Optimizing Nitrogen Fertilization to Improve Qualitative Performances and Physiological and Yield Responses of Potato (Solanum tuberosum L.)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experimental Design, Plant Material and Management Practices
2.2. Crop Physiology
2.3. Crop Yield and Its Components
2.4. Nitrogen Crop Uptake and Nitrogen Efficiency Indices
2.5. Tuber Chemical Composition
2.6. Statistical Analysis
2.7. Weather Conditions
3. Results and Discussion
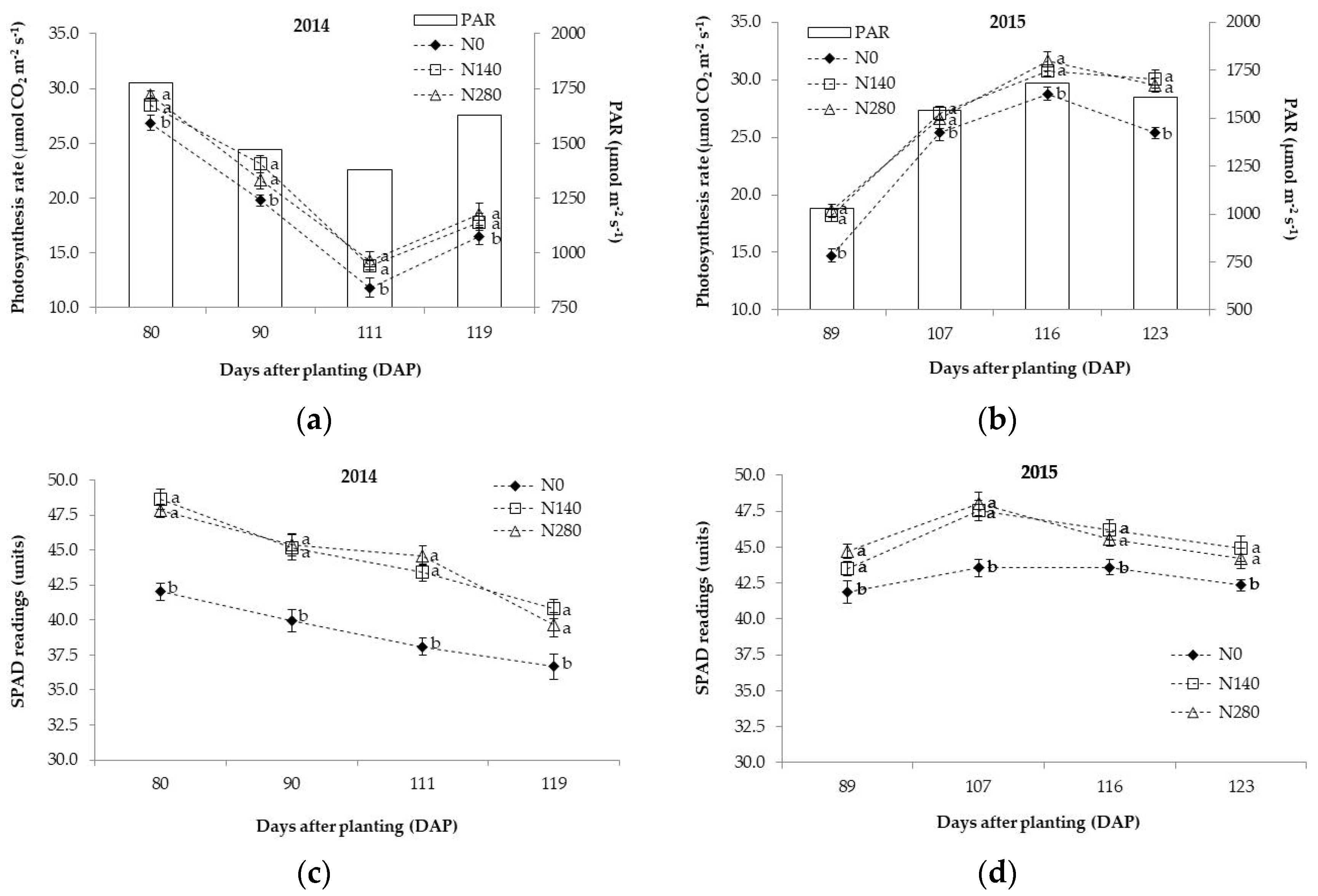
3.1. Physiological Traits
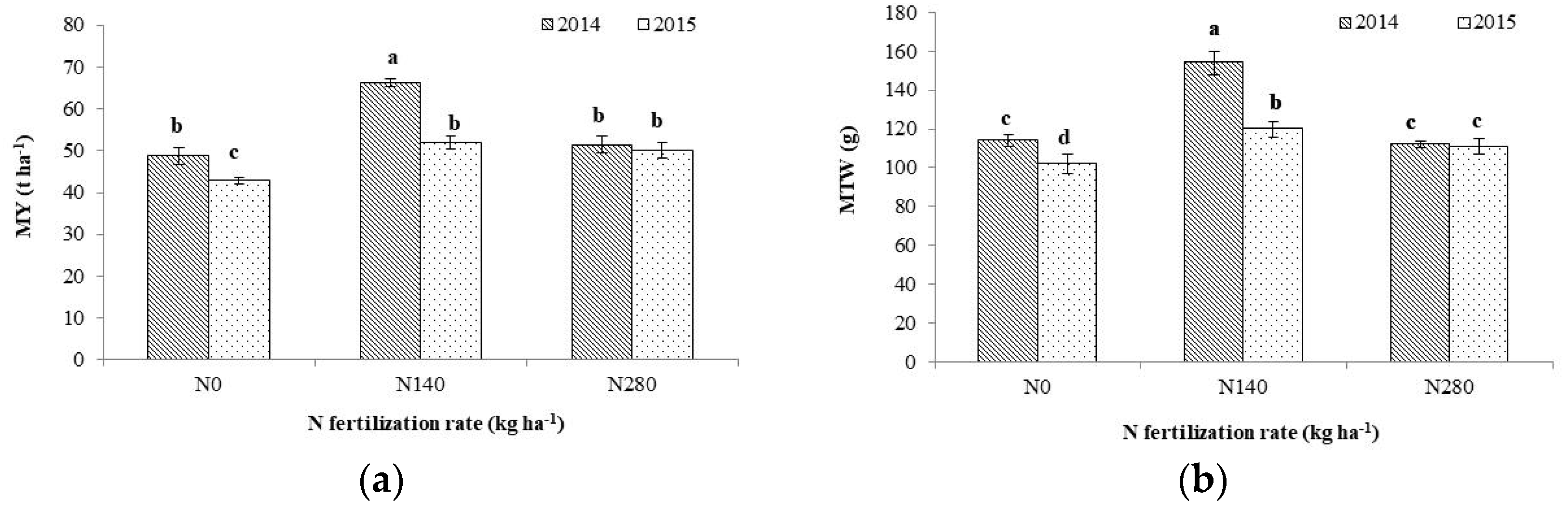
3.2. Aboveground Dry Biomass and Yield Components
3.3. Crop Nitrogen Uptake and N Efficiency Indices
3.4. Tuber Chemical Traits
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Savci, S. An agricultural pollutant: Chemical fertilizer. Int. J. Environ. Sci. Dev. 2012, 3, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Stefanelli, D.; Goodwin, I.; Jones, R. Minimal nitrogen and water use in horticulture: Effects on quality and content of selected nutrients. Food Res. Int. 2010, 43, 1833–1843. [Google Scholar] [CrossRef]
- Mengel, K.; Kirkby, E.A.; Kosegarten, H.; Appel, T. Principles of Plant Nutrition; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Ryan, J.; Masri, S.; Ceccarelli, S.; Grando, S.; Ibrikci, H. Differential responses of barley landraces and improved barley cultivars to nitrogen-phosphorus fertilizer. J. Plant Nutr. 2008, 31, 381–393. [Google Scholar] [CrossRef]
- Karyotis, T.; Güçdemir, İ.; Akgül, S.; Panagopoulos, A.; Karyoti, K.; Demir, S.; Kasacı, A. Nitrogen fertilization plans for the main crops of Turkey to mitigate nitrates pollution. Eurasian J. Soil Sci. 2014, 3, 13–24. [Google Scholar] [CrossRef] [Green Version]
- FAO Statistical Database. Available online: http://www.faostat.org/ (accessed on 5 July 2019).
