
Quantitative Determination of the Effects of He–Ne Laser Irradiation on Seed Thermodynamics, Germination Attributes and Metabolites of Safflower (Carthamus tinctorius L.) in Relation with the Activities of Germination Enzymes

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Measurement of Seed Thermodynamic Parameters
2.2. Estimation of Seed Germination Attributes
2.2.1. Germination Percentage
2.2.2. Days to 50% Germination
2.2.3. Mean Emergence Time
2.2.4. Coefficient of Uniformity of Emergence (CUE)
2.2.5. Emergence Index
2.2.6. Germination Energy
2.3. Activities of Seed Germination Enzymes
2.3.1. Estimation of Amylase Activity
2.3.2. Estimation of Protease Activity
2.3.3. Estimation of Glucosidase Activity
2.4. Estimation of the Levels of Different Seed Metabolites during Germination at Different Time Intervals
2.4.1. Estimation of Chl Content of Germinating Seeds
2.4.2. Estimation of Carotenoid Content of Germinating Seeds
2.4.3. Estimation of TSS and RS Contents of Germinating Seeds
2.4.4. Estimation of TSP and FAA Contents of Germinating Seeds
2.4.5. Estimation of Oil and FFA Contents of Germinating Seeds
2.5. Statistical Analysis
3. Results
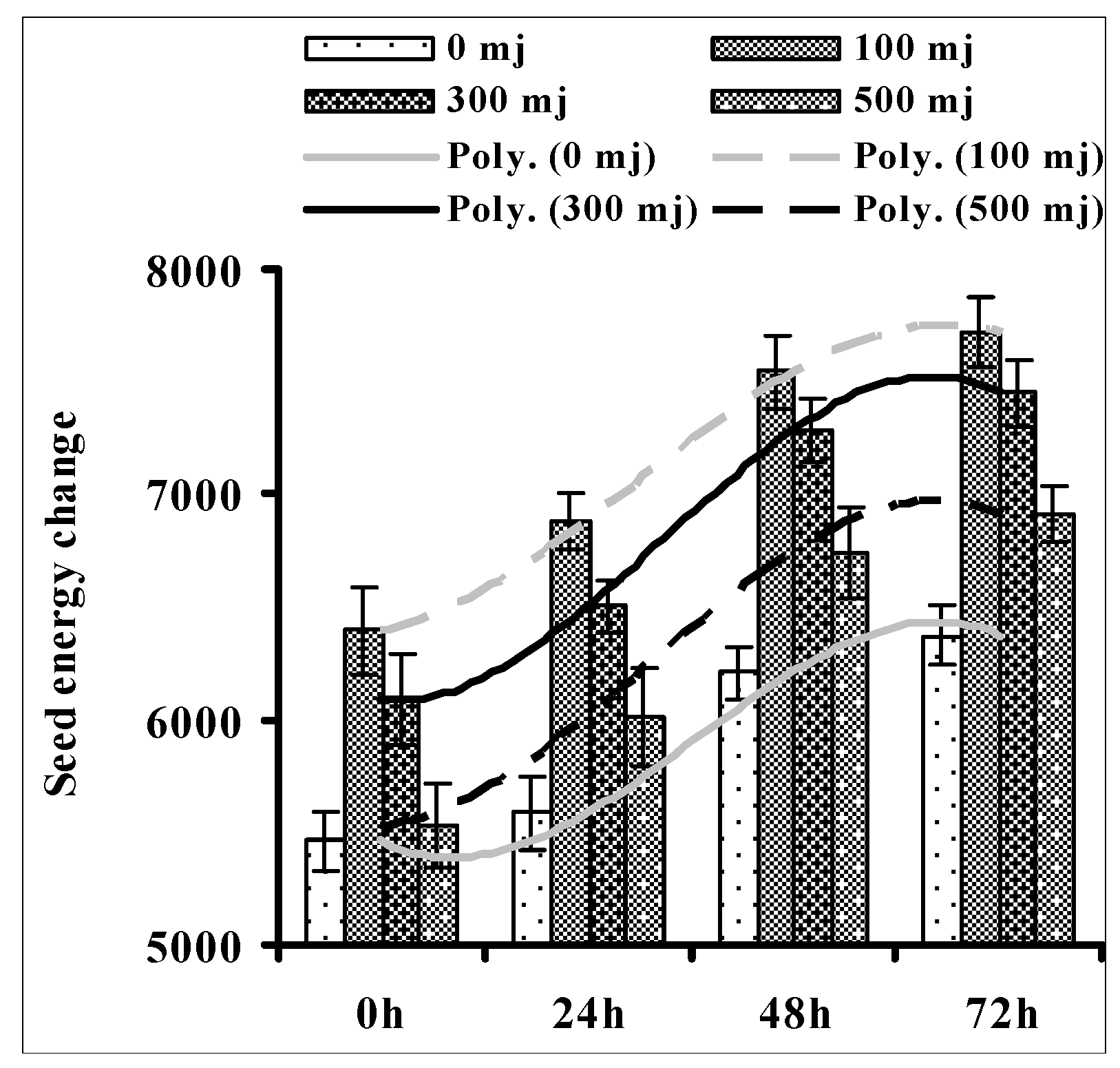
3.1. Effect of He–Ne Laser Energy on Seed Internal Energy during Germination
3.2. Effect of He–Ne Laser Energy on Seed Thermodynamics during Germination
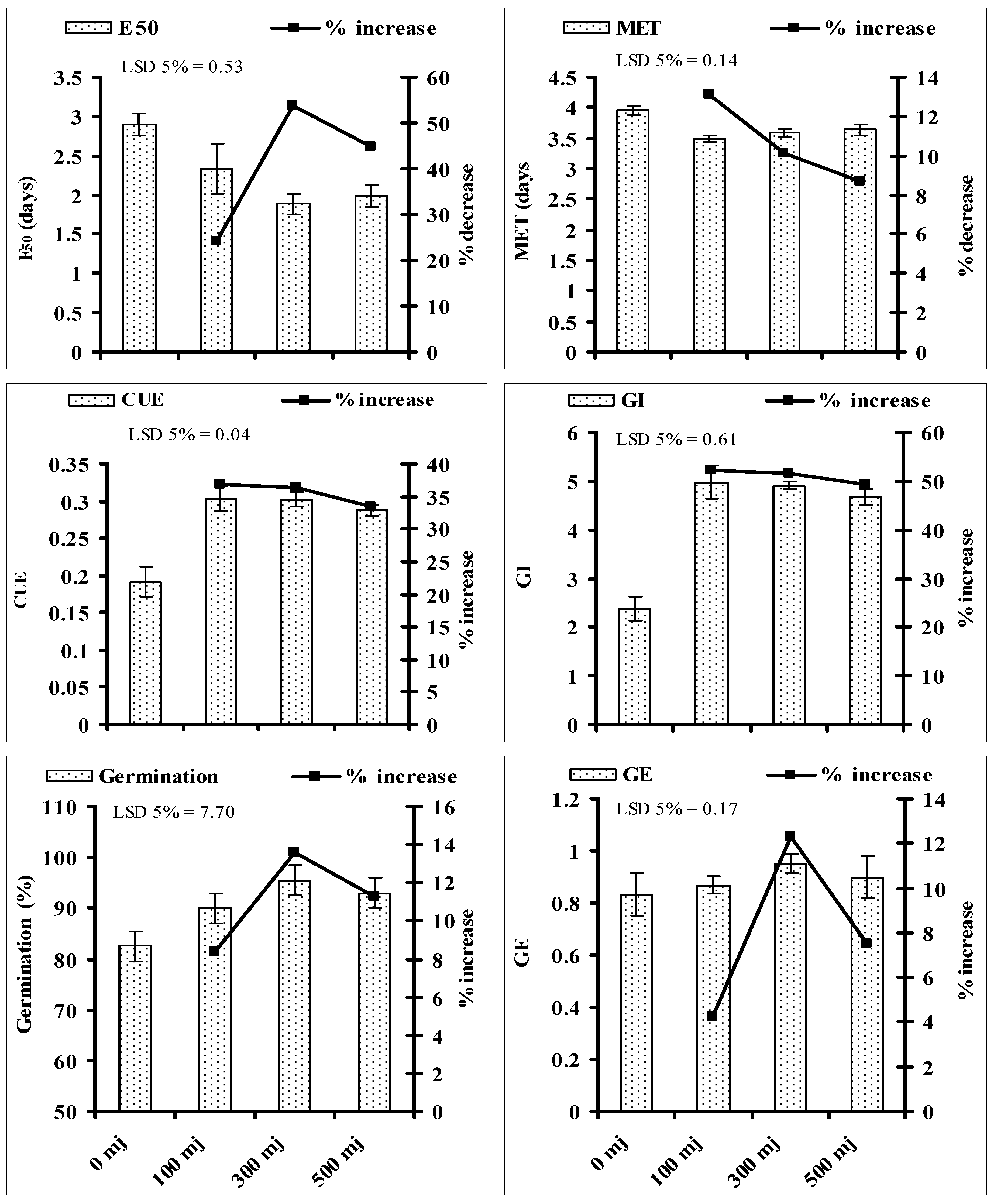
3.3. Effect of He–Ne Laser Energy on Seed Germination Related Attributes
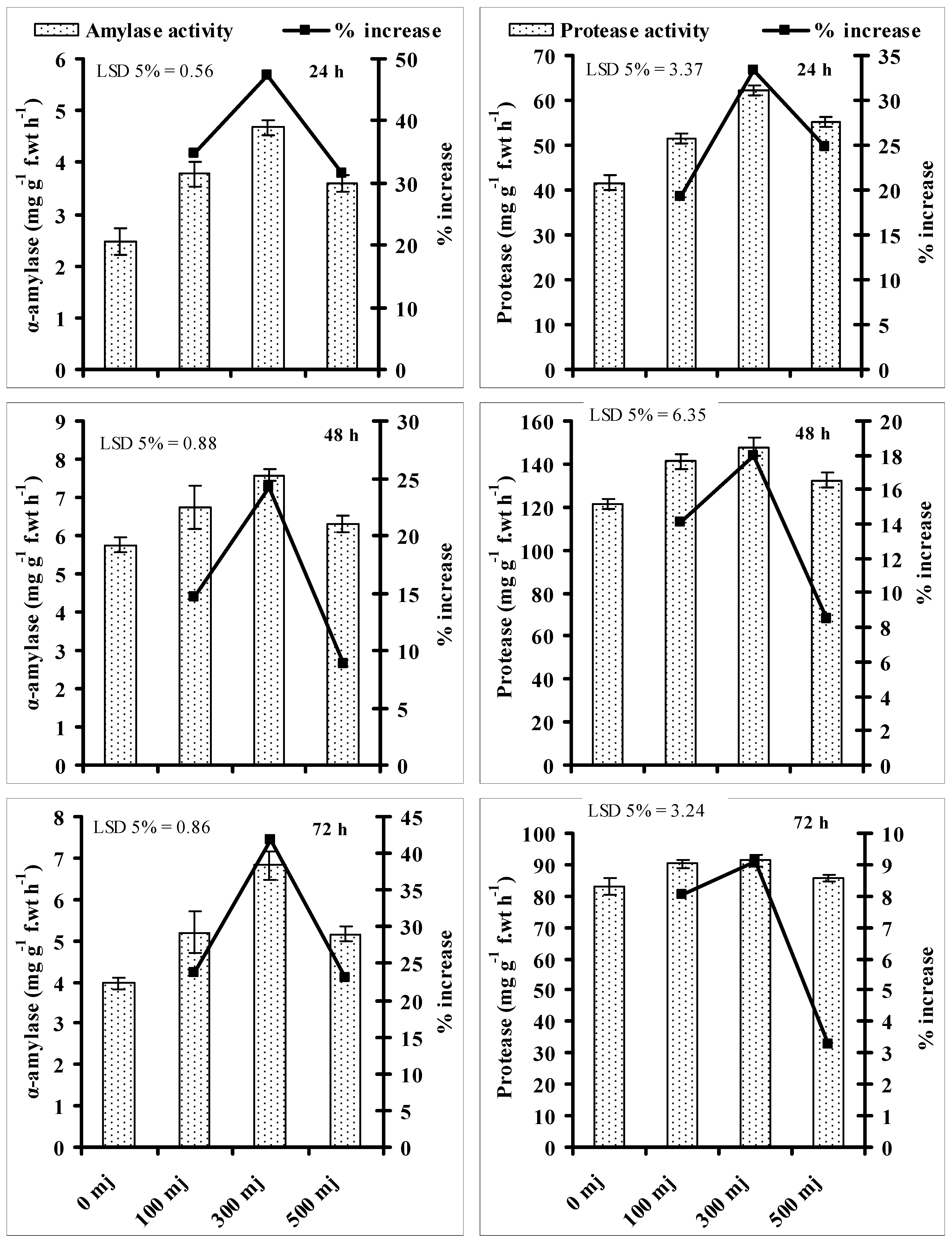
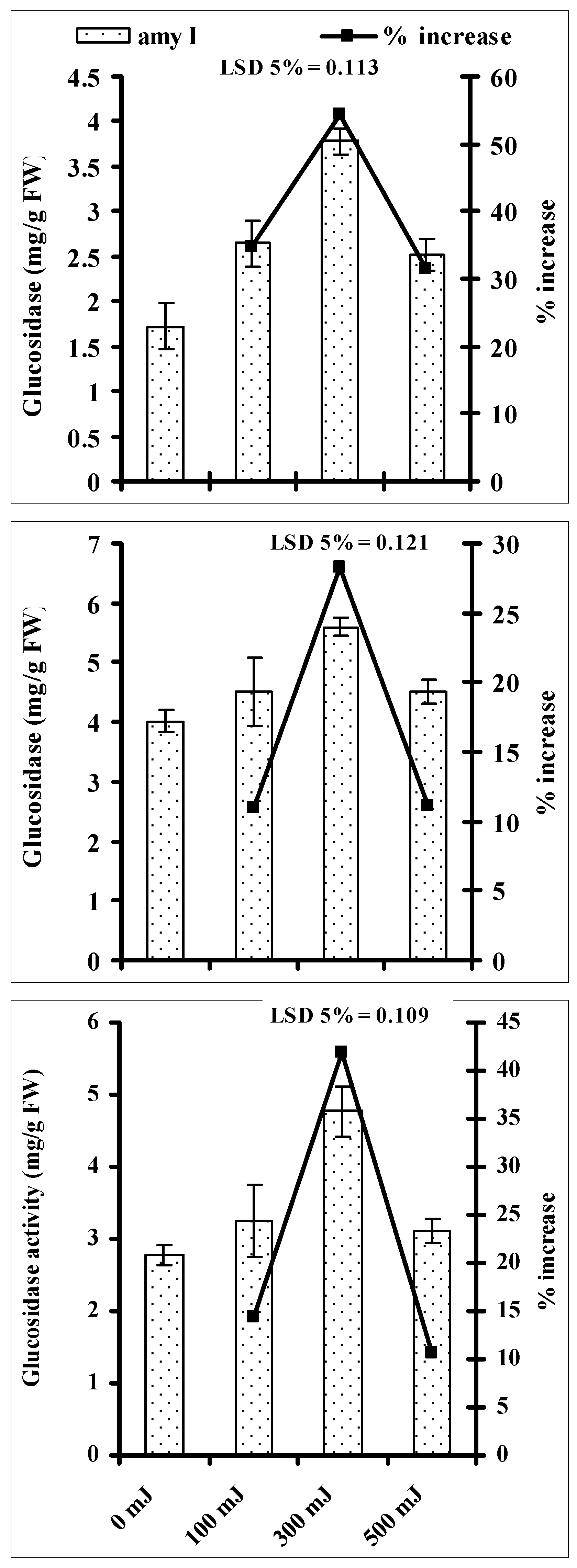
3.4. Effect of He–Ne Laser Energy on Activities of Seed Germination Enzymes
3.5. Effect of He–Ne Laser Energy on Seed Metabolites during Seed Germination
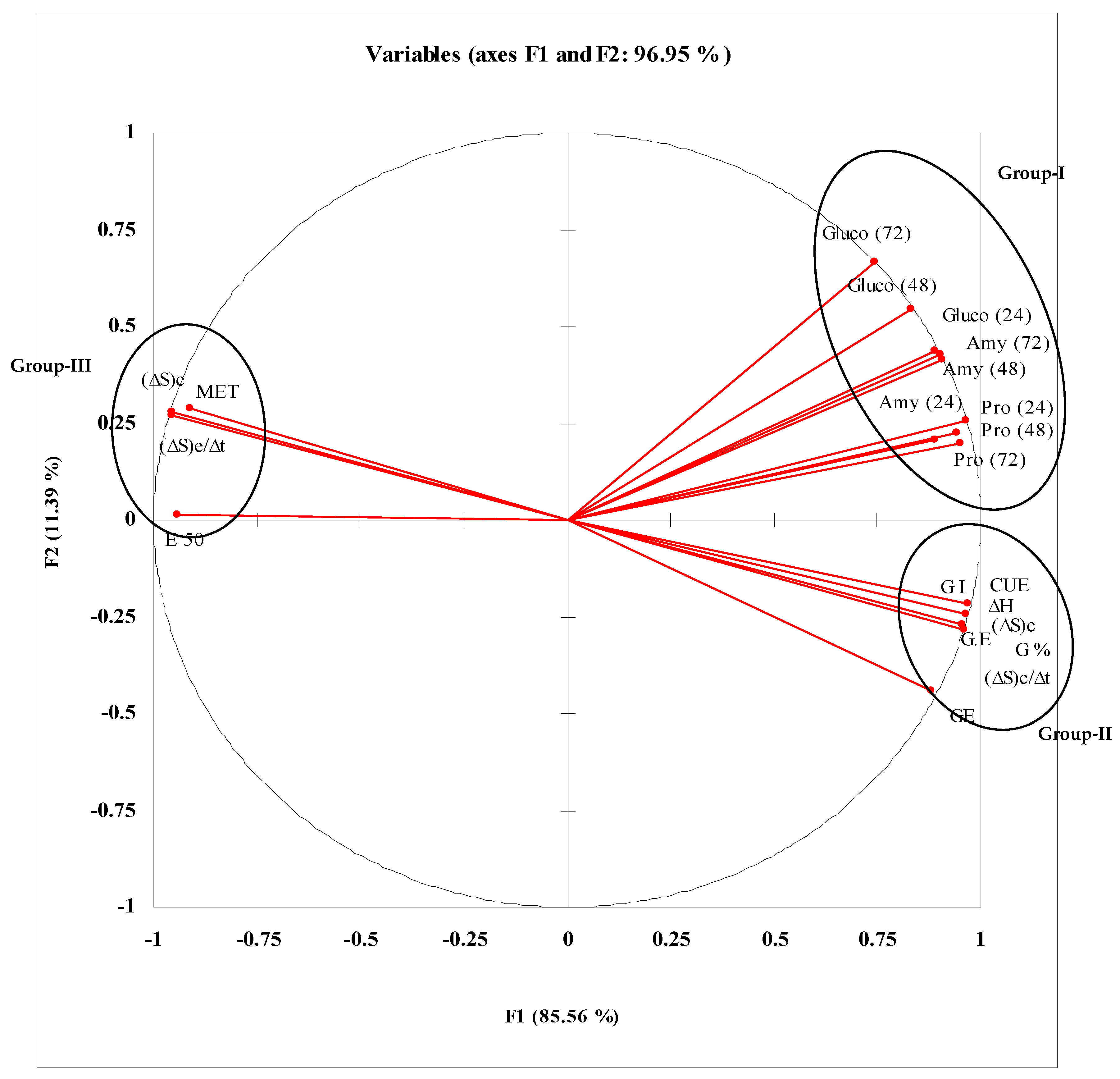
