 Aline Pereira Rank
Aline Pereira Rank Aline Koch
Aline Koch- Institute of Phytomedicine, University of Hohenheim, Stuttgart, Germany
The drastic loss of biodiversity has alarmed the public and raised sociopolitical demand for chemical pesticide-free plant production, which is now treated by governments worldwide as a top priority. Given this global challenge, RNAi-based technologies are rapidly evolving as a promising substitute to conventional chemical pesticides. Primarily, genetically modified (GM) crops expressing double-stranded (ds)RNA-mediating gene silencing of foreign transcripts have been developed. However, since the cultivation of GM RNAi crops is viewed negatively in numerous countries, GM-free exogenous RNA spray applications attract tremendous scientific and political interest. The sudden rise in demand for pesticide alternatives has boosted research on sprayable RNA biopesticides, generating significant technological developments and advancing the potential for field applications in the near future. Here we review the latest advances that could pave the way for a quick lab-to-field transition for RNA sprays, which, as safe, selective, broadly applicable, and cost-effective biopesticides, represent an innovation in sustainable crop production. Given these latest advances, we further discuss technological limitations, knowledge gaps in the research, safety concerns and regulatory requirements that need to be considered and addressed before RNA sprays can become a reliable and realistic agricultural approach.
Introduction
Modern agriculture faces diverse challenges related to the fact that the world population is expected to rise by 26% by 2050. Given this projection, agriculturists must boost productivity to meet emerging food needs (United Nations Department of Economics and Social Affairs, 2019). Major challenges to these production levels arise due to agricultural pests. Pests contribute estimated losses of up to 40% of crops worldwide, with insects serving as causal agents of damage and disease transmission (Douglas, 2018). So far, plant protection primarily relies on the use of chemical pesticides. However, the use of conventional pesticides contributes to soil, water, and air pollution and is predicted to be a major driver of an alarming loss of biodiversity worldwide (Wagner et al., 2021). In addition, improper and controlled chemical treatments detrimentally affect the environment, consumers, and users. Moreover, perennial applications result in the development of pesticide resistance in pathogens and pests (Fotoukkiaii et al., 2021).
Intergovernmental initiatives such as the EU’s farm-to-fork strategy (part of the European green deal) aim to rethink and redesign food systems, for example by developing frameworks for sustainable food production. A central goal is to reduce the overall use and risk of chemical pesticides by 50% by 2030 (European Commission, 2021). Given the social-political demand for an ideally pesticide-free agriculture, safe “green” alternatives are urgently desired to facilitate sustainability in crop protection. Thus, RNAi-based technologies are discussed as low risk pesticides that may enable reaching pesticide reduction and sustainability goals (Taning et al., 2021).
A promising alternative to chemical pesticides relies on harnessing the mechanistic of RNA interference (RNAi). Briefly, RNAi is a conserved cellular mechanism that regulates and protects eukaryotic cells against harmful nucleic acids. In this natural phenomenon, double-stranded RNAs (dsRNAs) are processed by Dicer enzymes into 21–24 nucleotide (nt) small interfering RNAs (siRNAs). These siRNAs either mediate transcriptional gene silencing (TGS) by ensuring inhibitory DNA and histone modifications as well as chromatin remodeling, or they facilitate post-transcriptional gene silencing (PTGS) by preventing translation of targeted mRNA transcripts. Both TGS and PTGS served as blueprints for the development and continuous improvement of RNAi toolkits. In agriculture, RNAi has proven to be an effective strategy in controlling pathogens such as viruses, bacteria, and fungi, as well as insect pests, mites, and nematodes (Fire et al., 1998; Baum et al., 2007; Zotti et al., 2018; Mezzetti et al., 2020; Koch and Wassenegger, 2021).
A decade ago, studies focused on the transgene-based endogenous expression and formation of dsRNA in host plants, termed host-induced gene silencing (HIGS) (Nowara et al., 2010). Nowadays, transgenic approaches are clouded by some major drawbacks. First, these approaches are laborious, complicated, and time-consuming, and their applicability is restricted by the transformability of the host plant. They are also very expensive; for example, the commercialization of a transgenic crop is estimated to reach 140 million dollars (Rosa et al., 2018). Finally, they are weakly accepted, as public distrust of GM plants persists, with an ongoing upward trend (Herman et al., 2021). Additionally, orders and species present completely different and variable demands, raising concerns about whether this technology is effective under field conditions (Scott et al., 2013; Yu et al., 2016; Cooper et al., 2019). Given this assessment, there is not only a need for alternatives that avoid the use of chemical pesticides but also increasing interest in finding alternatives to GM-based measures (Dalakouras et al., 2020; Das and Sherif, 2020). A breakthrough was achieved by demonstrating that dsRNAs effectively induce PTGS and confer disease resistance upon foliar spray (Nature, 2016), termed spray-induced gene silencing (SIGS) (Table 1; Koch et al., 2016; Wang et al., 2016). Compared to HIGS, SIGS is much faster, cheaper and easier to handle, and it might reach a broader range of hosts. Interestingly, a direct comparison of HIGS and SIGS revealed SIGS to be more efficient under lab conditions (Koch et al., 2019; Höfle et al., 2020). However, the lab-to-field transition will require optimization and further development of SIGS technology to increase stability (UV, rain) and specificity (off-target risks) under environmental conditions. In this review, we will highlight the most promising attempts to further develop SIGS for its lab-to-field transition and discuss significant achievements in improving stability and reducing environmental risks. In addition, we provide an overview of RNA spray approaches and their particularities, including nanomaterial-based delivery, efficiencies and durability.
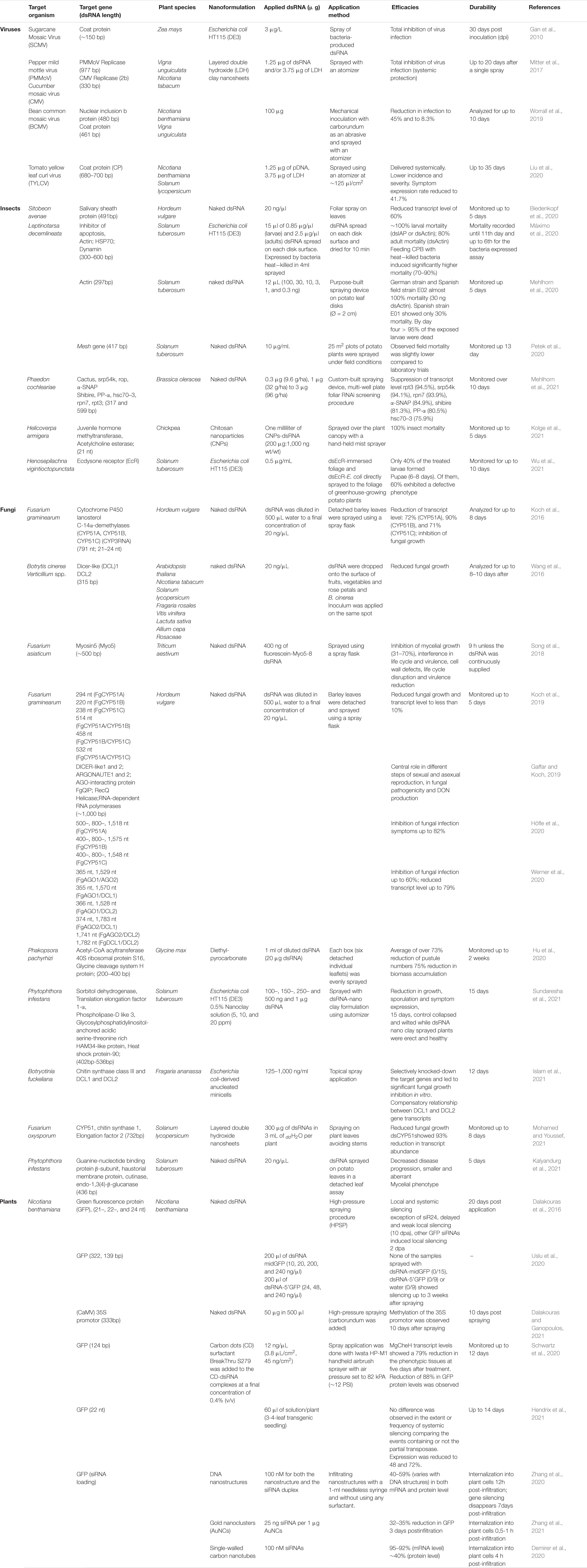
Table 1. Summary of RNA spray studies in plants.
Major Challenges and Recent Achievements in Transferring RNA Spray to Field Environments
The immense potential of sprayable RNA biopesticides for meeting current agro-economic challenges has prompted the development of RNAi technology. Initial euphoria was a major driver leading to a rapid increase of reports, resulting in 54 studies over the last 3 years (PubMed, August 2021) demonstrating academic proof-of-concept for several pathosystems (Table 1). However, at the same time, it became obvious that the molecular mechanism for uptake, processing and transport of sprayed RNA biopesticides was inadequately understood.
