
Are Copper Nanoparticles Toxic to All Plants? A Case Study on Onion (Allium cepa L.)


1
Agricultural Botany Department—Plant Physiology Division, Faculty of Agriculture, Cairo University, Giza 12613, Egypt
2
Agriculture Research Centre, Soil, Water and Environment Institute, Giza 12613, Egypt
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(5), 1006; https://doi.org/10.3390/agronomy11051006
Submission received: 9 April 2021
/
Revised: 7 May 2021
/
Accepted: 10 May 2021
/
Published: 19 May 2021
(This article belongs to the Special Issue Nano(bio)technologies and Nano(bio)materials Used in Plant Protection and Fertilization)
Abstract
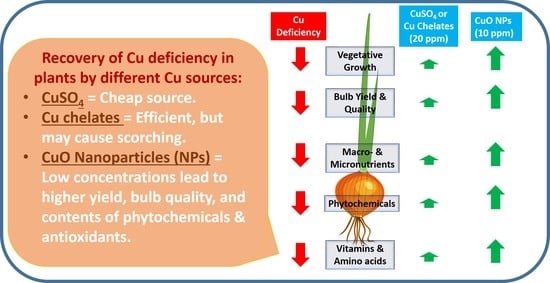
:Sandy soils with high alkalinity are characterized by low copper (Cu) contents that lead to many deficiency symptoms in plants. Cu deficiency in plants can be corrected using several cheap Cu sources. Nevertheless, the effects that novel sources, such as Cu nanoparticles (NPs), have on plants remain poorly studied. In the present work, we investigated the effect and efficiency of Cu supplementation to onion (Allium cepa L.) plants using Cu sulfate, chelate, or NPs, and compared their effects on bulb quality, yield, and contents of phytochemicals. Two successive seasons (2018/2019 and 2019/2020) of field experiments were conducted in newly reclaimed sandy soils, where plants were sprayed with either 10 ppm CuO NPs, 20 ppm CuSO4·5H2O, or 20 ppm of Cu chelates. Overall, Cu deficiency (control) resulted in a significant decrease in yield and all quality traits of onion plants. CuO NPs treatment significantly enhanced growth parameters, including plant height, number of leaves, fresh and dry weight, yield, and bulb quality, compared with Cu sulfate and chelates. This was also the case regarding chemical constituents such as macro- and micro-nutrients, total soluble solids, phytochemical compounds, vitamins, and amino acids. Although Cu sulfate is the cheapest form used for Cu supplementation, results of the present study suggest that CuO NPs was not only safe to use, but also was the treatment that led to the highest onion yield and quality.
1. Introduction
Onion is one of the most consumed vegetable crops around the world, with recent statistics estimating a worldwide production of about 96.8 million tons [1]. Besides its culinary uses, onion tissues have a wide variety of phytochemicals such as polyphenolics, flavonoids, ascorbic acid, sapogenins and saponins, sulfur compounds, anthocyanins, organic acids, aldehydes, and ketones [2]. These phytochemicals act in preventing and curing many diseases such as diabetes, hypertension, hypocholesterolemia, and atherosclerosis [3,4]. The levels of biochemical components and morpho-physiology of onion are closely related to the quality of the fertilization regime and soil conditions in which it is cultivated. A recent review by Khokhar [5] summarized the importance of micronutrients’ application to onion and reported the crucial role that iron (Fe), zinc (Zn), boron (B), and copper (Cu) have in increasing its yield and quality. Among these micronutrients, Cu is one of the least studied. Cu is an essential micronutrient that forms part of many important proteins and enzymes in plants and plays an important role in metabolism [6]. The effects of Cu deficiency manifest first in younger leaves because Cu is an immobile nutrient [7]. Onion plants are regarded as sensitive to Cu deficiency, and distinct symptoms thus become visible on Cu-deficient plants. Cu deficiency turns leaf tips white and bent at right angles. As Cu deficiency continues, leaves become chlorotic and necrotic while bulbs become pale and thin with fragile skins. Cu-deficient onion plants also mature early and produce greenish-bronze bulbs [8]. Cu deficiency is usually accentuated in sandy soils characterized by low contents of inorganic matter and high pH values, which are prevalent in many agricultural lands [9]. It is therefore important to adopt appropriate crop management practices in order to prevent Cu deficiency, increase both Cu use efficiency and onion yields, and reduce the overall costs of fertilizers.
Cu deficiency can be corrected by soil or foliar application of different Cu sources. A cheap and most commonly used source is Cu sulfate (CuSO4·5H2O), which contains approximately 25% elemental Cu. Soil application of Cu sulfate in sandy soils is reported to increase the thickness of onion scales and change their color from pale to brilliant brown [10]. However, foliar application of Cu sulfate may cause scorching to the leaves. Another alternative source is Cu chelates, which was reported to be more effective in soils of high pH values [8]. Recently, Cu nanoparticles (NPs) were introduced as a novel form of this micronutrient in agricultural studies. Nevertheless, Kasana et al. [11] reviewed the effects that Cu NPs have on crops and concluded that they do not have much effect on germination or growth of plants. Nevertheless, this conclusion has several shortcomings and is contradicted by several reports in the literature. First, treating lettuce seeds with 0.066% (w/w) Cu NPs was reported to increase shoot:root ratio of 15-day seedlings by 91% [12]. Second, many of the studies included in the above-mentioned review used extremely high doses of Cu, reaching levels of 1000 mg/L (ppm), which are very toxic to plants. Finally, and most importantly, the aforementioned review did not take into consideration that plants have different responses to Cu supplementation. This is important to consider because, while a certain Cu level might be toxic to one plant, a much higher concentration can be beneficial to another. For example, Arabidopsis thaliana seems to be very sensitive to Cu NPs supplementation, where only 2 mg/L of Cu markedly reduces its biomass [13]. This, however, is not the case in some Allium species that tolerate high doses of Cu. In fact, 50 ppm of Cu chelates was reported to significantly increase onion yields [14]. In addition, a recent study reported that greenhouse grown Allium fistulosum (green onions) positively responded to three different Cu nutrition sources (bCuO, CuO NPs, and CuSO4·5H2O) at high concentrations reaching up to 600 mg/kg, which enhanced the nutrient and allicin contents of treated plants [15].
Although many reports have described the effects of Cu sulfates and chelates on yield and quality of crops, very few studies are available regarding the effects that Cu NPs have on nutrient elements, vitamins, amino acids, yield, and quality of crops in general, and of onion in particular. We hypothesize that CuO NPs have a higher efficiency in enhancing agronomical and biochemical attributes of onion bulbs, compared with other Cu forms, due to their optimum size and penetration ability to biological tissues. Thus, the aim of this study was to compare the efficiency of CuO NPs with the traditional Cu sulfate and chelated fertilizer forms in accumulating macro- and micro-nutrients, vitamins, amino acids, as well as on yield and quality characteristics in onion bulbs. To the authors’ best knowledge, this is the first study that describes the effects and efficiency of using CuO NPs as a novel source to amend onion plants grown in Cu-deficient soils.
2. Materials and Methods
2.1. Experimental Location
The present investigation was carried out in an open field at the newly reclaimed desert area of Wadi El-Notron station, Beheira Governorate, Egypt (Longitude 28°54′ E, Latitude 28°20′ N, and Altitude 130 m), throughout two seasons (2018/19 and 2019/20). Before planting, physical and chemical analyses (Table 1) of the reclaimed soil were performed at the Soil, Water, and Environment Research Institute, Agriculture Research Centre (A.R.C.), Egypt, according to Richards [16] and Jackson [17].
2.2. Soil Preparation and Organic Matter
Before planting, the soil was mechanically ploughed at an approximate depth of 30–40 cm. Then, compost (plant residues from legume crops obtained from Agriculture Research Center, Giza, Egypt) was incorporated into the soil 14 days before planting at the rate of 2.5 t ha−1. Table 2 shows the chemical properties of the compost. Afterwards, the soil was planked twice until the soil surface had settled.
2.3. Irrigation System
Irrigation water was supplied through a drip irrigation network using (4 L h−1) drippers according to Karmeli and Keller [18].
2.4. Plant Material and Transplantation of Seedlings
Seeds of onion cultivar ‘Giza 20′ were sown in a nursery on 15 October in both seasons. The media for seed germination was a mixture of peat moss and vermiculite (1:3 ratio) (obtained from Agriculture Research Center, Giza, Egypt). On 1 December of each season, 15 cm high uniform seedlings were transplanted to the open field. Onion transplants were treated before cultivation with fungicides (Topsin M 70% w.p./vitavax 300% w.p., Wadi El-Nile Co., Giza, Egypt) at 2.5 g kg−1. This treatment was used to prevent any fungal damage during cultivation. The seedlings were sown at a distance of 20 cm between plants and 50 cm between rows. Herbicide (Stomp®, BASF PLC, Cheshire, UK) was sprayed after transplanting of seedlings and was followed by irrigation to check the growth of the weeds at the early growth stage. The recommended cultural practices to raise healthy onion bulbs were followed.