3.6. Correlations of Studied Attributes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali, Q.; Javed, M.T.; Noman, A.; Haider, M.Z.; Waseem, M.; Iqbal, N.; Waseem, M.; Shah, M.S.; Shahzad, F.; Perveen, R. Assessment of drought tolerance in mung bean cultivars/lines as depicted by the activities of germination enzymes, seedling’s antioxidative potential and nutrient acquisition. Arch. Agron. Soil. Sci. 2018, 64, 84–102. [Google Scholar] [CrossRef]
- Noman, A.; Ali, Q.; Maqsood, J.; Iqbal, N.; Javed, M.T.; Rasool, N.; Naseem, J. Deciphering physio-biochemical, yield, and nutritional quality attributes of waterstressed radish (Raphanus sativus L.) plants grown from Zn-Lys primed seeds. Chemosphere 2018, 195, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Ali, Q.; Ali, S.; Iqbal, N.; Javed, M.T.; Rizwan, M.; Khaliq, R.; Shahid, S.; Perveen, R.; Alamri, S.A.; Alyemeni, M.N.; et al. Alpha-tocopherol fertigation confers growth physio-biochemical and qualitative yield enhancement in field grown water deficit wheat (Triticum aestivum L.). Sci. Rep. 2019, 9, 12924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aladjadjiyan, A. The use of physical methods for plant growing stimulation in Bulgaria. J. Cent. Eur. Agric. 2007, 8, 369–380. [Google Scholar]
- Van Der Woud, W.J. A dimeric mechanism for the action of phytochrome: Evidence from photothermal interactions in lettuce seed germination. Photochem. Photobiol. 1985, 42, 655–661. [Google Scholar] [CrossRef]
- Shinomura, T. Phytochrome regulation of seed germination. J. Plant Res. 1997, 110, 151–161. [Google Scholar] [CrossRef]
- Muszyñski, S.; Gladyszewska, B. Representation of He–Ne laser irradiation effect on radish seeds with selected germination indices. Int. Agrophys. 2008, 22, 151–157. [Google Scholar]
- Perveen, R.; Ali, Q.; Ashraf, M.; Al-Qurainy, F.; Jamil, Y.; Ahmad, M.R. Effects of different doses of low power continuous wave He–Ne laser radiation on some seed thermodynamic and germination parameters and potential enzymes involved in seed germination of sunflower. Photochem. Photobiol. 2010, 86, 1050–1055. [Google Scholar] [CrossRef]