Initially, only the GM-based HIGS approach was considered because transgene-derived dsRNA was thought to be robust and reliable in the delivery of PTGS inducers. Naked dsRNA showed effectiveness under lab conditions but had the potential for instability when sprayed in fields, where it could be degraded by UV radiation or washed off by rain. To circumvent this, several studies, inspired by human RNAi therapy research (Swaminathan et al., 2021), attempted to increase stability and durability by establishing protective envelops around dsRNAs and siRNAs (Pugsley et al., 2021; Yan et al., 2021). In mammals, the delivery of siRNA therapeutics is mediated by liposome encapsulation (Liu and Huang, 2021; Zabel et al., 2021). Nanoparticle- and other carrier-based delivery of dsRNA/siRNA has been widely used, with remarkable success, in start-up companies such as “RNAissance1” which focus on broad, cost-efficient topical RNAi uses in agriculture. Their final dsRNA products are advertised as safe, ready to use and stabler than naked dsRNAs. This advancement represents a step toward lowering the amount of applied RNA biopesticides since it is estimated that 2–10 g of dsRNA are required to protect 1 ha (Das and Sherif, 2020). However, the application amount and frequency depend on several largely unknown factors that determine dsRNA persistence, distribution and dilutional and degradational processes in host plants as well as target species. Moreover, it is indisputable that mechanistic insights, which determine strength and limitations in a pathosystem-specific manner, will be required to optimize and further develop RNA sprays, as well as to anticipate obstacles that will appear when transferring RNA sprays to field environments.
Significant Advances of RNA Spray Controlling: Viruses
Given that the RNAi originally evolved as a primary antiviral defense in plants, it is not surprising that most studies have shown that RNAi is most effective at controlling viral pathogens. For example, across 75 studies, HIGS-based RNAi showed an average viral resistance of 90% (Gaffar and Koch, 2019; Koch and Wassenegger, 2021). Notably, similar efficacies were demonstrated when dsRNAs were applied exogenously (Table 1). Conclusive RNAi-based effects were reported in 2016 by Konakalla et al., who demonstrated that the exogenous application of dsRNA targeting the viral silencing suppressor p126 gene and the CP gene of TMV conferred virus resistance in tobacco (Table 1). The authors further showed systemic spreading of dsRNA from local to systemic tissues within an hour of dsRNA application using semi-quantitative RT-PCR (Konakalla et al., 2016). They found that p126 dsRNA levels continuously decreased in the local treated tissue, from 3 to 9 dpi, until dsRNA was no longer detectable. Another study supported these findings, showing that exogenously applied dsRNA derived from the HC-Pro and CP genes of ZYMV protects watermelon and cucumber against ZYMV and that it spread systemically over long distances in cucurbits (Kaldis et al., 2018). Further emphasizing the systemic spread of RNA biopesticides, the movement of sprayed dsRNA from barley leaves to stems and root tissues was demonstrated within 3 days after spray treatment (Biedenkopf et al., 2020). Systemic distribution is of key importance because it indicates that RNA biopesticides could be promising substitutes for systemic pesticides. Moreover, translocation from leaves (application sites) to roots suggests that foliar sprays may be able to target soil-borne infections, thus circumventing the need to develop soil-specific RNA treatments.
Interestingly, the induction of virus resistance by exogenous application had already been achieved 20 years ago (Tenllado and Díaz-Ruíz, 2001). However, “exogenous” does not necessarily imply a spray application. Thus, we must distinguish between different application strategies, as most of them work on a lab scale but are unsuitable and unpracticable for field applications (Table 1).
As already indicated, numerous exogenous application strategies rely on preparatory treatments, such as the mechanical inoculation of leaves, to guarantee efficient dsRNA uptake (Table 1). The rubbing or dusting of leaf surfaces with Carborundum (silicon carbide) as an abrasive is widely used as a pretreatment (Tenllado and Díaz-Ruíz, 2001; Tenllado et al., 2003; Yin et al., 2009; Konakalla et al., 2016; Necira et al., 2021). Given the unsuitability of such pretreatments on the field scale, subsequent research, inspired by human RNAi therapeutics where nanotechnology is unavoidable, has attempted to formulate dsRNA for efficient and targeted RNA delivery. Such formulations can help to increase cellular uptake, improve stability (and thus overcome environmental degradation by UV radiation or surface wash-off) and provide long-term protection against the targeted pathogen. Toward this, the first breakthrough was achieved by Mitter et al. (2017) by using positively charged layered double hydroxide (LDH) clay nanosheets as a dsRNA carrier (BioClay). This technology was originally developed for the delivery of siRNA therapeutics to mammalian cells (Ladewig et al., 2009, 2010). The authors found that loading dsRNA on LDH prolonged durability on the leaf surface for 30 days and increased stability through protection from nuclease degradation (Mitter et al., 2017). Moreover, they showed the uptake of dsRNA into plant cells and induction of endogenous RNA silencing, which mediated systemic protection against the targeted VSR 2b gene of CMV on cowpea and tobacco. In field BioClay allows sustained release of dsRNA on the leaf surface under ambient conditions (Ram Reddy et al., 2006; Xu et al., 2006; Reddy et al., 2008; Mitter et al., 2017). Notably, this study proved that the LDH nanocarrier can be completely degraded over time, thus resulting in a slow and sustained release of dsRNA under environmental conditions (Mitter et al., 2017). After this pioneering work on the biodegradability of clay-based nanomaterials, dsRNA-BioClay has been used to target the bean common mosaic virus (BCMV) to protect N. benthamiana and Vigna unguiculata (Worrall et al., 2019).
While this study provides a significant step forward in making RNA spray an applicable and sustainable approach for pathogen and pest control in agriculture, questions remain about how to produce efficient amounts of dsRNA for spray applications in field trials. Initially, Escherichia coli-based dsRNA production was used (Tenllado and Díaz-Ruíz, 2001; Tenllado et al., 2003; Yin et al., 2009; Gan et al., 2010) due to financial constraints and to the poor availability of suitable dsRNA synthesis kits. Interestingly, the latest studies showed that E. coli cannot only be used as a dsRNA factory but also provides adequate properties for dsRNA encapsulation (Islam et al., 2021; Necira et al., 2021). The authors of these studies concluded that the effects of E. coli-encapsulated dsRNA did not differ from the topical application of naked dsRNAs (Necira et al., 2021; Table 1). E. coli-encapsulation may nevertheless prove superior to naked dsRNA, as it provides a protective envelope conferring higher stability under field conditions, though host species are limited.
Numerous studies have addressed the question of how to reduce the production costs of dsRNA. Some have synthesized dsRNA using in vitro transcription kits, suitable for lab experiments but inconceivable for large-scale applications given their bad price-performance ratio ($700/mg). Thus, subsequent attempts have focused on large-scale production and on the purification of sprayable RNAs to make them commercially competitive and economically achievable, with great success. For example, recent research efforts have rapidly produced cost-effective, large-scale microbial-based dsRNA production using bacteria, such as E. coli (Voloudakis et al., 2015; Ahn et al., 2019; Bento et al., 2020; Niño-Sánchez et al., 2021) and Pseudomonas syringae (Niehl et al., 2018) and the yeast Yarrowia lipolytica (Timmons et al., 2001; Palli, 2014). Given these significant achievements, dsRNA costs per gram have dropped from $12,500 in 2008 to $100 in 2016, $60 in 2020 and finally $2 in 2021 (de Andrade and Hunter, 2016; Zotti et al., 2018; Dalakouras et al., 2020). Recently, large-scale cell-free production has further lowered the price to less than $0.50 per gram, making RNAi competitive in the market.2
Based on the plethora of proof-of-concept studies and recent achievements in nanomaterial-based dsRNA and siRNA delivery, which allow for highly effective virus control even under field conditions, fundamental knowledge on the molecular mechanisms and factors that determine uptake (over cuticle and cellular barriers), processing and translocation has begun to emerge.
Significant Advances of RNA Spray Controlling: Fungi
Since 2010 the number of HIGS-based studies demonstrating fungal disease control has continued to increase (Nowara et al., 2010; Koch and Wassenegger, 2021). The first reports that showed that RNA spray can fight fungus were directed against two necrotrophic ascomycetes Fusarium graminearum (Koch et al., 2016) and Botrytis cinerea (Wang et al., 2016). These case studies further energized the debate on whether RNA spray is a realistic approach for future field applications. Beyond cost concerns, the obvious question was how to enhance uptake of RNA biopesticides by plants, thus avoiding degradation under environmental conditions. The uptake of dsRNA depends on leaf surface stability, and efficient and subsequent uptake can prevent premature degradation or wash-off by rain. Notably, sprayed RNAs must overcome several physical and cellular barriers to reach their cognate mRNA targets. First, they must overcome the “outside-inside” or cuticle barrier at the leaf surface, which is especially relevant when targeting pathogens that replicate and grow intra- or intercellularly, such as viruses and fungi, or sap-sucking insects like aphids or whiteflies. Like viral pathogens, fungi maintain an intimate relationship with their hosts, acquiring nutrients in a biotrophic, hemibiotrophic, or necrotrophic manner. Given their different lifestyles, fungi may take up sprayed RNAs from extra-, inter- as well as intracellular space. Passing through this barrier was assumed to be passively facilitated by stomata opening (Koch et al., 2016). Given this assumption, one could ask whether stomata density and leaf architecture represent limiting factors in RNA uptake. This would explain why SIGS works well for some plant species and not for others. Considering this, formulations that facilitate the opening of stomata or increase leaf permeability may help to increase dsRNA uptake from foliar sprays.
However, before sprayed dsRNA can enter plant cells for processing and translocation, they reach another barrier, the “apoplast-symplast” interface. Entering plant cells is necessary for processing into siRNAs by plant DCLs. However, in particular cases, the uptake of unprocessed dsRNA may also occur, depending on the lifestyle of the targeted pathogen. For example, SIGS of the necrotroph F. graminearum require the uptake of unprocessed dsRNA precursors and their subsequent processing by the fungal RNAi machinery (Koch et al., 2016; Gaffar et al., 2019). Based on these findings, it has been hypothesized that the uptake of long, unprocessed dsRNA and its processing into many different inhibitory siRNAs by the target organism might lead to higher gene silencing efficiencies and increased disease resistance (Koch et al., 2019). However, there is some controversy about the uptake of dsRNA versus siRNA, for instance in terms of the relative efficacies and off-target risks (Koch and Wassenegger, 2021). Interestingly, preliminary data suggest that if sprayed dsRNA is too long, the length might interfere with sufficient cellular uptake (Höfle et al., 2019). To prove whether uptake of unprocessed dsRNA really confers stronger effects, it will be helpful to compare SIGS efficacies of biotrophic and necrotrophic pathogens. So far, the disease resistance level achieved by SIGS has been comparable (Hu et al., 2020) or even superior (Höfle et al., 2019; Koch et al., 2019) to HIGS-based efficiencies under lab conditions.
