Escherichia coli as Commensal and Pathogenic Bacteria among Food-Producing Animals: Health Implications of Extended Spectrum β-Lactamase (ESBL) Production


 , , , , and
, , , , and 
Abstract
:Simple Summary
Abstract
1. Pathogenicity of Escherichia coli
2. Antimicrobial Resistance Trends in E. coli
3. The Major Threat from ESBL-Producing E. coli
3.1. Contextualizing the Issue
3.2. CTX-M β-Lactamases and Their Relevance
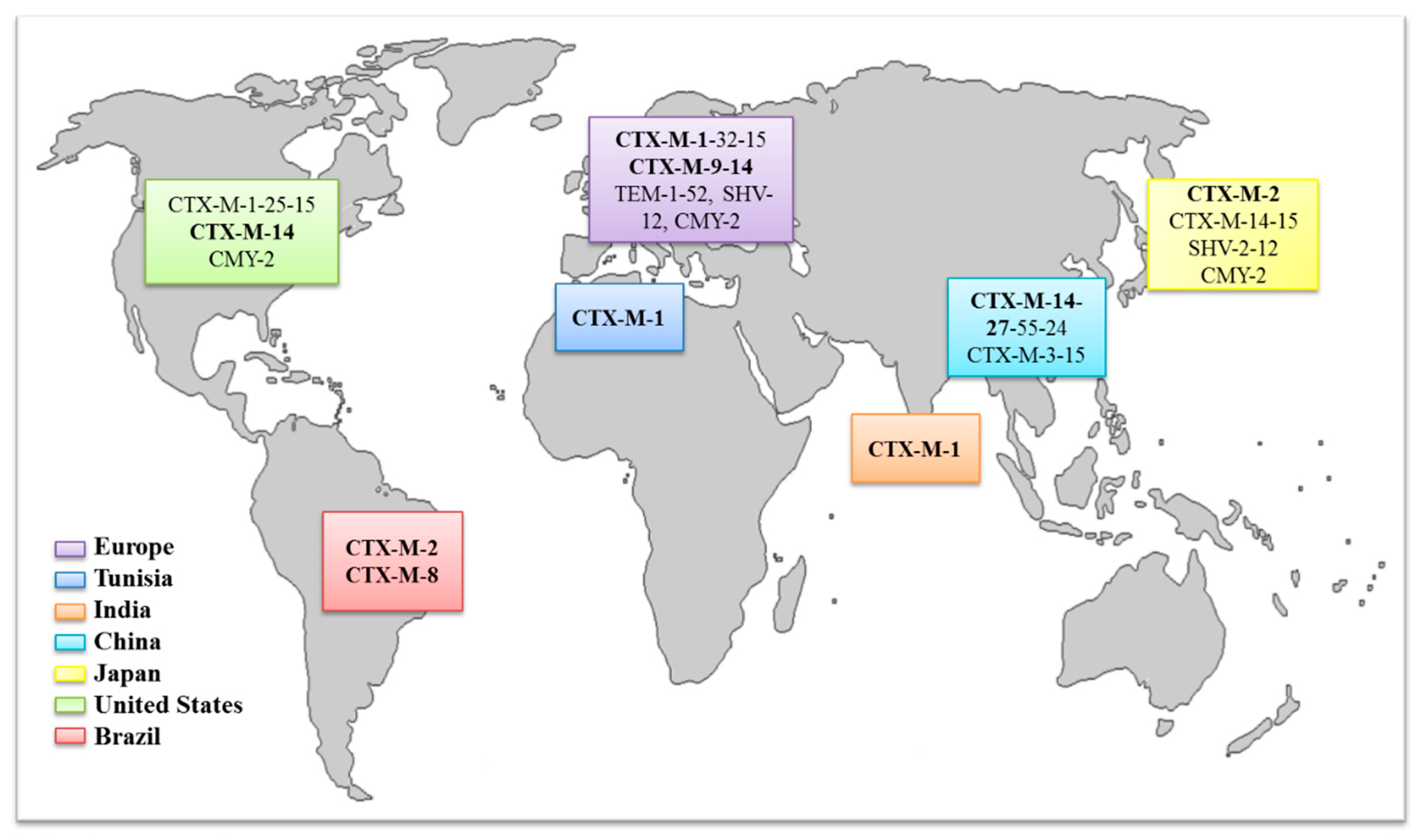
3.3. Dissemination of CTX-M and Its Implications
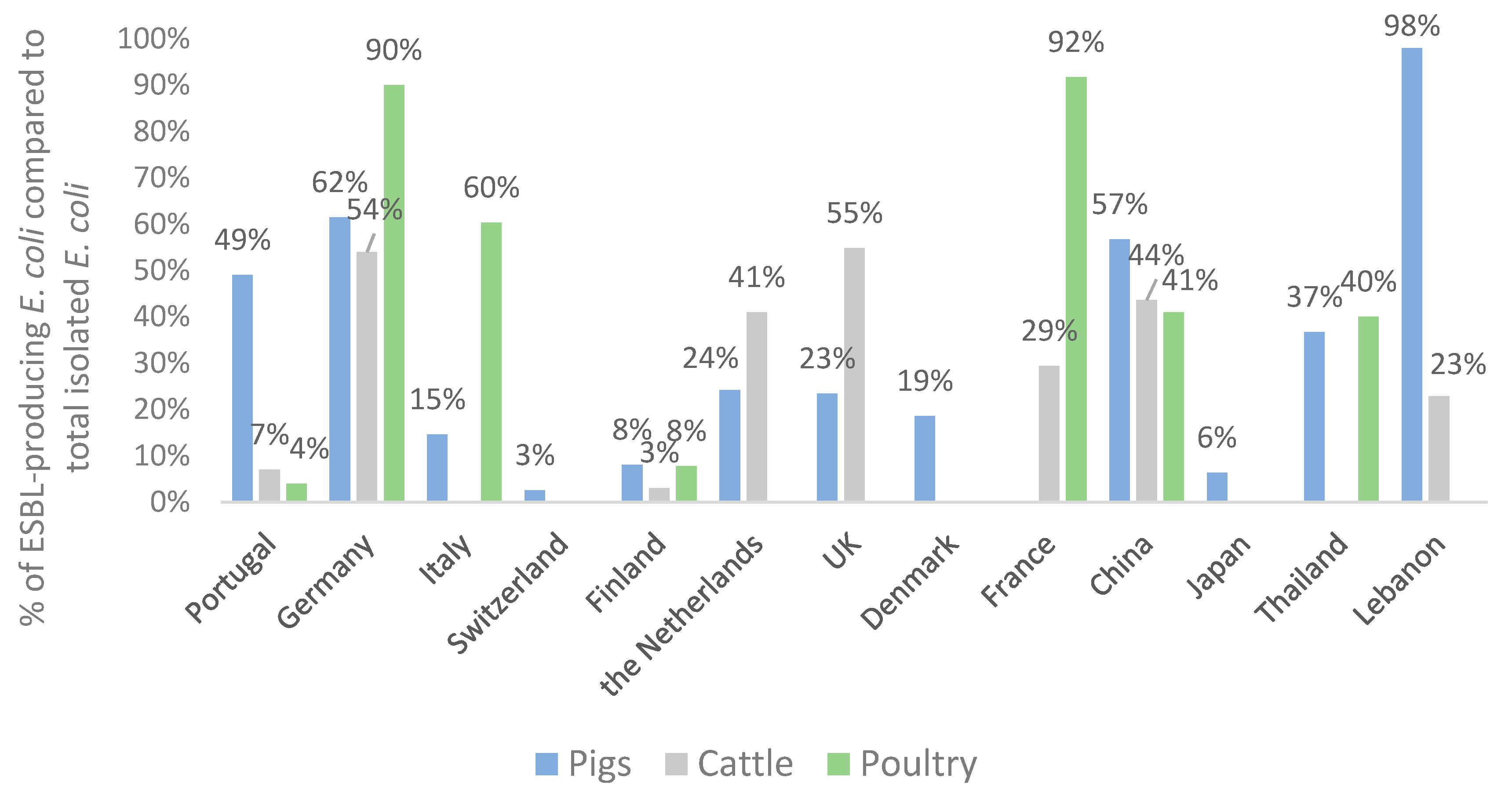
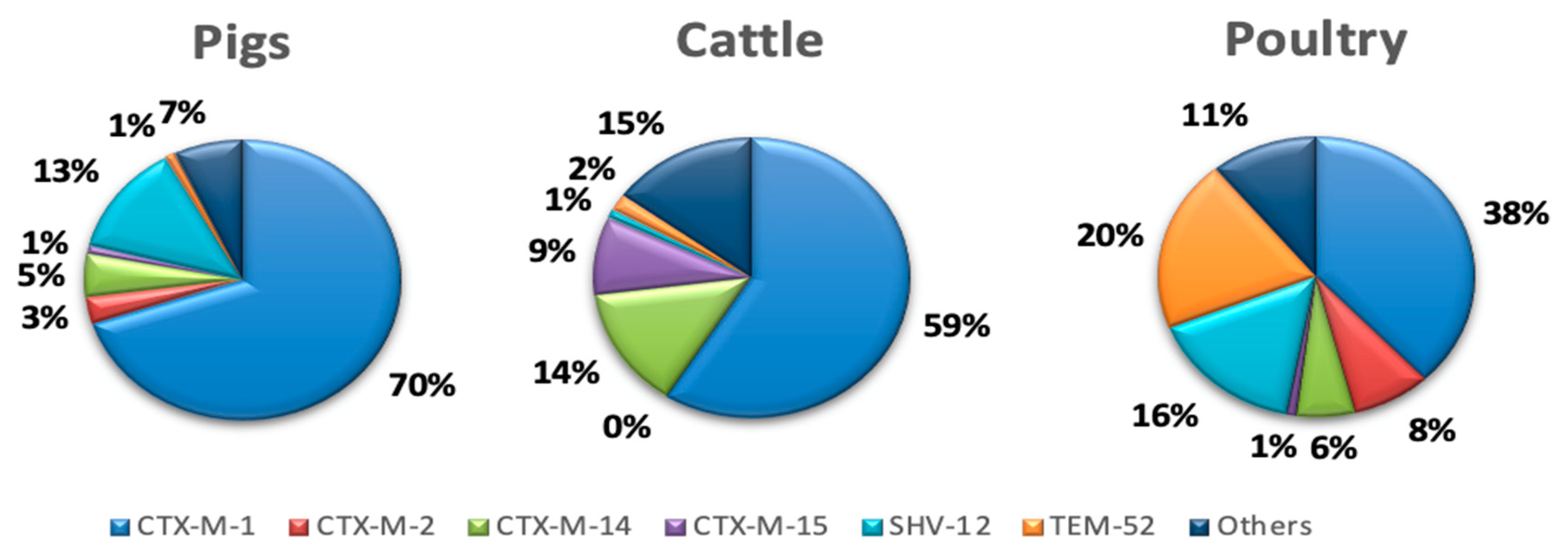
3.4. Distribution of ESBL Enzymes in E. coli Isolates from Food-Producing Animals
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lindstedt, B.-A.; Finton, M.D.; Porcellato, D.; Brandal, L.T. High frequency of hybrid Escherichia coli strains with combined Intestinal Pathogenic Escherichia coli (IPEC) and Extraintestinal Pathogenic Escherichia coli (ExPEC) virulence factors isolated from human faecal samples. BMC Infect. Dis. 2018, 18, 544. [Google Scholar] [CrossRef] [PubMed]
- Alizade, H.; Hosseini Teshnizi, S.; Azad, M.; Shojae, S.; Gouklani, H.; Davoodian, P.; Ghanbarpour, R. An overview of diarrheagenic Escherichia coli in Iran: A systematic review and meta-analysis. J. Res. Med. Sci. 2019, 24, 23. [Google Scholar] [PubMed]
- Basak Unver, K.; Ahmet, K. Vero-Toxigenic Escherichia coli (VTEC): To sum up all we know. J. Gastroenterol. Res. 2017, 1, 14–23. [Google Scholar] [CrossRef]
- Nileshkumar, V.; Hati, S.; Gawai, K.; Sreeja, V. Food borne pathogens: A threat to dairy industry. Res. Rev. J. Dairy Sci. Technol. 2015, 4, 28–36. [Google Scholar]
- Nobili, G.; Franconieri, I.; La Bella, G.; Basanisi, M.G.; La Salandra, G. Prevalence of Verocytotoxigenic Escherichia coli strains isolated from raw beef in southern Italy. Int. J. Food Microbiol. 2017, 257, 201–205. [Google Scholar] [CrossRef]
- Newell, D.G.; Koopmans, M.; Verhoef, L.; Duizer, E.; Aidara-Kane, A.; Sprong, H.; Opsteegh, M.; Langelaar, M.; Threfall, J.; Scheutz, F.; et al. Food-borne diseases—The challenges of 20 years ago still persist while new ones continue to emerge. Int. J. Food Microbiol. 2010, 139, S3–S15. [Google Scholar] [CrossRef]
- Nielsen, D.W.; Ricker, N.; Barbieri, N.L.; Allen, H.K.; Nolan, L.K.; Logue, C.M. Outer membrane protein A (OmpA) of extraintestinal pathogenic Escherichia coli. BMC Res. Notes 2020, 13, 51. [Google Scholar] [CrossRef]
- Stromberg, Z.R.; Johnson, J.R.; Fairbrother, J.M.; Kilbourne, J.; Van Goor, A.; Curtiss Rd, R.; Mellata, M. Evaluation of Escherichia coli isolates from healthy chickens to determine their potential risk to poultry and human health. PLoS ONE 2017, 12, e0180599. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R.; Johnson, J.R. Food-borne origins of Escherichia coli causing extraintestinal infections. Clin. Infect. Dis. 2012, 55, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R. Escherichia coli and urinary tract infections: The role of poultry-meat. Clin. Microbiol. Infect. 2016, 22, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R.; Johnson, J.R. Reservoirs of extraintestinal pathogenic Escherichia coli. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Meniño, I.; García, V.; Mora, A.; Díaz-Jiménez, D.; Flament-Simon, S.C.; Alonso, M.P.; Blanco, J.E.; Blanco, M.; Blanco, J. Swine enteric colibacillosis in Spain: Pathogenic potential of mcr-1 ST10 and ST131 E. coli isolates. Front. Microbiol. 2018, 9, 2659. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Stephan, R.; Nüesch-Inderbinen, M. Distribution of virulence factors in ESBL-producing Escherichia coli isolated from the environment, livestock, food and humans. Sci. Total Environ. 2016, 541, 667–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eibach, D.; Dekker, D.; Gyau Boahen, K.; Wiafe Akenten, C.; Sarpong, N.; Belmar Campos, C.; Berneking, L.; Aepfelbacher, M.; Krumkamp, R.; Owusu-Dabo, E.; et al. Extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae in local and imported poultry meat in Ghana. Vet. Microbiol. 2018, 217, 7–12. [Google Scholar] [CrossRef]