- Dirk, L.M.A.; Kumar, S.; Majee, M.; Downie, A.B. PHYTOCHROME INTERACTING FACTOR1 interactions leading to the completion or prolongation of seed germination. Plant Signal. Behav. 2018, 13. [Google Scholar] [CrossRef]
- Hartmann, K.M.; Grundy, A.C.; Market, R. Phytochrome- mediated long-term memory of seeds. Protoplasma 2005, 227, 47–52. [Google Scholar] [CrossRef]
- Azad, M.O.K.; Kim, W.W.; Park, C.H.; Cho, D.H. Effect of artificial LED light and far infrared irradiation on phenolic compound, isoflavones and antioxidant capacity in soybean (Glycine max L.) prout. Foods 2018, 7, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinoeve, S.; Antonov, M.; Stoyanov, T.; Georgieva, C. Spectral impact of low-power laser radiation on wheat and maize parameters. Prob. Engr. Cybern. Robot. 2004, 54, 74–85. [Google Scholar]
- Chen, Y.P.; Yuea, M.; Wang, X.L. Influence of He–Ne laser irradiation on seeds thermodynamic parameters and seedlings growth of Isatis indogotica. Plant Sci. 2005, 168, 601–606. [Google Scholar] [CrossRef]
- Cai, S.W.; Qi, Z.; Ma, X.L. The effect of He–Ne laser irradiation on soluble protein synthesis of corn seedling. Chin. J. Lasers 2000, 27, 284–288. [Google Scholar]
- Qi, Z.; Yue, M.; Wang, X.L. Laser pretreatment protects cells of broad bean from UV-B radiation damage. J. Photochem. Photobiol. 2000, 59, 33–37. [Google Scholar] [CrossRef]
- Qi, Z.; Yue, M.; Han, R.; Wang, X.L. The damage repair role of He–Ne laser on plants exposed to different intensities of UV-B irradiation. Photochem. Photobiol. 2002, 75, 680–686. [Google Scholar] [CrossRef]
- Han, R.; Wang, X.L.; Yue, M. Influence of He–Ne laser irradiation on the excision repair of cyclobutyl pyrimidine dimers in the wheat DNA. Chinese Sci. Bull. 2002, 47, 818–821. [Google Scholar] [CrossRef]