2.5. Fertilizer Application and Harvest
Fertilizers were applied at the dose recommended by the Ministry of Agriculture and Land Reclamation, Egypt. All experimental plots were supplemented with ammonium sulfate (20.5%), superphosphate (15.5%), and potassium sulfate (52%) at 690, 575, and 115 kg/ha, respectively (Obtained from Wadi El-Nile Co., Giza, Egypt). Onion bulbs were harvested in mid-May in both seasons. Harvesting of onion bulbs was performed according to the recommendations of the Ministry of Agriculture and Land reclamation, Egypt. Bulbs were manually harvested when 75% of plants showed neck fall or top die-down. The bulbs were dried (curing) in a naturally ventilated area, then the necks were trimmed, leaving 2–3 cm attached to the bulbs. The bulbs were roughed at this stage based upon their color, shape, and size. Any damaged, twin bulbs, or long necked bulbs, if any, were discarded.
2.6. Synthesis of Copper Oxide Nanoparticles
Reagents of analytical grade (Sigma, USA) were used to prepare CuO nanoparticles from its precursor chemical compound copper sulfate (CuSO4). Nanoparticles were obtained by top to bottom molecular chemical and physical combined methods according to Zhang et al. [19], with some modifications. 1 M sodium hydroxide (NaOH) solution was added slowly (drop by drop) to aqueous solution of copper sulfate in a molar ratio of 1:3 under vigorous stirring for 10 h, discontinuously (2 days). The obtained precipitate was then mixed with 2 N citric acid monohydrate for 3 h. The obtained precipitate was filtered and washed thoroughly with ionized water in a mixed water/hexane system using a high-speed stirrer, and then washed again with ionized water alone for 2 h. The precipitate was finally dried in an oven at 108 °C for 24 h, then exposed to 1.5 psi of pressure for 5 days, discontinuously (6 h per day).
2.7. Characterization of CuO Nanoparticles
The size and shape of CuO nanoparticles were detected directly by transmission electron microscopy (TEM) using an electron acceleration voltage of 60 kV. The TEM samples were prepared by dropping a few drops of the solution on a carbon-coated copper grid (., microgrid B, Oken Shoji Co. Ltd, Tokyo, Japan) and inspecting it under the TEM microscope. A representative image of the produced CuO nanoparticles is shown in Supplementary Figure S1.
2.8. Experimental Design
The four treatments were compared in a completely randomized block design (RBCD) of five replicates as follows:
- NPK+ without foliar spray (Control).
- NPK+ copper sulfate foliar spray at rate of 20 ppm L−1 (CuSO4).
- NPK+ EDTA chelated copper foliar spray at rate of 20 ppm L−1 (Cu-EDTA).
- NPK+ copper oxide nanoparticles (CuO-NPs) mixed with irrigation water at the first spray, then foliar spray at a rate of 10 ppm L−1.
The above-mentioned concentrations of the three Cu treatments (in ppm) represent the elemental Cu concentration. All copper treatments were applied at 20, 40, 60, and 80 days after planting.
2.9. Yield and Morphological Parameters
Five plants from each plot were randomly chosen 135 days after transplanting for measuring the following morphological traits, according to the recommendations of the Ministry of Agriculture and Land Reclamation, Egypt:
- Plant height (cm), using a measuring tape.
- Number of leaves per plant.
- Fresh weight of leaves (g), using a sensitive balance.
- Number of obtained bulbs per treatment.
- Neck diameter (cm), using a caliper.
- Bulb diameter (cm), using a caliper.
- Bulbing ratio (the diameter of the neck divided by the bulb diameter).
- Bulb weight (g), using a sensitive balance.
- Fresh weight (leaves and bulbs), using a sensitive balance.
- Yield (ton ha−1).
2.10. Chemical Analysis
Plant material was dried in an electric oven at 70 °C for 24 h according to AOAC [20], then finely ground for chemical determination of elements. The wet digestion of 0.2 g plant material with sulfuric and perchloric acids was carried out by adding 5 mL concentrated sulfuric acid to the samples and the mixture was heated for 10 min, then 0.5 mL perchloric acid was added and heated until a clear solution was obtained [17,20]. Total nitrogen content of the dried leaves was determined using the modified micro-Kjeldahl method as described by AOAC [20]. Phosphorus was determined colorimetrically by using the chlorostannous molybdophosphoric blue color method in sulfuric acid according to Jackson [17]. Potassium concentration was determined using the flame photometer apparatus (CORNING M 410, Germany). Concentrations of Fe, Cu, Zn, and Mn in plant bulbs were determined using an atomic absorption spectrophotometer with air-acetylene and fuel (PyeUnicam, model SP-1900, USA). Total soluble solids (T.S.S), protein, and fibers were assayed according to AOAC [20].
Total carbohydrates in bulbs were determined by the phosphomolybdic acid method according to AOAC [20]. Two grams of sample was crushed with 10 mL 80% ethanol in mortar and pestle, then filtered through Whatman filter paper. The filter and residue were collected separately. The alcohol residue was taken into a 250 mL conical flask and 150 mL distilled water, and 5 mL concentrated HCL was added to it. The residue was left to hydrolyze for 30 min and then cooled to room temperature. Na2CO3 was then added slowly until the extract became neutral (pH = 7). The extract was filtered, and the residue was discarded. The filtrate was taken into a conical flask and condensed in a hot water bath for 3–4 min. Distilled water was added to the filtrate, then filtered, residue was discarded, and the volume of filtrate was used for estimating content of reducing sugars. Of this filtrate, 20 mL was taken into a 150 mL conical flask and 2 mL of concentrated HCl was added to it. It was then hydrolyzed for 30 min and cooled at room temperature. Na2CO3 was slowly added until the extract became neutral (pH = 7). This extract was filtered, residue was discarded, and the final volume of the filtrate was measured and used for estimating content of total sugars. For the aliquot sample, 0.5 mL was taken into a test tube and 1 mL of Somogyi reagent was added. Test tubes were placed in a boiling water bath for 30 min, cooled to room temperature, and 1 mL of arsenomolybdate reagent was added. The content was mixed and diluted to a volume of 10 mL and its absorbance was spectrophotometerically measured at 560 nm.
Vitamins were determined in bulbs and estimated per 100 mL fresh weight. Vitamins B1 (thiamin) and B2 (riboflavin) were determined fluorometrically, whereas vitamins B3 (niacin) and C (ascorbic acid) were determined colorimetrically (see [21] for detailed protocols). Analysis of vitamins B5 (pantothenic acid), B6 (pyridoxine), and B9 (folate) were performed using the HPLC method (see [19,22,23] for detailed protocols). With respect to amino acids determination, bulbs were dried, weighed, and chopped. The obtained mixture was subsequently freeze-dried and stored at 4 °C in sealed vials. The amino acid extraction from each freeze-dried sample was performed according to Wayne [24], with some modifications. Twenty milliliters of distilled water was added to 1.0 g of freeze-dried onion sample. The obtained suspension was maintained under magnetic stirring for 3 min at 0 °C in an ice bath, and then centrifuged at 10,000 rpm for 15 min. This operation was consecutively repeated three times by re-suspending the pellet every time. The final solution was filtered under vacuum through a 0.45 µm nylon filter. Each extracted solution was lyophilized again and stored at 4 °C in sealed vials. Each extract was analyzed using HPLC-ELSD methodology [25]. Samples were prepared at a concentration of 25 mg/mL, filtered through a nylon 0.45 µm filter, and analyzed in triplicate. The mobile phase gradient was obtained from eluent A (7 mM HFBA in pure water) and eluent B (net MeCN) as follows: 0–10 min 100% A, 10–30 min from 100 up to 75% A, 30–38 min from 75 up to 70% A, 38–39 min 100% A, 39–70 min 100% A.
2.11. Data Analysis
The obtained data were analyzed using ANOVA at the 5% significance level. The difference between means of treatments were analyzed using Duncan’s Test at the 5% significance level.
3. Results
3.1. Soil Properties after Planting
Copper deficiency caused a marked increase in soil Cu values after two seasons of onion plantation (Table 3). In addition, soil alkalinity exhibited lower values at the end of the experiments compared with initial values before the first season.
3.2. Vegetative Growth
The effects of different sources of Cu on vegetative growth parameters of onion plants are shown in Table 4. Plant height, number and fresh weight of leaves, bulbs, and total number of plants significantly (p ≤ 0.05) increased under all Cu treatments as compared to control, where the highest values were obtained under CuO NPs.
3.3. Yield
Results of yield and bulb quality are presented in Table 5. Significant (p ≤ 0.05) increases in bulb diameter, neck diameter, bulbing ratio, bulb weight, and total yield were detected under all Cu supplementation treatments. In particular, CuO NPs treatment produced the best results by boosting bulb diameter, bulb weight, and total yield of onion as compared to control treatment. These findings were consistent in both experimental seasons.
3.4. Nitrogen, Phosphorus, and Potassium Contents of Onion Bulbs
Results in Table 6 clearly show that foliar application of different sources of copper had a significant (p ≤ 0.05) effect on the chemical composition of onion bulb tissues in the two seasons of study. Once again, the highest results of N, P, and K concentration were obtained using copper NPs as compared to other treatments. As expected, Cu deficiency led to the lowest values of N, P, and K concentration in control plants.
3.5. Trace Elements Content of Onion Bulbs
Results in Table 7 show that foliar application of different sources of Cu had a significant (p ≤ 0.05) effect on the nutritional values of onion tissues in the two seasons of study. Generally, it could be noted that the maximum values of Fe, Zn, Cu, and Mn were observed by using foliar application of CuO NPs as compared to other treatments, while the lowest values were observed under control conditions.
