Crocus sativus L. Ecotypes from Mediterranean Countries: Phenological, Morpho-Productive, Qualitative and Genetic Traits

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Experimental Design
2.2. Phenological and Morpho-Productive Traits
2.3. Qualitative Analysis
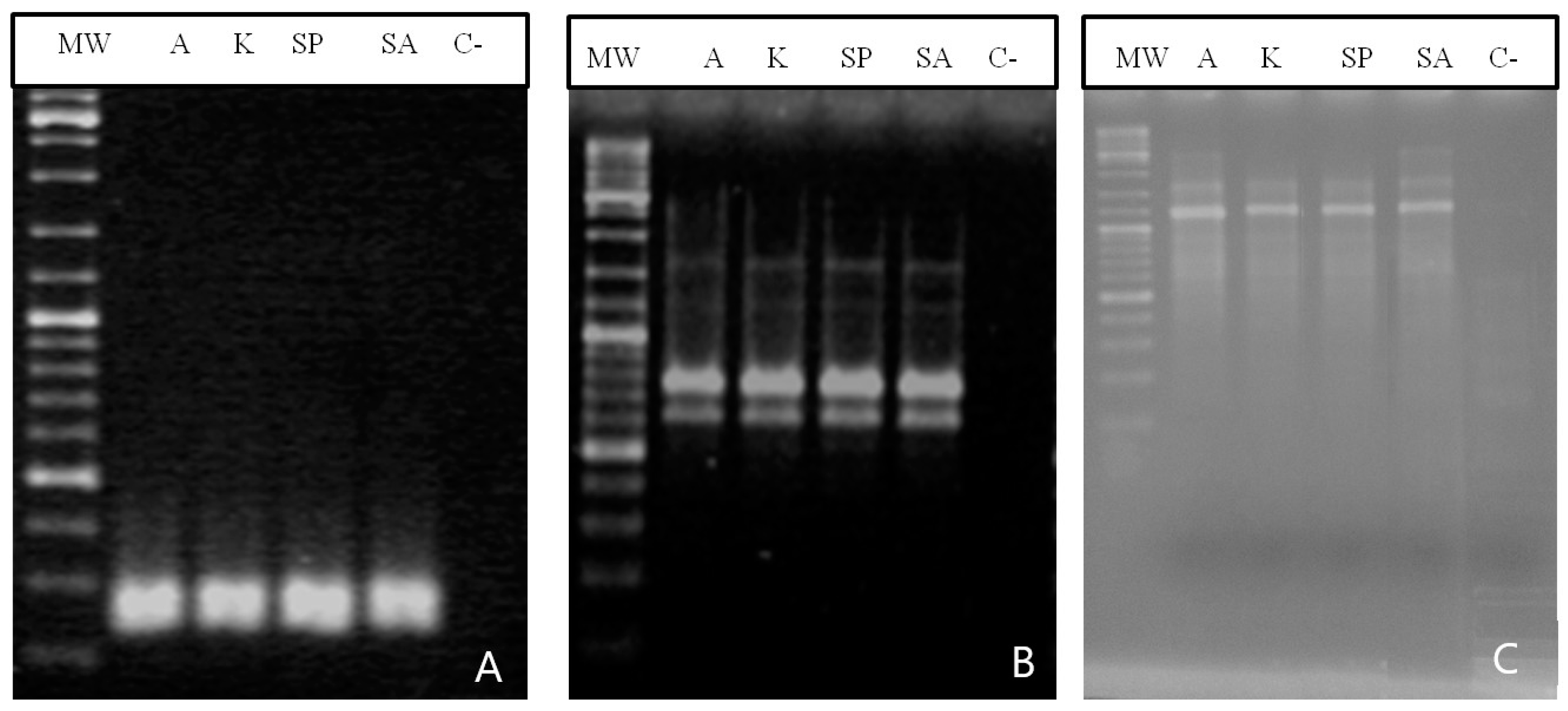
2.4. Molecular Analysis
2.4.1. SSRs Analysis
2.4.2. RAPD Analysis
2.4.3. SRAP Analysis
2.5. Statistical Analysis
3. Results
3.1. Climatic Data
3.2. Flower Morphological Traits and Colorimetric Coordinates of Tepals
3.3. Phenological, Productive and Vegetative Development Traits
3.4. Qualitative Traits
3.5. Molecular Analysis
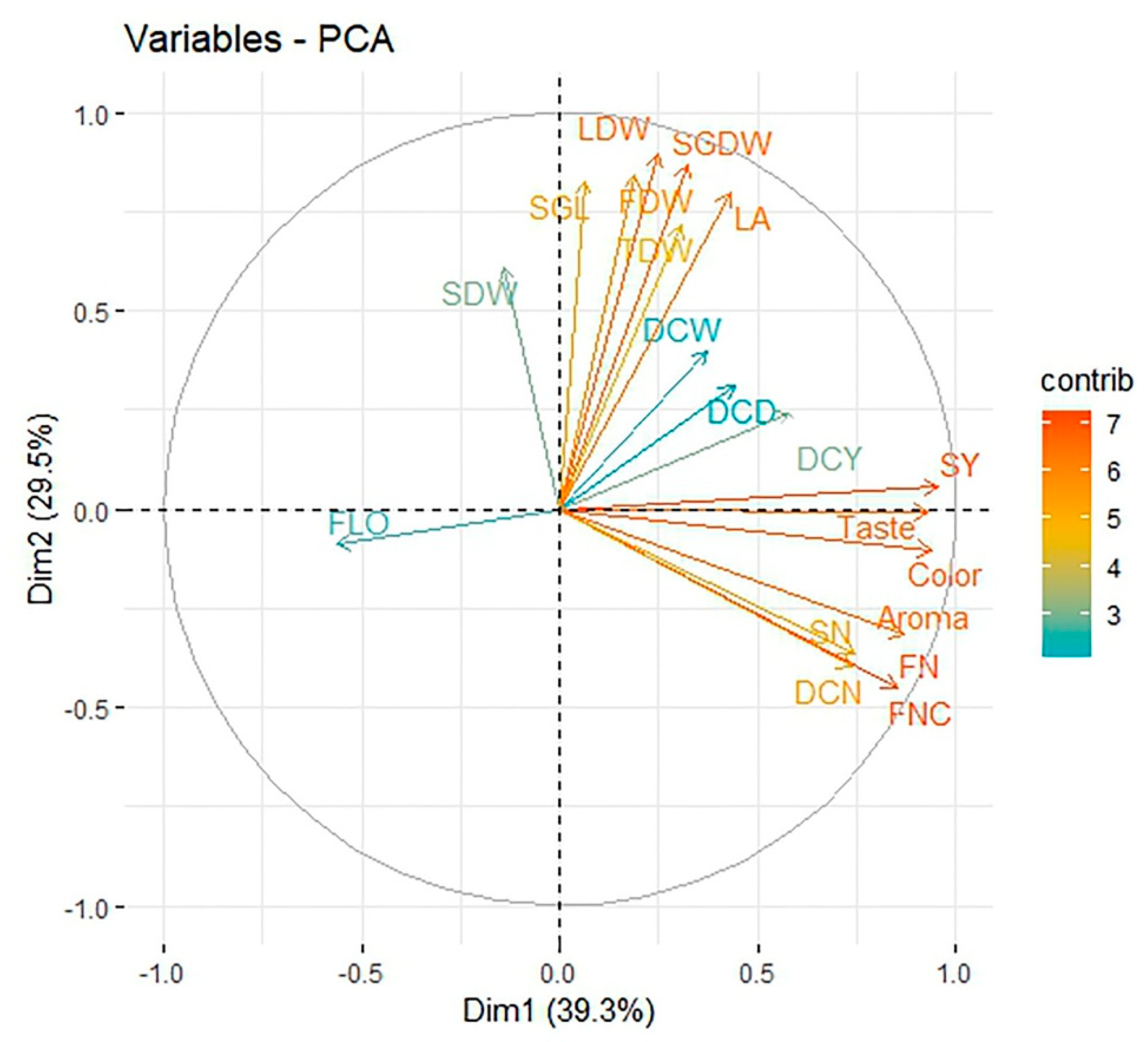
3.6. Principal Component Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bagur, M.J.; Salinas, G.L.A.; Jiménez-Monreal, A.M.; Chaouqi, S.; Llorens, S.; Martínez-Tomé, M.; Alonso, G.L. Saffron: An old medicinal plant and a potential novel functional food. Molecules 2018, 23, 30. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.Z.; Bathaie, S.Z. Historical uses of saffron: Identifying potential new avenues for modern research. Avicenna J. Phytomed. 2011, 1, 57–66. [Google Scholar]
- Nemati, Z.; Harpke, D.; Gemicioglu, A.; Kerndorff, H.; Blattner, F.R. Saffron (Crocus sativus) is an autotriploid that evolved in Attica (Greece) from wild Crocus cartwrightianus. Mol. Phylogenet. Evol. 2019, 136, 14–20. [Google Scholar] [CrossRef]
- Cardone, L.; Castronuovo, D.; Perniola, M.; Cicco, N.; Candido, V. Saffron (Crocus sativus L.), the king of spices: An overview. Sci. Hortic. 2020, 272, 109560. [Google Scholar] [CrossRef]
- De-Los-Mozos-Pascual, M.; Fernández, J.A.; Roldán, M. Preserving biodiversity in saffron: The Crocusbank project and the world saffron and Crocus collection. Acta Hortic. 2010, 850, 23–28. [Google Scholar] [CrossRef]
- Fernandez, J.A. Genetic resources of saffron and allies (Crocus spp.). Acta Hortic. 2007, 739, 167–185. [Google Scholar] [CrossRef]
- Husaini, A.M. Challenges of climate change: Omics-based biology of saffron plants and organic agricultural biotechnology for sustainable saffron production. GM Crops Food 2014, 5, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Mitsopoulou, T.; Tsimidou, M.Z. Morphological characteristics of greek saffron stigmas from kozani region. Acta Hortic. 2004, 650, 189–193. [Google Scholar] [CrossRef]