- Gładyszewska, B.; Koper, R.; Kornarzyski, K. Application and results of pre-sowing laser bio-stimulation of tomato seeds. Inz. Rol. 1998, 2, 37–44. [Google Scholar]
- Koper, R.; Rybak, P. Effect of pre-sowing laser biostimulation of glasshouse tomato seeds on physicochemical properties of the fruits. Inz. Rol. 2000, 5, 105–114. [Google Scholar]
- Koper, R.; £aziak, S.; WoŸniak, Z. A device for pre-sowing laser stimulation of seeds of chosen cultivated plants and the results of its application. Agric. Engin. 1997, 1, 63–69. (In Polish) [Google Scholar]
- Govil, S.R.; Agrawal, D.C.; Rai, K.P.; Thakur, S.N. Growth responses of Vigna radiata seeds to laser irradiation in the UV-A region. Physiol. Plant. 2006, 63, 133–134. [Google Scholar] [CrossRef]
- Wu, J.; Gao, X.; Zhang, S. Effect of laser pretreatment on germination and membrane lipid peroxidation of Chinese pine seeds under drought stress. Front. Biol. China 2007, 2, 314–317. [Google Scholar] [CrossRef]
- Jamil, Y.; Perveen, R.; Ashraf, M.; Ali, Q.; Iqbal, M.; Ahmad, M.R. He–Ne laser-induced changes in germination, thermodynamic parameters, internal energy, enzyme activities and physiological attributes of wheat during germination and early growth. Laser Phys. Lett. 2013, 10, 1–13. [Google Scholar] [CrossRef]
- Abu-Elsaoud, A.M.; Shahda, R.H. Role of He–Ne laser pre-treatment in protecting Zea mays against the deleterious effects of ultraviolet radiations. Egypt. J. Exp. Biol. 2017, 13, 403–422. [Google Scholar] [CrossRef]
- Tabatabaei, S.A.; Ansari, O. Predicting seed germination of safflower (carthamus tinctorius) cultivars using hydrotime model. Cercet. Agron. Mold. 2017, 1, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Association of Official seed Analysis (AOSA), Rules for testing seeds. J. Seed Technol. 1990, 12, 1–112.