3.6. Phytochemicals Content of Onion Bulbs
Results in Table 8 show the phytochemicals composition of onion treated by different sources of Cu. Exposure of onion plants to Cu deficiency markedly reduced the contents of carbohydrates, protein, dietary fibers, as well as the percentage of total soluble solids (TSS) of onion bulbs. The supplementation of Cu in the form of foliar spray had significant (p ≤ 0.05) impacts on these parameters and resulted in higher values compared to control, especially CuO NPs. Moreover, Cu deficiency caused a significant (p ≤ 0.05) increase in water content of bulbs, and the values were different for this parameter under different sources of Cu. The lowest value of water content was recorded under CuO NPs treatment.
3.7. Vitamin Contents of Onion Bulbs
Cu deficiency resulted in a marked (p ≤ 0.05) decrease in contents of vitamins of onion bulbs (Table 9). CuO NPs application resulted in more vitamin B3 compared to other Cu sources, which were inferior in this context. Nevertheless, the results of the rest of the studied vitamins were not consistent between the two seasons.
3.8. Amino Acid Contents of Onion Bulbs
Cu deficiency caused a notable decrease in amino acid contents of onion compared with all three Cu foliar treatments (Table 10). Exogenous application of Cu increased the contents of methionine, phenylalanine, leucine, tryptophan, and cysteine in onion, with the highest significant (p ≤ 0.05) increases attributed to CuO NPs treatment.
4. Discussion
This study investigated the response of onion plants grown in Cu-deficient soil to CuO NPs compared with CuSO4 and chelated Cu forms. The maximum recovery from Cu deficiency in the majority of the studied parameters was observed when onion plants were sprayed with 10 ppm CuO NPs.
4.1. Importance of Copper to Plants
Cu together with iron, zinc, boron, chlorine, manganese, and molybdenum are considered essential micronutrients that plants need to carry out many important physiological processes [6,26]. At the cellular level, Cu deficiency hinders electron transport in the thylakoid by affecting plastocyanin proteins [27]. In addition, Cu-deficient plants have a reduced cellular ability to scavenge reactive oxygen species (ROS) because Cu/Zn SOD enzymes become altered [28]. These damages affect plant growth and crop quality. Micronutrient deficiencies in vegetable crops have been widely studied and many strategies for increasing micronutrient availability in soils have been proposed [29,30,31]. One of the several causes for the appearance of Cu deficiency symptoms is cultivating crops in sandy soils of low organic matter content. This becomes exacerbated by additional environmental factors, including soil pH, calcium concentration, and the type of crop being cultivated [9]. Although total Cu content of most agricultural soils is sufficient, ranging 20–250 mg/kg soil, sandy-textured soils are more likely to be Cu-deficient than loams and clays, with Cu contents ranging 1–20 mg/kg soil [32]. The total Cu content in the soil used in this study (Table 1) was 12 ppm (mg/kg). This indicates a sub-optimal value for normal plant growth and development according to the above-mentioned established ranges. At the end of the experiments, the average of soil total Cu contents of the two seasons was 17 ppm (Table 3), a value also slightly below the optimal range. This indicates that the Cu levels used in this study were not toxic and would not contaminate the soil for future plantations. The soil used in this study was calcareous in nature with alkaline pH values, being 7.98 and 7.8 for before and after plantation, respectively (Table 1 and Table 3). It is possible that these conditions aggravated Cu deficiency in onion plants of this study, because Cu availability is reduced at a soil pH above 7.0 [33].
4.2. Cu Deficiency Negatively Affects Bulb Quality Parameters of Onion
Sensitivity to Cu deficiency differs widely between plant species. Alloway and Tills [34] reported that some field crops such as wheat are sensitive to Cu deficiency, while others like rapeseed are less sensitive. Regarding vegetable crops, the most sensitive species to Cu deficiency seem to be onions and carrots [35]. Generally, Cu deficiency symptoms in plants reflect the important functions that Cu carries out. Cu is a transition element that has an important role as an enzyme cofactor that catalyzes redox reactions. Additionally, Cu is involved in many pivotal metabolic functions (photosynthesis and respiration), protein synthesis, cell lignification, auxin regulation, and protection against oxidative stress [6]. In plants, Cu deficiency first appears in the younger leaves because Cu is an immobile nutrient. Fageria [9] reported that leaves become bluish-green and chlorotic near the tips, extending downwards at sides of the midrib. As deficiency continues, tips become necrotic with dark-brown color. In addition, new leaves appear as needle-shaped because they fail to unroll. In onion, leaf tips turn white and twisted or bend at right angles [8]. As Cu deficiency progresses, leaves may become chlorotic and die back. Regarding onion bulbs, they become pale with thin and brittle skins. This might be because of a decrease in expression of cell wall-related genes, as previously demonstrated in alfalfa [36]. It is worth noting that the root system is the first organ to encounter any soil abiotic stress, including nutrient deficiency, and its traits are affected to a higher degree compared to other plant organs [37,38]. Therefore, onion bulbs are usually greatly affected by soil abiotic stresses. Gupta [8] also reported that Cu deficiency in onion results in an early maturation of plants, manifested as smaller dried tops and bulbs lacking solidity and firmness with greenish bronze color. Our results of vegetative growth and yield seem to be in harmony with these findings (Table 4 and Table 5), where all bulb quality parameters were significantly affected by Cu deficiency.
4.3. CuO NPs Form Is the Most Efficient Treatment in Increasing Onion Yields
It is well-known that, up to a certain limit, plant biomass increases as photosynthetic rate increases. Cu deficiency affects both PS I and PS II activities in the chloroplast, which in turn reduce carbohydrate synthesis by almost 50% through affecting both chlorophyll content and leaf area [6]. These reductions were reported to be correlated with a reduction in plastocyanin levels in Cu-deficient leaves [39]. In addition, Cu-deficient plants show decreases in photosynthetic rates associated with perturbation in thylakoids, reduced stomatal conductance, and lower Rubisco activity and RuBP regeneration [40]. The reductions in bulb weight biomass detected in the control conditions of our study (Table 5) were almost 245%, as compared to plants sprayed with CuO NPs. This might indicate that Cu-deficient conditions produced maximal decreases in photosynthesis, which consequently decreased carbohydrate, fibers, and TSS % in affected bulbs (Table 8). As mentioned above, plants differ in their sensitivity to Cu deficiency and, thus, their response to Cu supplementation varies widely. Early greenhouse studies carried out by Gupta and MacLeod [41] reported responses to Cu supplementation and revealed differential yield increases of wheat, barley, and oats by 38%, 180%, and 500%, respectively. That was also the case in open field trials of wheat grown under Cu deficiency [42]. The CuO NPs form was the most efficient treatment to increase onion yields in our study. Increments in onion yield increased up to 72% when plants where sprayed with CuO NPs as compared to 9% and 35% in Cu sulfate and chelate, respectively (Table 5). This might indicate a greater effect of CuO NPs on the photosynthetic machinery as compared to sulfate and chelate forms. A recent study reported a 31% increase in total onion yield when plants were sprayed with 50 ppm of chelated Cu [14]. Using such high doses of Cu, however, is not sustainable to use in future plantations because this may cause toxicity to other crops of higher sensitivity to Cu. In addition, our study provided evidence that using a much lower dose of Cu (i.e., 10 ppm CuO NPs treatment) was greatly effective in achieving large yield increments compared with higher doses of Cu sulfate and chelates (i.e., 20 ppm).
4.4. CuO NPs Enhances the Content of Micro- and Macro-Elements in Onion Tissues
Although the morphological response of Cu-deficient onion plants has been described in the literature, little is known about its response regarding the accumulation of mineral and biochemical compounds in its tissues. It is clear from our results that onion plants were significantly affected by Cu deficiency. Cu is known to have several interactions with macro- and micro-elements that may consequently affect crop yields. Regarding macronutrients, nitrogen and/or phosphorus application were reported to aggravate Cu-deficiency in wheat [43,44]. In addition, foliar application of potassium was reported to reduce Cu content in pecan plants [45]. Furthermore, calcium was shown to reduce Cu uptake in wheat and lettuce [44,46]. Although CuO NPs showed a remarkable efficiency in increasing the accumulation of macro-elements in onion bulb tissues compared with Cu sulfate and chelates (Table 6), the reasons behind this remain to be elucidated. Concerning micronutrients, evidence supports that Cu uptake is also inhibited by application or presence of Zn [9]. Except for molybdenum, Cu requirement of plants is lower than the rest of the micronutrients. Generally, adequate Cu concentration in plant dry tissues is mostly between 2 and 20 mg kg−1 [33]. In onion, average Cu concentration in the whole plant ranges between 8 and 45 mg kg−1 in dry matter [47]. At higher levels of Cu supplementation, onion leaves and stems seem to accumulate more Cu than the bulbs [48]. Our results are in accordance with the aforementioned tissue Cu ranges (Table 7), where Cu supplementation in any of the three forms increased tissue Cu contents by almost 100%.
