
The Brown Marmorated Stink Bug (Halyomorpha halys Stål.) Influences Pungent and Non-Pungent Capsicum Cultivars’ Pre- and Post-Harvest Quality


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design
2.2. Treatments
- (1)
- control treatment (Control 1);
- (2)
- H. halys treatment (H. halys);
- (3)
- empty bag treatment (Control 2).
2.3. Pepper Picking
2.4. Storage Conditions
2.5. Extraction of Sugars and Organic Acids
2.6. Extraction of Phenolics and Capsaicinoids
2.7. Chemicals
2.8. Statistical Analysis
3. Results
3.1. Visual Appearance of Pepper Fruit and Water Loss
3.2. Indiviudal and Total Sugar Contents
3.3. Individual and Total Organic Acid Contents
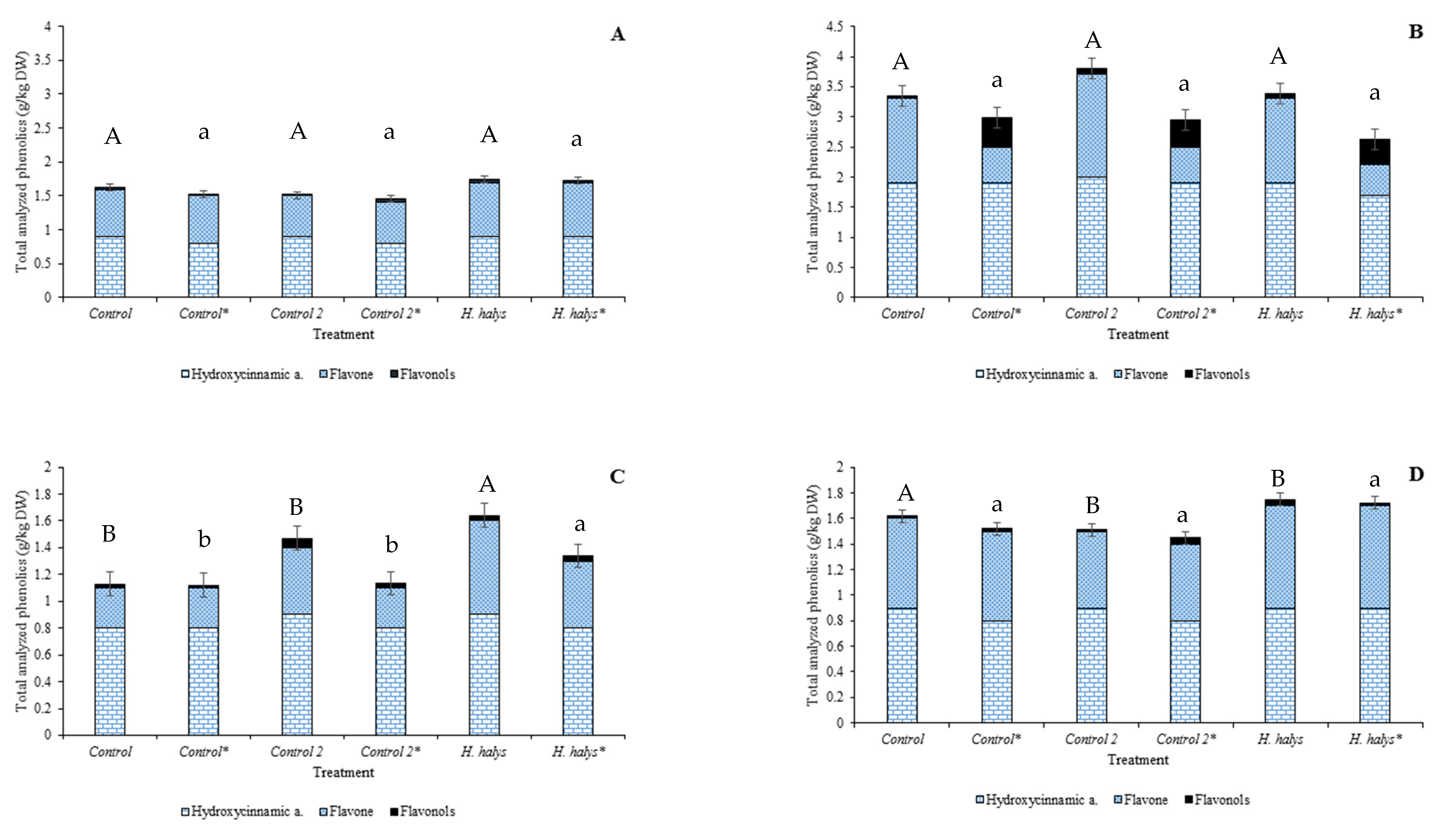
3.4. Individual and Total Phenolics Contents
3.5. Individual and Total Capsaicinoid Contents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Penella, C.; Calatayud, A. Pepper Crop under Climate Change: Grafting as an Environmental Friendly Strategy. In Climate Resilient Agriculture: Strategies and Perspectives; IntechOpen: London, UK, 2018. [Google Scholar]
- Bicikliski, O.; Trajkova, F.; Mihajlov, L.; Jordanovska, S.; Tashev, K. Vitamin C and Total Antioxidant Content in Pepper Fruits (Capsicum annuum L.): Comparative Analysis of Peppers Grown in Conventional and Organic Agricultural Systems. Annu. Res. Rev. Biol. 2018, 27, 1–11. [Google Scholar] [CrossRef]
- Cronin, J.R. The Chili Pepper’s Pungent Principle: Capsaicin Delivers Diverse Health Benefits. Altern. Complement. Ther. 2002, 8, 110–113. [Google Scholar] [CrossRef]
- Zamljen, T.; Zupanc, V.; Slatnar, A. Influence of irrigation on yield and primary and secondary metabolites in two chilies species, Capsicum annuum L. and Capsicum chinense Jacq. Agric. Water Manag. 2020, 234, 106104. [Google Scholar] [CrossRef]
- Daryanto, A.; Syukur, M.; Hidayat, P.; Maharijaya, A. Antixenosis and antibiosis based resistance of chili pepper to melon aphid. J. Appl. Hort 2017, 19, 147–151. [Google Scholar] [CrossRef]
- Johari, A.; Herlinda, S.; Irsan, C.; Pujiastuti, Y. Phenomenon of Thrips (Thysanoptera) Attack on Chili Plant (Capsicum annuum L.). Am. J. Agric. Biol. Sci. 2016, 11, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Moreau, T.L.; Isman, M.B. Combining reduced-risk products, trap crops and yellow sticky traps for greenhouse whitefly (Trialeurodes vaporariorum) management on sweet peppers (Capsicum annum). Crop Prot. 2012, 34, 42–46. [Google Scholar] [CrossRef]