- Coolbear, P.; Francis, A.; Grierson, D. The effect of low temperature pre-sowing treatment under germination performance and membrane integrity of artificially aged tomato seeds. J. Exp. Bot. 1984, 35, 1609–1617. [Google Scholar] [CrossRef]
- Ellis, R.A.; Roberts, E.H. The qualification of aging and survival in orthodox seeds. Seed Sci. Technol. 1981, 9, 373–409. [Google Scholar]
- Bewley, J.D.; Black, M. Seed Physiology of Development and Germination; Plenum Press: New York, NY, USA, 1985. [Google Scholar]
- Association of Official Seed Analysis (AOSA). Seed Vigor Testing Handbook, 1st ed; Contribution No. 32 to the handbook on seed testing; Association of Official Seed Analysis; AOSA: East Lasing, MI, USA, 1983. [Google Scholar]
- Ruan, S.; Xue, Q.; Tylhowska, K. The influence of priming on germination of rice (Oryza sativa L.) seeds and seedlings emergence and performance in flooded soils. Seed Sci. Technol. 2002, 30, 61–67. [Google Scholar]
- Chrispeel, M.J.; Vaner, K.E. Gibberelic acid enhanced synthesis and release of alpha-amylase and ribonuclease by isolated barley aleurone layers. Plant Physiol. 1967, 42, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Ainouz, I.I.; Filho, J.X.; Filho, E.G. Atividade proteolitica msementes de Vigna sinensis cv. serida Cienc. Cult. Sao Panlo Suppl. 1972, 24, 104. [Google Scholar]
- Lowry, O.H.; Roebrough, N.J.; Randall, R.J.; Farr, A.L. Protein measurement with folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Dawson, R.C.M.; Elliot, D.C.; Elliot, W.H.; Jones, K.M. Data for Biochemical Research; Oxford at the Claren don Press: London, UK, 1969; p. 484. [Google Scholar]
- Lloyd, J.B.; Whelan, W.J. An improved method for enzymatic determination of glucose in the presence of maltose. Anal. Biochem. 1969, 30, 467–469. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzyme in isolated chloroplast Polyphenol oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirk, J.T.O.; Allen, R.L. Dependence of chloroplast pigment synthesis on protein synthesis: Effect of actidione. Biochem. Biophys. Res. Commun. 1965, 21, 523–530. [Google Scholar] [CrossRef]
- Tonguç, M.; Elkoyunu, R.; Erbaş, S.; Karakurt, Y. Changes in seed reserve composition during germination and initial seedling development of safflower (Carthamus tinctorius L.). Turk. J. Biol. 2012, 36, 107–112. [Google Scholar]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Somogyi, M. Notes on sugar determination. J. Biol. Chem. 1952, 195, 19–23. [Google Scholar] [CrossRef]
- Larson, L.A.; Beevers, H. Amino acid metabolism in young pea seedlings. Plant Physiol. 1963, 40, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Noctor, G.; Bergot, G.L.; Mauve, C.; Thominet, D.; Lelarge-Trouverie, C.; Prioul, J.L. A comparative study of amino acid measurement in leaf extracts by gas chromatography-time of flight-mass spectrometry and high-performance liquid chromatography with fluorescence detection. Metabolomics 2007, 3, 161–174. [Google Scholar] [CrossRef]
- Lee, P.Y.; Takahashi, T. An improved colorimetric determination of amino acids with the use of ninhydrin. Anal. Biochem. 1966, 14, 71–77. [Google Scholar] [CrossRef]
- Lowry, R.R.; Tinsley, I.J. Rapid colorimetric determination of free fatty acids. J. Am. Oil Chem. Soc. 1976, 53, 470–472. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, K.M.; Mollwo, A. The action spectrum for maximal photosensitivity of germination. Sci. Nat. 2000, 87, 398–403. [Google Scholar] [CrossRef]
- Criddle, R.S.; Breidenbach, R.W.; Hansen, L.D. Plant calorimetry: How to quantitatively compare apples and oranges. Thermochim. Acta 1991, 193, 67–90. [Google Scholar] [CrossRef]
- Dziwulska, A.; Koper, R. Effects of pre-sowing laser bio-stimulation on germination of Lucerne seeds. Acta Agrophysica 2003, 82, 33–39. (In Polish) [Google Scholar]
- Saleem, M.H.; Rehman, M.; Zahid, M.; Imran, M.; Xiang, W.; Liu, L. Morphological changes and antioxidative capacity of jute (Corchorus capsularis, Malvaceae) under different color light-emitting diodes. Braz. J. Bot. 2019, 42, 581–590. [Google Scholar] [CrossRef]
- Raffa, R.B. Thermodynamics of the drug receptor interaction. Life Sci. 1999, 59, 967–980. [Google Scholar] [CrossRef]
- Yan, L.F.; Zhan, Y.L. Molecular Biology; Agriculture University of China Press: Beijing, China, 1997. (In Chinese) [Google Scholar]