- Fernández, J.A.; Santana, O.; Guardiola, J.L.; Molina, R.V.; Heslop-Harrison, P.; Borbely, G.; Branca, F.; Argento, S.; Maloupa, E.; Talou, T.; et al. The world saffron and Crocus collection: Strategies for establishment, management, characterisation and utilisation. Genet. Resour. Crop. Evol. 2011, 58, 125–137. [Google Scholar] [CrossRef] [Green Version]
- CROCUSBANK: Genetic Resources of Saffron and Allies. Available online: http://www.crocusbank.org (accessed on 18 November 2020).
- Fallahi, H.R.; Aghhavani-Shajari, M.; Sahabi, H.; Behdani, M.A.; Sayyari-Zohan, M.H.; Vatandoost, S. Influence of some pre and post-harvest practices on quality of saffron stigmata. Sci. Hortic. 2020, 278, 109846. [Google Scholar] [CrossRef]
- Baghalian, K.; Sheshtamand, M.S.; Jamshidi, A.H. Genetic variation and heritability of agro-morphological and phytochemical traits in Iranian saffron (Crocus sativus L.) populations. Ind. Crops Prod. 2010, 31, 401–406. [Google Scholar] [CrossRef]
- Amirnia, R.; Bayat, M.; Gholamian, A. Influence of corm provenance and sowing dates on stigma yield and yield components in saffron (Crocus sativus L.). Turkish J. Field Crops 2013, 18, 198–204. [Google Scholar]
- Cardone, L.; Castonuovo, D.; Perniola, M.; Cicco, N.; Candido, V. Evaluation of corm origin and climatic conditions on saffron (Crocus sativus L.) yield and quality. J. Sci. Food Agric. 2019, 99, 5858–5869. [Google Scholar] [CrossRef] [PubMed]
- Soukrat, S.; Metougui, M.L.; Gabone, F.; Nehvi, F.; Abousalim, S.; Benlahabib, O. Study of diversity in some Moroccan population of saffron (Crocus sativus L.). Afr. J. Agric. Res. 2019, 14, 759–769. [Google Scholar]
- Mondini, L.; Noorani, A.; Pagnotta, M.A. Assessing plant genetic diversity by molecular tools. Diversity 2009, 1, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Siracusa, L.; Gresta, F.; Avola, G.; Albertini, E.; Raggi, L.; Marconi, G.; Lombardo, G.M.; Ruberto, G. Agronomic, chemical and genetic variability of saffron (Crocus sativus L.) of different origin by LC-UV-vis-DAD and AFLP analyses. Genet. Resour. Crop. Evol. 2013, 60, 711–721. [Google Scholar] [CrossRef]