- Wermelinger, B.; Wyniger, D.; Forster, B. First records of an invasive bug in Europe: Halyomorpha halys Stal (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Mitt. Schweiz. Entomol. Ges. 2007, 81, 1. [Google Scholar]
- Zhou, Y.; Giusti, M.M.; Parker, J.; Salamanca, J.; Rodriguez-Saona, C. Frugivory by Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) Alters Blueberry Fruit Chemistry and Preference by Conspecifics. Environ. Entomol. 2016, 45, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Schumm, Z.R.; Alston, D.G.; Spears, L.R.; Manlove, K. Impact of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) Feeding on Tart Cherry (Rosales: Rosaceae) Quality and Yield in Utah. J. Econ. Entomol. 2020, 113, 2328–2334. [Google Scholar] [CrossRef] [PubMed]
- Mensah-Bonsu, M.; Dingha, B.N.; Jackai, L.E.N.; Adjei-Fremah, S.; Worku, M. Evaluation of preference of brown marmorated stink bug, Halyomorpha halys (Stål) for different colour bell peppers and the role of plant protein. Arthropod-Plant Interact. 2020, 14, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Heath, J.J.; Cipollini, D.F.; Stireman Iii, J.O. The role of carotenoids and their derivatives in mediating interactions between insects and their environment. Arthropod-Plant Interact. 2013, 7, 1–20. [Google Scholar] [CrossRef]
- Zobel, E.S.; Hooks, C.R.R.; Dively, G.P. Seasonal Abundance, Host Suitability, and Feeding Injury of the Brown Marmorated Stink Bug, Halyomorpha halys (Heteroptera: Penatomidae), in Selected Vegetables. J. Econ. Entomol. 2016, 109, 1289–1302. [Google Scholar] [CrossRef] [PubMed]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, Ecology, and Management of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, A1–A13. [Google Scholar] [CrossRef]
- Rokaya, P.; Baral, D.; Gautam, D.M.; Shrestha, A.; Paudyal, K. Effect of Postharvest Treatments on Quality and Shelf Life of Mandarin (Citrus reticulata Blanco). Am. J. Plant Sci. 2016, 7, 1098–1105. [Google Scholar] [CrossRef] [Green Version]
- Samira, A.; Woldetsadik, K.; Workneh, T.S. Postharvest quality and shelf life of some hot pepper varieties. J. Food Sci. Technol. 2013, 50, 842–855. [Google Scholar] [CrossRef] [Green Version]