- Alam, H.; Khattak, J.Z.K.; Ksiksi, T.S.; Saleem, M.H.; Fahad, S.; Sohail, H.; Ali, Q.; Zamin, M.; El-Esawi, M.A.; Saud, S. Negative impact of long-term exposure of salinity and drought stress on native Tetraena mandavillei L. Physiol. Plant. 2020, 1–16. [Google Scholar] [CrossRef]
- Shinomura, T.; Nagatani, A.; Chory, J.; Furuya, M. The induction of seed germination in Arabidopsis thaliana is regulated principally by phytochrome B and secondarily by phytochrome A. Plant Physl. 1994, 104, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Shichijo, C.; Katada, K.; Tanaka, O.; Hashimoto, T. Phytochrome A-mediated inhibition of seed germination in tomato. Planta 2001, 213, 764–769. [Google Scholar] [CrossRef]
- Perveen, R.; Jamil, Y.; Ashraf, M.; Ali, Q.; Iqbal, M.; Ahmad, M.R. He–Ne laser-induced improvement in biochemical, physiological, growth and yield characteristics in sunflower (Helianthus annuus L.). Photochem. Photobiol. 2011, 87, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- Gladyszewska, B.; Koper, R. Determination of free radical concentration in tomato seeds biostimulated by laser irradiation (in Polish). Inynieria Rol. 2000, 4, 35–42. [Google Scholar]
- Podleoeny, J.; Misiak, L.; Koper, R. Concentration of free radicals in faba bean seeds after the pre-sowing treatment of the seeds with laser light. Int. Agrophys. 2001, 15, 185–189. [Google Scholar]
- Salyaev, R.K.; Dudareva, L.V.; Lankevich, S.V.; Ekimova, E.G.; Sumtsova, V.M. Effect of low-intensity laser radiation on the lipid peroxidation in wheat callus culture. Russ. J. Plant Physl. 2003, 50, 498–500. [Google Scholar] [CrossRef]
- Rubinov, A.N. Physical grounds for biological effect of laser radiation. J. Phys. D Appl. Phys. 2003, 6, 2317–2330. [Google Scholar] [CrossRef]
- Ali, Q.; Ahmar, S.; Sohail, M.A.; Kamran, M.; Ali, M.; Saleem, M.H.; Rizwan, M.; Ahmed, A.M.; Mora-Poblete, F.; do Amaral Júnior, A.T. Research advances and applications of biosensing technology for the diagnosis of pathogens in sustainable agriculture. Environ. Sci. Poll. Res. 2021, 1–18. [Google Scholar]
- Samuilov, F.D.; Garifullina, R.L. Effect of laser irradiation on microviscosity of aqueous medium in imbibing maize seeds as studied with a spin probe method. Russ. J. Plant Physl. 2007, 54, 128–131. [Google Scholar] [CrossRef]
- Rassam, Y.Z.; Mashhadani, F.A.A.; Boya, A.F. Laser treatment may enhance growth and resistance to fungal infection of hard wheat Seeds. J. Agric. Vet. Sci. 2012, 2, 47–51. [Google Scholar]
- Khalifa, N.S.; El-Ghandoor, H. Investigate the effect of Nd-Yag laser beam on soybean (Glycin max) leaves at the protein level. Int. J. Biol. 2011, 3, 135. [Google Scholar] [CrossRef] [Green Version]
- Hameed, A.; Akram, N.A.; Saleem, M.H.; Ashraf, M.; Ahmed, S.; Ali, S.; Abdullah Alsahli, A.; Alyemeni, M.N. Seed Treatment with α-Tocopherol Regulates Growth and Key Physio-Biochemical Attributes in Carrot (Daucus carota L.) Plants under Water Limited Regimes. Agronomy 2021, 11, 469. [Google Scholar] [CrossRef]
- Vasilevski, G. Perspectives of the application of biophysical methods in sustainable agriculture. Bulg. J. Plant Physliol. 2003, 29, 179–186. [Google Scholar]
- Li, Y.W.; Feng, W.X. The effects of He–Ne laser treatment on seeds germination and growth of atractylodes macrocephala. Chin. J. Appl. Laser 1996, 16, 37–41. [Google Scholar]
- Qiu, Z.B.; Zhu, X.J.; Li, F.M.; Liu, X.; Yue, M. The optical effect of a semiconductor laser on protecting wheat from UV-B radiation damage. Photochem. Photobiol. Sci. 2007, 6, 788–793. [Google Scholar] [CrossRef]
- Kuznetsov, V.l.V.; Dmitrieva, G.A. Fiziologiya Rastenii (Plant Physiology); Vysshaya Shkola: Moscow, Russia, 2005. [Google Scholar]
- Shakhov, A.A. Fotoenergetika Rastenii i Urozhai (Photoenergetics of Plants and the Yield); Nauka: Moscow, Russia, 1993. [Google Scholar]
- Koper, R.; Dziwulska, A. Effects of laser stimulation of white lupine seeds. Acta Agrophysica 2003, 82, 99–106. (In Polish) [Google Scholar]
- Zhang, H.; Gao, Z.; Li, Z.; Du, H.; Lin, B.; Cui, M.; Yin, Y.; Lei, F.; Yu, C.; Meng, C. Laser radiation induces growth and lipid accumulation in the seawater microalga Chlorella pacifica. Energies 2017, 10, 1671. [Google Scholar] [CrossRef] [Green Version]
- Liburdy, R.P. Microwave bioeffects in erythrocytes. Bioelectromagnetics 1985, 6, 283–289. [Google Scholar]