- Chakraborty, A.; Saralaya, V.; Adhikari, P.; Shenoy, S.; Baliga, S.; Hegde, A. Characterization of Escherichia coli phylogenetic groups associated with extraintestinal infections in South Indian population. Ann. Med. Health Sci. Res. 2015, 5, 241–246. [Google Scholar]
- Koga, V.L.; Tomazetto, G.; Cyoia, P.S.; Neves, M.S.; Vidotto, M.C.; Nakazato, G.; Kobayashi, R.K.T. Molecular screening of virulence genes in extraintestinal pathogenic Escherichia coli isolated from human blood culture in Brazil. Biomed. Res. Int. 2014, 2014, 465054. [Google Scholar] [CrossRef] [Green Version]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Raimondi, S.; Righini, L.; Candeliere, F.; Musmeci, E.; Bonvicini, F.; Gentilomi, G.; Erjavec, M.S.; Amaretti, A.; Rossi, M. Antibiotic resistance, virulence factors, phenotyping, and genotyping of E. coli isolated from the feces of healthy subjects. Microorganisms 2019, 7, 251. [Google Scholar] [CrossRef] [Green Version]
- Nyirabahizi, E.; Tyson, G.H.; Dessai, U.; Zhao, S.; Kabera, C.; Crarey, E.; Womack, N.; Crews, M.K.; Strain, E.; Tate, H. Evaluation of Escherichia coli as an indicator for antimicrobial resistance in Salmonella recovered from the same food or animal ceca samples. Food Control 2020, 115, 107280. [Google Scholar] [CrossRef]
- ECDC. Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-Net) 2017; ECDC: Stockholm, Sweden, 2018. [Google Scholar]
- EFSA. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J. 2019, 17, 5598. [Google Scholar]
- Niero, G.; Bortolaia, V.; Vanni, M.; Intorre, L.; Guardabassi, L.; Piccirillo, A. High diversity of genes and plasmids encoding resistance to third-generation cephalosporins and quinolones in clinical Escherichia coli from commercial poultry flocks in Italy. Vet. Microbiol. 2018, 216, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Hille, K.; Ruddat, I.; Schmid, A.; Hering, J.; Hartmann, M.; von Münchhausen, C.; Schneider, B.; Messelhäusser, U.; Friese, A.; Mansfeld, R.; et al. Cefotaxime-resistant E. coli in dairy and beef cattle farms—Joint analyses of two cross-sectional investigations in Germany. Prev. Vet. Med. 2017, 142, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.P.; Nguyen, T.A.D.; Le, T.H.; Tran, N.M.D.; Ngo, T.P.; Dang, V.C.; Kawai, T.; Kanki, M.; Kawahara, R.; Jinnai, M.; et al. Dissemination of extended-spectrum β-lactamase- and AmpC β-Lactamase-producing Escherichia coli within the food distribution system of Ho Chi Minh City, Vietnam. Biomed. Res. Int. 2016, 2016, 8182096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, K.W.; Lee, Y.J. Characterization of plasmid mediated quinolone resistance determinants in ciprofloxacin resistant-Escherichia coli from chicken meat produced by integrated broiler operations in Korea. Int. J. Food Microbiol. 2019, 307, 108274. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, E.; Van Meervenne, E.; Boon, N.; Van de Wiele, T.; Wattiau, P.; Herman, L.; Heyndrickx, M.; Van Coillie, E. Characterization of cefotaxime- and ciprofloxacin-resistant commensal Escherichia coli originating from Belgian farm animals indicates high antibiotic resistance transfer rates. Microb. Drug Resist. 2017, 24, 707–717. [Google Scholar] [CrossRef]
- Wasyl, D.; Hoszowski, A.; Zając, M.; Szulowski, K. Antimicrobial resistance in commensal Escherichia coli isolated from animals at slaughter. Front. Microbiol. 2013, 4, 221. [Google Scholar] [CrossRef] [Green Version]