- Torricelli, R.; Javan, I.Y.; Albertini, E.; Venanzoni, R.; Hosseinzadeh, Y.G. Morphological and molecular characterization of Italian, Iranian and Spanish saffron (Crocus sativus L.) accessions. Appl. Ecol. Environ. Res. 2019, 17, 1875–1887. [Google Scholar] [CrossRef]
- Grilli-Caiola, M.; Caputo, P.; Zanier, R. RAPD analysis in Crocus sativus L. accensions and related Crocus species. Bio. Plant. 2004, 48, 375–380. [Google Scholar] [CrossRef]
- Babaei, S.; Talebia, M.; Bahara, M.; Zeinali, H. Analysis of genetic diversity among saffron (Crocus sativus) accession from different regions of Iran as revealed by SRAP markers. Sci. Hortic. 2014, 171, 27–31. [Google Scholar] [CrossRef]
- Rubio-Moraga, A.; Castillo-López, R.; Gómez-Gómez, L.; Ahrazem, O. Saffron is a monomorphic species as revealed by RAPD, ISSR and microsatellite analyses. BMC Res. Notes 2009, 2, 189–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemati, Z.; Zeinalabedini, M.; Mardi, M.; Pirseyediand, S.M.; Marashi, S.H.; Khayam Nekoui, S.M. Isolation and characterization of a first set of polymorphic microsatellite markers in saffron, Crocus sativus (Iridaceae). Am. J. Bot. 2012, 99, e340–e343. [Google Scholar] [CrossRef] [PubMed]
- Fluch, S.; Hohl, K.; Stierschneider, M.; Kopecky, D.; Kaar, B. Crocus sativus L.—Molecular evidence on its clonal origin. Acta Hortic. 2010, 850, 41–46. [Google Scholar] [CrossRef]
- Keify, F.; Beiki, A.H. Exploitation of random amplified polymorphic DNA (RAPD) and sequence-related amplified polymorphism (SRAP) markers for genetic diversity of saffron collection. J. Med. Plants Res. 2012, 6, 2761–2768. [Google Scholar]
- Anabat, M.M.; Riahi, H.; Sheidai, M.; Koohdar, F. Population genetic study and barcoding in Iran saffron (Crocus sativus L.). Ind. Crops Prod. 2020, 143, 111915. [Google Scholar] [CrossRef]
- Mir, M.A.; Mansoor, S.; Sugapriya, M.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Deciphering genetic diversity analysis of saffron (Crocus sativus L.) using RAPD and ISSR markers. Saudi J. Biol. Sci. 2020, 28, 1308–1317. [Google Scholar] [CrossRef]
- Zalacain, A.; Ordoudi, S.A.; Díaz-Plaza, E.M.; Carmona, M.; Blázquez, I.; Tsimidou, M.Z.; Alonso, G.L. Near-infrared spectroscopy in saffron quality control: Determination of chemical composition and geographical origin. J. Agric. Food Chem. 2005, 53, 9337–9341. [Google Scholar] [CrossRef]