Interestingly, a recent study that assessed the dsRNA uptake ability of different fungi revealed that dsRNA uptake efficiencies varied across the tested fungal and oomycete species (Qiao et al., 2021). For example, the authors demonstrated efficient dsRNA uptake for Botrytis cinerea, Sclerotinia sclerotiorum, Rhizoctonia solani, Aspergillus niger, and Verticillium dahliae. They found no uptake for Colleotrichium gloeosporioides, modest uptake for the non-pathogenic fungus Trichoderma virens and limited uptake for the oomycete Phytophthora infestans (Qiao et al., 2021). This study is of great significance, as it confirms and extends previous findings (Koch et al., 2016; Wang et al., 2016; McLoughlin et al., 2018) and further supports the idea that necrotrophic fungi exhibit a stronger response to exogenous RNA applications. Whether this is because they can take up unprocessed dsRNA, which may lead to higher gene silencing efficiencies - based on the finding that F. graminearum DCLs are required for SIGS (Koch et al., 2016; Gaffar et al., 2019) – or whether the overall amount of dsRNA absorbed in less time correlates with their necrotrophic lifestyle needs further verification. Supporting this idea, Hailing Jin and her team observed dsRNA uptake as early as 6 h after YFP-dsRNA in vitro treatment of the necrotrophs B. cinerea, S. sclerotinia, and R. solani, suggesting faster dsRNA uptake compared to A. niger (10 hpt) and V. dahliae (12 hpt). In addition, they demonstrated the antifungal activity of topically applied dsRNAs targeting fungal vesicle pathway genes or DCL genes in B. cinerea, S. sclerotinia, R. solani, and A. niger (Qiao et al., 2021). However, as the dsRNAs were dropped onto the surface of each plant or fruit sample and the fungi were applied directly to the dsRNA-treated area, we do not know from this study whether the uptake of plant DCL-processed siRNA would have made a difference. In other words, the type of RNA offered for uptake was limited to dsRNA, bringing us back to the initial question of whether the uptake of unprocessed dsRNA is a crucial determinant of SIGS efficacy, and thus whether the uptake of dsRNA by the plant represents a limiting factor regarding the durability of protective RNA spray effects.
Regarding this question, we need to differentiate between sprayed RNAs that stick to the plant (or fruit or vegetable) surface and those that enter plant cells. We predict that surface interaction of dsRNA or siRNA with microbial targets will require intense stabilization formulas, with potential risks for environmental accumulation and pollution. Cellular uptake, on the other hand, may have a protective effect, but promoting fast processing or even bearing the risk of enzymatic and lytic degradation together with dilution over distance. Although we seek RNA-stabilizing formulas, at the same time we question RNA degradability as a selling point. Recently, dsRNA spray effects against B. cinerea infection were found to last for 7 days after spraying onto tomato plants in a greenhouse, indicating a quite rapid degradation. Notably, the authors of this study concluded that dsRNA gets degraded by the environment, suggesting no cellular uptake by the plant (Qiao et al., 2021). Although short longevity can be determined by degradation, it remains unknown whether this degradation occurs outside or inside the sprayed plant.
So far, most SIGS studies demonstrating the control of fungal pathogens were conducted with naked, un-formulated dsRNA (Table 1). However, formulations that increase cellular uptake (especially for plants that showed no absorption of naked dsRNA from their surfaces) or surface stability (especially for post-harvest products such as fruits and vegetables) will boost broad applicability and the lab-to-field transition. A recent study showed that E. coli-derived anucleated minicells can be used as a cost-effective platform for dsRNA production and encapsulation, shielding dsRNA from RNase degradation (Islam et al., 2021). The authors demonstrated that the protection of strawberries from Botryotinia fuckeliana infection was prolonged to 12 days under greenhouse conditions, further emphasizing the added value of encapsulation for dsRNA stability. Although extending protection against the gray mold of tomatoes by 7 days (naked dsRNA) (Qiao et al., 2021) and strawberries by 12 days (Minicell-based) (Islam et al., 2021) already represent positive results, there is still room for further improvement. Thus, data on durability and persistence together with systemic distribution are highly desired for “lab-to-field” technology transfers. Moreover, we need to know which parameters promote or restrict dsRNA and siRNA uptake in a plant species-specific context at the “outside-inside” barrier and the “apoplast-symplast” interface, as well as in a pathosystem specific manner at the “plant-fungus” interface. However, the mechanism by which sprayed RNA overcomes the apoplast-symplast barrier is largely unknown. It is hypothesized that endocytosis and extracellular vesicles (EVs) play a crucial role in the uptake and translocation of HIGS- and SIGS-associated RNAs (Wytinck et al., 2020a; Koch and Wassenegger, 2021; Santos et al., 2021). Notably, recent data suggest that SIGS, in contrast to HIGS, might not involve EVs for siRNA delivery and uptake, at least in the F. graminearum-barley pathosystem (Schlemmer et al., 2021a,b). Based on insights into the mode of uptake, we have the chance to optimize and develop dsRNA and siRNA delivery and cellular uptake in the future.
Significant Advances of RNA Spray Controlling: Insects
Host-induced gene silencing-mediated control of insect pests has been proven effective with an average of 50% conferred resistance (Koch and Wassenegger, 2021). Exogenous dsRNA and siRNA applications (e.g., feeding, injection, and oral delivery) to various insect pests and mites is routinely conducted to study gene function or identify RNAi targets (Lü et al., 2020; Mehlhorn et al., 2020, 2021; Máximo et al., 2020). In line with the above discussion about RNAs that remain on sprayed surfaces and those that enter plant cells, insect pests and mites can ingest dsRNA and siRNAs by chewing-biting or piercing-sucking feeding behaviors. In addition to the control of viruses and fungi, insect pests are also generally accessible to RNA sprays (Table 1). For example, a recent report demonstrated the efficiency of RNA sprays against Henosepilachna vigintioctopunctata (28-spotted ladybird) in the greenhouse (Wu et al., 2021). Spraying E. coli-expressed dsRNAs targeting the ecdysone receptor (EcR) gene onto the foliage of greenhouse-growing potato plants provoked the death of third and fourth instar larvae and reduced leaf damage (Wu et al., 2021). Confirming these results, another study showed exogenous dsRNA application as a promising alternative to chemical pesticides for controlling H. vigintioctopunctata (Lü et al., 2021). RNA sprays have also been shown to be effective in controlling the Colorado potato beetle, Leptinotarsa decemlineata (Mehlhorn et al., 2020). Even more importantly, the authors investigated geographical variation in RNAi sensitivity in the second instar larvae of 14 different European populations of field-collected L. decemlineata and found only minor variability in RNAi sensitivity between populations. This baseline study provides the first valuable insights on the broad applicability and transferability of RNA sprays over a geographic range in Europe (Mehlhorn et al., 2020). While the accessibility of phyllophagous pests by RNA sprays may seem trivial, a breakthrough was achieved by providing the first laboratory evidence that feeding dsRNA-coated oilseed rape buds to the pollen beetle Brassicogethes aeneus diminishes pollen beetle survival rate (Willow et al., 2021). This study is significant because many pathogens and pests that tremendously impact agriculture infect ears and buds rather than leaves.
Given the fact that we lack reliable results from field trials, or even from data generated under simulated field test conditions, lab-to-field transfer will require improvements of stability and adherence of dsRNA and siRNA to resist UV radiation and rain wash-off. For these purposes, encapsulation is pursued to guarantee and prolong the longevity of RNA sprays in the field. For example, E. coli is not only utilized for cost-effective dsRNA production but also represents a protective envelope for efficient dsRNA delivery (Lü et al., 2020; Máximo et al., 2020; Wang et al., 2021; Wu et al., 2021). Interestingly, encapsulation of dsRNA, if large enough, may prevent cellular uptake (plant intake) and thus shield dsRNA from plant DCL processing, as indicated by previous breakthroughs demonstrating that the expression of dsRNA in chloroplasts (DCL-free organelles) targeting the ß-actin gene of L. decemlineata (Colorado potato beetle) caused larval lethality (Zhang et al., 2015).