- Cummings, K.J.; Aprea, V.A.; Altier, C. Antimicrobial resistance trends among Escherichia coli isolates obtained from dairy cattle in the Northeastern United States, 2004–2011. Foodborne Pathog. Dis. 2013, 11, 61–67. [Google Scholar] [CrossRef]
- Myrenås, M.; Slettemeås, J.S.; Thorsteinsdottir, T.R.; Bengtsson, B.; Börjesson, S.; Nilsson, O.; Landén, A.; Sunde, M. Clonal spread of Escherichia coli resistant to cephalosporins and quinolones in the Nordic broiler production. Vet. Microbiol. 2018, 213, 123–128. [Google Scholar] [CrossRef]
- Hang, B.P.T.; Wredle, E.; Börjesson, S.; Sjaunja, K.S.; Dicksved, J.; Duse, A. High level of multidrug-resistant Escherichia coli in young dairy calves in southern Vietnam. Trop. Anim. Health Prod. 2019, 51, 1405–1411. [Google Scholar] [CrossRef]
- Callens, B.; Cargnel, M.; Sarrazin, S.; Dewulf, J.; Hoet, B.; Vermeersch, K.; Wattiau, P.; Welby, S. Associations between a decreased veterinary antimicrobial use and resistance in commensal Escherichia coli from Belgian livestock species (2011–2015). Prev. Vet. Med. 2018, 157, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Belaynehe, K.M.; Shin, S.W.; Hong-Tae, P.; Yoo, H.S. Occurrence of aminoglycoside-modifying enzymes among isolates of Escherichia coli exhibiting high levels of aminoglycoside resistance isolated from Korean cattle farms. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slettemeås, J.S.; Sunde, M.; Ulstad, C.R.; Norström, M.; Wester, A.L.; Urdahl, A.M. Occurrence and characterization of quinolone resistant Escherichia coli from Norwegian turkey meat and complete sequence of an IncX1 plasmid encoding qnrS1. PLoS ONE 2019, 14, e0212936. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.A.E.E.; Zhong, L.-L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.-B. Colistin and its role in the era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrijver, R.; Stijntjes, M.; Rodríguez-Baño, J.; Tacconelli, E.; Babu Rajendran, N.; Voss, A. Review of antimicrobial resistance surveillance programmes in livestock and meat in EU with focus on humans. Clin. Microbiol. Infect. 2018, 24, 577–590. [Google Scholar] [CrossRef] [Green Version]
- Azam, H.; Ghezeljeh, S.M.; Mahmoud, S. Prevalence of class 1 and 2 integrons among the multidrug resistant uropathogenic strains of Escherichia coli. Asian Biomed. 2017, 9, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Wyrsch, E.R.; Reid, C.J.; DeMaere, M.Z.; Liu, M.Y.; Chapman, T.A.; Roy Chowdhury, P.; Djordjevic, S.P. Complete sequences of multiple-drug resistant IncHI2 ST3 plasmids in Escherichia coli of porcine origin in Australia. Front. Sustain. Food Syst. 2019, 3, 18. [Google Scholar] [CrossRef]
- Mbelle, N.M.; Feldman, C.; Osei Sekyere, J.; Maningi, N.E.; Modipane, L.; Essack, S.Y. The resistome, mobilome, virulome and phylogenomics of multidrug-resistant Escherichia coli clinical isolates from Pretoria, South Africa. Sci. Rep. 2019, 9, 16457. [Google Scholar] [CrossRef]
- Reid, C.J.; Wyrsch, E.R.; Roy Chowdhury, P.; Zingali, T.; Liu, M.; Darling, A.E.; Chapman, T.A.; Djordjevic, S.P. Porcine commensal Escherichia coli: A reservoir for class 1 integrons associated with IS26. Microb. Genom. 2017, 3, e000143. [Google Scholar] [CrossRef]