- Hameed, R.; Malik, A.; Khan, A.S.; Imran, M.; Umar, M.; Riaz, R. Evaluating the effect of different storage conditions on quality of green chillies (Capsicum annuum L.). Trop. Agric. Res. 2015, 24, 391–399. [Google Scholar] [CrossRef]
- Lama, K.; Alkalai-Tuvia, S.; Chalupowicz, D.; Fallik, E. Extended Storage of Yellow Pepper Fruits at Suboptimal Temperatures May Alter Their Physical and Nutritional Quality. Agronomy 2020, 10, 1109. [Google Scholar] [CrossRef]
- Wiman, N.; Parker, J.; Rodriguez-Saona, C.; Walton, V. Characterizing Damage of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) in Blueberries. J. Econ. Entomol. 2015, 108, 1156–1163. [Google Scholar] [CrossRef]
- Rehman, F.; Khan, F.A.; Badruddin, S.M.A. Role of Phenolics in Plant Defense Against Insect Herbivory. In Chemistry of Phytopotentials: Health, Energy and Environmental Perspectives; Khemani, L.D., Srivastava, M.M., Srivastava, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 309–313. [Google Scholar]
- Li, B.; Yang, M.; Shi, R.; Ye, M. Insecticidal Activity of Natural Capsaicinoids Against Several Agricultural Insects. Nat. Prod. Commun. 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Bai, P.; Wei, L.; Kang, R.; Chen, L.; Zhang, M.; Tan, E.K.; Liu, W. Capsaicin Functions as Drosophila Ovipositional Repellent and Causes Intestinal Dysplasia. Sci. Rep. 2020, 10, 9963. [Google Scholar] [CrossRef]
- Soriano, I.R.; Asenstorfer, R.E.; Schmidt, O.; Riley, I.T. Inducible Flavone in Oats (Avena sativa) Is a Novel Defense Against Plant-Parasitic Nematodes. Phytopathology 2004, 94, 1207–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Othman, Z.A.A.; Ahmed, Y.B.H.; Habila, M.A.; Ghafar, A.A. Determination of Capsaicin and Dihydrocapsaicin in Capsicum Fruit Samples using High Performance Liquid Chromatography. Molecules 2011, 16, 8919–8929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, Z.; Daood, H.; Ambrózy, Z.; Helyes, L. Determination of Polyphenols, Capsaicinoids, and Vitamin C in New Hybrids of Chili Peppers. J. Anal. Methods Chem. 2015, 2015, 102125. [Google Scholar] [CrossRef] [Green Version]
- Ministry of Agriculture, Forestry and Food. Integrated Production Guidelines 2021. Available online: https://www.gov.si/en/state-authorities/ministries/ministry-of-agriculture-forestry-and-food/ (accessed on 22 October 2020).