4.5. Cu Enhances Vitamin Contents in Onion Bulbs
A novel result in our study is the effect that Cu deficiency has on the contents of vitamins and amino acids of onion plants (Table 9 and Table 10, respectively). Onion has a rich variety of bioactive components that induce many beneficial effects on human health [2,4]. It is known that onion is a rich source of vitamins B6 (pyridoxine), C (ascorbic acid), and B9 (folate) [49]. These vitamins play important roles in plant metabolism and many molecular events linked to induced resistance [50]. These include, but are not limited to, acting as a co-enzyme in chemical reactions and induction of DNA and protein synthesis [3]. Apparently, these vitamins contained in onion seem to have a beneficial effect on treating or preventing hyperhomocyteinemia and protecting against oxidative damage in humans [51]. The most studied vitamin regarding its relation to Cu supplementation is ascorbic acid (vitamin C). The effect of Cu supplementation on plant tissue levels of ascorbic acid seems to be species-dependent. For example, in vegetable crops, levels of ascorbic acid in cabbage, carrots, cauliflower, eggplants, lettuce, and spinach increase with Cu supplementation [52]. On the other hand, Cu supplementation had no effect on the levels of ascorbic acid of mandarin and tomato plants, while its levels decreased in spleen amaranth plants [52]. Before starting the present study, it was expected that onion plants might respond to Cu supplementation and increase its vitamin C contents in the same manner as the above-mentioned Cu-responsive plants, such as spinach and carrots. Nevertheless, our results on onion indicated that, although levels of vitamin C increased under all three Cu sources used in this study, these increases were only detected as significant in the second season of field experiments (Table 9). These results are non-conclusive and, thus, further trials using additional Cu concentrations are needed to confirm the effect that Cu has on ascorbic acid contents in onion plants. Similar conclusions could be drawn on values of vitamin B1, B2, B5, B6, and B9 (Table 9), where their changes were inconsistent between the two seasons as well. Conversely, Cu supplementation had a significant effect on B3 (niacin) contents of onion plants in both seasons (Table 9). Little is known about vitamin B3 biosynthesis pathways in plants [53] and, therefore, the way that Cu causes such increases in vitamin B3 remains to be elucidated. In all cases, elevated vitamin B3 levels in onion under Cu supplementation might be beneficial for preventing and treating human diseases such as pellagra and high blood pressure [54].
4.6. Cu Enhances the Accumulation of Amino Acids in Onion Bulbs
In contrast to vitamins, Cu supplementation remarkably increased the content of amino acids in onion in both seasons. The effects of Cu deficiency and/or its supplementation on amino acid contents in plants are vastly understudied. Early reports, however, established that each amino acid responded differently to Cu deficiency in tomato [55]. The author reported that tomato plants subjected to Cu deficiency had increased levels of aspartate, glutamate, asparagine, arginine, β-alanine, valine, and proline. On the other hand, amino acid levels of histidine, lysine, serine, glycine, threonine, α-alanine, γ-aminobutyric acid, ethanolamine, and phenylalanine decreased in Cu-deficient tomato plants. The levels of the rest of the amino acids in the aforementioned study remained the same. Our results in onion show similar effects for phenylalanine, where it is severely decreased under Cu deficiency (Table 10). The rest of the amino acids in our study (methionine, leucine, tryptophan, and cysteine) were increased by Cu supplementation, reaching the highest levels under CuO NPs treatment. Cu is present in many plant amino acids. It may be bound with nitrogen in histidine in an oxidized state (Cu2+) or by sulfur in cysteine or methionine in a reduced state (Cu+) [27]. Cu also forms part of many important proteins such as plastocyanin, cytochrome c oxidase, Cu/Zn Super oxide dismutase, ethylene receptors, laccase, ascorbate oxidase, amine oxidase, phytocyanin, and polyphenol oxidase [56]. Consequently, any Cu deficiency would affect total protein content in plants. Our study might support this notion, as the contents of protein in Cu-deficient onion plants were significantly reduced (Table 8).
4.7. Is There a Clear Limit between the Stimulating and Inhibiting/Toxic Concentrations of Cu NPs in Plants?
One reason why CuO NPs might be more effective and efficient in supplementing Cu to plants is that nanoscale Cu has a higher surface per volume compared with sulfate and chelate forms. This gives CuO NPs new physical and chemical properties that might not be present in bulk form. For instance, compared with bulk materials, NPs have the ability of penetrating thick cell walls and exerting their functions inside seeds, resulting in improved germination rates in some plants [57]. Some NPs affect photosynthesis directly by interacting with carbonic anhydrase and Rubisco [58].
Nonetheless, are Cu NPs beneficial or toxic to plants? A considerable body of literature, including the present work, indicates that the application of Cu nanoparticles has many beneficial effects on certain plant species. Cu NPs significantly increase fruit quality and bioactive compounds of tomato under normal conditions [59] and under salinity stress [60]. Priming maize plants with Cu NPs increases photosynthetic pigments and total grain yield under drought stress [61]. Recent reviews by Mittal et al. [62] and Paramo et al. [63] documented several works that emphasize the beneficial role of Cu NPs in the agro-industry and in producing high-quality food for human nutrition. In these studies, Cu NPs play clear roles as elicitors of many bioactive compounds of high nutritional quality [64] in cucumber [65], ashwaghanda [66], basil [67], and stevia [68]. On the other hand, many reports document the inhibitory/toxic effects that Cu NPs have on some plants [11,69]. After all, we cannot ignore works reporting that levels as low as 2 mg L−1 of Cu NPs seem to greatly inhibit biomass production in certain plant species, such as Arabidopsis [3]. In fact, the use of Cu NPs as antifungal and antibacterial agents is proof of this toxicity at certain concentrations, at least to microorganisms [70,71]. Nevertheless, with few exceptions, most of the reviewed studies related to the toxicity of Cu NPs seem to have a toxicological focus, and very high doses of Cu NPs are used in them ([72], and references therein). It is thus appropriate to state with reasonable confidence that the stimulating/toxic effect of Cu NPs differs widely among plants. Our results show that, in the case of onion, low concentrations of CuO NPs (10 ppm) were able to produce very positive results on bulb yield and quality compared with Cu sulfate and chelates in sandy soils characterized by high alkalinity. Moreover, onion plants benefited from this low concentration of Cu, with very little impact on soil Cu concentration at the end of experiments. This makes Cu NPs a very safe option for supplementing Cu to onion plants without polluting the environment and soil ecosystem, making it an attractive resource for precision and sustainable agriculture [73,74]. Furthermore, the availability of several biological materials to biosynthesize Cu NPs, including plants, bacteria, and fungi [75], provides several options for its low-cost and environmentally safe production. In all cases, future experiments studying the effects of higher doses of Cu NPs on onion are needed.
Another reasonable question that begs an answer is what is the limit between the stimulating and inhibiting/toxic concentrations of Cu NPs in plants? Unfortunately, there is no easy and direct answer to that question because that limit is not universal in plants and depends on many factors. It might be tempting to answer that question by describing that the limit of stimulation/inhibition of Cu NPs in tomato, for example, seems to be between 250 and 500 mg L−1 [59]. Nevertheless, many factors other than NPs concentration seem to play a major role in defining that limit. For example, factors related to the NPs itself such as nanoparticle size and size distribution, morphology, agglomeration, crystal structure, or surface charge might be potential contributors to that limit [63]. Interaction of the NPs with other molecules is another factor that should be taken into consideration. For instance, using CTAB-assisted formulations of Cu NPs enhance their cytotoxic and genotoxic potential [76]. In addition, sensitivity to Cu seems to differ widely between plant species and genera, as mentioned before. Cu also has a very narrow optimum range of concentration in which it exerts its beneficial effects [6,8]. When Cu levels fall below or exceed these limits, deficiency or toxicity symptoms immediately show on these plants, respectively. Furthermore, different experimental conditions and methodologies introduce additional variation in the results and their subsequent interpretation. For example, it would be risky to draw conclusions from in vitro studies or seed nano-priming experiments and compare them with results from plants foliar-sprayed with NPs in field trials. Finally, a more reasonable and cautious conclusion regarding Cu NPs is that their beneficial or toxic effects depend on many factors, including the concentration and properties of Cu NPs, plant species being studied, and experimental conditions. In all cases, future screening of field, horticultural, medicinal, and ornamental crops in multi-local field studies are needed to determine the exact stimulation/inhibition limit of NPs of each of these crops. It is also recommended to establish a common framework outlining the guidelines related to these future studies.
5. Conclusions
Recovery from Cu deficiency symptoms can be achieved by using various Cu sources and forms. CuSO4 is known to be a cheap and common Cu form to achieve that goal. Nevertheless, the results presented in this study provide evidence to prove the hypothesis that low concentrations of CuO NPs not only are safe to apply to onion plants, but also lead to higher yields, antioxidants, and phytochemicals accumulation in onion compared with CuSO4 and chelated Cu forms. Using 10 ppm of CuO NPs also boosted yield of onion by mainly increasing the number of bulbs and bulb weight per ha. CuO NPs also maximized bulb contents of TSS, minerals, vitamins, and amino acids compared with CuSO4 and chelated Cu. CuO NPs were not only safe to use but showed similar or superior results to the other Cu forms evaluated in this study. Future studies on the effect of CuO NPs on other crops are needed to further elucidate their role in recovery from Cu deficiency.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agronomy11051006/s1, Figure S1: Image from TEM microscopy showing copper oxide nanoparticles.