- Anastasaki, E.; Kanakis, C.; Pappas, C.; Maggi, L.; del Campo, C.P.; Carmona, M.; Polissiou, M.G. Differentiation of saffron from four countries by mid-infrared spectroscopy and multivariate analysis. Eur. Food Res. Technol. 2010, 230, 571–577. [Google Scholar] [CrossRef]
- Anastasaki, E.; Kanakis, C.; Pappas, C.; Maggi, L.; Del Campo, C.P.; Carmona, M.; Alonso, G.L.; Polissiou, M.G. Geographical differentiation of saffron by GC–MS/FID and chemometrics. Eur. Food Res. Technol. 2009, 229, 899–905. [Google Scholar] [CrossRef]
- Carmona, M.; Martinez, J.; Zalacain, A.; Rodriguez-Mendez, M.L.; de Saja, J.A.; Alonso, G.L. Analysis of saffron volatile fraction by TD-GC-MS and e-nose. Eur. Food Res. Technol. 2006, 223, 96–101. [Google Scholar] [CrossRef]
- Ghanbari, J.; Khajoei-Nejad, G.; van Ruth, S.M. Effect of saffron (Crocus sativus L.) corm provenance on its agro-morphological traits and bioactive compounds. Sci. Hortic. 2019, 256, 108605. [Google Scholar] [CrossRef]
- D’Archivio, A.A.; Di Pietro, L.; Maggi, M.A.; Rossi, L. Optimization using chemometrics of HS-SPME/GC-MS profiling of saffron aroma and identification of geographical volatile markers. Eur. Food Res. Technol. 2018, 244, 1605–1613. [Google Scholar] [CrossRef]
- Bhandari, H.R.; Bhanu, A.N.; Srivastava, K.; Singh, M.N.; Shreya, H.A. Assessment of genetic diversity in crop plants—An overview. Adv. Plants Agric. Res. 2017, 7, 00255. [Google Scholar]
- Small, E. Saffron (Crocus sativus) the eco-friendly spice. Biodiversity 2016, 17, 162–170. [Google Scholar] [CrossRef]
- Khanali, M.; Farahani, S.S.; Shojaei, H.; Elhami, B. Life cycle environmental impacts of saffron production in Iran. Environ. Sci. Pollut. Res. 2017, 24, 4812–4821. [Google Scholar] [CrossRef] [Green Version]
- Abolhassani, L.; Khorramdel, S.; Reed, M.; Saghaian, S. Environmental Economic Analysis of Saffron Production. In Saffron: Science, Technology and Health; Koocheki, A., Khajeh-Hosseini, M., Eds.; Woodhead Publishing: Sawston, UK, 2020; pp. 367–390. [Google Scholar]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Soil Survery Staff. Keys to Soil Taxonomy; United States Department of Agriculture, Soil Conservation Service: Washington, DC, USA, 2010.