Targeting piercing-sucking insects by RNA spray requires cellular uptake and systemic distribution via the phloem. Thus, knowledge of the paths used by dsRNA and siRNA as SIGS inducers is a prerequisite for further developing and applying RNA sprays to the field. Given this assumption, previous reports revealed the systemic spread of sprayed RNAs (Koch et al., 2016; Konakalla et al., 2016; Kaldis et al., 2018; Biedenkopf et al., 2020). Spraying a fluorescent-labeled dsRNA onto barley leaves and subsequently examining longitudinal leaf sections revealed that the fluorescence was not confined to the apoplast but also was present in the symplast of phloem parenchyma cells, companion cells and mesophyll cells, as well as in trichomes and stomata (Koch et al., 2016). The finding that RNA biopesticides systemically spread through the phloem was confirmed by spraying fluorescent-labeled dsRNA followed by phloem sampling by stylectomy (excision of a stylet, typically that of an aphid) of the distal, non-sprayed leaf parts. Using CLSM, a green fluorescent signal was detected after cutting off the stylet tip of feeding aphids (Biedenkopf et al., 2020). Even more importantly, it was shown that sprayed dsRNA moved from barley leaves over stems to the root tissue within 3 days of spray treatment (Biedenkopf et al., 2020). This finding is especially interesting for the control of root pathogens because so far dsRNA applied to soil has immediately degraded (Dubelman et al., 2014; Parker et al., 2019; Bachman et al., 2020; Qiao et al., 2021). However, systemic distribution may result in dilution of SIGS signals; thus, further research is needed to prove the activity of dsRNA and siRNA in systemic tissues and to address the question of how much or how often RNA biopesticides need to be applied to confer robust and durable SIGS-based disease resistance. Given these challenges, previous studies have proposed utilizing the symbiosis of plants with bacteria, for example by engineering symbionts that serve as dsRNA vectors to maintain long-lasting dsRNA production in planta (Whitten et al., 2016; Whitten and Dyson, 2017). However, emerging data suggest that pest control via RNA sprays does not necessarily require plant passage for uptake and SIGS induction (Thairu et al., 2017; Niu et al., 2019; Linyu et al., 2021). Due to the limited uptake of dsRNA in hemiptera insects (Niu et al., 2019) originally developed and described gene silencing induction in aphids upon foliar spray on pea aphids themselves to verify potent RNAi targets in aphids. More recently, another confirmatory report used nanocarrier SPc (star polycation) transdermal delivery systems to verify gene silencing efficiency of selected RNAi targets in Aphis gossypii (Linyu et al., 2021). However, both studies imply sensitivity of RNA aphid sprays, which may circumvent plant passage and uptake from the phloem when applied in the field. Moreover, the development of formulations that allow transdermal delivery in a species-specific context is very valuable, especially in light of risk assessment and regulations (avoiding off-target effects).
We predict that nanomaterial-based formulations can solve major challenges regarding lab-to-field transitions by increasing stability and warranting specificity and selectivity. Not only surface stability, but also rather the ability to resist enzymatic degradation by RNase or extreme pH-values, which are prominent in saliva and gut of insects (Christiaens et al., 2014; Peng et al., 2020a,b). Given these challenges, a recent breakthrough was achieved, demonstrating that chitosan nanoparticles-mediated dsRNA delivery in Helicoverpa armigera protects from degradation by nucleases and insect gut pH (Kolge et al., 2021). Beyond shielding dsRNA from RNase degradation, the silencing of a dsRNA ribonuclease improved oral RNAi efficacy in the southern green stinkbug (Sharma et al., 2021). This finding indicates that the simultaneous application of protective agents such as RNase inhibitors or the silencing of dsRNases might encourage targeted detoxification mediated by P450s, ABC transporters and others (Wang et al., 2021). Notably, a nanomaterial-based formulation may help to increase selectivity not only by shielding dsRNA from the environment but also by providing target-site-specific RNA release (e.g., pH-dependent release kinetics), or selectivity facilitated by attractants that are incorporated in the envelope of (nano)-capsules following the proven attract and kill principle.
Significant Advances of RNA Spray Silencing Plant Genes
Overcoming cuticle and cellular barriers is a prerequisite if RNA sprays are to substitute for conventional chemical herbicides or be used to study the gene function of plants. This is fundamentally different from RNA biopesticides that lay on plant, fruit or vegetable surfaces, in which cases sprays of naked dsRNA exhibit strong effects, for example, in controlling B. cinerea and Botryotinia fuckeliana (Wang et al., 2016; Islam et al., 2021; Qiao et al., 2021). However, previous attempts at spraying naked dsRNA to silence the expression of transgenes gave contrasting results, starting a controversial debate on the future herbicide uses of RNA sprays (Uslu and Wassenegger, 2020). For example, uptake of naked dsRNA and transgene silencing was demonstrated for the model plant Arabidopsis thaliana (Mitter et al., 2017; Dubrovina and Kiselev, 2019). However, attempts to silence transgene-expressed green fluorescence protein (GFP) in Nicotiana benthamiana by a high-pressure spray of naked dsRNA failed (Uslu et al., 2020). Notably, high-pressure sprays were specifically developed for RNAi applications and usage in N. benthamiana, resulting in transgene silencing (Dalakouras et al., 2016, 2018). Remarkably, based on their RNA-seq results the authors concluded that failure in transgene silencing correlates with the absence of dsRNA-derived specific siRNAs (Uslu et al., 2020). This finding raises the question of whether there was cellular uptake of high-pressure sprayed dsRNA, which is required for processing by plant DCLs. Notably, the spraying of 22 nt synthetic siRNA (matching the GFP sequence, position 164–187) as a positive control induced transgene silencing, as previously described (Dalakouras et al., 2016). Regarding this, another recent report showed induced methylation of the 35S promotor (which controls GFP transgene expression) upon 35S-dsRNA high pressure spray indicating cellular uptake of dsRNA (Dalakouras and Ganopoulos, 2021). Nevertheless, in contrast to siRNA spraying, cellular uptake of sprayed long dsRNAs seems to be less efficient, especially entering the nucleus to trigger RNA-directed DNA methylation is challenging. Moreover, another previous study demonstrated cellular uptake and plant DCL-mediated siRNA generation of a naked dsRNA sprayed onto barley leaves; however, this did not target a transgene (Koch et al., 2016). Nevertheless, these findings clearly illustrate how different plant species respond to RNA sprays, thus exemplifying the importance and urgency of research into the mechanistic basis (uptake, processing, and translocation) of RNA spray applications.
As much as spraying naked dsRNA highlights the simplicity of this technology, there are limits and restrictions that we have just begun to unravel. Given that sufficient RNA delivery is a continuing challenge in RNA spray applications, significant advances in nanomaterial-based formulations and the development of other smart delivery platforms have boosted technological development. Utilizing different biological, physical, and chemical-assisted delivery methods such as Bacterium-mediated RNAi (Goodfellow et al., 2019), high-pressure sprays (Dalakouras et al., 2016, 2018; Uslu et al., 2020), lipid nanoparticles, cationic polymers, cell-penetrating peptides and clay nanosheets has been shown to provide protection from nucleases and pH and to improve cellular uptake increasing plant resistance to pathogens and pests (Table 1). Recent studies have demonstrated efficient siRNA delivery into intact plants facilitated by nanomaterials such as carbon dots (Schwartz et al., 2020), single-walled carbon nanotubes (Demirer et al., 2019, 2020), DNA nanostructure carriers (Zhang et al., 2019, 2020) and gold-nanoclusters (Zhang et al., 2021; Table 1). Based on their small size (3.9 nm) carbon dots can pass through the cell wall (size exclusion limit 3–10 nm) (Carpita et al., 1979) mediating siRNA delivery into plant cells (Schwartz et al., 2020). Notably, passing through a plant cell wall is not the only prerequisite for efficient silencing. As discussed above, RNAi effectors face several barriers in reaching their mRNA cognates. Thus, cellular uptake requires overcoming the apoplast-symplast barrier mediated by endocytosis through the plasma membrane and subsequent release from endomembrane vesicles (Wytinck et al., 2020b). Given these later barriers, the properties of carbon dots are found to be suboptimal (Schwartz et al., 2020). Although carbon-dots-mediated siRNA delivery needs further improvement for efficient endogenous gene silencing, the delivery of dsRNAs over plant cell walls that reach the apoplast might be sufficient for controlling pathogens that require the uptake of unprocessed dsRNAs for SIGS induction (Koch et al., 2016; Gaffar et al., 2019). Due to their ultrasmall size (2 nm) and easier and faster synthesis, gold-nanoclusters (AuNCs) have been adapted for the delivery of siRNAs in mature plants, inducing efficient transgene as well as endogenous gene silencing in N. benthamiana (Zhang et al., 2021).
However, as naked dsRNA/siRNA delivery is restricted by largely unknown parameters that prevent RNA sprays from reaching their full potential, usage of different delivery methods, as advantageous and promising they may be, have limitations, as indicated above (for a detailed comparison of delivery platforms, see also: Zhang et al., 2021). Regarding this, identification of chemicals (e.g., Sortin1) that increased RNAi potency by enhancing siRNA accumulation and loading into AGOs may boost the development of powerful RNA biopesticide formulations (Jay et al., 2019). Moreover, while all of these studies provided proof-of-concept, the suitability of nanomaterial-based RNAi effector delivery as well as application of chemical enhancers of PTGS needs to be approved under field conditions. In addition, toxicity and biodegradability, together with applicability to other plant species and transferability for delivering dsRNA targeting foreign genes, needs further research in order to be verified.
Conclusion and Future Perspectives
Despite the numerous proof-of-concept studies demonstrating the great potential underlying RNAi-based plant protection, especially GMO-free RNA sprays, we are still far from field applications or even product launches. Currently, we are facing complex and multi-layered challenges comprising technical-biological prospects (stability, selectivity, and broad applicability) as well as social-political demands (acceptance and regulations). To meet these challenges, significant advances in developing RNA sprays that guarantee stability in the field and minimize off-target risks for non-target organisms have been made, for example, by using nanomaterial-based formulations and the fusion of different technologies. However, we lack a sufficient data basis for understanding how nano-formulated RNA biopesticides will behave under field conditions. Moreover, we urgently require knowledge on durability, persistence and systemic effects of RNA sprays to develop recommendations for handling in the field. For example, when and how often do RNA biopesticides need to be applied or renewed to confer maximum protection? Based on the answer, we could calculate real costs for each pathosystem. To achieve broad applicability, which is often used as a selling point over GMO-based RNAi strategies (HIGS), we need to clarify the question of why RNA sprays work well for some pathosystems and not for others. If we know what determines SIGS efficiencies, we may have a starting point for developing solutions that demonstrate broad transferability. Elucidating the molecular mechanisms of uptake, processing and translocation of spray-delivered RNAs is crucial for making RNA sprays realistic and achievable in future field applications. In addition, emerging data that suggest extending RNA sprays to exploit the mechanisms of TGS that will allow epigenetic modifications of plant genes (Dalakouras and Vlachostergios, 2021) should be considered and discussed regarding the potential risk of introducing unintended off-target effects by modifying the plant’s epigenome (Dalakouras and Papadopoulou, 2020).