- Danie, R.M. The upper limits of enzyme thermal stability. Enzyme Microb. Technol. 1996, 19, 74–79. [Google Scholar] [CrossRef]
- Podlesny, J. Effect of laser irradiation on the biochemical changes in seeds and the accumulation of dry matter in the faba bean. Int. Agrophys. 2002, 16, 209–213. [Google Scholar]
- Xu, M.F. Biological effects of lasers in wheat. Appl. Laser 1995, 153, 131. (In Chinese) [Google Scholar]
- Hong, F.; Lin, L. Effects of laser irradiation on amylase in the seeds of Bailan melon during germination. J. Southw. Uni. Natl. 2005, 2. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| He–Ne Laser Energy Doses (mJ) | |||||
|---|---|---|---|---|---|
| 0 | 100 | 300 | 500 | LSD 5% | |
| ∆H (J) | 108.57 ± 3.43 a | 158.15 ± 2.98 b | 162.83 ± 3.82 b | 164.58 ± 4.60 b | 10.01 |
| (∆S)c (J/K) | 0.36 ± 0.011 a | 0.530 ± 0.12 b | 0.54 ± 0.013 b | 0.55 ± 0.015 b | 0.03 |
| (∆S)e (J/K) | −0.36 ± 0.011 a | −0.531 ± 0.012 b | −0.546 ± 0.013 b | −0.552 ± 0.015 b | 0.03 |
| (∆S)c/∆t (μW/K) | 1.41 ± 0.045 a | 2.048 ± 0.040 b | 2.108 ± 0050 b | 2.13 ± 0.061 b | 0.13 |
| (∆S)e/∆t (μW/K) | −1.41 ± 0.045 a | −2.05 ± 0.038 b | −2.108 ± 0.050 b | −2.14 ± 0.052 b | 0.12 |
| Chlorophyll (mg/g of Germinating Seeds) (LSD 5% = 0.25) | Carotenoids (μg/g of Germinating Seeds) (LSD 5% = 0.065) | |||||||
| Time Interval (h) | 0 mJ | 100 mJ | 300 mJ | 500 mJ | 0 mJ | 100 mJ | 300 mJ | 500 mJ |
| 0 | w 0 c | w 0 b | w 0 c | w 0 d | w 0 c | w 0 c | w 0 c | w 0 c |
| 24 | w 0.09 ± 0.002 c | w 0.16 ± 0.070 b | w 0.122 ± 0.005 c | w 0.114 ± 0.005 d | w 0.014 ± 0.006 c | w 0.042 ± 0.003 b | w 0.034 ± 0.001 c | w 0.028 ± 0.002 c |
| 48 | w 0.19 ± 0.010 c | w 0.42 ± 0.037 b | x 0.710 ± 0.029 b | w 0.410 ± 0.031 c | x 0.054 ± 0.003 c | x 0.110 ± 0.006 b | w 0.142 ± 0.005 b | x 0.113 ± 0.006 b |
| 72 | w 0.61 ± 0.028 b | x 1.09 ± 0.073 a | x 1.221 ± 0.065 a | x 1.082 ± 0.060 b | z 0.150 ± 0.021 b | y 0.260 ± 0.015 a | w 0.349 ± 0.016 a | x 0.280 ± 0.014 a |
| 96 | w,x 1.08 ± 0.055 a | w,x 1.28 ± 0.076 a | w 1.429 ± 0.069 a | w 1.371 ± 0.069 a | y 0.280 ± 0.013 a | x 0.320 ± 0.016 a | w 0.407 ± 0.017 a | x 0.329 ± 0.018 a |
| Total Soluble Sugars (mg/g of Germinating Seeds) (LSD 5% = 6.56) | Reducing Sugars (mg/g of Germinating Seeds) (LSD 5% = 4.10) | |||||||
| Time Interval (h) | 0 mJ | 100 mJ | 300 mJ | 500 mJ | 0 mJ | 100 mJ | 300 mJ | 500 mJ |
| 0 | w 16.25 ± 0.92 e | w 18.24 ± 0.88 e | w 20.35 ± 0.75 e | w 16.45 ± 0.88 e | w 5.90 ± 0.25 d | w 5.99 ± 0.31 d | w 6.15 ± 0.33 d | w 6.05 ± 0.33 d |
| 24 | y 17.31 ± 0.88 d | x 24.33 ± 0.99 d | w 32.25 ± 1.33 d | x,y 21.12 ± 0.55 d | w 7.25 ± 0.28 c,d | w 7.12 ± 0.33 d | w 8.11 ± 0.35 d | w 7.43 ± 0.25 d |
| 48 | x 34.33 ± 1.55 c | x 39.55 ± 1.95 c | w 52.21 ± 2.15 c | x 38.23 ± 1.55 c | y 11.35 ± 0.58 c | yx 14.65 ± 0.61 c | w 20.23 ± 0.61 c | x 15.65 ± 0.55 c |
| 72 | x 47.56 ± 1.99 b | x 51.65 ± 2.25 b | w 67.65 ± 1.99 b | x 49.33 ± 1.55 b | z 16.35 ± 0.91 b | x,y 34.15 ± 1.82 b | w 42.11 ± 1.55 b | y 31.12 ± 0.99 b |
| 96 | x 58.65 ± 1.97 a | x 64.33 ± 3.01 a | w 72.15 ± 3.15 a | x 61.23 ± 1.85 a | y 32.11 ± 0.99 a | x 43.00 ± 1.98 a | w 53.23 ± 1.99 a | x 41.25 ± 2.01 a |
| Total Protein (mg/kg of Germinating Seeds) (LSD 5% = 62.40) | Total Free Amino Acids (mg/kg of Germinating Seeds) (LSD 5% = 51.70) | |||||||
| Time Interval (h) | 0 mJ | 100 mJ | 300 mJ | 500 mJ | 0 mJ | 100 mJ | 300 mJ | 500 mJ |
| 0 | w 688 ± 8.9 a | w 699 ± 5.56 a | w 711 ± 4.45 a | w 710 ± 5.6 a | w 55 ± 1.99 d | w 67 ± 2.33 e | w 63 ± 2.34 e | w 67 ± 2.34 d |
| 24 | w 600 ± 5.7 b | w 570 ± 4.99 b | w 540 ± 3.65 b | w 592 ± 4.3 b | x 75 ± 1.65 d | w,x 121 ± 3.21 d | w 135 ± 3.15 d | w,x 99 ± 3.25 d |
| 48 | w 460 ± 4.5 c | x,y 340 ± 7.11 c | y 305 ± 4.35 c | x 401 ± 6.3 c | x 175 ± 2.24 c | w,x 225 ± 3.22 c | w 242 ± 2.99 c | w,x 201 ± 3.43 c |
| 72 | w 290 ± 6.3 d | w,x 240 ± 5.11 d | x 215 ± 7.12 d | w,x 262 ± 8.0 d | x 345 ± 2.65 b | w 405 ± 3.21 b | w 435 ± 3.45 b | w 399 ± 3.45 b |
| 96 | w 230 ± 5.2 d | w,x 190 ± 4.10 e | x,y 165 ± 4.98 d | y 215 ± 8.1 e | y 499 ± 3.11 a | w 601 ± 2.44 a | w 605 ± 3.65 a | x 561 ± 3.55 a |
| Oil Content (LSD 5% = 3.36) | Free Fatty Acids (LSD 5% = 7.61) | |||||||
| Time Interval (h) | 0 mJ | 100 mJ | 300 mJ | 500 mJ | 0 mJ | 100 mJ | 300 mJ | 500 mJ |
| 0 | w 33.05 ± 2.16 a | w 33.1 ± 0.13 a | w 33.05 ± 0.12 a | w 32.95 ± 0.14 a | w 45 ± 2.33 d | w 38 ± 2.44 d | w,x 41 ± 2.45 e | x 36 ± 2.78 d |