Author Contributions
A.W.M.M. and A.Z.A.H. conceived the experiments and methodology; A.W.M.M., A.Z.A.H., S.A.M. and R.E.-B. performed the experiments, measurements, and chemical analyses; A.W.M.M., A.Z.A.H. and S.A.M. analyzed the data; S.A.M. and A.W.M.M. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO-United Nations Statistics Division. Statistics Division of Food and Agriculture Organization of the United Nations; FAO-United Nations Statistics Division: Rome, Italy, 2018. [Google Scholar]
- Teshika, J.D.; Zakariyyah, A.M.; Zaynab, T.; Zengin, G.; Rengasamy, K.R.; Pandian, S.K.; Fawzi, M.M. Traditional and modern uses of onion bulb (Allium cepa L.): A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 59, S39–S70. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, R.K. Nutraceutical, pharmaceutical and therapeutic uses of Allium cepa: A review. Int. J. Green Pharm. 2016, 10, 46–64. [Google Scholar]
- Marrelli, M.; Amodeo, V.; Statti, G.; Conforti, F. Biological properties and bioactive components of Allium cepa L.: Focus on potential benefits in the treatment of obesity and related comorbidities. Molecules 2019, 24, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khokhar, K.M. Mineral nutrient management for onion bulb crops—A review. J. Hortic. Sci. Biotechnol. 2019, 94, 703–717. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of Nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 191–248. [Google Scholar]
- Rahimi, A.; Bussler, W. Physiologische Voraussetzungen für die Bildung der Kupfermangelsymptome. Z. Pflanz. Bodenkd. 1973, 136, 25–32. [Google Scholar] [CrossRef]
- Gupta, U.C. Copper in Crop and Plant Nutrition. In Handbook of Copper Compounds and Applications; Marcel Dekker, Inc.: New York, NY, USA, 1997; pp. 203–229. [Google Scholar]
- Fageria, N.K. The Use of Nutrients in Crop Plants; CRC press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Purvis, E.; Carolus, R. Nutrient Deficiencies in Vegetable Crops. In Hunger Signs in Crops; MacKay & Company, Inc.: New York, USA, 1964; pp. 245–286. [Google Scholar]
- Kasana, R.C.; Panwar, N.R.; Kaul, R.K.; Kumar, P. Biosynthesis and effects of copper nanoparticles on plants. Environ. Chem. Lett. 2017, 15, 233–240. [Google Scholar] [CrossRef]
- Shah, V.; Belozerova, I. Influence of metal nanoparticles on the soil microbial community and germination of lettuce seeds. Water Air Soil Pollut. 2009, 197, 143–148. [Google Scholar] [CrossRef]
- Nair, P.M.G.; Chung, I.M. Impact of copper oxide nanoparticles exposure on Arabidopsis thaliana growth, root system development, root lignificaion, and molecular level changes. Environ. Sci. Pollut. Res. 2014, 21, 12709–12722. [Google Scholar] [CrossRef]
- Fouda, K.F. Response of onion yield and Its chemical content to NPK fertilization and foliar application of some micronutrients. Egypt. J. Soil Sci. 2016, 56, 549–561. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, C.; Cota-Ruiz, K.; Peralta-Videa, J.R.; Sun, Y.; Rawat, S.; Tan, W.; Reyes, A.; Hernandez-Viezcas, J.A.; Niu, G. Improvement of nutrient elements and allicin content in green onion (Allium fistulosum) plants exposed to CuO nanoparticles. Sci. Total Environ. 2020, 725, 138387. [Google Scholar] [CrossRef]
- Richards, L.A. Diagnosis and Improvement of Saline and Alkali Soils; Government Printing Office (Superintendent of Documents): Washington, DC, USA, 1954. [Google Scholar]
- Jackson, M. Soil Chemical Analysis Prentice; Hall of India Private Limited: New Delhi, India, 1967; p. 498. [Google Scholar]
- Keller, J.; Karmeli, D. Trickle Irrigation Design; Rain Bird Sprinkler Manufacturing Corporation: Glendora, CA, USA, 1975. [Google Scholar]
- Zhang, Q.; Zhang, K.; Xu, D.; Yang, G.; Huang, H.; Nie, F.; Liu, C.; Yang, S. CuO nanostructures: Synthesis, characterization, growth mechanisms, fundamental properties, and applications. Prog. Mater. Sci. 2014, 60, 208–337. [Google Scholar] [CrossRef]
- The Association of Official Analytical Chemists. Official Methods of Analysis: Changes in Official Methods of Analysis Made at the Annual Meeting, Supplement; Association of Official Analytical Chemists: Washington, DC, USA, 1990; Volume 15. [Google Scholar]
- Katoch, R. Analytical Techniques in Biochemistry and Molecular Biology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- de Brouwer, V.; Storozhenko, S.; Stove, C.P.; van Daele, J.; van der Straeten, D.; Lambert, W.E. Ultra-performance liquid chromatography–tandem mass spectrometry (UPLC–MS/MS) for the sensitive determination of folates in rice. J. Chromatogr. B 2010, 878, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Nazmul, H.; Akhtaruzzaman, M.; Zakir, S. Estimation of Vitamins B-Complex (B 2, B 3, B 5 and B 6) of Some Leafy Vegetables Indigenous to Bangladesh by HPLC Method. J. Anal. Sci. Methods Instrum. 2013, 3, 24–29. [Google Scholar]
- Wayne, W. A reliable methodology for quantitative extraction of fruit and vegetable physiological amino acids and their subsequent analysis with commonly available HPLC systems. Food Nutr. Sci. 2012, 3, 863–871. [Google Scholar]
- Sardella, R.; Lisanti, A.; Marinozzi, M.; Ianni, F.; Natalini, B.; Blanch, G.P.; del Castillo, M.L.R. Combined monodimensional chromatographic approaches to monitor the presence of D-amino acids in cheese. Food Control 2013, 34, 478–487. [Google Scholar] [CrossRef] [Green Version]
- Andresen, E.; Peiter, E.; Küpper, H. Trace metal metabolism in plants. J. Exp. Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef]
- Pilon, M.; Abdel-Ghany, S.E.; Cohu, C.M.; Gogolin, K.A.; Ye, H. Copper cofactor delivery in plant cells. Curr. Opin. Plant Biol. 2006, 9, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, H.; Pilon, M.; Shikanai, T. How do plants respond to copper deficiency? Plant Signal. Behav. 2008, 3, 231–232. [Google Scholar] [CrossRef] [Green Version]
- Masunaga, T.; Fong, J.D.M. Strategies for Increasing Micronutrient Availability in Soil for Plant Uptake. In Plant Micronutrient Use Efficiency; Elsevier: Amsterdam, The Netherlands, 2018; pp. 195–208. [Google Scholar]
- Mahmoud, A.W.M.; Abdeldaym, E.A.; Abdelaziz, S.M.; El-Sawy, M.B.; Mottaleb, S.A. Synergetic Effects of Zinc, Boron, Silicon, and Zeolite Nanoparticles on Confer Tolerance in Potato Plants Subjected to Salinity. Agronomy 2020, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Shambhavi, S.; Kumar, R.; Kumar, R.; Singh, M. Micronutrients Deficiency in Vegetable Crops and Their Management. Diseases of Fruits and Vegetable Crops: Recent Management Approaches; Apple Academic Press: Palm Bay, FL, USA, 2020; p. 382. [Google Scholar]
- Locascio, S. The relationship of copper availability and soil pH. Solutions 1978, 30–42. [Google Scholar]
- Mengel, K.; Kirkby, E.A.; Kosegarten, H.; Appel, T. Fertilizer Application. In Principles of Plant Nutrition; Springer: New York, NY, USA, 2001; pp. 337–396. [Google Scholar]
- Alloway, B.; Tills, A.R. Copper deficiency in world crops. Outlook Agric. 1984, 13, 32–42. [Google Scholar] [CrossRef]
- Price, G. Australian Soil Fertility Manual; CSIRO Publishing: Clayton South, VIC, Australia, 2006. [Google Scholar]
- Printz, B.; Guerriero, G.; Sergeant, K.; Audinot, J.-N.; Guignard, C.; Renaut, J.; Lutts, S.; Hausman, J.-F. Combining-omics to unravel the impact of copper nutrition on alfalfa (Medicago sativa) stem metabolism. Plant Cell Physiol. 2016, 57, 407–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darwish, E.; Mottaleb, S.A.; Omara, M.; Safwat, G. Effect of salt stress on root plasticity and expression of ion transporter genes in tomato plants. Int. J. Bot. Res. 2016, 6, 13–26. [Google Scholar]
- Mottaleb, S.A.; Darwish, E.; Mostafa, M.; Safwat, G. Phenotyping Root System Architecture of Cotton (Gossypium barbadense L.) Grown Under Salinity. Agriculture (Pol’nohospodárstvo) 2017, 63, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Shahbaz, M.; Ravet, K.; Peers, G.; Pilon, M. Prioritization of copper for the use in photosynthetic electron transport in developing leaves of hybrid poplar. Front. Plant. Sci. 2015, 6, 407. [Google Scholar] [CrossRef] [Green Version]
