infected tomato (Solanum lycopersicum) roots reveals complex gene expression profiles and metabolic networks of both host and nematode during susceptible and resistance responses. Mol. Plant Pathol. 19, 615–633. doi: 10.1111/mpp.12547
Sikora, R., and Fernández, E. (2005). “Nematode parasites of vegetables” in Plant parasitic nematodes in subtropical and tropical agriculture. eds. M. Luc, R. A. Sikora, and J. Bridge (Wallingford, UK: CABI International), 319–392.
Song, Y., Chen, D., Lu, K., Sun, Z., and Zeng, R. (2015). Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front. Plant Sci. 6:786. doi: 10.3389/fpls.2015.00786
Stirling, G. (2014). Biological control of plant-parasitic nematodes: Soil ecosystem management in sustainable agriculture. 2nd Edn. Wallingford, UK: CABI International.
Talavera, M., Sayadi, S., Chirosa-Ríos, M., Salmerón, T., Flor-Peregrín, E., and Verdejo-Lucas, S. (2012). Perception of the impact of root-knot nematode-induced diseases in horticultural protected crops of south-eastern Spain. Nematology 14, 517–527. doi: 10.1163/156854112X635850
Vieira Dos Santos, M. C., Curtis, R., and Abrantes, I. (2014). The combined use of Pochonia chlamydosporia with plant defence activators. A potential sustainable control strategy for Meloidogyne chitwoodi. Phytopathol. Mediterr. 53, 66–74. doi: 10.14601/Phytopathol_Mediterr-12186
Wesemael, W. M. L., Viaene, N., and Moens, M. (2011). Root-knot nematodes (Meloidgoyne spp.) in Europe. Nematology 13, 3–16. doi: 10.1163/138855410X526831
Whitehead, A. G., and Hemming, J. R. (1965). A comparison of some quantitative methods of extracting small vermiform nematodes from soil. Ann. Appl. Biol. 55, 25–38. doi: 10.1111/j.1744-7348.1965.tb07864.x
Zavala-Gonzalez, E. A., Escudero, N., Lopez-Moya, F., Aranda-Martinez, A., Exposito, A., Ricaño-Rodríguez, J., et al. (2015). Some isolates of the nematophagous fungus Pochonia chlamydosporia promote root growth and reduce flowering time in tomato. Ann. Appl. Biol. 166, 472–483. doi: 10.1111/aab.12199
Zavala-Gonzalez, E. A., Rodríguez-Cazorla, E., Escudero, N., Aranda-Martinez, A., Martínez-Laborda, A., Ramírez-Lepe, M., et al. (2017). Arabidopsis thaliana root colonization by the nematophagous fungus Pochonia chlamydosporia is modulated by jasmonate signaling and leads to accelerated flowering and improved yield. New Phytol. 213, 351–364. doi: 10.1111/nph.14106
Keywords: Cucumis sativus, induced resistance, root endophytes, root-knot nematodes, Solanum lycopersicum
Citation: Ghahremani Z, Escudero N, Saus E, Gabaldón T and Sorribas FJ (2019) Pochonia chlamydosporia Induces Plant-Dependent Systemic Resistance to Meloidogyne incognita. Front. Plant Sci. 10:945. doi: 10.3389/fpls.2019.00945
Edited by:
Danny Coyne, International Institute of Tropical Agriculture (IITA), KenyaReviewed by:
Zafar Ahmad Handoo, Agricultural Research Service, United StatesMaria Clara Vieira Dos Santos, University of Minho, Portugal
Copyright © 2019 Ghahremani, Escudero, Saus, Gabaldón and Sorribas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: F. Javier Sorribas, francesc.xavier.sorribas@upc.edu