| 24 | w 32.9 ± 3.25 a,b | x 28.7 ± 0.08 b | y 24.65 ± 0.32 b | w,x 30.85 ± 0.22 a | w 65 ± 1.55 c | w 67 ± 2.34 c | w 65 ± 2.44 c | w 70 ± 1.56 c |
| 48 | w 26.1 ± 2.14 b | x 20.5 ± 0.11 c | y 15.90 ± 0.22 c | w 24.90 ± 0.11 b | x 105 ± 2.34 b | w 120 ± 2.54 b | w 115 ± 2.45 b | w 115 ± 1.67 b |
| 72 | w 18.9 ± 0.15 c | x 14.6 ± 0.21 d | y 8.80 ± 0.23 d | w,x 15.85 ± 0.10 c | x,y 125 ± 1.65 a | x 131 ± 2.56 a | w 201 ± 2.36 a | y 121 ± 1.22 a |
| 96 | w 12.5 ± 0.08 d | w 10.2 ± 0.11 e | x 5.45 ± 0.12 e | w 9.40 ± 0.04 d | x 120 ± 1.45 a | w 128 ± 2.45 a | x 120 ± 2.34 b | wx 123 ± 2.33 a |
| Variables | Amy (24) | Amy (48) | Amy (72) | Pro (24) | Pro (48) | Pro (72) | Gluco (24) | Gluco (48) | Gluco (72) |
|---|---|---|---|---|---|---|---|---|---|
| Amy (24) | 1 | 0.972 *** | 0.982 *** | 0.967 *** | 0.972 *** | 0.916 *** | 0.982 *** | 0.942 *** | 0.890 *** |
| Amy (48) | 0.972 *** | 1 | 0.977 *** | 0.897 *** | 0.972 *** | 0.939 *** | 0.985 *** | 0.962 *** | 0.945 *** |
| Amy (72) | 0.982 *** | 0.977 *** | 1 | 0.961 *** | 0.932 *** | 0.863 *** | 0.999 *** | 0.988 *** | 0.955 *** |
| Pro (24) | 0.967 *** | 0.897 *** | 0.961 *** | 1 | 0.882 *** | 0.787 *** | 0.950 *** | 0.924 *** | 0.852 *** |
| Pro (48) | 0.972 *** | 0.972 *** | 0.932 *** | 0.882 *** | 1 | 0.985 *** | 0.941 *** | 0.882 *** | 0.841 *** |
| Pro (72) | 0.916 *** | 0.939 *** | 0.863 *** | 0.787 *** | 0.985 *** | 1 | 0.880 *** | 0.809 *** | 0.780 *** |
| Gluco (24) | 0.982 *** | 0.985 *** | 0.999 *** | 0.950 *** | 0.941 *** | 0.880 *** | 1 | 0.988 *** | 0.960 *** |
| Gluco (48) | 0.942 *** | 0.962 *** | 0.988 *** | 0.924 *** | 0.882 *** | 0.809 *** | 0.988 *** | 1 | 0.988 *** |
| Gluco (72) | 0.890 *** | 0.945 *** | 0.955 *** | 0.852 *** | 0.841 *** | 0.780 *** | 0.960 *** | 0.988 *** | 1 |
| E 50 | −0.902 *** | −0.781 *** | −0.871 *** | −0.972 *** | −0.801 *** | −0.694 ** | −0.852 *** | −0.810 *** | −0.708 *** |
| MET | −0.811 *** | −0.727 ** | −0.687 ** | −0.738 ** | −0.866 *** | −0.870 *** | −0.691 ** | −0.571 * | −0.473 * |
| CUE | 0.885 *** | 0.784 *** | 0.784 *** | 0.854 *** | 0.892 *** | 0.860 *** | 0.781 *** | 0.679 ** | 0.575 * |
| G index | 0.874 *** | 0.767 *** | 0.770 *** | 0.849 *** | 0.878 *** | 0.844 *** | 0.766 *** | 0.663 ** | 0.555 * |
| G % | 0.854 *** | 0.728 ** | 0.752 *** | 0.855 *** | 0.839 *** | 0.791 *** | 0.743 ** | 0.643 ** | 0.525 * |
| G.E | 0.855 *** | 0.718 ** | 0.763 *** | 0.880 *** | 0.816 *** | 0.751 *** | 0.751 *** | 0.660 ** | 0.539 * |
| ∆H | 0.855 *** | 0.718 ** | 0.763 *** | 0.880 *** | 0.816 *** | 0.751 *** | 0.751 *** | 0.660 ** | 0.539 * |
| (∆S)c | 0.855 *** | 0.718 ** | 0.763 *** | 0.880 *** | 0.816 *** | 0.751 *** | 0.751 *** | 0.660 ** | 0.539 * |
| (∆S)e | −0.855 *** | −0.718 ** | −0.763 *** | −0.880 *** | −0.816 *** | −0.751 *** | −0.751 *** | −0.660 ** | −0.539 * |
| (∆S)c/∆t | 0.855 *** | 0.718 ** | 0.763 *** | 0.880 *** | 0.816 *** | 0.752 *** | 0.751 *** | 0.660 ** | 0.539 * |
| (∆S)e/∆t | −0.850 *** | −0.711 ** | −0.757 *** | −0.877 *** | −0.809 *** | −0.744 ** | −0.744 ** | −0.653 ** | −0.530 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perveen, R.; Wang, X.; Jamil, Y.; Ali, Q.; Ali, S.; Zakaria, M.Q.; Afzaal, M.; Kasana, R.A.; Saleem, M.H.; Fiaz, S. Quantitative Determination of the Effects of He–Ne Laser Irradiation on Seed Thermodynamics, Germination Attributes and Metabolites of Safflower (Carthamus tinctorius L.) in Relation with the Activities of Germination Enzymes. Agronomy 2021, 11, 1411. https://doi.org/10.3390/agronomy11071411
Perveen R, Wang X, Jamil Y, Ali Q, Ali S, Zakaria MQ, Afzaal M, Kasana RA, Saleem MH, Fiaz S. Quantitative Determination of the Effects of He–Ne Laser Irradiation on Seed Thermodynamics, Germination Attributes and Metabolites of Safflower (Carthamus tinctorius L.) in Relation with the Activities of Germination Enzymes. Agronomy. 2021; 11(7):1411. https://doi.org/10.3390/agronomy11071411
Chicago/Turabian StylePerveen, Rashida, Xiukang Wang, Yasir Jamil, Qasim Ali, Shafaqat Ali, Muhammad Qaisar Zakaria, Muhammad Afzaal, Rehan Ahmed Kasana, Muhammad Hamzah Saleem, and Sajid Fiaz. 2021. "Quantitative Determination of the Effects of He–Ne Laser Irradiation on Seed Thermodynamics, Germination Attributes and Metabolites of Safflower (Carthamus tinctorius L.) in Relation with the Activities of Germination Enzymes" Agronomy 11, no. 7: 1411. https://doi.org/10.3390/agronomy11071411