- Frusciante, L.; Barone, A.; Carputo, D.; Ranalli, P. Breeding and physiological aspects of potato cultivation in the Mediterranean region. Potato Res. 1999, 42, 265–277. [Google Scholar] [CrossRef]
- Ierna, A.; Pandino, G.; Lombardo, S.; Mauromicale, G. Tuber yield, water and fertilizer productivity in early potato as affected by a combination of irrigation and fertilization. Agric. Water Manag. 2011, 101, 35–41. [Google Scholar] [CrossRef]
- Cantore, V.; Wassar, F.; Yamac, S.S.; Sellami, M.H.; Albrizio, R.; Stellacci, A.M.; Todorovic, M. Yield and water use efficiency of early potato grown under different irrigation regimes. Int. J. Plant Prod. 2014, 8, 409–428. [Google Scholar]
- Meise, P.; Seddig, S.; Uptmoor, R.; Ordon, F.; Schum, A. Impact of nitrogen supply on leaf water relations and physiological traits in a set of potato (Solanum tuberosum L.) cultivars under drought stress. J. Agron. Crop. Sci. 2018, 204, 59–374. [Google Scholar] [CrossRef]
- Lombardo, S.; Lo Monaco, A.; Pandino, G.; Parisi, B.; Mauromicale, G. The phenology: Yield and tuber composition of ‘early’ crop potatoes: A comparison between organic and conventional cultivation systems. Renew. Agric. Food Syst. 2013, 28, 50–58. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Mauromicale, G. The influence of growing environment on the antioxidant and mineral content of “early” crop potato. J. Food Compos. Anal. 2013, 32, 28–35. [Google Scholar] [CrossRef]
- Ierna, A.; Mauromicale, G. Potato growth, yield and water productivity response to different irrigation and fertilization regimes. Agric. Water Manag. 2018, 201, 21–26. [Google Scholar] [CrossRef]
- Izmirlioglu, G.; Demirci, A. Enhanced bio-ethanol production from industrial potato waste by statistical medium optimization. Int. J. Mol. Sci. 2015, 16, 24490–24505. [Google Scholar] [CrossRef] [Green Version]
- Jagatee, S.; Behera, S.; Dash, P.K.; Sahoo, S.; Mohanty, R.C. Bioprospecting starchy feedstocks for bioethanol production: A future perspective. JMRR 2015, 3, 24–42. [Google Scholar]
- Koch, M.; Naumann, M.; Pawelzik, E.; Gransee, A.; Thiel, H. The importance of nutrient management for potato production Part I: Plant nutrition and yield. Potato Res. 2019. [Google Scholar] [CrossRef] [Green Version]
- Mauromicale, G.; Signorelli, P.; Ierna, A.; Foti, S. Effects of intraspecific competition on yield of early potato grown in Mediterranean environment. Am. J. Potato Res. 2003, 80, 281–288. [Google Scholar] [CrossRef]
- Ierna, A.; Mauromicale, G. Sustainable and profitable nitrogen fertilization management of potato. Agronomy 2019, 10, 582. [Google Scholar] [CrossRef] [Green Version]
- Milroy, S.P.; Wang, P.; Sodras, V.O. Defining upper limits of nitrogen uptake and nitrogen use efficiency of potato in response to crop N supply. Field Crop. Res. 2019, 239, 38–46. [Google Scholar] [CrossRef]
- Ospina, C.A.; Lammerts van Bueren, E.T.; Allefs, J.J.H.M.; Engel, B.; van der Putten, P.E.L.; van der Linden, C.G.; Struik, P.C. Diversity of crop development traits and nitrogen use efficiency among potato cultivars grown under contrasting nitrogen regimes. Euphytica 2014, 199, 13–29. [Google Scholar] [CrossRef]
- Vos, J.; van der Putten, P.E.L. Effect of nitrogen supply on leaf growth, leaf nitrogen economy and photosynthetic capacity in potato. Field Crop. Res. 1998, 59, 63–72. [Google Scholar] [CrossRef]