- Morales, F.; Pavlovič, A.; Abadía, A.; Abadía, J. Photosynthesis in Poor Nutrient Soils, in Compacted Soils, and Under Drought. In The Leaf: A Platform for Performing Photosynthesis; Springer: New York, NY, USA, 2018; pp. 371–399. [Google Scholar]
- Gupta, U.C.; Macleod, L. Response to copper and optimum levels in wheat, barley and oats under greenhouse and field conditions. Can. J. Soil Sci. 1970, 50, 373–378. [Google Scholar] [CrossRef]
- Brennan, R. Effectiveness of some copper compounds applied as foliar sprays in alleviating copper deficiency of wheat grown on copper-deficient soils of Western Australia. Aust. J. Exp. Agric. 1990, 30, 687–691. [Google Scholar] [CrossRef]
- Fleming, G.A.; Delaney, J. Copper and nitrogen in the nutrition of wheat on cutaway peat. Ir. J. Agric. Res. 1961, 1, 81–82. [Google Scholar]
- Brown, J.C. Calcium Movement in Barley and Wheat as Affected by Copper and Phosphorus 1. Agron. J. 1965, 57, 617–621. [Google Scholar] [CrossRef]
- Mengel, K.; Barker, A.; Pilbeam, D. Handbook of Plant Nutrition; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Cheng, T.; Allen, H.E. Prediction of uptake of copper from solution by lettuce (Lactuca sativa Romance). Environ. Toxicol. Chem. Int. J. 2001, 20, 2544–2551. [Google Scholar] [CrossRef]
- Ginocchio, R.; Rodríguez, P.H.; Badilla-Ohlbaum, R.; Allen, H.E.; Lagos, G.E. Effect of soil copper content and pH on copper uptake of selected vegetables grown under controlled conditions. Environ. Toxicol. Chem. Int. J. 2002, 21, 1736–1744. [Google Scholar] [CrossRef]
- Badilla-Ohlbaum, R.; Ginocchio, R.; Rodríguez, P.H.; Céspedes, A.; González, S.; Allen, H.E.; Lagos, G.E. Relationship between soil copper content and copper content of selected crop plants in central Chile. Environ. Toxicol. Chem. Int. J. 2001, 20, 2749–2757. [Google Scholar] [CrossRef]
- Kwiatkowska, C.A.; Finglas, P.M.; Faulks, R.M. The vitamin content of retail vegetables in the UK. J. Hum. Nutr. Diet. 1989, 2, 159–172. [Google Scholar] [CrossRef]
- Boubakri, H.; Gargouri, M.; Mliki, A.; Brini, F.; Chong, J.; Jbara, M. Vitamins for enhancing plant resistance. Planta 2016, 244, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Bahram-Parvar, M.; Lim, L.T. Fresh-cut onion: A review on processing, health benefits, and shelf-life. Compr. Rev. Food Sci. Food Saf. 2018, 17, 290–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozafar, A. Plant. Vitamins; CRC press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Roje, S. Vitamin B biosynthesis in plants. Phytochemistry 2007, 68, 1904–1921. [Google Scholar] [CrossRef]
- Wan, P.; Moat, S.; Anstey, A. Pellagra: A review with emphasis on photosensitivity. Br. J. Dermatol. 2011, 164, 1188–1200. [Google Scholar] [CrossRef]
- Possingham, J. The effect of mineral nutrition on the content of free amino acids and amides in tomato plants IA comparison of the effects of deficiencies of copper, zinc, manganese, iron, and molybdenum. Aust. J. Biol. Sci. 1956, 9, 539–551. [Google Scholar] [CrossRef]
- Burkhead, J.L.; Gogolin Reynolds, K.A.; Abdel-Ghany, S.E.; Cohu, C.M.; Pilon, M. Copper homeostasis. New Phytol. 2009, 182, 799–816. [Google Scholar] [CrossRef]
- Pourkhaloee, A.; Haghighi, M.; Saharkhiz, M.J.; Jouzi, H.; Doroodmand, M.M. Carbon nanotubes can promote seed germination via seed coat penetration. Seed Technol. 2011, 33, 155–169. [Google Scholar]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Faisal, M.; Al Sahli, A.A. Nano-silicon dioxide mitigates the adverse effects of salt stress on Cucurbita pepo L. Environ. Toxicol. Chem. 2014, 33, 2429–2437. [Google Scholar] [CrossRef] [PubMed]
- López-Vargas, E.R.; Ortega-Ortíz, H.; Cadenas-Pliego, G.; de Alba Romenus, K.; Cabrera de la Fuente, M.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Foliar application of copper nanoparticles increases the fruit quality and the content of bioactive compounds in tomatoes. Appl. Sci. 2018, 8, 1020. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Hernández, H.; González-Morales, S.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Juárez-Maldonado, A. Effects of chitosan–PVA and Cu nanoparticles on the growth and antioxidant capacity of tomato under saline stress. Molecules 2018, 23, 178. [Google Scholar] [CrossRef] [Green Version]
- van Nguyen, D.; Nguyen, H.M.; Le, N.T.; Nguyen, K.H.; Nguyen, H.T.; Le, H.M.; Nguyen, A.T.; Dinh, N.T.T.; Hoang, S.A.; van Ha, C. Copper nanoparticle application enhances plant growth and grain yield in maize under drought stress conditions. J. Plant. Growth Regul. 2021, 2021, 1–12. [Google Scholar]
- Mittal, D.; Kaur, G.; Singh, P.; Yadav, K.; Ali, S. Nanoparticle-based sustainable agriculture and food science: Recent advances and future outlook. Front. Nanotechnol. 2020, 2020, 2. [Google Scholar]
- Paramo, L.A.; Feregrino-Pérez, A.A.; Guevara, R.; Mendoza, S.; Esquivel, K. Nanoparticles in Agroindustry: Applications, Toxicity, Challenges, and Trends. Nanomaterials 2020, 10, 1654. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Montejo, S.d.J.; Vargas-Hernandez, M.; Torres-Pacheco, I. Nanoparticles as Novel Elicitors to Improve Bioactive Compounds in Plants. Agriculture 2021, 11, 134. [Google Scholar] [CrossRef]
- Zhao, L.; Hu, J.; Huang, Y.; Wang, H.; Adeleye, A.; Ortiz, C.; Keller, A.A. 1H NMR and GC–MS based metabolomics reveal nano-Cu altered cucumber (Cucumis sativus) fruit nutritional supply. Plant. Physiol. Biochem. 2017, 110, 138–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, O.S.; Pant, N.C.; Laishram, L.; Tewari, M.; Dhoundiyal, R.; Joshi, K.; Pandey, C. Effect of CuO nanoparticles on polyphenols content and antioxidant activity in Ashwagandha (Withania somnifera L. Dunal). J. Pharmacogn. Phytochem. 2018, 7, 3433–3439. [Google Scholar]
- Abbasifar, A.; Shahrabadi, F.; ValizadehKaji, B. Effects of green synthesized zinc and copper nano-fertilizers on the morphological and biochemical attributes of basil plant. J. Plant. Nutr. 2020, 43, 1104–1118. [Google Scholar] [CrossRef]
- Ahmad, M.A.; Javed, R.; Adeel, M.; Rizwan, M.; Ao, Q.; Yang, Y. Engineered ZnO and CuO nanoparticles ameliorate morphological and biochemical response in tissue culture regenerants of candyleaf (Stevia rebaudiana). Molecules 2020, 25, 1356. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Cao, W.; Rui, Y. Interactions between nanoparticles and plants: Phytotoxicity and defense mechanisms. J. Plant. Interact. 2017, 12, 158–169. [Google Scholar] [CrossRef]
- Rai, M.; Ingle, A.P.; Pandit, R.; Paralikar, P.; Shende, S.; Gupta, I.; Biswas, J.K.; da Silva, S.S. Copper and copper nanoparticles: Role in management of insect-pests and pathogenic microbes. Nanotechnol. Rev. 2018, 7, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Hasan, M.; Ahammed, G.J.; Li, M.; Yin, H.; Zhou, J. Applications of nanotechnology in plant growth and crop protection: A review. Molecules 2019, 24, 2558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajput, V.; Minkina, T.; Suskova, S.; Mandzhieva, S.; Tsitsuashvili, V.; Chapligin, V.; Fedorenko, A. Effects of copper na-noparticles (CuO NPs) on crop plants: A mini review. BioNanoScience 2018, 8, 36–42. [Google Scholar] [CrossRef]
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The new perspective in precision agriculture. Biotechnol. Rep. 2017, 15, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Sharma, A.; Bhardwaj, S.K.; Arya, S.K.; Bhardwaj, N.; Khatri, M. Recent advances in the applications of nano-agrochemicals for sustainable agricultural development. Environ. Sci. Process. Impacts 2021, 23, 213–239. [Google Scholar] [CrossRef]
- Rasool, A.; Shah, W.H.; Tahir, I.; Rehman, R.U. Application of Nanoparticles in Crop Production and Protection. In Plant Nanobionics; Springer: New York, NY, USA, 2019; pp. 235–253. [Google Scholar]
- Singh, Z.; Singh, I. CTAB surfactant assisted and high pH nano-formulations of CuO nanoparticles pose greater cytotoxic and genotoxic effects. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
Table 1.