- Falagas, M.E.; Karageorgopoulos, D.E. Extended-spectrum beta-lactamase-producing organisms. J. Hosp. Infect. 2009, 73, 345–354. [Google Scholar] [CrossRef]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzshcmar, M.E.; Devlesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Schwaber, M.J.; Carmeli, Y. Mortality and delay in effective therapy associated with extended-spectrum beta-lactamase production in Enterobacteriaceae bacteraemia: A systematic review and meta-analysis. J. Antimicrob. Chemother. 2007, 60, 913–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mughini-Gras, L.; Dorado-García, A.; van Duijkeren, E.; van den Bunt, G.; Dierikx, C.M.; Bonten, M.J.M.; Bootsma, M.C.J.; Schmitt, H.; Hald, T.; Evers, E.G.; et al. Attributable sources of community-acquired carriage of Escherichia coli containing β-lactam antibiotic resistance genes: A population-based modelling study. Lancet Planet Health 2019, 3, e357–e369. [Google Scholar]
- Cantón, R.; Novais, A.; Valverde, A.; Machado, E.; Peixe, L.; Baquero, F.; Coque, T.M. Prevalence and spread of extended-spectrum β-lactamase-producing Enterobacteriaceae in Europe. Clin. Microbiol. Infect. 2008, 14, 144–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezabih, Y.M.; Sabiiti, W.; Alamneh, E.; Bezabih, A.; Peterson, G.M.; Bezabhe, W.M.; Roujeinikova, A. The global prevalence and trend of human intestinal carriage of ESBL-producing Escherichia coli in the community. J. Antimicrob. Chemother. 2020, dkaa399. [Google Scholar] [CrossRef]
- Mueller, L.; Cimen, C.; Poirel, L.; Descombes, M.-C.; Nordmann, P. Prevalence of fosfomycin resistance among ESBL-producing Escherichia coli isolates in the community, Switzerland. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 945–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, M.; Anjum, M.; Andersson, D.I.; Sandegren, L. Combinations of mutations in envZ, ftsI, mrdA, acrB and acrR can cause high-level carbapenem resistance in Escherichia coli. J. Antimicrob. Chemother. 2016, 71, 1188–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, D.L.; Bonomo, R.A. Extended-spectrum β-lactamases: A clinical update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [Green Version]
- Sawa, T.; Kooguchi, K.; Moriyama, K. Molecular diversity of extended-spectrum β-lactamases and carbapenemases, and antimicrobial resistance. J. Intensive Care 2020, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Rawat, D.; Nair, D. Extended-spectrum β-lactamases in Gram negative bacteria. J. Glob. Infect. Dis. 2010, 2, 263–274. [Google Scholar] [CrossRef]
- Smet, A.; Martel, A.; Persoons, D.; Dewulf, J.; Heyndrickx, M.; Herman, L.; Haesebrouck, F.; Butaye, P. Broad-spectrum β-lactamases among Enterobacteriaceae of animal origin: Molecular aspects, mobility and impact on public health. FEMS Microbiol. Rev. 2010, 34, 295–316. [Google Scholar] [CrossRef] [Green Version]
- Cantón, R.; González-Alba, J.M.; Galán, J.C. CTX-M enzymes: Origin and diffusion. Front. Microbiol. 2012, 3, 110. [Google Scholar] [CrossRef] [Green Version]
- Jena, J.; Sahoo, R.K.; Debata, N.K.; Subudhi, E. Prevalence of TEM, SHV, and CTX-M genes of extended-spectrum β-lactamase-producing Escherichia coli strains isolated from urinary tract infections in adults. 3 Biotech. 2017, 7, 244. [Google Scholar] [CrossRef] [PubMed]
- Manyahi, J.; Moyo, S.J.; Tellevik, M.G.; Ndugulile, F.; Urassa, W.; Blomberg, B.; Langeland, N. Detection of CTX-M-15 beta-lactamases in Enterobacteriaceae causing hospital- and community-acquired urinary tract infections as early as 2004, in Dar es Salaam, Tanzania. BMC Infect. Dis. 2017, 17, 282. [Google Scholar] [CrossRef]
- Barrios, H.; Garza-Ramos, U.; Mejia-Miranda, I.; Reyna-Flores, F.; Sánchez-Pérez, A.; Mosqueda-García, D.; Silva-Sanchez, J. ESBL-producing Escherichia coli and Klebsiella pneumoniae: The most prevalent clinical isolates obtained between 2005 and 2012 in Mexico. J. Glob. Antimicrob. Resist. 2017, 10, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Falgenhauer, L.; Imirzalioglu, C.; Ghosh, H.; Gwozdzinski, K.; Schmiedel, J.; Gentil, K.; Bauerfeind, R.; Kämpfer, P.; Seifert, H.; Michael, G.B.; et al. Circulation of clonal populations of fluoroquinolone-resistant CTX-M-15-producing Escherichia coli ST410 in humans and animals in Germany. Int. J. Antimicrob. Agents 2016, 47, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, T.; Cantamessa, R.; Melo, L.; Fernandes, M.R.; Fraga, E.; Dropa, M.; Sato, M.I.Z.; Cerdeira, L.; Lincopan, N. International high-risk clones of Klebsiella pneumoniae KPC-2/CC258 and Escherichia coli CTX-M-15/CC10 in urban lake waters. Sci. Total Environ. 2017, 598, 910–915. [Google Scholar] [CrossRef]