- Mauromicale, G.; Ierna, A.; Marchese, M. Chlorophyll fluorescence and chlorophyll content in field-grown potato as affected by nitrogen supply, genotype, and plant age. Photosynthetica 2006, 44, 76–82. [Google Scholar] [CrossRef]
- Ahmed, A.; Zaki, M.; Shafeek, M.; Helmy, Y.; El-Baky, M.A. Integrated use of farmyard manure and inorganic nitrogen fertilizer on growth, yield and quality of potato (Solanum tuberosum L.). Int. J. Curr. Microbiol. App. Sci. 2015, 4, 325–349. [Google Scholar]
- Fontes, P.C.R.; Braun, H.; Busato, C.; Cecon, P.R. Economic optimum nitrogen fertilization rates and nitrogen fertilization rate effects on tuber characteristics of potato cultivars. Potato Res. 2010, 53, 167–179. [Google Scholar] [CrossRef]
- Foti, S.; Mauromicale, G.; Ierna, A. Influence of irrigation regimes on growth and yield of potato cv. Spunta. Potato Res. 1995, 38, 307–318. [Google Scholar] [CrossRef]
- Getahun, B.B. Potato breeding for nitrogen-use efficiency: Constraints, achievements, and future prospects. J. Crop. Sci. Biotechnol. 2018, 21, 269–281. [Google Scholar] [CrossRef]
- Tiwari, J.K.; Plett, D.; Garnett, T.; Chakrabarti, S.K.; Singh, R.K. Integrated genomics, physiology and breeding approaches for improving nitrogen use efficiency in potato: Translating knowledge from other crops. Funct. Plant Biol. 2018, 45, 587–605. [Google Scholar] [CrossRef] [Green Version]
- Bélanger, G.; Walsh, J.R.; Richards, J.E.; Milburn, P.H.; Ziadi, N. Yield response of two potato cultivars to supplemental irrigation and N fertilization in New Brunswick. Am. J. Potato Res. 2000, 77, 11–21. [Google Scholar] [CrossRef]
- Soil Survey Staff. Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; U.S. Gov. Print. Office: Washington, DC, USA, 1999.
- Fierotti, G.; Dazzi, C.; Raimondi, S. Carta dei suoli della Sicilia 1:250.000; Regione Siciliana and Università degli Studi di Palermo: Palermo, Italy, 1988. [Google Scholar]
- Foti, S. Early potatoes in Italy with particular reference to Sicily. Potato Res. 1999, 42, 229–240. [Google Scholar] [CrossRef]
- Italian Society of Soil Science. Metodi Normalizzati di Analisi del Suolo; Edagricole: Bologna, Italy, 1985. [Google Scholar]
- UNICHIM. Analisi dei Terreni Agrari; Unichim: Milano, Italy, 1985. [Google Scholar]
- Mauromicale, G.; Ierna, A. Patata primaticcia. In Fisionomia e profili di qualità dell’orticoltura meridional; Bianco, V.V., La Malfa, G., Tudisca, G., Eds.; Arti Grafiche Siciliane: Palermo, Italy, 1999; pp. 275–296. [Google Scholar]
- Uddling, J.; Gelang-Alfredsson, J.; Piikki, K.; Pleijel, H. Evaluating the relationship between leaf chlorophyll concentration and SPAD-502 chlorophyll meter readings. Photosynth. Res. 2007, 91, 37–46. [Google Scholar] [CrossRef]
- Meise, P.; Seddig, S.; Uptmoor, R.; Ordon, F.; Schum, A. Assessment of yield and yield components of starch potato cultivars (Solanum tuberosum L.) under nitrogen deficiency and drought stress conditions. Potato Res. 2019, 62, 193–220. [Google Scholar] [CrossRef]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and interpretation of factors which contribute to efficiency of nitrogen utilization. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]