Some physical and chemical properties of experimental soil before planting.
| Physical Properties | Value | Chemical Properties | Value | ||
|---|---|---|---|---|---|
| Particle Size Distribution (%) | Electrical conductivity (EC ds/m) | 1.70 | |||
| Coarse sand (2000-200µ) | 79.20 | pH (1:2.5) soil:water suspension | 7.98 | ||
| Fine sand (200-20 µ) | 13.80 | Soluble cations (meq/L) | |||
| Silt (20-2 µ) | 4.75 | Ca2+ | 5.30 | ||
| Clay (< 2 µ) | 2.25 | Mg2+ | 4.32 | ||
| Bulk density (g/cm3) | 1.56 | K+ | 2.39 | ||
| Total porosity (%) | 54.70 | Na+ | 5.40 | ||
| Pore Size Distribution as % of Total Porosity | Soluble Anions (meq/L) | ||||
| Macro (drainable)-pores (>28.8 µ) | 80.84 | CO3 2- | Nil | ||
| Micropores (<28.8 µ) | 19.16 | HCO3- | 1.65 | ||
| Water holding capacity | 21.56 | Cl- | 3.65 | ||
| Field capacity | 8.68 | SO42- | 12.11 | ||
| Wilting percentage | 4.15 | Total carbonate (%) | 0.20 | ||
| Available moisture | 4.53 | Organic matter (%) | 0.26 | ||
| Hydraulic conductivity (cm/h) | 6.30 | Micronutrients (ppm) | |||
| Cu | Zn | Mn | Fe | ||
| 12.05 | 15.63 | 14.61 | 42.26 | ||
Table 2.
Chemical properties of applied compost.
| Property | Value |
|---|---|
| Moisture content (%) | 25 |
| PH (1:5) | 7.5 |
| EC (1:5 extract) dsm−1 | 3.1 |
| Organic C (%) | 33.11 |
| Organic matter (%) | 70 |
| Total N (%) | 1.82 |
| Total K (%) | 1.25 |
| C/N ratio | 15:1 |
| Total P (%) | 1.29 |
| Fe (ppm) | 97.5 |
| Mn (ppm) | 18.40 |
| Cu (ppm) | 5.25 |
| Zn (ppm) | 14.33 |
| Total content of bacteria (CFU g−1) | 2.5 × 107 |
| Phosphate dissolving bacteria (CFU g−1) | 2.5 × 106 |
| Weed seeds | 0 |
Table 3.
Physical and chemical properties of experimental soil after planting.
| Physical Properties | Value | Chemical Properties | Value | ||
|---|---|---|---|---|---|
| Particle Size Distribution (%) | Electrical conductivity (EC ds/m) | 1.64 | |||
| Coarse sand (2000–200µ) | 81.30 | pH (1:2.5) soil:water suspension | 7.80 | ||
| Fine sand (200–20 µ) | 12.40 | Soluble cations (meq/L) | |||
| Silt (20–2 µ) | 4.30 | Ca2+ | 5.13 | ||
| Clay (<2 µ) | 2.00 | Mg2+ | 4.22 | ||
| Bulk density (g/cm3) | 1.60 | K+ | 1.93 | ||
| Total porosity (%) | 53.78 | Na+ | 5.24 | ||
| Pore Size Distribution as % of Total Porosity | Soluble Anions (meq/L) | ||||
| Macro (drainable)-pores (>28.8 µ) | 82.54 | CO3 2- | Nil | ||
| Micropores (<28.8 µ) | 17.46 | HCO3- | 1.60 | ||
| Water holding capacity | 13.07 | Cl- | 3.44 | ||
| Field capacity | 8.46 | SO42- | 11.46 | ||
| Wilting percentage | 4.61 | Total carbonates (%) | 0.19 | ||
| Available moisture | 3.85 | Organic matter (%) | 0.22 | ||
| Hydraulic conductivity (cm/h) | 6.12 | Micronutrients (ppm) | |||
| Cu | Zn | Mn | Fe | ||
| 17.65 | 12.71 | 9.58 | 37.11 | ||
Table 4.
Effects of different sources of Cu on vegetative growth parameters of onion in two seasons (2018/2019 and 2019/2020).
Table 4.
Effects of different sources of Cu on vegetative growth parameters of onion in two seasons (2018/2019 and 2019/2020).
| Season 1 (2018–2019) | Season 2 (2019–2020) | |||||||
|---|---|---|---|---|---|---|---|---|
| Plant Height (cm) | No. of Leaves/Plant | Fresh Weight of Leaves (g) | No. of Bulbs | Plant Height (cm) | No. of Leaves/Plant | Fresh Weight of Leaves (g) | No. of Bulbs | |
| Control | 50.2 ± 0.32 b | 9.20 ± 0.25 a | 30.2 ± 0.12 b | 103 ± 2.5 c | 51.5 ± 1.5 b | 9.59 ± 1.3 a | 32.4 ± 1.2 b | 107 ± 2.1 c |
| CuSO4 | 52.1 ± 0.46 a | 9.60 ± 0.13 a | 32.5 ± 0.22 a | 124 ± 3.3 b | 52.8 ± 0.9 b | 9.73 ± 0.8 a | 33.6 ± 1.5 b | 125 ± 1.5 b |
| Cu-EDTA | 53.0 ± 1.02 a | 9.85 ± 0.57 a | 33.7 ± 0.15 a | 125 ± 2.4 b | 53.5 ± 0.7 b | 9.80 ± 1.1 a | 33.9 ± 1.1 b | 126 ± 2.1 b |
| Cu-NPs | 55.2 ± 1.32 a | 10.20 ± 0.41 a | 38.9 ± 0.36 a | 130 ± 1.9 a | 55.7 ± 0.6 a | 10.15 ± 2.3 a | 37.8 ± 1.6 a | 132 ± 1.3 a |
Each value represents the mean ± standard error. Values with different letters show significant differences at p ≤ 0.05 (Duncan’s Test).
Table 5.
Effects of different sources of Cu on the yield and bulb quality of onion in two seasons (2018/2019 and 2019/2020).
Table 5.
Effects of different sources of Cu on the yield and bulb quality of onion in two seasons (2018/2019 and 2019/2020).
| Season 1 (2018–2019) | Season 2 (2019–2020) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Bulb Diameter (cm) | Neck Diameter (cm) | Bulbing Ratio | Bulb Weight (g) | Total Yield (ton/ha) | Bulb Diameter (cm) | Neck Diameter (cm) | Bulbing Ratio | Bulb Weight (g) | Total Yield (ton/ha) | |
| Control | 5.70 ± 0.04 b | 1.56 ± 0.02 b | 0.27 ± 0.0001 c | 85.3 ± 4 c | 18.6 ± 0.3 c | 5.93 ± 0.03 b | 1.58 ± 0.03 c | 0.27 ± 0.0001 b | 87.9 ± 7 c | 19.0 ± 0.4 c |
| CuSO4 | 5.86 ± 0.06 b | 1.63 ± 0.04 b | 0.28 ± 0.0001 b | 90.5 ± 6 c | 20.2 ± 0.9 c | 5.91 ± 0.04 b | 1.85 ± 0.02 b | 0.31 ± 0.0002 a | 96.4 ± 11 c | 26.4 ± 1.6 b |
| Cu-EDTA | 5.96 ± 0.04 b | 1.74 ± 0.05 b | 0.29 ± 0.0001 b | 130.2 ± 8 b | 25.2 ± 0.2 b | 6.03 ± 0.02 a | 1.86 ± 0.01 b | 0.31 ± 0.0001 a | 135.5 ± 7 b | 30.0 ± 0.9 b |
| Cu-NPs | 6.55 ± 0.07 a | 2.04 ± 0.02 a | 0.31 ± 0.0001 a | 156.1 ± 10 a | 32.0 ± 0.3 a | 6.93 ± 0.03 a | 2.19 ± 0.06 a | 0.32 ± 0.0002 a | 158.7 ± 9 a | 33.6 ± 0.5 a |
Each value represents the mean ± standard error. Values with different letters show significant differences at p ≤ 0.05 (Duncan’s Test).
Table 6.
Effects of different sources of Cu on N, P, and K % of onion in two seasons (2018/2019 and 2019/2020).
Table 6.
Effects of different sources of Cu on N, P, and K % of onion in two seasons (2018/2019 and 2019/2020).
| Season 1 (2018–2019) | Season 2 (2019–2020) | |||||
|---|---|---|---|---|---|---|
| N % | P % | K % | N % | P % | K % | |
| Control | 1.70 ± 0.02 c | 0.14 ± 0.005 c | 1.35 ± 0.01 c | 1.79 ± 0.05 b | 0.15 ± 0.003 c | 1.25 ± 0.02 c |
| CuSO4 | 1.77 ± 0.01 b | 0.18 ± 0.002 b | 1.42 ± 0.03 b | 1.90 ± 0.02 b | 0.19 ± 0.001 b | 1.32 ± 0.01 b |
| Cu-EDTA | 1.75 ± 0.02 b | 0.21 ± 0.004 a | 1.47 ± 0.02 b | 2.08 ± 0.01 a | 0.22 ± 0.001 b | 1.39 ± 0.02 b |
| Cu-NPs | 1.92 ± 0.03 a | 0.22 ± 0.006 a | 1.61 ± 0.01 a | 2.30 ± 0.03 a | 0.25 ± 0.002 a | 1.46 ± 0.01 a |
Each value represents the mean ± standard error. Values with different letters show significant differences at p ≤ 0.05 (Duncan’s Test).
Table 7.
Effect of different sources of Cu on trace elements contents of onion in two seasons (2018/2019 and 2019/2020).
Table 7.