In contrast to lab experiments, where dsRNA and siRNA effects are tested in isolation and on small scales, field environments are unpredictable and highly dynamic. For example, the occurrence of pathogens and pests, as well as their coincidences, represents a major challenge for determining application time and frequencies. Moreover, plants grown under environmental conditions develop differently from plants grown in greenhouses, especially in terms of cuticle properties, which may impact adherence, stability, and cellular uptake. Thus, we need smart application and formulation strategies, such as multi-targeting and site-directed targeting, to ensure high efficacies on the species level. For example, combining RNA biopesticides with distinct molecular targets may not only confer stronger effects but also allow high efficiencies on a species-specific scale. In other words, the majority of highly effective dsRNAs and siRNAs target highly conserved genes, which bear high off-target risks. They are suitable candidates when conducting proof-of-concept studies but are inappropriate for field trials. Thus, identifying molecular targets on species level and their simultaneous combinatory (multi-targeting) applications may exhibit strong effects and at the same time minimize off-target risks. However, lab-to-field transitions require further precision and accuracy in off-target predictions and their controllability for adequate risk assessment. Currently, data requirements specific for RNA spray-based plant protection products are not yet in place under the regulatory frameworks for pesticides and plant protection products in the United States and EU (Dietz-Pfeilstetter et al., 2021). The OECD (Organization for Economic Cooperation and Development) organized a meeting on this question, which represented a major step forward in terms of testing regulation and research direction for future external dsRNA products (OECD, 2020). It is important to note that siRNAs display small gene regulatory units, which needs to be considered when developing directives that ensure appropriate risk assessment. Consistent with this, a recent report reviewed the evidence for and against the transfer of diet-derived miRNAs from plants, meat, milk and exosome and their putative molecular regulator roles as well as pharmacological opportunities for cross-kingdom regulation in the consuming organism (Mar-Aguilar et al., 2020; del Pozo-Acebo et al., 2021). The authors concluded that the transfer of miRNAs from the diet to the blood is still inconclusive and that the main source of controversy in plant studies is the lack of reproducibility of the findings. Currently, a risk assessment by the European Food Safety Authority (EFSA) classified RNA biopesticides as safe regarding low risks that sprayable RNAs pose for animals/humans (Olivier et al., 2018). The decisive argument was that oral uptake of RNAi products by consumers bears a low risk for interference with gene expression in humans, as too many biological and physical barriers have to be overcome (Schiemann et al., 2019; Kleter, 2020). However, so far, the available scientific data on environmental, consumer and user safety evaluations of RNA spray applications are still scarce. Preliminary data on the environmental fate (persistence and degradability) of sprayed dsRNA suggest a short shelf-life after application and absorption to soil (Parker et al., 2019; Bachman et al., 2020). However, ongoing nanomaterial-based attempts to increase stability may prolong environmental persistence and increase the risk to local ecosystems. In addition, it is still unclear whether and to what extent sprayed dsRNAs and siRNAs can accumulate along the food chain and how nanomaterials affect this accumulation. Thus, the environmental risk assessment of RNA biopesticides needs to be reinforced in order to facilitate their future placement on the market. Moreover, the length of the pesticide authorization process should be reduced to support a just transition. Given this, it is just a matter of time since first RNA biopesticides (e.g., Monsanto’s/Bayer’s BioDirectTM technology) will access the market. Finally, we need to develop educational work and information campaigns for public outreach and transparency as early as possible.
Author Contributions
Both authors contributed to wrote the manuscript and approved the submitted version.
Funding
This work was supported by the Ministry for Food and Rural Areas, Baden-Württemberg (Grant Number: 54-8214.07-FP20-137/1) to AR and AK.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Frontiers in Plant Science for invitation.
Footnotes
References
Ahn, S.-J., Donahue, K., Koh, Y., Martin, R. R., and Choi, M.-Y. (2019). Microbial-Based Double-Stranded RNA Production to Develop Cost-Effective RNA Interference Application for Insect Pest Management. Int. J. Insect Sci. 11:117954331984032. doi: 10.1177/1179543319840323
Bachman, P., Fischer, J., Song, Z., Urbanczyk-Wochniak, E., and Watson, G. (2020). Environmental Fate and Dissipation of Applied dsRNA in Soil, Aquatic Systems, and Plants. Front. Plant Sci. 11:21. doi: 10.3389/fpls.2020.00021
Baum, J. A., Bogaert, T., Clinton, W., Heck, G. R., Feldmann, P., Ilagan, O., et al. (2007). Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 25, 1322–1326. doi: 10.1038/nbt1359
Bento, F. M. M., Marques, R. N., Campana, F. B., Demétrio, C. G. B., Leandro, R. A., Parra, J. R. P., et al. (2020). Gene silencing by RNAi via oral delivery of dsRNA by bacteria in the South American tomato pinworm, Tuta absoluta. Pest Manag. Sci. 76, 287–295. doi: 10.1002/ps.5513
Biedenkopf, D., Will, T., Knauer, T., Jelonek, L., Furch, A. C. U., Busche, T., et al. (2020). Systemic spreading of exogenous applied RNA biopesticides in the crop plant Hordeum vulgare. ExRNA 2:12. doi: 10.1186/s41544-020-00052-3
Carpita, N., Sabularse, D., Montezinos, D., and Delmer, D. P. (1979). Determination of the pore size of cell walls of living plant cells. Science 205, 1144–1147. doi: 10.1126/science.205.4411.1144
Christiaens, O., Swevers, L., and Smagghe, G. (2014). DsRNA degradation in the pea aphid (Acyrthosiphon pisum) associated with lack of response in RNAi feeding and injection assay. Peptides 53, 307–314. doi: 10.1016/j.peptides.2013.12.014
Cooper, A. M. W., Silver, K., Zhang, J., Park, Y., and Zhu, K. Y. (2019). Molecular mechanisms influencing efficiency of RNA interference in insects. Pest Manag. Sci. 75, 18–28. doi: 10.1002/ps.5126
Dalakouras, A., and Ganopoulos, I. (2021). Induction of Promoter DNA Methylation Upon High-Pressure Spraying of Double-Stranded RNA in Plants. Agron 11:789. doi: 10.3390/agronomy11040789
Dalakouras, A., and Papadopoulou, K. K. (2020). Epigenetic Modifications: an Unexplored Facet of Exogenous RNA Application in Plants. Plants 9:673. doi: 10.3390/plants9060673
Dalakouras, A., and Vlachostergios, D. (2021). Epigenetic approaches to crop breeding: current status and perspectives. J. Exp. Bot. 72, 5356–5371. doi: 10.1093/jxb/erab227
Dalakouras, A., Jarausch, W., Buchholz, G., Bassler, A., Braun, M., Manthey, T., et al. (2018). Delivery of hairpin RNAs and small RNAs into woody and herbaceous plants by trunk injection and petiole absorption. Front. Plant Sci. 9:1253. doi: 10.3389/fpls.2018.01253
Dalakouras, A., Wassenegger, M., Dadami, E., Ganopoulos, I., Pappas, M. L., and Papadopoulou, K. (2020). Genetically modified organism-free RNA interference: exogenous application of RNA molecules in plants. Plant Physiol. 182, 38–50. doi: 10.1104/pp.19.00570
Dalakouras, A., Wassenegger, M., McMillan, J. N., Cardoza, V., Maegele, I., Dadami, E., et al. (2016). Induction of silencing in plants by high-pressure spraying of In vitro-synthesized small RNAs. Front. Plant Sci. 7:1327. doi: 10.3389/fpls.2016.01327
Das, P. R., and Sherif, S. M. (2020). Application of Exogenous dsRNAs-induced RNAi in Agriculture: challenges and Triumphs. Front. Plant Sci.11:946. doi: 10.3389/fpls.2020.00946
de Andrade, E. C., and Hunter, W. B. (2016). “RNA Interference – Natural Gene-Based Technology for Highly Specific Pest Control (HiSPeC)” in RNA Interference, ed. Y. A. Ibrokhim (InTechOpen). doi: 10.5772/61612
del Pozo-Acebo, L., López de las Hazas, M.-C., Margollés, A., Dávalos, A., and García-Ruiz, A. (2021). Eating microRNAs: pharmacological opportunities for cross-kingdom regulation and implications in host gene and gut microbiota modulation. Br. J. Pharmacol. 178, 2218–2245. doi: 10.1111/bph.15421
Demirer, G. S., Zhang, H., Goh, N. S., Pinals, R. L., Chang, R., and Landry, M. P. (2020). Carbon nanocarriers deliver siRNA to intact plant cells for efficient gene knockdown. Sci. Adv. 6:eaaz0495. doi: 10.1126/sciadv.aaz0495
Demirer, G. S., Zhang, H., Matos, J. L., Goh, N. S., Cunningham, F. J., Sung, Y., et al. (2019). High aspect ratio nanomaterials enable delivery of functional genetic material without DNA integration in mature plants. Nat. Nanotechnol. 14, 456–464. doi: 10.1038/s41565-019-0382-5
Dietz-Pfeilstetter, A., Mendelsohn, M., Gathmann, A., and Klinkenbuß, D. (2021). Considerations and Regulatory Approaches in the USA and in the EU for dsRNA-Based Externally Applied Pesticides for Plant Protection. Front. Plant Sci. 12:682387. doi: 10.3389/fpls.2021.682387
Douglas, A. E. (2018). Strategies for Enhanced Crop Resistance to Insect Pests. Annu. Rev. Plant Biol. 69, 637–660. doi: 10.1146/annurev-arplant-042817-040248
Dubelman, S., Fischer, J., Zapata, F., Huizinga, K., Jiang, C., Uffman, J., et al. (2014). Environmental fate of double-stranded RNA in agricultural soils. PLoS One 9:e93155. doi: 10.1371/journal.pone.0093155
Dubrovina, A. S., and Kiselev, K. V. (2019). Exogenous RNAs for gene regulation and plant resistance. Int. J. Mol. Sci. 20:ijms20092282. doi: 10.3390/ijms20092282
European Commission. (2021). EU Biodiversity Strategy for 2030. Available online at: https://op.europa.eu/en/publication-detail/-/publication/31e4609f-b91e-11eb-8aca-01aa75ed71a1 (accessed September 2021).