- Zamljen, T.; Jakopič, J.; Hudina, M.; Veberič, R.; Slatnar, A. Influence of intra and inter species variation in chilies (Capsicum spp.) on metabolite composition of three fruit segments. Sci. Rep. 2021, 11, 4932. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Medic, A.; Jakopic, J.; Hudina, M.; Solar, A.; Veberic, R. Identification and quantification of the major phenolic constituents in Juglans regia L. peeled kernels and pellicles, using HPLC–MS/MS. Food Chem. 2021, 352, 129404. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Schmitzer, V.; Jakopic, J.; Cunja, V.; Veberic, R.; Munda, A.; Stampar, F. Phenolic compounds as defence response of pepper fruits to Colletotrichum coccodes. Physiol. Mol. Plant Pathol. 2013, 84, 138–145. [Google Scholar] [CrossRef]
- Sagun, S.; Collins, E.; Martin, C.; Nolan, E.J.; Horzempa, J. Alarm Odor Compounds of the Brown Marmorated Stink Bug Exhibit Antibacterial Activity. J. Pharmacogn. Nat. Prod. 2016, 2, 119. [Google Scholar] [CrossRef] [Green Version]
- Esquivel, J.F. Stink bug rostrum length vs. stylet penetration potential. Entomol. Exp. Appl. 2019, 167, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Kissinger, M.; Tuvia-Alkalai, S.; Shalom, Y.; Fallik, E.; Elkind, Y.; Jenks, M.; Goodwin, M. Characterization of Physiological and Biochemical Factors Associated with Postharvest Water Loss in Ripe Pepper Fruit during Storage. J. Am. Soc. Hortic. Sci. 2005, 130, 735–741. [Google Scholar] [CrossRef] [Green Version]
- Tano, K.; Kamenan, A.; Arul, J. Respiration and transpiration characteristics of selected fresh fruits and vegetables. Agron. Afr. 2009, 17, 103–115. [Google Scholar] [CrossRef]
- Morkunas, I.; Ratajczak, L. The role of sugar signaling in plant defense responses against fungal pathogens. Acta Physiol. Plant. 2014, 36, 1607–1619. [Google Scholar] [CrossRef] [Green Version]
- Wind, J.; Smeekens, S.; Hanson, J. Sucrose: Metabolite and signaling molecule. Phytochemistry 2010, 71, 1610–1614. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Dias, L.; Vilanova, S.; Fita, A.; Prohens, J.; Rodríguez-Burruezo, A. Genetic diversity, population structure, and relationships in a collection of pepper (Capsicum spp.) landraces from the Spanish centre of diversity revealed by genotyping-by-sequencing (GBS). Hortic. Res. 2019, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- Marchiosi, R.; dos Santos, W.D.; Constantin, R.P.; de Lima, R.B.; Soares, A.R.; Finger-Teixeira, A.; Mota, T.R.; de Oliveira, D.M.; Foletto-Felipe, M.d.P.; Abrahão, J. Biosynthesis and metabolic actions of simple phenolic acids in plants. Phytochem. Rev. 2020, 19, 865–906. [Google Scholar] [CrossRef]
- Antoniali, S.; Leal, P.A.M.; Magalhães, A.M.d.; Fuziki, R.T.; Sanches, J. Physico-chemical characterization of ‘Zarco HS’ yellow bell pepper for different ripeness stages. Sci. Agric. 2007, 64, 19–22. [Google Scholar] [CrossRef]