- Ministero per le Politiche Agricole. Metodi Ufficiali di Analisi Chimica del Suolo; D.M. del 13/09/99, Gazzetta Ufficiale n. 248 del 21/10/99; Ministero per le Politiche Agricole: Rome, Italy, 1999. [Google Scholar]
- Saleem, M.H.; Ali, S.; Rehman, M.; Rana, M.S.; Rizwan, M.; Kamran, M.; Imran, M.; Riaz, M.; Soliman, M.H.; Elkelish, A.; et al. Influence of phosphorus on copper phytoextraction via modulating cellular organelles in two jute (Corchorus capsularis L.) varieties grown in a copper mining soil of Hubei Province, China. Chemosphere 2020, 248, 126032. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.H.; Fahad, S.; Khan, S.U.; Ahmar, S.; Khan, M.H.U.; Rehman, M.; Maqbool, Z.; Liu, L. Morpho-physiological traits, gaseous exchange attributes, and phytoremediation potential of jute (Corchorus capsularis L.) grown in different concentrations of copper-contaminated soil. Ecotoxicol. Environ. Safe. 2020, 189, 109915. [Google Scholar] [CrossRef]
- Hameed, A.; Akram, N.A.; Saleem, M.H.; Ashraf, M.; Ahmed, S.; Ali, S.; Alsahli, A.A.; Alyemeni, M.N. Seed Treatment with α-Tocopherol Regulates Growth and Key Physio-Biochemical Attributes in Carrot (Daucus carota L.) Plants under Water Limited Regimes. Agronomy 2021, 11, 469. [Google Scholar] [CrossRef]
- Cardone, L.; Castronuovo, D.; Perniola, M.; Scrano, L.; Cicco, N.; Candido, V. The Influence of Soil Physical and Chemical Properties on Saffron (Crocus sativus L.) Growth, Yield and Quality. Agronomy 2020, 10, 1154. [Google Scholar] [CrossRef]
- Konica Minolta. 2003. Available online: https://www.konicaminolta.eu/fileadmin/content/eu/Measuring Instruments/4 Learning Centre/C A/PRECISE COLORCOMMUNICATION/pcc english 13.pdf (accessed on 3 December 2020).
- International Organization for Standardization. ISO 3632-1. Spices—Saffron (Crocus sativus L.). Part 1: Specification, 2nd ed; International Organization for Standardization: Geneva, Switzerland, 2011; p. 6. [Google Scholar]
- International Organization for Standardization. ISO 3632-2. Spices—Saffron (Crocus sativus L.). Part 2: Test Methods, 1st ed; International Organization for Standardization: Geneva, Switzerland, 2010; p. 42. [Google Scholar]
- Van de Wouw, M.; Kik, C.; van Hintum, T.; van Treuren, R.; Visser, B. Genetic erosion in crops: Concept, research results and challenges. Plant Genet. Resour. 2010, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Edreva, A.; Velikova, V.; Tsonev, T.; Dagnon, S.; Gürel, A.; Aktaş, L.; Gesheva, E. Stress-protective role of secondary metabolites: Diversity of functions and mechanisms. Gen. Appl. Plant Physiol. 2008, 34, 67–78. [Google Scholar]
- Molina, R.V.; Valero, M.; Navarro, Y.; Guardiola, J.L.; Garcia-Luis, A. Temperature effects on flower formation in saffron (Crocus sativus L.). Sci. Hortic. 2005, 103, 361–379. [Google Scholar] [CrossRef]
- Grimes, S.J.; Phillips, T.D.; Hahn, V.; Capezzone, F.; Graeff-Hönninger, S. Growth, Yield Performance and Quality Parameters of Three Early Flowering Chia (Salvia hispanica L.) Genotypes Cultivated in Southwestern Germany. Agriculture 2018, 8, 154. [Google Scholar] [CrossRef] [Green Version]