- Gariglio, N.F.; Pilatti, R.A.; Baldi, B.L. Using nitrogen balance to calculate fertilization in strawberries. HortTechnology 2000, 10, 147–150. [Google Scholar] [CrossRef] [Green Version]
- Ceccon, P.; Fagnano, M.; Grignani, C.; Monti, M.; Orlandini, S. Agronomia; Edises: Napoli, Italy, 2017. [Google Scholar]
- Snyder, J.C.; Desborough, S.L. Rapid estimation of potato tuber total protein content with coomassie brilliant blue g-250. Appl. Genet. 1978, 52, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, S.; Pandino, G.; Mauromicale, G. The effect on tuber quality of an organic versus a conventional cultivation system in the early crop potato. J. Food Comp. Anal. 2017, 62, 189–196. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Mauromicale, G. The nutraceutical response of two globe artichoke cultivars to contrasting NPK fertilizer regimes. Food Res. Int. 2015, 76, 852–859. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 22, 25–30. [Google Scholar] [CrossRef]
- Wilhelm, W.W.; Arnold, S.L.; Schepers, J.S. Using a nitrate specific ion electrode to determine stalk nitrate-nitrogen concentration. Agron. J. 2000, 92, 186–189. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. Analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Levene, H. Robust tests for equality of variances. In Contributions to Probability and Statistics; Olkin, I., Ghurye, S.G., Hoeffding, W., Madow, W.G., Mann, H.B., Eds.; Stanford University Press: Stanford, CA, USA, 1960; pp. 278–292. [Google Scholar]
- Busato, C.; Fontes, P.C.R.; Braun, H.; Cecon, P.R. Seasonal variation and threshold values for chlorophyll meter readings on leaves of potato cultivars. J. Plant Nutr. 2010, 33, 2148–2156. [Google Scholar] [CrossRef]
- Gianquinto, G.; Goffart, J.P.; Olivier, M.; Guarda, G.; Colauzzi, M.; Dalla Costa, L.; Delle Vedove, G.; Vos, J.; MacKerron, D.K.L. The use of hand-held chlorophyll meters as a tool to assess the nitrogen status and to guide nitrogen fertilization of potato crop. Potato Res. 2004, 47, 35–80. [Google Scholar] [CrossRef]
- Darwish, T.M.; Atallah, T.W.; Hajhasan, S.; Haidar, A. Nitrogen and water use efficiency of fertigated processing potato. Agric. Water Manag. 2006, 85, 95–104. [Google Scholar] [CrossRef]
- Badr, M.A.; El-Tohamy, W.A.; Zaghloul, A.M. Yield and water use efficiency of potato grown under different irrigation and nitrogen levels in an arid region. Agric. Water Manag. 2012, 110, 9–15. [Google Scholar] [CrossRef]
- Zebarth, B.J.; Rosen, C.J. Research perspective on nitrogen bmp development for potato. Am. J. Potato Res. 2007, 84, 3–18. [Google Scholar] [CrossRef]
- De la Morena, I.; Guillén, A.; del Moral, L.F.G. Yield development in potatoes as influenced by cultivar and the timing and level of nitrogen fertilization. Am. Potato J. 1994, 71, 165–173. [Google Scholar] [CrossRef]
- Dalla Costa, L.; Delle Vedove, G.; Gianquinto, G.; Giovanardi, R.; Peressotti, A. Yield, water use efficiency and nitrogen uptake in potato: Influence of drought stress. Potato Res. 1997, 40, 19–34. [Google Scholar] [CrossRef]
- Neumann, A.; Torstensson, G.; Aronsson, H. Nitrogen and phosphorus leaching losses from potatoes with different harvest times and following crops. Field Crop. Res. 2012, 133, 130–138. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [Green Version]