- Araujo, W.L.; Nunes-Nesi, A.; Nikoloski, Z.; Sweetlove, L.J.; Fernie, A.R. Metabolic control and regulation of the tricarboxylic acid cycle in photosynthetic and heterotrophic plant tissues. Plant Cell Environ. 2012, 35, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zushi, K.; Matsuzoe, N. Postharvest Changes in Sugar, Organic Acid, Glutamic Acid and Antioxidant Contents in Tomato Fruit Grown under Salinity Stress. Environ. Control Biol. 2006, 44, 111–117. [Google Scholar] [CrossRef]
- Leiss, K.A.; Maltese, F.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G.L. Identification of chlorogenic acid as a resistance factor for thrips in chrysanthemum. Plant Physiol. 2009, 150, 1567–1575. [Google Scholar] [CrossRef] [Green Version]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Yactayo-Chang, J.P.; Tang, H.V.; Mendoza, J.; Christensen, S.A.; Block, A.K. Plant Defense Chemicals against Insect Pests. Agronomy 2020, 10, 1156. [Google Scholar] [CrossRef]
- Reyes-Escogido, M.D.L.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and pharmacological aspects of capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aley, J.P.; Adams, N.J.; Ladyman, R.J.; Fraser, D.L. The efficacy of capsaicin as an equine repellent for chewing wood. J. Vet. Behav. 2015, 10, 243–247. [Google Scholar] [CrossRef]
- Cowles, R.S.; Keller, J.E.; Miller, J.R. Pungent spices, ground red pepper, and synthetic capsaicin as onion fly ovipositional deterrents. J. Chem. Ecol. 1989, 15, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Olszewska, J.; Tęgowska, E. Opposite effect of capsaicin and capsazepine on behavioral thermoregulation in insects. J. Comp. Physiol. A 2011, 197, 1021–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maliszewska, J.; Marcinkowska, S.; Nowakowska, A.; Kletkiewicz, H.; Rogalska, J. Altered heat nociception in cockroach Periplaneta americana L. exposed to capsaicin. PLoS ONE 2018, 13, e0194109. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Before Storage | After Storage | ||||||
|---|---|---|---|---|---|---|---|
| Control | H. halys | Control 2 | Control | H. halys | Control 2 | ||
| Pungent ‘Eris F1’ | Pericarp | 87.2 ± 1.0 a ꜞ | 84.5 ± 1.3 b | 84.2 ± 1.2 b | 86.4 ± 1.2 a | 83.7 ± 1.3 b | 83.8 ± 1.2 b |
| Placenta | 89.4 ± 2.1 | 86.9 ± 1.7 | 88.4 ± 1.9 | 85.0 ± 1.0 | 84.8 ± 1.1 | 86.2 ± 0.9 | |
| Non-pungent ‘Lombardo tago’ | Pericarp | 87.9 ± 1.7 | 86.2 ± 1.4 | 86.6 ± 1.9 | 87.0 ± 1.2 | 86.0 ± 1.2 | 86.2 ± 1.4 |
| Placenta | 89.4 ± 2.2 | 87.7 ± 2.0 | 86.5 ± 2.4 | 87.4 ± 0.9 | 85.8 ± 1.0 | 85.9 ± 1.0 | |
| Before Storage | After Storage | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | H. halys | Control 2 | Control | H. halys | Control 2 | |||
| Pungent ‘Eris F1′ | Pericarp | Sucrose | 118.2 ± 4.3 | 126.3 ± 8.4 | 81.0 ± 9.3 | 98.1 ± 6.7 | 129.2 ± 16.9 | 96.0 ± 5.9 |
| Glucose | 224.5 ± 8.4 a ꜞ | 215.1 ± 17.5 a | 141.4 ± 8.8 b | 150.2 ± 4.7 b | 232.1 ± 19.0 a | 177.5 ± 10.5 a | ||