- Naseer, U.; Sundsfjord, A. The CTX-M Conundrum: Dissemination of plasmids and Escherichia coli clones. Microb. Drug Resist. 2011, 17, 83–97. [Google Scholar] [CrossRef]
- Lee, K.; Yong, D.; Jeong, S.H.; Tulgaa, K.; Docquier, J.-D.; Rossolini, G.M.; Chong, Y. Genetic and biochemical characterisation of CTX-M-37 extended-spectrum β-lactamase from an Enterobacter cloacae clinical isolate from Mongolia. J. Glob. Antimicrob. Resist. 2017, 10, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Akya, A.; Ahmadi, M.; Khodamoradi, S.; Rezaei, M.R.; Karani, N.; Elahi, A.; Lorestani, R.C.; Rezaei, M. Prevalence of blaCTX-M, blaCTX-M-2, blaCTX-M-8, blaCTX-M-25 and blaCTX-M-3 genes in Escherichia coli isolated from urinary tract infection in Kermanshah City, Iran. J. Clin. Diagnostic Res. 2019, 13, DC04–DC07. [Google Scholar] [CrossRef]
- Poirel, L.; Naas, T.; Nordmann, P. Genetic support of extended-spectrum β-lactamases. Clin. Microbiol. Infect. 2008, 14, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Hamamoto, K.; Hirai, I. Characterisation of chromosomally-located blaCTX-M and its surrounding sequence in CTX-M-type extended-spectrum β-lactamase-producing Escherichia coli isolates. J. Glob. Antimicrob. Resist. 2019, 17, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Stohr, J.J.J.M.; Kluytmans-van den Bergh, M.F.Q.; Wedema, R.; Friedrich, A.W.; Kluytmans, J.A.J.W.; Rossen, J.W.A. Detection of extended-spectrum beta-lactamase (ESBL) genes and plasmid replicons in Enterobacteriaceae using PlasmidSPAdes assembly of short-read sequence data. bioRxiv 2019, 863316. [Google Scholar] [CrossRef]
- Li, P.; Shen, K.; Zhang, Y.; Ying, J.; Zhu, T.; Liu, Y.; Xu, L.; Lin, C.; Zhang, K.; Li, P.; et al. Characterization of a Novel blaKLUC variant with reduced β-lactam resistance from an IncA/C group plasmid in a clinical Klebsiella pneumoniae isolate. Front. Microbiol. 2018, 9, 1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.-H.; Hu, Z.-Q. Epidemiology and genetics of CTX-M extended-spectrum β-lactamases in Gram-negative bacteria. Crit. Rev. Microbiol. 2013, 39, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Seiffert, S.N.; Hilty, M.; Perreten, V.; Endimiani, A. Extended-spectrum cephalosporin-resistant gram-negative organisms in livestock: An emerging problem for human health? Drug Resist. Updat. 2013, 16, 22–45. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.S.; Singhal, N.; Virdi, J.S. Genetic environment of blaTEM-1, blaCTX-M-15, blaCMY-42 and characterization of integrons of Escherichia coli isolated from an Indian urban aquatic environment. Front. Microbiol. 2018, 9, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebana, E.; Carattoli, A.; Coque, T.M.; Hasman, H.; Magiorakos, A.-P.; Mevius, D.; Peixe, L.; Poirel, L.; Schuepbach-Regula, G.; Torneke, K.; et al. Public health risks of enterobacterial isolates producing extended-epectrum β-lactamases or AmpC β-lactamases in food and food-producing animals: An EU perspective of epidemiology, analytical methods, risk factors, and control options. Clin. Infect. Dis. 2012, 56, 1030–1037. [Google Scholar] [CrossRef] [Green Version]