- Iwama, K. Physiology of the potato: New insights into root system and repercussions for crop management. Potato Res. 2008, 51, 333–353. [Google Scholar] [CrossRef]
- Tyler, K.B.; Broadbent, F.E.; Bishop, J.C. Efficiency of nitrogen uptake by potatoes. Am. Potato J. 1983, 60, 261–269. [Google Scholar] [CrossRef]
- Zvomuya, F.; Rosen, C.J.; Creighton Miller, J. Response of Russet Norkotah clonal selections to nitrogen fertilization. Am. J. Potato Res. 2002, 79, 231–239. [Google Scholar] [CrossRef]
- Solaiman, A.H.M.; Nishizawa, T.; Roy, T.S.; Rahman, M.; Chakraborty, R.; Choudhury, J.; Choudhury, J.; Sarkar, M.D.; Hasanuzzama, M. Yield, dry matter: Specific gravity and color of three Bangladesh local potato cultivars as influenced by stage of maturity. J. Plant Sci. 2015, 10, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Dean, B.B.; Thomton, R.E. The Specific Gravity of Potatoes; Washington State Univ. Cooperative Extension Bulletin #1541: Pullman, WA, USA, 1992. [Google Scholar]
- Cacace, J.E.; Huarte, M.A.; Monti, M.C. Evaluation of potato cooking quality in Argentina. Am. Potato J. 1994, 71, 145–153. [Google Scholar] [CrossRef]
- Pereira, A.B.; Villa Nova, N.A.; Ramos, V.J.; Pereira, A.R. Potato potential yield based on climatic elements and cultivar characteristics. Bragantia 2008, 67, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Locascio, S.J.; Wiltbank, W.J.; Gull, D.D.; Maynard, D.N. Fruit and vegetable quality as affected by nitrogen nutrition. In Nitrogen in Crop Production; Hauck, R.D., Ed.; American Society of Agronomy: Madison, WI, USA, 1984; pp. 617–641. [Google Scholar]
- Bryant, J.P.; Chapin, F.S.; Klein, D.R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 1983, 40, 357–368. [Google Scholar] [CrossRef] [Green Version]
- van Eck, H.A. Genetics of morphological and tuber traits. In Potato Biology and Biotechnology: Advances and Perspectives; Vreugdenhil, D., Bradshaw, J., Gebhardt, C., Govers, F., MacKerron, D.K.L., Taylor, M., Ross, H., Eds.; Elsevier: Oxford, UK, 2007; pp. 91–111. [Google Scholar]
- Bártová, V.; Bárta, J.; Diviš, J.; Švajner, J.; Peterka, J. Crude protein content in tubers of starch processing potato cultivars in dependence on different agro-ecological conditions. J. Cent. Eur. Agric. 2009, 10, 57–66. [Google Scholar]
- Wang, Z.; Li, S.; Malhi, S. Effects of fertilization and other agronomic measures on nutritional quality of crops. J. Sci. Food Agric. 2008, 88, 7–23. [Google Scholar] [CrossRef]
- Sadej, W.; Namiotko, A. Nitrates content in potato tubers cultivated under various fertilization systems. Ecol. Chem. Eng. A 2011, 18, 1123–1130. [Google Scholar]
- Santamaria, P. Nitrate in vegetable: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Licciardello, F.; Lombardo, S.; Rizzo, V.; Pitino, I.; Pandino, G.; Strano, M.G.; Muratore, G.; Restuccia, C.; Mauromicale, G. Integrated agronomical and technological approach for the quality maintenance of ready-to-fry potato sticks during refrigerated storage. Postharvest Biol. Technol. 2018, 136, 23–30. [Google Scholar] [CrossRef]
- Rizzo, V.; Amoroso, L.; Licciardello, F.; Mazzaglia, A.; Muratore, G.; Restuccia, C.; Lombardo, S.; Pandino, G.; Strano, M.G.; Mauromicale, G. The effect of sous vide packaging with rosemary essential oil on storage quality of fresh-cut potato. LWT Food Sci. Technol. 2018, 94, 111–118. [Google Scholar] [CrossRef]