Fire, A., Xu, S., Montgomery, M. K., Kostas, S. A., Driver, S. E., and Mello, C. C. (1998). Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391, 806–811. doi: 10.1038/35888
Fotoukkiaii, S. M., Wybouw, N., Kurlovs, A. H., Tsakireli, D., Pergantis, S. A., Clark, R. M., et al. (2021). High-resolution genetic mapping reveals cis-regulatory and copy number variation in loci associated with cytochrome P450-mediated detoxification in a generalist arthropod pest. PLoS Genet. 17:e1009422. doi: 10.1371/journal.pgen.1009422
Gaffar, F. Y., and Koch, A. (2019). Catch me if you can! RNA silencing-based improvement of antiviral plant immunity. Viruses 11:v11070673. doi: 10.3390/v11070673
Gaffar, F. Y., Imani, J., Karlovsky, P., Koch, A., and Kogel, K.-H. (2019). Different Components of the RNA Interference Machinery Are Required for Conidiation, Ascosporogenesis, Virulence, Deoxynivalenol Production, and Fungal Inhibition by Exogenous Double-Stranded RNA in the Head Blight Pathogen Fusarium graminearum. Front. Microbiol. 10:1662. doi: 10.3389/fmicb.2019.01662
Gan, D., Zhang, J., Jiang, H., Jiang, T., Zhu, S., and Cheng, B. (2010). Bacterially expressed dsRNA protects maize against SCMV infection. Plant Cell Rep, 29, 1261–1268. doi: 10.1007/s00299-010-0911-z
Goodfellow, S., Zhang, D., Wang, M. B., and Zhang, R. (2019). Bacterium-mediated RNA interference: potential application in plant protection. Plants 8, 1–18. doi: 10.3390/plants8120572
Hendrix, B., Hoffer, P., Sanders, R., Schwartz, S., Zheng, W., Eads, B., et al. (2021). Systemic gfp silencing is associated with high transgene expression in Nicotiana benthamiana. PLoS One 16:e0245422. doi: 10.1371/journal.pone.0245422
Herman, R. A., Storer, N. P., Anderson, J. A., Amijee, F., Cnudde, F., and Raybould, A. (2021). Transparency in risk-disproportionate regulation of modern crop-breeding techniques. GM Crops Food 12, 376–381. doi: 10.1080/21645698.2021.1934353
Höfle, L., Biedenkopf, D., Werner, B. T., Shrestha, A., Jelonek, L., and Koch, A. (2020). Study on the efficiency of dsRNAs with increasing length in RNA-based silencing of the Fusarium CYP51 genes. RNA Biol. 17, 463–473. doi: 10.1080/15476286.2019.1700033
Höfle, L., Shrestha, A., Werner, B., Jelonek, L., and Koch, A. (2019). Study on the efficacy of dsRNAs with increasing length in RNAi-based silencing of the Fusarium CYP51 genes. Biorxiv doi: 10.1101/824953
Hu, D., Chen, Z. Y., Zhang, C., and Ganiger, M. (2020). Reduction of Phakopsora pachyrhizi infection on soybean through host- and spray-induced gene silencing. Mol. Plant Pathol. 21, 794–807. doi: 10.1111/mpp.12931
Islam, M. T., Davis, Z., Chen, L., Englaender, J., Zomorodi, S., Frank, J., et al. (2021). Minicell-based fungal RNAi delivery for sustainable crop protection. Microbial. Biotechnol. 14, 1847–1856. doi: 10.1111/1751-7915.13699
Jay, F., Vitel, M., Brioudes, F., Louis, M., Knobloch, T., and Voinnet, O. (2019). Chemical enhancers of posttranscriptional gene silencing in Arabidopsis. Rna 25, 1078–1090. doi: 10.1261/rna.068627.118
Kaldis, A., Berbati, M., Melita, O., Reppa, C., Holeva, M., Otten, P., et al. (2018). Exogenously applied dsRNA molecules deriving from the Zucchini yellow mosaic virus (ZYMV) genome move systemically and protect cucurbits against ZYMV. Mol. Plant Pathol. 19, 883–895. doi: 10.1111/mpp.12572
Kalyandurg, P. B., Sundararajan, P., Dubey, M., Ghadamgahi, F., Zahid, M. A., Whisson, S., et al. (2021). Spray-induced gene silencing as a potential tool to control potato late blight disease. Phytopathology doi: 10.1094/PHYTO-02-21-0054-SC [Epub ahead of print].
Kleter, G. A. (2020). Food safety assessment of crops engineered with RNA interference and other methods to modulate expression of endogenous and plant pest genes. Pest Manag. Sci. 76, 3333–3339. doi: 10.1002/ps.5957
Koch, A., and Wassenegger, M. (2021). Host-induced gene silencing – mechanisms and applications. New. Phytol. 231, 54–59. doi: 10.1111/nph.17364
Koch, A., Biedenkopf, D., Furch, A., Weber, L., Rossbach, O., Abdellatef, E., et al. (2016). An RNAi-Based Control of Fusarium graminearum Infections Through Spraying of Long dsRNAs Involves a Plant Passage and Is Controlled by the Fungal Silencing Machinery. PLoS Pathog. 12:e1005901. doi: 10.1371/journal.ppat.1005901
Koch, A., Höfle, L., Werner, B. T., Imani, J., Schmidt, A., Jelonek, L., et al. (2019). SIGS vs HIGS: a study on the efficacy of two dsRNA delivery strategies to silence Fusarium FgCYP51 genes in infected host and non-host plants. Mol. Plant Pathol. 20, 1636–1644. doi: 10.1111/mpp.12866
Kolge, H., Kadam, K., Galande, S., Lanjekar, V., and Ghormade, V. (2021). New Frontiers in Pest Control: chitosan Nanoparticles-Shielded dsRNA as an Effective Topical RNAi Spray for Gram Podborer Biocontrol. ACS Appl. Bio Mater. 4, 5145–5157. doi: 10.1021/acsabm.1c00349
Konakalla, N. C., Kaldis, A., Berbati, M., Masarapu, H., and Voloudakis, A. E. (2016). Exogenous application of double-stranded RNA molecules from TMV p126 and CP genes confers resistance against TMV in tobacco. Planta 244, 961–969. doi: 10.1007/s00425-016-2567-6
Ladewig, K., Niebert, M., Xu, Z. P., Gray, P. P., and Lu, G. Q. M. (2010). Efficient siRNA delivery to mammalian cells using layered double hydroxide nanoparticles. Biomaterials 31, 1821–1829. doi: 10.1016/j.biomaterials.2009.10.058
Ladewig, K., Zhi, P. X., and Gao, Q. L. (2009). Layered double hydroxide nanoparticles in gene and drug delivery. Expert Opin. Drug Deliv. 6, 907–922. doi: 10.1517/17425240903130585
Linyu, W., Lianjun, Z., Ning, L., Xiwu, G., and Xiaoning, L. (2021). Effect of RNAi targeting CYP6CY3 on the growth, development and insecticide susceptibility of Aphis gossypii by using nanocarrier-based transdermal dsRNA delivery system. Pest. Biochem. Physiol. 177:104878. doi: 10.1016/j.pestbp.2021.104878
Liu, Q., Li, Y., Xu, K., Li, D., Hu, H., Zhou, F., et al. (2020). Clay nanosheet-mediated delivery of recombinant plasmids expressing artificial miRNAs via leaf spray to prevent infection by plant DNA viruses. Hortic. Res. 7:179. doi: 10.1038/s41438-020-00400-2
Liu, Y., and Huang, L. (2021). “Preparation and Characterization of siRNA-Loaded Liposomes” in Design and Delivery of SiRNA Therapeutics. eds H. J. Ditzel, M. Tuttolomondo, and S. Kauppinen (New York, NY: Springer). 159–169. doi: 10.1007/978-1-0716-1298-9_10
Lü, J., Liu, Z., Guo, W., Guo, M., Chen, S., Li, H., et al. (2020). Feeding delivery of dsHvSnf7 is a promising method for management of the pest Henosepilachna vigintioctopunctata (Coleoptera: Coccinellidae). Insects 11:34. doi: 10.3390/insects11010034
Lü, J., Yang, C., Liu, Z., Vélez, A. M., Guo, M., Chen, S., et al. (2021). Dietary RNAi toxicity assay suggests α and γ subunits of HvCOPI as novel molecular targets for Henosepilachna vigintioctopunctata, an emerging coccinellid pest. J. Pest Sci. 94, 1–14. doi: 10.1007/s10340-021-01350-x
Mar-Aguilar, F., Arreola-Triana, A., Mata-Cardona, D., Gonzalez-Villasana, V., Rodríguez-Padilla, C., and Reséndez-Pérez, D. (2020). Evidence of transfer of miRNAs from the diet to the blood still inconclusive. PeerJ. 8:e9567. doi: 10.7717/peerj.9567
Máximo, W. P. F., Howell, J. L., Mogilicherla, K., Basij, M., Chereddy, S. C. R. R., and Palli, S. R. (2020). Inhibitor of apoptosis is an effective target gene for RNAi-mediated control of Colorado potato beetle, Leptinotarsa decemlineata. Arch. Insect Biochem. Physiol. 104:e21685. doi: 10.1002/arch.21685
McLoughlin, A. G., Wytinck, N., Walker, P. L., Girard, I. J., Rashid, K. Y., de Kievit, T., et al. (2018). Identification and application of exogenous dsRNA confers plant protection against Sclerotinia sclerotiorum and Botrytis cinerea. Sci. Rep. 8:7320. doi: 10.1038/s41598-018-25434-4
Mehlhorn, S. G., Geibel, S., Bucher, G., and Nauen, R. (2020). Profiling of RNAi sensitivity after foliar dsRNA exposure in different European populations of Colorado potato beetle reveals a robust response with minor variability. Pest. Biochem. Physiol. 166:104569. doi: 10.1016/j.pestbp.2020.104569
Mehlhorn, S., Ulrich, J., Baden, C. U., Buer, B., Maiwald, F., Lueke, B., et al. (2021). The mustard leaf beetle, Phaedon cochleariae, as a screening model for exogenous RNAi-based control of coleopteran pests. Pest. Biochem. Physiol. 176:104870. doi: 10.1016/j.pestbp.2021.104870
Mezzetti, B., Smagghe, G., Arpaia, S., Christiaens, O., Dietz-Pfeilstetter, A., Jones, H., et al. (2020). RNAi: what is its position in agriculture? J. Pest Sci. 93, 1125–1130. doi: 10.1007/s10340-020-01238-2