- Shafiq, M.; Huang, J.; Ur Rahman, S.; Shah, J.M.; Chen, L.; Gao, Y.; Wang, M.; Wang, L. High incidence of multidrug-resistant Escherichia coli coharboring mcr-1 and bla (CTX-M-15) recovered from pigs. Infect. Drug Resist. 2019, 12, 2135–2149. [Google Scholar] [CrossRef] [Green Version]
- Woodford, N.; Turton, J.F.; Livermore, D.M. Multiresistant Gram-negative bacteria: The role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 736–755. [Google Scholar] [CrossRef] [Green Version]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-spectrum β-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: A global perspective. Clin. Microbiol. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Flament-Simon, S.-C.; Nicolas-Chanoine, M.-H.; García, V.; Duprilot, M.; Mayer, N.; Alonso, M.P.; García-Meniño, I.; Blanco, J.E.; Blanco, M.; Blanco, J. Clonal structure, virulence factor-encoding genes and antibiotic resistance of Escherichia coli causing urinary tract infections and other extraintestinal infections in humans in Spain and France during 2016. Antibiotics 2020, 9, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, P.N.A.; Ben Zakour, N.L.; Roberts, L.W.; Wailan, A.M.; Zowawi, H.M.; Tambyah, P.A.; Lye, D.C.; Jureen, R.; Lee, T.H.; Yin, M.; et al. Whole genome analysis of cephalosporin-resistant Escherichia coli from bloodstream infections in Australia, New Zealand and Singapore: High prevalence of CMY-2 producers and ST131 carrying blaCTX-M-15 and blaCTX-M-27. J Antimicrob. Chemother. 2018, 73, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi, S.K.S.; Najar-Peerayeh, S.; Mobarez, A.M.; Parizi, M.K. Characterization of uropathogenic E. coli O25b-B2-ST131, O15:K52:H1, and CGA: Neutrophils apoptosis, serum bactericidal assay, biofilm formation, and virulence typing. J. Cell. Physiol. 2019, 234, 18272–18282. [Google Scholar] [CrossRef] [PubMed]
- Usein, C.-R.; Papagheorghe, R.; Oprea, M.; Condei, M.; Strãuţ, M. Molecular characterization of bacteremic Escherichia coli isolates in Romania. Folia Microbiol. (Praha) 2016, 61, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, J.E.; Redondo, L.M.; Figueroa Espinosa, R.A.; Cejas, D.; Gutkind, G.O.; Chacana, P.A.; Di Conza, J.A.; Fernández Miyakawa, M.E. Simultaneous carriage of mcr-1 and other antimicrobial resistance determinants in Escherichia coli from poultry. Front. Microbiol. 2018, 9, 1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barilli, E.; Vismarra, A.; Villa, Z.; Bonilauri, P.; Bacci, C. ESβL E. coli isolated in pig’s chain: Genetic analysis associated to the phenotype and biofilm synthesis evaluation. Int. J. Food Microbiol. 2019, 289, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Dohmen, W.; Dorado-García, A.; Bonten, M.J.M.; Wagenaar, J.A.; Mevius, D.; Heederik, D.J.J. Risk factors for ESBL-producing Escherichia coli on pig farms: A longitudinal study in the context of reduced use of antimicrobials. PLoS ONE 2017, 12, e0174094. [Google Scholar] [CrossRef]
- Ramos, S.; Silva, N.; Dias, D.; Sousa, M.; Capelo-Martinez, J.L.; Brito, F.; Caniça, M.; Igrejas, G.; Poeta, P. Clonal diversity of ESBL-producing Escherichia coli in pigs at slaughter level in Portugal. Foodborne Pathog. Dis. 2012, 10, 74–79. [Google Scholar] [CrossRef]
- Randall, L.P.; Lemma, F.; Rogers, J.P.; Cheney, T.E.A.; Powell, L.F.; Teale, C.J. Prevalence of extended-spectrum-β-lactamase-producing Escherichia coli from pigs at slaughter in the UK in 2013. J. Antimicrob. Chemother. 2014, 69, 2947–2950. [Google Scholar] [CrossRef] [Green Version]
- Hering, J.; Hille, K.; Frömke, C.; von Münchhausen, C.; Hartmann, M.; Schneider, B.; Friese, A.; Roesler, U.; Merle, R.; Kreienbrock, L. Prevalence and potential risk factors for the occurrence of cefotaxime resistant Escherichia coli in German fattening pig farms—A cross-sectional study. Prev. Vet. Med. 2014, 116, 129–137. [Google Scholar] [CrossRef]