- Brown, C.R. Antioxidants in potato. Am. J. Potato Res. 2005, 82, 163–172. [Google Scholar] [CrossRef]
- Lee, S.K.; Kader, A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Lachman, J.; Hamouz, K.; Čepl, J.; Pivec, V.; Šulc, M.; Dvořák, P. The effect of selected factors on polyphenol content and antioxidant activity in potato tubers. Chem. Listy 2006, 100, 522–527. [Google Scholar]
- Hamouz, K.; Čepl, J.; Dvořák, P. Influence of environmental conditions on the quality of potato tubers. Hortic. Sci. 2005, 32, 89–95. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Month | 2014 | 2015 | Long-Term Period | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Rainfall (mm) | Min. Air Temp. (°C) | Max. Air Temp. (°C) | Rainfall (mm) | Min. Air Temp. (°C) | Max. Air Temp. (°C) | Rainfall (mm) | Min. Air Temp. (°C) | Max. Air Temp. (°C) | |
| January | 21 | 11.1 | 16.5 | 46 | 8.8 | 15.2 | 65 | 7.1 | 15.4 |
| February | 23 | 11.0 | 16.9 | 73 | 8.5 | 13.7 | 38 | 7.6 | 16.2 |
| March | 15 | 10.7 | 16.6 | 31 | 9.5 | 16.1 | 25 | 8.8 | 17.7 |
| April | 8 | 13.2 | 19.7 | 0 | 13.4 | 18.7 | 31 | 10.9 | 20.2 |
| May | 1 | 16.4 | 22.0 | 10 | 15.3 | 22.8 | 20 | 14.4 | 24.3 |
| Total/mean | 68 | 12.5 | 18.3 | 160 | 11.4 | 17.3 | 179 | 9.8 | 18.8 |
| Source of Variation | ||||
|---|---|---|---|---|
| Variable | Year | Nitrogen Fertilization Rate (N) | Measurement Time (M) | (N) × (M) |
| Photosynthesis Rate | 2014 | *** | *** | * |
| *** | *** | * | ||
| SPAD Readings | 2015 | *** | *** | *** |
| *** | *** | ** | ||
| Source of Variation | |||
|---|---|---|---|
| Variable a | Nitrogen fertilization Rate (N) | Year (Y) | (N) × (Y) |
| ADB | * | ** | NS |
| MY | *** | ** | * |
| MTW | *** | *** | *** |
| NTP | ** | NS | NS |
| CNU | *** | *** | ** |
| NUE | *** | ** | * |
| NUpE | *** | *** | *** |
| NUtE | NS | *** | ** |
| FUE | *** | ** | *** |
| DM | ** | NS | * |
| Starch | *** | *** | * |
| Sucrose | *** | ** | NS |
| Glucose | ** | NS | NS |
| Fructose | ** | NS | NS |
| Total protein | *** | NS | NS |
| Total polyphenols | *** | *** | * |
| Nitrate | *** | *** | * |
| Ascorbic acid | *** | ** | *** |
| Antioxidant activity | *** | *** | ** |
| Year | N Fertilization Rate | ||||
|---|---|---|---|---|---|
| Variable | 2014 | 2015 | N0 | N140 | N280 |
| ADB (t ha−1 DM) | 1.02 ± 0.02 a | 0.85 ± 0.04 b | 0.78 ± 0.02 c | 0.99 ± 0.03 b | 1.10 ± 0.03 a |
| MY (t ha−1) a | 55.5 ± 1.2 a | 48.3 ± 2.0 b | 45.8 ± 1.0 c | 59.1 ± 1.3 a | 50.8 ± 2.0 b |
| MTW (g) | 127 ± 4 a | 111 ± 6 b | 108 ± 4 b | 137 ± 4 a | 112 ± 5 b |
| NTP (no. plant−1) | 9.7 ± 0.3 a | 9.6 ± 0.2 a | 9.4 ± 0.3 b | 9.6 ± 0.2 b | 10.1 ± 0.4 a |
| CNU (kg ha−1) | 145 ± 5 a | 128 ± 3 b | 113 ± 4 b | 147 ± 4 a | 148 ± 5 a |
| NUE (kg tuber DW kg N−1) | 66.0 ± 1.0 a | 56.6 ± 1.3 b | 113.3 ± 0.9 a | 46.8 ± 0.4 b | 23.8 ± 1.0 c |
| NUpE (kg N kg N−1) | 0.98 ± 0.08 a | 0.85 ± 0.036 b | 1.61 ± 0.09 a | 0.70 ± 0.08 b | 0.42 ± 0.04 c |
| NUtE (kg tuber DW kg N−1) | 64.4 ± 0.7 a | 64.3 ± 0.6 a | 70.3 ± 0.8 a | 66.6 ± 0.6 a | 56.2 ± 0.7 b |
| FUE (kg kg−1) | 67.7 ± 1.3 a | 45.0 ± 1.0 b | - | 95.0 ± 1.6 a | 17.7 ± 1.0 b |
| Dry matter (DM) (%) | 16.8 ± 1.0 a | 16.8 ± 0.9 a | 17.2 ± 1.0 a | 16.6 ± 0.5 b | 16.4 ± 0.6 b |