Mitter, N., Worrall, E. A., Robinson, K. E., Li, P., Jain, R. G., Taochy, C., et al. (2017). Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nature Plants 3:1607. doi: 10.1038/nplants.2016.207
Mohamed, M. A., and Youssef, K. (2021). Topical delivery of host induced RNAi silencing by layered double hydroxide nanosheets: an efficient tool to decipher pathogenicity gene function of Fusarium crown and root rot in tomato. Physiol. Mol. Plant Pathol. 115:101684. doi: 10.1016/j.pmpp.2021.101684
Necira, K., Makki, M., Sanz-García, E., Canto, T., Djilani-Khouadja, F., and Tenllado, F. (2021). Topical application of Escherichia coli-encapsulated dsRNA induces resistance in Nicotiana benthamiana to potato viruses and involves rdr6 and combined activities of dcl2 and dcl4. Plants 10:644. doi: 10.3390/plants10040644
Niehl, A., Soininen, M., Poranen, M. M., and Heinlein, M. (2018). Synthetic biology approach for plant protection using dsRNA. Plant Biotechnol. J. 16, 1679–1687. doi: 10.1111/pbi.12904
Niño-Sánchez, J., Chen, L. H., de Souza, J. T., Mosquera, S., and Stergiopoulos, I. (2021). Targeted delivery of gene silencing in fungi using genetically engineered bacteria. J. Fungi 7, 1–22. doi: 10.3390/jof7020125
Niu, J., Yang, W. J., Tian, Y., Fan, J. Y., Ye, C., Shang, F., et al. (2019). Topical dsRNA delivery induces gene silencing and mortality in the pea aphid. Pest Manag. Sci. 75, 2873–2881. doi: 10.1002/ps.5457
Nowara, D., Schweizer, P., Gay, A., Lacomme, C., Shaw, J., Ridout, C., et al. (2010). HIGS: host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 22, 3130–3141. doi: 10.1105/tpc.110.077040
OECD (2020). Considerations for the Environmental Risk Assessment of the Application of Sprayed or Externally Applied ds-RNA-Based Pesticides. United Kingdom: Organisation for Economic Co-operation and Development.
Olivier. C., Teodora, D., Kaloyan, K., Salvatore, A., Mallikarjuna, R. J., Isabella, U. et al. (2018). Literature Review of Baseline Information on RNAi to Support the Environmental Risk Assessment of RNAi-Based GM Plants. Italy: European Food Safety Authority.
Palli, S. R. (2014). RNA interference in Colorado potato beetle: steps toward development of dsRNA as a commercial insecticide. Curr. Opin. Insect Sci. 6, 1–8. doi: 10.1016/j.cois.2014.09.011
Parker, K. M., Barragán Borrero, V., van Leeuwen, D. M., Lever, M. A., Mateescu, B., and Sander, M. (2019). Environmental Fate of RNA Interference Pesticides: adsorption and Degradation of Double-Stranded RNA Molecules in Agricultural Soils. Environ. Scie. Technol. 53, 3027–3036. doi: 10.1021/acs.est.8b05576
Peng, Y., Wang, K., Chen, J., Wang, J., Zhang, H., Ze, L., et al. (2020a). Identification of a double-stranded RNA-degrading nuclease influencing both ingestion and injection RNA interference efficiency in the red flour beetle Tribolium castaneum. Insect Biochem. Mol. Biol. 125:103440. doi: 10.1016/j.ibmb.2020.103440
Peng, Y., Wang, K., Zhu, G., Han, Q., Chen, J., Elzaki, M. E. A., et al. (2020b). Identification and characterization of multiple dsRNases from a lepidopteran insect, the tobacco cutworm, Spodoptera litura (Lepidoptera: Noctuidae). Pest. Biochem. Physiol. 162, 86–95. doi: 10.1016/j.pestbp.2019.09.011
Petek, M., Coll, A., Ferenc, R., Razinger, J., and Gruden, K. (2020). Validating the Potential of Double-Stranded RNA Targeting Colorado Potato Beetle Mesh Gene in Laboratory and Field Trials. Front. Plant Sci. 11:1250. doi: 10.3389/fpls.2020.01250
Pugsley, C. E., Isaac, R. E., Warren, N. J., and Cayre, O. J. (2021). Recent Advances in Engineered Nanoparticles for RNAi-Mediated Crop Protection Against Insect Pests. Front. Agron. 3:652981. doi: 10.3389/fagro.2021.652981
Qiao, L., Lan, C., Capriotti, L., Ah-Fong, A., Nino Sanchez, J., Hamby, R., et al. (2021). Spray-induced gene silencing for disease control is dependent on the efficiency of pathogen RNA uptake. Plant Biotechnol. J. 19, 1756–1768. doi: 10.1111/pbi.13589
Ram Reddy, M. K., Xu, Z. P., Lu, G. Q., and da Costa, J. C. D. (2006). Layered double hydroxides for CO2 capture: structure evolution and regeneration. Industr. Eng. Chem. Res. 45, 7504–7509. doi: 10.1021/ie060757k
Reddy, M. K. R., Xu, Z. P., Lu, G. Q., and Diniz Da Costa, J. C. (2008). Effect of SOx adsorption on layered double hydroxides for CO2 capture. Industr. Eng. Chem. Res. 47, 7357–7360. doi: 10.1021/ie8004226
Rosa, C., Kuo, Y. W., Wuriyanghan, H., and Falk, B. W. (2018). RNA interference mechanisms and applications in plant pathology. Annu. Rev. Phytopathol. 56, 581–610. doi: 10.1146/annurev-phyto-080417-050044
Santos, D., Remans, S., van den Brande, S., and Broeck, J. V. (2021). RNAs on the go: extracellular transfer in insects with promising prospects for pest management. Plants 10, 1–22. doi: 10.3390/plants10030484
Schiemann, J., Dietz-Pfeilstetter, A., Hartung, F., Kohl, C., Romeis, J., and Sprink, T. (2019). Risk Assessment and Regulation of Plants Modified by Modern Biotechniques: current Status and Future Challenges. Annu. Rev. Plant Biol. 70, 699–726. doi: 10.1146/annurev-arplant-050718-100025
Schlemmer, T., Barth, P., Weipert, L., Preußer, C., Hardt, M., Möbus, A., et al. (2021a). Isolation and Characterization of Barley (Hordeum vulgare) Extracellular Vesicles to Assess Their Role in RNA Spray-Based Crop Protection. Int. J. Mol. Sci. 22:7212. doi: 10.3390/ijms22137212
Schlemmer, T., Weipert, L., Barth, P., Werner, B., Preußer, C., Hardt, M., et al. (2021b). Host-induced gene silencing involves Arabidopsis ESCRT-III pathway for the transfer of dsRNA-derived siRNA. Biorxiv 945154. doi: 10.1101/2020.02.12.945154
Schwartz, S. H., Hendrix, B., Hoffer, P., Sanders, R. A., and Zheng, W. (2020). Carbon dots for efficient small interfering RNA delivery and gene silencing in plants. Plant Physiol. 184, 647–657. doi: 10.1104/pp.20.00733
Scott, J. G., Michel, K., Bartholomay, L. C., Siegfried, B. D., Hunter, W. B., Smagghe, G., et al. (2013). Towards the elements of successful insect RNAi. J. Insect Physiol. 59, 1212–1221. doi: 10.1016/j.jinsphys.2013.08.014
Sharma, R., Taning, C. N. T., Smagghe, G., and Christiaens, O. (2021). Silencing of double-stranded ribonuclease improves oral RNAi efficacy in southern green stinkbug Nezara viridula. Insects 12, 1–16. doi: 10.3390/insects12020115
Song, X. S., Gu, K. X., Duan, X. X., Xiao, X. M., Hou, Y. P., Duan, Y. B., et al. (2018). Secondary amplification of siRNA machinery limits the application of spray-induced gene silencing. Mol. Plant Pathol. 19, 2543–2560. doi: 10.1111/mpp.12728
Sundaresha, S., Sharma, S., Bairwa, A., Tomar, M., Kumar, R., Bhardwaj, V., et al. (2021). Spraying of dsRNA molecules derived from Phytophthora infestans, as an effective plant protection strategies for the management of potato late blight. Life Sci. 2021:2021020280. doi: 10.20944/preprints202102.0280.v1
Swaminathan, G., Shigna, A., Kumar, A., Byroju, V. V., Durgempudi, V. R., and Dinesh Kumar, L. (2021). RNA Interference and Nanotechnology: a Promising Alliance for Next Generation Cancer Therapeutics. Front. Nanotechnol. 3:694838. doi: 10.3389/fnano.2021.694838
Taning, C. N. T., Mezzetti, B., Kleter, G., Smagghe, G., and Baraldi, E. (2021). Does RNAi-Based Technology Fit within EU Sustainability Goals? Trends Biotechnol. 39, 644–647. doi: 10.1016/j.tibtech.2020.11.008
Tenllado, F., and Díaz-Ruíz, J. R. (2001). Double-Stranded RNA-Mediated Interference with Plant Virus Infection. J. Virol. 75, 12288–12297. doi: 10.1128/JVI.75.24.12288-12297.2001
Tenllado, F., Martínez-García, B., Vargas, M., and Díaz-Ruíz, J. R. (2003). Crude extracts of bacterially expressed dsRNA can be used to protect plants against virus infections. BMC Biotechnol. 3:3. doi: 10.1186/1472-6750-3-3
Thairu, M. W., Skidmore, I. H., Bansal, R., Nováková, E., Hansen, T. E., Li-Byarlay, H., et al. (2017). Efficacy of RNA interference knockdown using aerosolized short interfering RNAs bound to nanoparticles in three diverse aphid species. Insect Mol. Biol. 26, 356–368. doi: 10.1111/imb.12301
Timmons, L., Court, D. L., and Fire, A. (2001). Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans. Gene 263, 103–112. doi: 10.1016/S0378-1119(00)00579-5
United Nations Department of Economics and Social Affairs (2019). World Population Prospects 2019. Available online at: http://www.ncbi.nlm.nih.gov/pubmed/12283219 (accessed September 2021).