- Ramos, S.; Igrejas, G.; Silva, N.; Jones-Dias, D.; Capelo-Martinez, J.-L.; Caniça, M.; Poeta, P. First report of CTX-M producing Escherichia coli, including the new ST2526, isolated from beef cattle and sheep in Portugal. Food Control 2013, 31, 208–210. [Google Scholar] [CrossRef]
- Haenni, M.; Châtre, P.; Métayer, V.; Bour, M.; Signol, E.; Madec, J.-Y.; Gay, E. Comparative prevalence and characterization of ESBL-producing Enterobacteriaceae in dominant versus subdominant enteric flora in veal calves at slaughterhouse, France. Vet. Microbiol. 2014, 171, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Velasova, M.; Smith, R.P.; Lemma, F.; Horton, R.A.; Duggett, N.A.; Evans, J.; Tongue, S.C.; Anjum, M.F.; Randall, L.P. Detection of extended-spectrum β-lactam, AmpC and carbapenem resistance in Enterobacteriaceae in beef cattle in Great Britain in 2015. J. Appl. Microbiol. 2019, 126, 1081–1095. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Feng, C.; Xu, H.; Yu, X.; Guo, L.; Jiang, X.; Song, X. Detection and characterization of ESBL-producing Escherichia coli expressing mcr-1 from dairy cows in China. J. Antimicrob. Chemother. 2018, 74, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Gonggrijp, M.A.; Santman-Berends, I.M.G.A.; Heuvelink, A.E.; Buter, G.J.; van Schaik, G.; Hage, J.J.; Lam, T.J.G.M. Prevalence and risk factors for extended-spectrum β-lactamase- and AmpC-producing Escherichia coli in dairy farms. J. Dairy Sci. 2016, 99, 9001–9013. [Google Scholar] [CrossRef]
- Boonyasiri, A.; Tangkoskul, T.; Seenama, C.; Saiyarin, J.; Tiengrim, S.; Thamlikitkul, V. Prevalence of antibiotic resistant bacteria in healthy adults, foods, food animals, and the environment in selected areas in Thailand. Pathog. Glob. Health 2014, 108, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Nuangmek, A.; Rojanasthien, S.; Chotinun, S.; Yamsakul, P.; Tadee, P.; Thamlikitkul, V.; Tansakul, N.; Patchanee, P. Antimicrobial resistance in ESBL-producing Escherichia coli isolated from layer and pig farms in Thailand. Acta Sci. Vet. 2018, 46, 1538. [Google Scholar] [CrossRef] [Green Version]
- Reich, F.; Schill, F.; Atanassova, V.; Klein, G. Quantification of ESBL-Escherichia coli on broiler carcasses after slaughtering in Germany. Food Microbiol. 2016, 54, 1–5. [Google Scholar] [CrossRef]
- Casella, T.; Nogueira, M.C.L.; Saras, E.; Haenni, M.; Madec, J.-Y. High prevalence of ESBLs in retail chicken meat despite reduced use of antimicrobials in chicken production, France. Int. J. Food Microbiol. 2017, 257, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Wang, Y.; Shi, X.; Wang, S.; Ren, H.; Shen, Z.; Wang, Y.; Lin, J.; Wang, S. Rapid rise of the ESBL and mcr-1 genes in Escherichia coli of chicken origin in China, 2008–2014. Emerg. Microbes Infect. 2018, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhai, Z.; Li, Q.; Liu, L.; Guo, S.; Li, Q.; Yang, L.; Ye, C.; Chang, W.; Zhai, J. Characterization of extended-spectrum β-lactamase-producing Escherichia coli isolates from pigs and farm workers. J. Food Prot. 2016, 79, 1630–1634. [Google Scholar] [CrossRef] [PubMed]
- Norizuki, C.; Kawamura, K.; Wachino, J.; Suzuki, M.; Nagano, N.; Kondo, T.; Arakawa, Y. Detection of Escherichia coli producing CTX-M-1-group extended-spectrum β-lactamases from pigs in Aichi prefecture, Japan, between 2015 and 2016. Jpn. J. Infect. Dis. 2017, 71, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diab, M.; Hamze, M.; Madec, J.-Y.; Haenni, M. High Prevalence of non-ST131 CTX-M-15-producing Escherichia coli in healthy cattle in Lebanon. Microb. Drug Resist. 2016, 23, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Manageiro, V.; Clemente, L.; Graça, R.; Correia, I.; Albuquerque, T.; Ferreira, E.; Caniça, M. New insights into resistance to colistin and third-generation cephalosporins of Escherichia coli in poultry, Portugal: Novel blaCTX-M-166 and blaESAC genes. Int. J. Food Microbiol. 2017, 263, 67–73. [Google Scholar] [CrossRef]