| Starch (g kg−1 DM) | 632 ± 8 a | 604 ± 7 b | 640 ± 8 a | 634 ± 6 a | 580 ± 6 b |
| Sucrose (g kg−1 DM) | 11.7 ± 0.5 a | 11.9 ± 0.6 a | 13.1 ± 0.6 a | 11.4 ± 0.4 b | 11.0 ± 0.3 b |
| Glucose (g kg−1 DM) | 6.1 ± 0.1 a | 5.9 ± 0.3 a | 6.7 ± 0.4 a | 5.8 ± 0.2 b | 5.6 ± 0.3 b |
| Fructose (g kg−1 DM) | 2.0 ± 0.2 a | 2.1 ± 0.3 a | 2.4 ± 0.4 a | 1.8 ± 0.1 b | 1.9 ± 0.1 b |
| Total protein(g kg−1 DM) | 89 ± 4 a | 90 ± 3 a | 82 ± 4 b | 86 ± 4 b | 100 ± 5 a |
| Total polyphenols (g kg−1 DM) | 3.17 ± 0.07 b | 3.83 ± 0.10 a | 3.86 ± 0.09 a | 3.31 ± 0.08 b | 3.33 ± 0.07 b |
| Nitrate (g kg−1 DM) | 1.03 ± 0.7 a | 0.88 ± 0.09 b | 0.86 ± 0.09 c | 0.93 ± 0.06 b | 1.08 ± 0.04 a |
| Ascorbic acid (g kg−1 DM) | 0.60 ± 0.05 a | 0.63 ± 0.05 a | 0.71 ± 0.04 a | 0.61 ± 0.07 b | 0.52 ± 0.08 c |
| Antioxidant activity (%inhibition DPPH) | 55.6 ± 1.2 b | 61.1 ± 1.2 a | 62.3 ± 1.2 a | 58.3 ± 1.0 b | 54.4 ± 1.3 c |
| Year | N Fertilization Rate | CNU (kg ha−1) | NUE (kg Tuber DM kg N−1) | NUpE (kg N kg N−1) | NUtE (kg Tuber DM kg N−1) | FUE (kg kg−1) |
|---|---|---|---|---|---|---|
| 2014 | N0 | 120.5 ± 2.5 d | 122.1 ± 1.3 a | 1.72 ± 0.06 a | 70.9 ± 2.3 a | - |
| N140 | 162.6 ± 3.0 a | 52.0 ± 1.8 c | 0.77 ± 0.08 c | 67.1 ± 2.0 b | 125.7 ± 2.0 a | |
| N280 | 150.9 ± 4.0 b | 23.8 ± 1.0 e | 0.43 ± 0.05 e | 55.2 ± 1.9 d | 9.6 ± 0.3 d | |
| 2015 | N0 | 104.8 ± 2.9 e | 104.5 ± 1.8 b | 1.5084 ± 0.05 b | 69.7 ± 1.6 a | - |
| N140 | 132.2 ± 3.0 c | 41.6 ± 1.5 d | 0.63 ± 0.05 d | 66.0 ± 2.5 b | 64.3 ± 1.2 b | |
| N280 | 145.7 ± 2.2 bc | 23.8 ± 1.2 e | 0.42 ± 0.05 e | 57.1 ± 2.5 c | 25.7 ± 1.8 c |
| Year | N Fertilization Rate | Dry Matter (DM) (%) | Starch (g kg−1 DM) | Nitrate (g kg−1 DM) | Ascorbic Acid (g kg−1 DM) | Total Polyphenols (g kg−1 DM) | Antioxidant Activity (%inhibition DPPH) |
|---|---|---|---|---|---|---|---|
| 2014 | N0 | 17.5 ± 1.0 a | 647 ± 4 ab | 0.94 ± 0.06 c | 0.67 ± 0.02 b | 3.42 ± 0.11 b | 58.2 ± 1.2 bc |
| N140 | 16.5 ± 1.5 cd | 657 ± 8 a | 1.01 ± 0.03 b | 0.58 ± 0.04 d | 3.04 ± 0.05 c | 55.9 ± 0.4 c | |
| N280 | 16.3 ± 0.9 d | 592 ± 9 cd | 1.15 ± 0.09 a | 0.54 ± 0.06 e | 3.05 ± 0.08 c | 52.5 ± 0.8 d | |
| 2015 | N0 | 17.0 ± 0.5 b | 632 ± 5 b | 0.78 ± 0.06 e | 0.75 ± 0.04 a | 4.31 ± 0.20 a | 66.4 ± 0.7 a |
| N140 | 16.8 ± 0.7 bc | 612 ± 11 c | 0.85 ± 0.04 d | 0.64 ± 0.03 c | 3.58 ± 0.13 b | 60.7 ± 0.6 b | |
| N280 | 16.6 ± 0.1 c | 569 ± 10 d | 1.02 ± 0.04 b | 0.49 ± 0.05 f | 3.61 ± 0.07 b | 56.3 ± 0.4 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lombardo, S.; Pandino, G.; Mauromicale, G. Optimizing Nitrogen Fertilization to Improve Qualitative Performances and Physiological and Yield Responses of Potato (Solanum tuberosum L.). Agronomy 2020, 10, 352. https://doi.org/10.3390/agronomy10030352
Lombardo S, Pandino G, Mauromicale G. Optimizing Nitrogen Fertilization to Improve Qualitative Performances and Physiological and Yield Responses of Potato (Solanum tuberosum L.). Agronomy. 2020; 10(3):352. https://doi.org/10.3390/agronomy10030352
Chicago/Turabian StyleLombardo, Sara, Gaetano Pandino, and Giovanni Mauromicale. 2020. "Optimizing Nitrogen Fertilization to Improve Qualitative Performances and Physiological and Yield Responses of Potato (Solanum tuberosum L.)" Agronomy 10, no. 3: 352. https://doi.org/10.3390/agronomy10030352