| Fructose | 285.2 ± 14.2 | 249.3 ± 18.8 | 212.2 ± 11.2 | 207.2 ± 6.3 | 269.3 ± 24.6 | 224.3 ± 7.3 | ||
| Total sugars | 627.9 ± 31.7 a | 590.7 ± 25.1 a | 439.6 ± 14.4 b | 455.5 ± 21.7 b | 634.6 ± 51.3 a | 497.8 ± 41.6 b | ||
| Placenta | Sucrose | 143.4 ± 5.0 | 124.3 ± 31.0 | 73.2 ± 12.2 | 76.0 ± 10.2 | 109.0 ± 9.6 | 89.0 ± 8.2 | |
| Glucose | 222.3 ± 8.4 | 160.0 ± 21.8 | 68.6 ± 8.9 | 83.5 ± 14.7 | 190.4 ± 21.2 | 98.0 ± 12.7 | ||
| Fructose | 333.1 ± 10.2 a | 222.1 ± 34.2 b | 141.0 ± 11.7 c | 136.2 ± 18.4 b | 267.2 ± 30.5 a | 154.1 ± 11.5 b | ||
| Total sugars | 698.8 ± 52.7 a | 506.4 ± 24.5 b | 282.8 ± 19.4 c | 295.7 ± 25.8 b | 566.6 ± 42.6 a | 341.2 ± 36.2 b | ||
| Non-pungent ‘Lombardo tago’ | Pericarp | Sucrose | 68.0 ± 2.0 | 75.5 ± 6.8 | 77.1 ± 6.2 | 55.3 ± 14.4 | 81.3 ± 5.3 | 66.2 ± 5.4 |
| Glucose | 265.5 ± 5.4 a | 228.3 ± 13.0 b | 261.6 ± 12.7 a | 152.1 ± 29.1 b | 277.1 ± 19.6 a | 172.1 ± 20.2 b | ||
| Fructose | 351.0 ± 8.7 | 332.1 ± 16.5 | 372.2 ± 11.5 | 229.3 ± 47.4 b | 366.5 ± 14.1 a | 257.1 ± 33.4 b | ||
| Total sugars | 684.5 ± 36.0 a | 635.9 ± 52.3 b | 710.9 ± 55.3 a | 436.7 ± 24.2 b | 724.9 ± 63.4 a | 495.4 ± 32.8 b | ||
| Placenta | Sucrose | 95.4 ± 6.7 a | 88.2 ± 8.6 a | 52.3 ± 5.6 b | 49.0 ± 2.4b | 78.0 ± 6.2a | 86.1 ± 12.7a | |
| Glucose | 194.5 ± 13.2 a | 125.5 ± 5.2 b | 127.5 ± 12.2 b | 104.2 ± 6.7 | 118.2 ± 20.9 | 99.3 ± 11.6 | ||
| Fructose | 323.0 ± 21.6 a | 199.1 ± 7.5 b | 232.1 ± 19.1 a | 176.1 ± 24.8 | 185.2 ± 26.4 | 174.4 ± 8.6 | ||
| Total sugars | 617.9 ± 52.9 a | 412.8 ± 19.8 b | 411.9 ± 24.3 b | 329.3 ± 37.0 | 381.4 ± 32.8 | 359.8 ± 28.0 | ||
| Before Storage | After Storage | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | H. halys | Control 2 | Control | H. halys | Control 2 | |||
| Pungent ‘Eris F1′ | Pericarp | Oxalic acid | / | / | / | / | / | / |
| Citric acid | 46.5 ± 2.4 | 45.4 ± 2.4 | 47.7 ± 8.2 | 23.9 ± 1.7 b | 27.2 ± 1.3 ab | 31.6 ± 2.3 a | ||
| Malic acid | 14.2 ± 0.6 | 13.3 ± 0.5 | 14.5 ± 2.6 | 17.4 ± 1.3 b | 26.6 ± 3.4 a | 17.2 ± 1.2 b | ||
| Qunic acid | 21.6 ± 2.2 | 15.6 ± 1.2 | 14.3 ± 2.2 | 16.6 ± 1.5 | 13.4 ± 1.3 | 14.4 ± 1.5 | ||
| Succinic acid | 5.2 ± 0.5 | 5.0 ± 0.8 | 5.8 ± 0.5 | 5.0 ± 0.2 | 5.4 ± 0.5 | 6.0 ± 0.3 | ||
| Fumaric acid ♦ | 42.4 ± 3.8 | 46.6 ± 3. 5 | 45.2 ± 9.3 | 52.4 ± 7.6 b | 110.2 ± 2.3 a | 63.4 ± 6.8 b | ||
| Ascorbic acid | 35.2 ± 4.3 | 33.5 ± 5.3 | 44.5 ± 3.6 | 23.4 ± 1.4 | 19.2 ± 1.3 | 19.2 ± 1.6 | ||
| Total acids | 122.7 ±13.6 | 111.2 ±13.5 | 125.3 ±13.7 | 85.7 ±5.3 b | 91.2 ±4.6 a | 88.5 ±2.3 b | ||
| Placenta | Oxalic acid ♦ | 217.2 ± 36.3 | 148.6 ± 9.6 | 211.8 ± 10.2 | 468.4 ± 68.2 | 419.6 ± 5.6 | 454.4 ± 2.5 | |
| Citric acid | 67.6 ± 2.7 a ꜞ | 46.2 ± 6.8 b | 45.4 ± 3.3 b | 36.5 ± 4.6 | 32.3 ± 2.7 | 29.3 ± 3.5 | ||
| Malic acid | 26.3 ± 1.3 | 20.7 ± 3.6 | 21.5 ± 3.8 | 41.7 ± 6.5 | 42.7 ± 8.2 | 31.6 ± 4.2 | ||
| Qunic acid | 12.2 ± 0.8 | 9.9 ± 1.3 | 9.7 ± 1.7 | 12.3 ± 1.4 | 12.9 ± 0.3 | 11.5 ± 2.6 | ||