- Wehner, T.C.; Guner, N. Growth stage, flowering pattern, yield, and harvest date prediction of four types of cucumber tested at 10 planting dates. Acta Hortic. 2004, 637, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Ojaq, S.M.; Mozafari, H.; Jabbari, H.; Sani, B. Evaluation of yield of safflower (Carthamus tinctorius L.) genotypes under semi-arid conditions. Plant Genet. Resour. 2020, 18, 270–277. [Google Scholar] [CrossRef]
- Yerima, A.R.I.B.; Achigan-Dako, E.G.; Aissata, M.; Sekloka, E.; Billot, C.; Adje, C.O.; Barnaud, A.; Bakasso, Y. Agromorphological Characterization Revealed Three Phenotypic Groups in a Region-Wide Germplasm of Fonio (Digitaria exilis (Kippist) Stapf) from West Africa. Agronomy 2020, 10, 1653. [Google Scholar] [CrossRef]
- Bayat, M.; Amirnia, R.; Tajbakhsh, M.; Ramezani, M. Evaluation of Saffron Ecotypes for Stigma Yield and Yield Components Using Different Maternal Corm Weights. J. Plant Physiol. Breed. 2016, 6, 53–64. [Google Scholar]
- Soheilivand, S.; Agayev, Y.; Shakib, A.M.; Fathi, M.; Rezvani, E. Comparison of diversity in flowering rate of two saffron (Crocus sativus) populations of Iran. Acta Hortic. 2007, 739, 303–307. [Google Scholar] [CrossRef]
- Renau-Morata, B.; Nebauer, S.G.; Sánchez, M.; Molina, R.V. Effect of corm size, water stress and cultivation conditions on photosynthesis and biomass partitioning during the vegetative growth of saffron (Crocus sativus L.). Ind. Crop. Prod. 2012, 39, 40–46. [Google Scholar] [CrossRef]
- García-Rodríguez, M.V.; López-Córcoles, H.; Alonso, G.L.; Pappas, C.S.; Polissiou, M.G.; Tarantilis, P.A. Comparative evaluation of an ISO 3632 method and an HPLC-DAD method for safranal quantity determination in saffron. Food Chem. 2017, 221, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Masi, E.; Taiti, C.; Heimler, D.; Vignolini, P.; Romani, A.; Mancuso, S. PTR-TOF-MS and HPLC analysis in the characterization of saffron (Crocus sativus L.) from Italy and Iran. Food Chem. 2016, 192, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Lage, M.; Cantrell, C.L. Quantification of saffron (Crocus sativus L.) metabolites crocins, picrocrocin and safranal for quality determination of the spice grown under different environmental Moroccan conditions. Sci. Hortic. 2009, 121, 366–373. [Google Scholar] [CrossRef]
- Cossignani, L.; Urbani, E.; Simonetti, M.S.; Maurizi, A.; Chiesi, C.; Blasi, F. Characterisation of secondary metabolites in saffron from central Italy (Cascia, Umbria). Food Chem. 2014, 143, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Kanakis, C.D.; Daferera, D.J.; Tarantilis, P.A.; Polissiou, M.G. Qualitative determination of volatile compounds and quantitative evaluation of safranal and 4-hydroxy-2,6,6-trimethyl-1-cyclohexene-1-carboxaldehyde (HTCC) in Greek saffron. J. Agric. Food Chem. 2004, 52, 4515–4521. [Google Scholar] [CrossRef]
- Gresta, F.; Avola, G.; Lombardo, G.M.; Siracusa, L.; Ruberto, G. Analysis of flowering, stigmas yield and qualitative traits of saffron (Crocus sativus L.) as affected by environmental conditions. Sci. Hortic. 2009, 119, 320–324. [Google Scholar] [CrossRef]
- Grilli Caiola, M.; Di Somma, D.; Lauretti, P. Comparative study of pollen and pistil of Crocus sativus L. (Iridaceae) and its allied species. Ann. Bot. 2001, 1, 73–82. [Google Scholar]
- Casadesús, J.; Noyer-Weidner, M. Epigenetics. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 500–503. ISBN 9780080961569. [Google Scholar] [CrossRef]