Uslu, V. V., and Wassenegger, M. (2020). Critical view on RNA silencing-mediated virus resistance using exogenously applied RNA. Curr. Opin. Virol. 42, 18–24. doi: 10.1016/j.coviro.2020.03.004
Uslu, V. V., Bassler, A., Krczal, G., and Wassenegger, M. (2020). High-Pressure-Sprayed Double Stranded RNA Does Not Induce RNA Interference of a Reporter Gene. Front. Plant Sci. 11:534391. doi: 10.3389/fpls.2020.534391
Voloudakis, A. E., Holeva, M. C., Sarin, L. P., Bamford, D. H., Vargas, M., Poranen, M. M., et al. (2015). Efficient double-stranded RNA production methods for utilization in plant virus control. Methods Mol. Biol. 1236, 255–274. doi: 10.1007/978-1-4939-1743-3_19
Wagner, D. L., Grames, E. M., Forister, M. L., Berenbaum, M. R., and Stopak, D. (2021). Insect decline in the Anthropocene: death by a thousand cuts. Proc. Natl. Acad. Sci. U. S. A. 118:e2023989118. doi: 10.1073/pnas.2023989118
Wang, K., Li, B., Yu, Y., Wei, J., Zhu, J., Wang, J., et al. (2021). Bacterial Rdl2 dsRNA increased the insecticidal activity of GABAR blockers and allosteric modulators against Plutella xylostella. Biorxiv doi: 10.1101/2021.03.24.436736
Wang, M., Weiberg, A., Lin, F. M., Thomma, B. P. H. J., Huang, H. D., and Jin, H. (2016). Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2:16151. doi: 10.1038/nplants.2016.151
Werner, B. T., Gaffar, F. Y., Schuemann, J., Biedenkopf, D., and Koch, A. M. (2020). RNA-Spray-Mediated Silencing of Fusarium graminearum AGO and DCL Genes Improve Barley Disease Resistance. Front. Plant Sci. 11:476. doi: 10.3389/fpls.2020.00476
Whitten, M. M. A., Facey, P. D., del Sol, R., Fernández-Martínez, L. T., Evans, M. C., Mitchell, J. J., et al. (2016). Symbiont-mediated RNA interference in insects. Proc. R. Soc. B Biol. Sci. 283:20160042. doi: 10.1098/rspb.2016.0042
Whitten, M., and Dyson, P. (2017). Gene silencing in non-model insects: overcoming hurdles using symbiotic bacteria for trauma-free sustainable delivery of RNA interference: sustained RNA interference in insects mediated by symbiotic bacteria: applications as a genetic tool and as a biocid. Bioessays 39:1600247. doi: 10.1002/bies.201600247
Willow, J., Soonvald, L., Sulg, S., Kaasik, R., Silva, A. I., Taning, C. N. T., et al. (2021). RNAi efficacy is enhanced by chronic dsRNA feeding in pollen beetle. Commun. Biol. 4, 1–8. doi: 10.1038/s42003-021-01975-9
Worrall, E. A., Bravo-Cazar, A., Nilon, A. T., Fletcher, S. J., Robinson, K. E., Carr, J. P., et al. (2019). Exogenous application of RNAi-inducing double-stranded RNA inhibits aphid-mediated transmission of a plant virus. Front. Plant Sci. 10:265. doi: 10.3389/fpls.2019.00265
Wu, J. J., Mu, L. L., Kang, W. N., Ze, L. J., Shen, C. H., Jin, L., et al. (2021). RNA interference targeting ecdysone receptor blocks the larval–pupal transition in Henosepilachna vigintioctopunctata. Insect Sci. 28, 419–429. doi: 10.1111/1744-7917.12777
Wytinck, N., Manchur, C. L., Li, V. H., Whyard, S., and Belmonte, M. F. (2020a). DsRNA uptake in plant pests and pathogens: insights into RNAi-based insect and fungal control technology. Plants 9, 1–17. doi: 10.3390/plants9121780
Wytinck, N., Sullivan, D. S., Biggar, K. T., Crisostomo, L., Pelka, P., Belmonte, M. F., et al. (2020b). Clathrin mediated endocytosis is involved in the uptake of exogenous double-stranded RNA in the white mold phytopathogen Sclerotinia sclerotiorum. Sci. Rep. 10, 12773. doi: 10.1038/s41598-020-69771-9
Xu, Z. P., Stevenson, G., Lu, C. Q., and Lu, G. Q. (2006). Dispersion and size control of layered double hydroxide nanoparticles in aqueous solutions. J. Phys. Chem. B 110, 16923–16929. doi: 10.1021/jp062281o
Yan, S., Ren, B. Y., and Shen, J. (2021). Nanoparticle-mediated double-stranded RNA delivery system: a promising approach for sustainable pest management. Insect Sci. 28, 21–34. doi: 10.1111/1744-7917.12822
Yin, G., Sun, Z., Liu, N., Zhang, L., Song, Y., Zhu, C., et al. (2009). Production of double-stranded RNA for interference with TMV infection utilizing a bacterial prokaryotic expression system. Appl. Microbiol. Biotechnol. 84, 323–333. doi: 10.1007/s00253-009-1967-y
Yu, X. D., Liu, Z. C., Huang, S. L., Chen, Z. Q., Sun, Y. W., Duan, P. F., et al. (2016). RNAi-mediated plant protection against aphids. Pest. Manag. Sci. 72, 1090–1098. doi: 10.1002/ps.4258
Zabel, M. D., Mollnow, L., and Bender, H. (2021). “siRNA Therapeutics for Protein Misfolding Diseases of the Central Nervous System” in Design and Delivery of SiRNA Therapeutics, eds H. J. Ditzel, M. Tuttolomondo, and S. Kauppinen (New York, NY: Springer). 377–394. doi: 10.1007/978-1-0716-1298-9_20
Zhang, H., Cao, Y., Xu, D., Goh, N. S., Demirer, G. S., Cestellos-Blanco, S., et al. (2021). Gold-Nanocluster-Mediated Delivery of siRNA to Intact Plant Cells for Efficient Gene Knockdown. Am. Chem. Soc. 13, 5859–5866. doi: 10.1101/2021.03.17.435890
Zhang, H., Demirer, G. S., Zhang, H., Ye, T., Goh, N. S., Aditham, A. J., et al. (2019). DNA nanostructures coordinate gene silencing in mature plants. Proc. Natl. Acad. Sci. U. S. A. 116, 7543–7548. doi: 10.1073/pnas.1818290116
Zhang, H., Zhang, H., Demirer, G. S., González-Grandío, E., Fan, C., and Landry, M. P. (2020). Engineering DNA nanostructures for siRNA delivery in plants. Nat. Protocols 15, 3064–3087. doi: 10.1038/s41596-020-0370-0
Zhang, J., Khan, S. A., Hasse, C., Ruf, S., Heckel, D. G., and Bock, R. (2015). Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science 347, 991L–994. doi: 10.1126/science.1261680
Keywords: RNAi-based plant protection, RNA biopesticides, dsRNA, siRNA, nanomaterial-based siRNA delivery, spray-induced gene silencing (SIGS)
Citation: Rank AP and Koch A (2021) Lab-to-Field Transition of RNA Spray Applications – How Far Are We? Front. Plant Sci. 12:755203. doi: 10.3389/fpls.2021.755203
Received: 08 August 2021; Accepted: 15 September 2021;
Published: 15 October 2021.
Edited by:
Antje Dietz-Pfeilstetter, Julius Kühn-Institut - Braunschweig, GermanyReviewed by:
Athanasios Dalakouras, Institute of Industrial and Forage Crops, Hellenic Agricultural Organisation, GreeceClauvis Nji Tizi Taning, Ghent University, Belgium
Copyright © 2021 Rank and Koch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aline Koch, aline.koch@uni-hohenheim.de