| Succinic acid | 6.7 ± 0.2 | 5.4 ± 0.5 | 6.2 ± 1.7 | 6.5 ± 0.2 | 5.7 ± 0.5 | 6.2 ± 0.2 | ||
| Fumaric acid ♦ | 70.4 ± 4.6 b | 114.5 ± 13.3 a | 136.7 ± 13.6 a | 125.4 ± 9.8 | 204.2 ± 23.3 | 215.8 ± 8.6 | ||
| Ascorbic acid | 20.3 ± 1.3 a | 11.6 ± 1.8 b | 7.7 ± 1.5 b | 12.2 ± 1.3 | 7.4 ± 1.6b | 8.1 ± 1.3b | ||
| Total acids | 131.2 ±10.6 a | 92.4 ±4.3 b | 90.5 ±5.6 b | 109.6 ±9.5 a | 100.2 ±3.8 a | 87.5 ±2.2 a | ||
| Non-pungent ‘Lombardo tago’ | Pericarp | Oxalic acid | / | / | / | / | / | / |
| Citric acid | 36.1 ± 1.2 | 42.4 ± 2.4 | 40.5 ± 2.3 | 34.2 ± 4.4 | 35.2 ± 1.2 | 27.3 ± 6.2 | ||
| Malic acid | 16.4 ± 1.5 | 17.3 ± 1.6 | 17.6 ± 0.6 | 25.4 ± 2.2 b | 35.8 ± 2.6 a | 21.5 ± 5.5 b | ||
| Qunic acid | 15.2 ± 0.2b | 17.5 ± 1.1a | 12.2 ± 1.5b | 45.5 ± 3.6 a | 26.3 ± 3.4 b | 22.8 ± 2.6 b | ||
| Succinic acid | 4.0 ± 0.2 | 5.0 ± 0.4 | 5.1 ± 0.2 | 5.2 ± 0.1 | 5.7 ± 0.4 | 5.2 ± 0.3 | ||
| Fumaric acid ♦ | 34.2 ± 1.6 b | 89.5 ± 8.2 a | 86.8 ± 7.3 a | 63.8 ± 7.7 b | 100. 5± 14.2 a | 71.9 ± 6.2 b | ||
| Ascorbic acid | 30.5 ± 1.7 | 31.2 ± 1.5 | 33.4 ± 0.3 | 26.2 ± 0.6 | 23.2 ± 1.2 | 25.4 ± 1.4 | ||
| Total acids | 102.4 ±5.4 b | 112.6± 10.1 a | 107.2 ±9.0 b | 135.7 ±11.2 | 125.1 ±10.6 | 101.4 ±8.0 | ||
| Placenta | Oxalic acid ♦ | 103.4 ± 10.6 | 84.7 ± 2.8 | 70.6 ± 1.0 | 320. 4± 10.0 | 540.6 ± 9.3 | 439.3 ± 8.0 | |
| Citric acid | 40.3 ± 3.3 | 37.5 ± 3.4 | 32.8 ± 4.2 | 34.2 ± 5.3 | 36.5 ± 2.4 | 31.2 ± 2.2 | ||
| Malic acid | 23.6 ± 2.5 | 32.3 ± 7.2 | 19.2 ± 3.5 | 30.5 ± 2.5 | 40.2 ± 6.7 | 22.6 ± 4.5 | ||
| Qunic acid | 7.3 ± 1.8 | 5.4 ± 1.9 | 5.4 ± 1.3 | 8.1 ± 3.7 | 17.4 ± 2.3 | 15.2 ± 3.7 | ||
| Succinic acid | 5.8 ± 0.2 | 5.6 ± 0.3 | 5.1 ± 0.2 | 5.2 ± 0.4 | 5.2 ± 0.3 | 6.4 ± 0.4 | ||
| Fumaric acid ♦ | 71.5 ± 0.4 b | 152.3 ± 17.4 a | 128.6 ± 3.3 a | 172.3 ± 5.8 | 162.5 ± 12.7 | 198.6 ± 5.4 | ||
| Ascorbic acid | 15.4 ± 0.7 a | 11.1 ± 0.7 b | 11.3 ± 1.4 b | 14.4 ± 0.3 a | 10.5 ± 0.1 b | 11.2 ± 1.0 b | ||
| Total acids | 90.8 ±4.3 | 91.9 ±6.5 | 71.3 ±2.6 | 92.4 ±4.3 b | 109.5 ±3.7 a | 86.2 ±5.6 b | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zamljen, T.; Veberic, R.; Hudina, M.; Slatnar, A. The Brown Marmorated Stink Bug (Halyomorpha halys Stål.) Influences Pungent and Non-Pungent Capsicum Cultivars’ Pre- and Post-Harvest Quality. Agronomy 2021, 11, 2252. https://doi.org/10.3390/agronomy11112252
Zamljen T, Veberic R, Hudina M, Slatnar A. The Brown Marmorated Stink Bug (Halyomorpha halys Stål.) Influences Pungent and Non-Pungent Capsicum Cultivars’ Pre- and Post-Harvest Quality. Agronomy. 2021; 11(11):2252. https://doi.org/10.3390/agronomy11112252
Chicago/Turabian StyleZamljen, Tilen, Robert Veberic, Metka Hudina, and Ana Slatnar. 2021. "The Brown Marmorated Stink Bug (Halyomorpha halys Stål.) Influences Pungent and Non-Pungent Capsicum Cultivars’ Pre- and Post-Harvest Quality" Agronomy 11, no. 11: 2252. https://doi.org/10.3390/agronomy11112252