- Busconi, M.; Soffritti, G.; Stagnati, L.; Marocco, A.; Martínez, J.M.; Pascual, M.D.L.M.; Fernandez, J.A. Epigenetic stability in Saffron (Crocus sativus L.) accessions during four consecutive years of cultivation and vegetative propagation under open field conditions. Plant Sci. 2018, 277, 1–10. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ecotype | City (Province)-Region | Country | Altitude | Coordinates | Climate 1 |
|---|---|---|---|---|---|
| Abruzzo | Navelli (L’Aquila)-Abruzzo | Italy | 805 m asl | 42°14′ N–13°43′ E | Cfb |
| Sardinia | San Gavino Monreale (Medio Campidano)-Sardinia | Italy | 100 m asl | 39°33′ N–8°48′ E | Csa |
| Castilla-La Mancha | Albacete-Castilla-La Mancha | Spain | 686 m asl | 38°59′ N–1°51′ W | BSk |
| Kozani | Krokos-Kozani | Greece | 720 m asl | 40°18′ N–21°47′ E | Cfa |
| Sep | Oct | Nov | Dec | Jan | Feb | Mar | Apr | May | Mean | |
|---|---|---|---|---|---|---|---|---|---|---|
| 2016–2017 | ||||||||||
| T.min (°C) | 16.0 | 12.9 | 9.8 | 5.3 | 1.8 | 6.4 | 7.9 | 9.0 | 13.6 | 9.2 |
| T.mean (°C) | 19.5 | 15.8 | 12.1 | 8.1 | 4.0 | 9.3 | 11.6 | 12.5 | 17.9 | 12.3 |
| T.max (°C) | 24.8 | 20.5 | 15.3 | 11.6 | 7.1 | 13.4 | 16.6 | 17.3 | 23.8 | 16.7 |
| Rainfall (mm) | 80.2 | 37.0 | 34.8 | 25.4 | 68.0 | 49.0 | 15.8 | 40.0 | 41.8 | 395.0 |
| 2017–2018 | ||||||||||
| T.min (°C) | 15.8 | 12.8 | 8.6 | 4.8 | 6.3 | 3.8 | 6.8 | 12.1 | 14.8 | 9.5 |
| T.mean (°C) | 19.6 | 16.2 | 11.1 | 7.9 | 9.2 | 6.7 | 9.9 | 16.1 | 18.9 | 12.9 |
| T.max (°C) | 25.2 | 21.3 | 14.6 | 13.8 | 15.2 | 12.2 | 15.9 | 23.2 | 25.0 | 18.5 |
| Rainfall (mm) | 52.4 | 14.0 | 62.6 | 26.4 | 14.4 | 62.4 | 47.8 | 10.8 | 47.4 | 338.2 |
| Experimental Factors 1 | Stigma Length (mm) | Stamen Length (mm) | Stigma Fresh Weight (g) | Stigma Dry Weight (g) | Stamen Fresh Weight (g) | Stamen Dry Weight (g) | Tepal Fresh Weight (g) | Tepal Dry Weight (g) | Flower Fresh Weight (g) | Flower Dry Weight (g) | L* | a* | b* |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ecotype (E) | |||||||||||||
| Abruzzo | 40.57 a | 23.92 | 0.0438 a | 0.0078 a | 0.0270 a | 0.0091 a | 0.2878 a | 0.0359 a | 0.3586 a | 0.0528 a | 49.31 | 25.13 | −20.92 |
| Sardinia | 36.80 b | 22.75 | 0.0360 b | 0.0065 b | 0.0269 a | 0.0082 b | 0.2829 a | 0.0324 ab | 0.3430 a | 0.0471 ab | 52.95 | 27.94 | −24.79 |
| Kozani | 36.63 bc | 22.65 | 0.0302 bc | 0.0055 c | 0.0241 a | 0.0075 bc | 0.2395 b | 0.0292 b | 0.2968 b | 0.0422 b | 50.07 | 26.37 | −20.64 |
| Spain | 34.50 c | 23.00 | 0.0260 c | 0.0061 bc | 0.0168 b | 0.0069 c | 0.2542 b | 0.0318 ab | 0.2970 b | 0.0448 b | 48.51 | 28.58 | −22.17 |
| Significance2 | ** | ns | ** | ** | * | ** | ** | * | ** | ** | ns | ns | ns |
| Year (Y) | |||||||||||||
| 2016 | 37.14 | 23.27 | 0.0352 | 0.0067 | 0.0265 | 0.0082 | 0.275 | 0.0348 | 0.3366 | 0.0497 | 47.54 | 28.27 | −22.22 |
| 2017 | 37.11 | 22.89 | 0.0328 | 0.0062 | 0.0209 | 0.0076 | 0.257 | 0.0299 | 0.3111 | 0.0437 | 52.87 | 25.74 | −22.04 |
| Significance | ns | ns | ns | ns | * | ns | ns | * | ns | * | ** | ** | ns |
| E × Y | |||||||||||||
| Significance | ** | ns | ** | ** | ** | ** | ns | ** | ns | ** | ** | ** | ns |
| Experimental Factors 1 | Days to Flower (d) | Flowering Interval (d) | Flower Number Corm−1 | Flower Number (n m−2) | Stigma Yield (kg ha−1) | Daughter Corm Yield (t ha−1) |
|---|---|---|---|---|---|---|
| Ecotype (E) | ||||||
| Abruzzo | 41.0 ab | 21.7 a | 2.08 c | 83.12 c | 6.58 b | 18.68 a |