Effect of different sources of Cu on trace elements contents of onion in two seasons (2018/2019 and 2019/2020).
| Season 1 (2018–2019) | Season 2 (2019–2020) | |||||||
|---|---|---|---|---|---|---|---|---|
| Fe (ppm) | Cu (ppm) | Zn (ppm) | Mn (ppm) | Fe (ppm) | Cu (ppm) | Zn (ppm) | Mn (ppm) | |
| Control | 143.7 ± 2 c | 7.50 ± 0.12 b | 22.9 ± 0.1 c | 19.9 ± 0.2 b | 137.6 ± 2 c | 7.98 ± 0.21 c | 23.9 ± 0.3 b | 20.4 ± 0.5 b |
| CuSO4 | 150.7 ± 1 b | 7.92 ± 0.25 b | 24.3 ± 0.2 b | 21.7 ± 0.1 a | 154.8 ± 3 b | 8.60 ± 0.25 b | 25.3 ± 0.1 a | 22.8 ± 0.2 a |
| Cu-EDTA | 157.1 ± 2 a | 8.56 ± 0.32 a | 24.9 ± 0.3 b | 23.0 ± 0.1 a | 166.3 ± 3 a | 8.80 ± 015 b | 26.0 ± 0.3 a | 23.1 ± 0.2 a |
| Cu-NPs | 159.6 ± 3 a | 8.90 ± 0.28 a | 25.9 ± 0.2 a | 23.4 ± 0.2 a | 169.1 ± 4 a | 9.39 ± 0.19 a | 26.8 ± 0.5 a | 23.8 ± 0.4 a |
Each value represents the mean ± standard error. Values with different letters show significant differences at p ≤ 0.05 (Duncan’s Test).
Table 8.
Effect of different sources of Cu on phytochemical contents of onion in two seasons (2018/2019 and 2019/2020).
Table 8.
Effect of different sources of Cu on phytochemical contents of onion in two seasons (2018/2019 and 2019/2020).
| Season 1 (2018–2019) | Season 2 (2019–2020) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TSS (%) | Carbohydrates (mg/g) | Protein (mg/g) | Dietary fibers (mg/g) | Water (mg/g) | TSS (%) | Carbohydrates (mg/g) | Protein (mg/g) | Dietary fibers (mg/g) | Water (mg/g) | |
| Control | 7.02 ± 0.1 c | 92.0 ± 0.2 c | 9.0 ± 0.2 b | 16.0 ± 0.2 b | 873.5 ± 3 a | 6.03 ± 0.3 b | 93.3 ± 0.4 b | 8.9 ± 0.02 c | 15.3 ± 0.3 c | 853.5 ± 9 a |
| CuSO4 | 7.23 ± 0.1 b | 93.5 ± 0.1 b | 10.3 ± 0.1 a | 16.4 ± 0.0 a | 862.3 ± 5 a | 6.50 ± 0.2 a b | 95.8 ± 0.9 b | 11.3 ± 0.00 b | 16.1 ± 0.2 b | 834.3 ± 5 b |
| Cu-EDTA | 7.63 ± 0.4 b | 94.3 ± 0.3 b | 10.5 ± 0.3 a | 16.8 ± 0.1 a | 857.8 ± 6 a | 6.67 ± 0.3 a | 101.3 ± 0.5 a | 11.5 ± 0.00 a | 16.5 ± 0.3 b | 816.6 ± 3 b |
| Cu-NPs | 8.23 ± 0.1 a | 95.3 ± 0.1 a | 11.1 ± 0.6 a | 17.1 ± 0.1 a | 820.0 ± 3 b | 6.75 ± 0.4 a | 102.8 ± 0.7 a | 11.6 ± 0.01 a | 17.0 ± 0.1 a | 800.0 ± 2 c |
Each value represents the mean ± standard error. Values with different letters show significant differences at p ≤ 0.05 (Duncan’s Test).
Table 9.
Effect of different sources of Cu on vitamin contents of onion in two seasons (2018/2019 and 2019/2020).
Table 9.
Effect of different sources of Cu on vitamin contents of onion in two seasons (2018/2019 and 2019/2020).
| Season 1 (2018–2019) | Season 2 (2019–2020) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B1 (mg) | B2 (mg) | B3 (mg) | B5 (mg) | B6 (mg) | B9 (µg) | C (mg) | B1 (mg) | B2 (mg) | B3 (mg) | B5 (mg) | B6 (mg) | B9 (µg) | C (mg) | |
| Control | 0.039(2) c | 0.023(1) c | 0.113(1) c | 0.129(2) a | 0.14 ± 0.02 a | 20 ± 0.1 a | 7.0 ± 0.2 a | 0.043(1) a | 0.030(1) a | 0.111(2) b | 0.124(6) a | 0.12 ± 0.01 b | 17 ± 0.2 b | 6.7 ± 0.2 b |
| CuSO4 | 0.041(1) b | 0.025(2) b | 0.115(3) b | 0.123(4) c | 0.11 ± 0.01 b | 19 ± 0.2 b | 7.2 ± 0.4 a | 0.040(1) b | 0.024(2) b | 0.110(2) b | 0.120(5) b | 0.16 ± 0.01 a | 15 ± 0.1 b | 7.9 ± 0.1 a |
| Cu-EDTA | 0.043(3) b | 0.026(2) b | 0.117(2) b | 0.125(3) b | 0.12 ± 0.01 b | 18 ± 0.3 b | 7.4 ± 0.2 a | 0.044(2) a | 0.020(4) c | 0.118(4) a | 0.125(6) a | 0.11 ± 0.02 b | 18 ± 0.2 a | 7.6 ± 0.3 a |
| Cu-NPs | 0.048(2) a | 0.029(1) a | 0.120(1) a | 0.120(2) c | 0.14 ± 0.03 a | 22 ± 0.3 a | 7.8 ± 0.6 a | 0.045(1) a | 0.031(7) a | 0.117(1) a | 0.121(4) b | 0.15 ± 0.01 a | 20 ± 0.5 a | 7.9 ± 0.2 a |
Each value represents the mean ± standard error. Values with different letters show significant differences at p ≤ 0.05 (Duncan’s Test)
Table 10.
Effect of different sources of Cu on amino acid contents of onion in two seasons (2018/2019 and 2019/2020).
Table 10.
Effect of different sources of Cu on amino acid contents of onion in two seasons (2018/2019 and 2019/2020).
| Season 1 (2018–2019) | Season 2 (2019–2020) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Methionine (µg/mL) | Phenylalanine (µg/mL) | Leucine (µg/mL) | Tryptophan (µg/mL) | Cysteine( µg/mL) | Methionine (µg/mL) | Phenylalanine (µg/mL) | Leucine (µg/mL) | Tryptophan (µg/mL) | Cysteine (µg/mL) | |
| Control | 40.1 ± 0.3 c | 3.1 ± 0.2 c | 31.2 ± 0.3 c | 25.4 ± 0.6 b | 48.6 ± 0.1 c | 39.9 ± 0.2 c | 3.2 ± 0.7 d | 33.2 ± 0.3 b | 29.7 ± 0.5 a | 46.7 ± 0.5 c |
| CuSO4 | 44.3 ± 0.5 b | 11.2 ± 0.4 c | 33.4 ± 0.2 c | 20.1 ± 0.5 c | 50.0 ± 0.2 b | 42.8 ± 0.1 b | 11.2 ± 0.2 c | 34.5 ± 0.4 b | 22.4 ± 0.2 c | 49.2 ± 0.4 b |
| Cu-EDTA | 43.1 ± 0.9 b | 13.3 ± 0.1 b | 36.1 ± 0.6 b | 24.2 ± 0.2 b | 50.0 ± 0.5 b | 45.1 ± 0.6 a | 13.6 ± 0.4 b | 33.9 ± 0.7 b | 26.4 ± 03 b | 49.3 ± 0.2 b |
| Cu-NPs | 48.2 ± 0.6 a | 15.9 ± 0.6 a | 40.1 ± 0.8 a | 28.7 ± 0.4 a | 55.3 ± 0.3 a | 46.9 ± 0.5 a | 16.1 ± 0.2 a | 41.2 ± 0.2 a | 29.1 ± 0.4 a | 56.0 ± 0.3 a |
Each value represents the mean ± standard error. Values with different letters show significant differences at p ≤ 0.05 (Duncan’s Test).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mottaleb, S.A.; Hassan, A.Z.A.; El-Bahbohy, R.; Mahmoud, A.W.M. Are Copper Nanoparticles Toxic to All Plants? A Case Study on Onion (Allium cepa L.). Agronomy 2021, 11, 1006. https://doi.org/10.3390/agronomy11051006
AMA Style
Mottaleb SA, Hassan AZA, El-Bahbohy R, Mahmoud AWM. Are Copper Nanoparticles Toxic to All Plants? A Case Study on Onion (Allium cepa L.). Agronomy. 2021; 11(5):1006. https://doi.org/10.3390/agronomy11051006
Chicago/Turabian StyleMottaleb, Shady Abdel, Ahmed Z. A. Hassan, Reham El-Bahbohy, and Abdel Wahab M. Mahmoud. 2021. "Are Copper Nanoparticles Toxic to All Plants? A Case Study on Onion (Allium cepa L.)" Agronomy 11, no. 5: 1006. https://doi.org/10.3390/agronomy11051006
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.