- Päivärinta, M.; Pohjola, L.; Fredriksson-Ahomaa, M.; Heikinheimo, A. Low occurrence of extended-spectrum β-lactamase-producing Escherichia coli in Finnish food-producing animals. Zoonoses Public Health 2016, 63, 624–631. [Google Scholar] [CrossRef]
- Apostolakos, I.; Mughini-Gras, L.; Fasolato, L.; Piccirillo, A. Assessing the occurrence and transfer dynamics of ESBL/pAmpC-producing Escherichia coli across the broiler production pyramid. PLoS ONE 2019, 14, e0217174. [Google Scholar] [CrossRef]
- Kraemer, J.G.; Pires, J.; Kueffer, M.; Semaani, E.; Endimiani, A.; Hilty, M.; Oppliger, A. Prevalence of extended-spectrum β-lactamase-producing Enterobacteriaceae and methicillin-resistant Staphylococcus aureus in pig farms in Switzerland. Sci. Total Environ. 2017, 603–604, 401–405. [Google Scholar] [CrossRef] [Green Version]
- WHO. Global Antimicrobial Resistance Surveillance System (GLASS) Report: Early Implementation 2017–2018; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Poirel, L.; Madec, J.-Y.; Lupo, A.; Schink, A.-K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.-T.; Song, F.-J.; Zou, M.; Zhang, Q.-D.; Shan, H. High incidence of Escherichia coli strains coharboring mcr-1 and blaNDM from chickens. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Silva, N.; Carvalho, I.; Currie, C.; Sousa, M.; Igrejas, G.; Poeta, P. Extended-spectrum-β-lactamase and carbapenemase-producing Enterobacteriaceae in food-producing animals in Europe. Antibiot. Drug Resist. 2019, 12, 261–273. [Google Scholar]
- Carattoli, A. Animal reservoirs for extended spectrum β-lactamase producers. Clin. Microbiol. Infect. 2008, 14, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente, L.; Manageiro, V.; Correia, I.; Amaro, A.; Albuquerque, T.; Themudo, P.; Ferreira, E.; Caniça, M. Revealing mcr-1-positive ESBL-producing Escherichia coli strains among Enterobacteriaceae from food-producing animals (bovine, swine and poultry) and meat (bovine and swine), Portugal, 2010–2015. Int. J. Food Microbiol. 2019, 296, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Day, M.J.; Hopkins, K.L.; Wareham, D.W.; Toleman, M.A.; Elviss, N.; Randall, L.; Teale, C.; Cleary, P.; Wiuff, C.; Doumith, M.; et al. Extended-spectrum β-lactamase-producing Escherichia coli in human-derived and food chain-derived samples from England, Wales, and Scotland: An epidemiological surveillance and typing study. Lancet Infect. Dis. 2019, 19, 1325–1335. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, S.; Silva, V.; Dapkevicius, M.d.L.E.; Caniça, M.; Tejedor-Junco, M.T.; Igrejas, G.; Poeta, P. Escherichia coli as Commensal and Pathogenic Bacteria among Food-Producing Animals: Health Implications of Extended Spectrum β-Lactamase (ESBL) Production. Animals 2020, 10, 2239. https://doi.org/10.3390/ani10122239
Ramos S, Silva V, Dapkevicius MdLE, Caniça M, Tejedor-Junco MT, Igrejas G, Poeta P. Escherichia coli as Commensal and Pathogenic Bacteria among Food-Producing Animals: Health Implications of Extended Spectrum β-Lactamase (ESBL) Production. Animals. 2020; 10(12):2239. https://doi.org/10.3390/ani10122239
Chicago/Turabian StyleRamos, Sónia, Vanessa Silva, Maria de Lurdes Enes Dapkevicius, Manuela Caniça, María Teresa Tejedor-Junco, Gilberto Igrejas, and Patrícia Poeta. 2020. "Escherichia coli as Commensal and Pathogenic Bacteria among Food-Producing Animals: Health Implications of Extended Spectrum β-Lactamase (ESBL) Production" Animals 10, no. 12: 2239. https://doi.org/10.3390/ani10122239