| Sardinia | 37.0 b | 23.0 a | 3.61 a | 144.23 a | 9.26 a | 18.77 a |
| Kozani | 43.0 a | 16.0 b | 2.10 c | 82.68 c | 4.59 c | 15.13 b |
| Spain | 40.5 ab | 16.0 b | 2.83 b | 113.27 b | 6.89 b | 19.03 a |
| Significance2 | * | * | ** | ** | ** | * |
| Year (Y) | ||||||
| 2016 | 38.75 | 20.3 | 2.69 | 105.86 | 7.11 | 16.18 |
| 2017 | 42.00 | 18.1 | 2.64 | 105.79 | 6.54 | 19.63 |
| Significance | ns | ns | ns | ns | ns | * |
| E × Y | ||||||
| Significance | ns | ns | ns | ns | ns | ** |
| Experimental Factors 1 | Leaf Number (n Plant−1) | Leaf Length (cm) | Leaf Fresh Weight (g) | Leaf Dry Weight (g) | Leaf Area (cm2 Plant−1) | Shoot Number (n Plant−1) | Daughter Corm Weight (g) | Daughter Corm Number (n Plant−1) | Daughter Corm Diameter (cm) |
|---|---|---|---|---|---|---|---|---|---|
| Ecotype (E) | |||||||||
| Abruzzo | 40.45 | 33.90 | 14.48 a | 5.14 a | 240.58 a | 6.28 b | 7.63 a | 6.37 b | 2.31 a |
| Sardinia | 42.25 | 30.37 | 9.36 b | 3.21 b | 197.63 b | 8.63 a | 6.76 ab | 9.12 a | 2.22 a |
| Kozani | 41.38 | 30.80 | 7.15 c | 2.30 c | 149.82 d | 6.22 b | 4.17 b | 6.57 b | 1.82 b |
| Spain | 41.17 | 31.65 | 10.11 b | 3.29 b | 185.34 c | 8.25 a | 4.72 b | 8.58 a | 2.18 a |
| Significance2 | ns | ns | ** | ** | ** | * | * | * | * |
| Year (Y) | |||||||||
| 2016 | 34.93 | 27.51 | 9.70 | 3.42 | 183.62 | 7.67 | 4.14 | 8.08 | 1.96 |
| 2017 | 47.69 | 35.85 | 10.85 | 3.55 | 203.07 | 7.03 | 7.50 | 7.24 | 2.30 |
| Significance | ** | ** | ns | ns | ** | ns | ** | ns | * |
| E × Y | |||||||||
| Significance | ** | ** | * | ns | ** | * | ** | * | * |
| Experimental Factors 1 | Moisture and Volatile Matter (%) 3 | A1%1cm 4 (440 nm) | ISO 5 Reference (Crocetin Esters) | A1%1cm 6 (257 nm) | ISO 7 Reference (Picrocrocin) | A1%1cm 8 (330 nm) | ISO 9 Reference (Safranal) |
|---|---|---|---|---|---|---|---|
| Ecotype (E) | |||||||
| Abruzzo | 4.58 | 242.53 c | I | 88.61 b | I | 26.06 b | I |
| Sardinia | 4.67 | 266.03 a | I | 96.95 a | I | 30.19 a | I |
| Kozani | 4.04 | 223.83 d | I | 80.38 c | I | 24.94 b | I |
| Spain | 4.04 | 251.07 b | I | 89.48 b | I | 28.14 ab | I |
| Significance2 | ns | ** | ** | * | |||
| Year (Y) | |||||||
| 2016 | 4.29 | 244.01 | I | 87.74 | I | 25.88 | I |
| 2017 | 4.35 | 247.73 | I | 89.99 | I | 28.66 | I |
| Significance | ns | ns | ns | ||||
| E × Y | |||||||
| Significance | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardone, L.; Castronuovo, D.; Perniola, M.; Cicco, N.; Molina, R.V.; Renau-Morata, B.; Nebauer, S.G.; Candido, V. Crocus sativus L. Ecotypes from Mediterranean Countries: Phenological, Morpho-Productive, Qualitative and Genetic Traits. Agronomy 2021, 11, 551. https://doi.org/10.3390/agronomy11030551
Cardone L, Castronuovo D, Perniola M, Cicco N, Molina RV, Renau-Morata B, Nebauer SG, Candido V. Crocus sativus L. Ecotypes from Mediterranean Countries: Phenological, Morpho-Productive, Qualitative and Genetic Traits. Agronomy. 2021; 11(3):551. https://doi.org/10.3390/agronomy11030551
Chicago/Turabian StyleCardone, Loriana, Donato Castronuovo, Michele Perniola, Nunzia Cicco, Rosa V. Molina, Begoña Renau-Morata, Sergio G. Nebauer, and Vincenzo Candido. 2021. "Crocus sativus L. Ecotypes from Mediterranean Countries: Phenological, Morpho-Productive, Qualitative and Genetic Traits" Agronomy 11, no. 3: 551. https://doi.org/10.3390/agronomy11030551