
Possible Dietary Effects of Insect-Based Diets across Zebrafish (Danio rerio) Generations: A Multidisciplinary Study on the Larval Phase

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
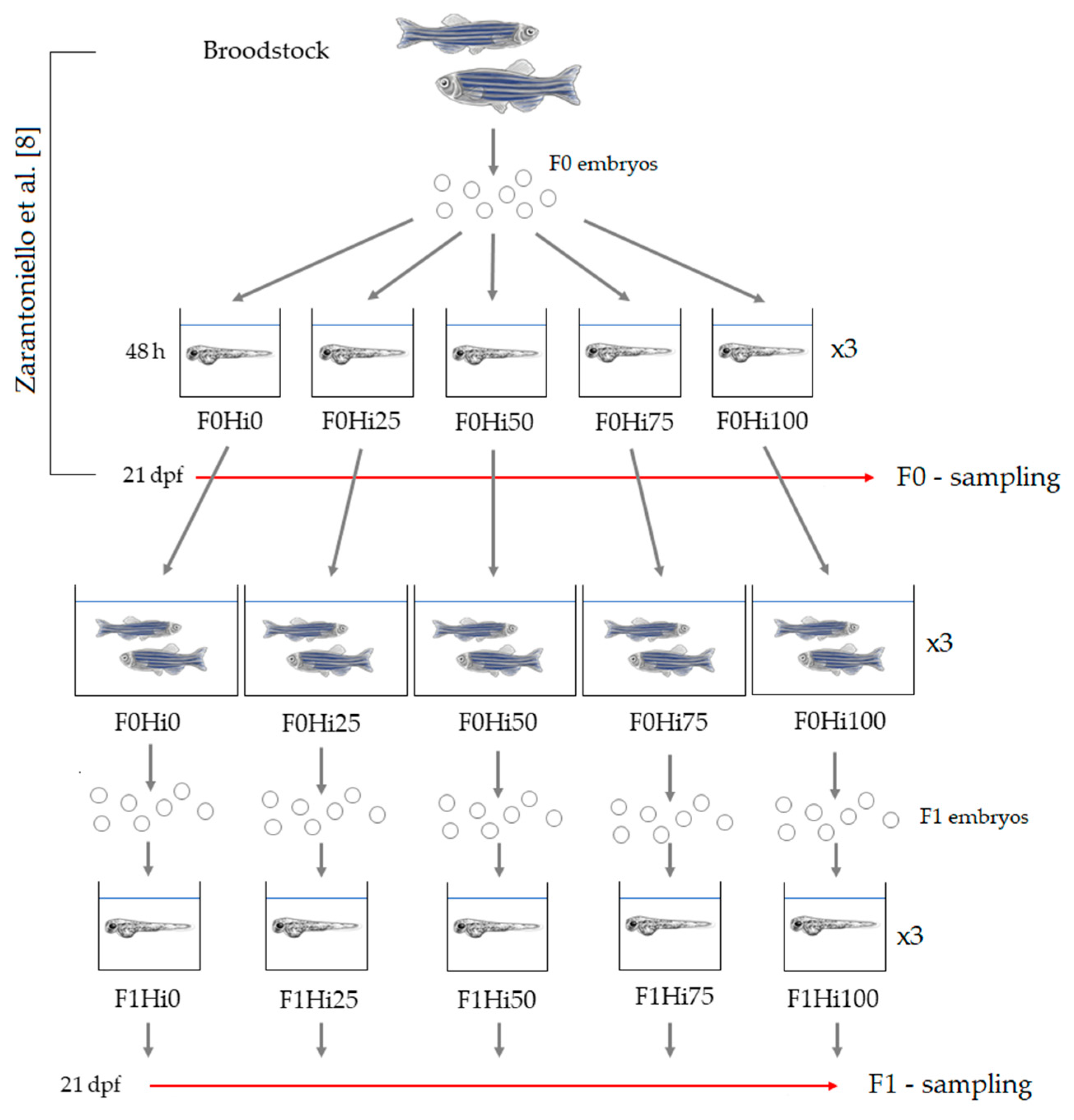
2. Materials and Methods
2.1. Insect Rearing
2.2. Fish Diets
2.3. Broodstock Rearing and F0 Production
2.4. F1 Zebrafish Larvae
2.5. Biometry
2.6. Fatty Acid Composition
2.7. Histology
2.8. Total RNA Extraction and cDNA Synthesis
2.9. Real-Time PCR
2.10. Statistical Analyses
3. Results
3.1. F1 Zebrafish Larvae—Growth and Survival
3.2. F1 Zebrafish Larvae—Fatty Acid Content and Composition
3.3. F1 Zebrafish Larvae—Histology
3.4. F1 Zebrafish Larvae—Real-Time PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Oliva-Teles, A.; Enes, P.; Peres, H. Replacing fishmeal and fish oil in industrial aquafeeds for carnivorous fish. In Feed and Feeding Practices in Aquaculture; Davis, D.A., Ed.; Woodhead Publishing: Cambridge, UK, 2015; pp. 203–233. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Metian, M. Feed matters: Satisfying the feed demand of aquaculture. Rev. Fish. Sci. Aquac. 2015, 23, 1–10. [Google Scholar] [CrossRef]
- Gasco, L.; Finke, M.; van Huis, A. Can diets containing insects promote animal health? J. Insects Food Feed 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Berggren, Å.; Jansson, A.; Low, M. Approaching Ecological Sustainability in the Emerging Insects-as-Food Industry. Trends Ecol. Evol. 2019, 34, 132–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parodi, A.; De Boer, I.J.M.; Gerrits, W.J.J.; Van Loon, J.J.A.; Heetkamp, M.J.W.; Van Schelt, J.; Bolhuis, J.E.; Van Zanten, H.H.E. Bioconversion efficiencies, greenhouse gas and ammonia emissions during black soldier fly rearing—A mass balance approach. J. Clean. Prod. 2020, 271, 122488. [Google Scholar] [CrossRef]
- Barroso, F.G.; Sánchez-Muros, M.J.; Rincón, M.Á.; Rodriguez-Rodriguez, M.; Fabrikov, D.; Morote, E.; Guil-Guerrero, J.L. Production of n-3-rich insects by bioaccumulation of fishery waste. J. Food Compos. Anal. 2019, 82, 103237. [Google Scholar] [CrossRef]
- Truzzi, C.; Giorgini, E.; Annibaldi, A.; Antonucci, M.; Illuminati, S.; Scarponi, G.; Riolo, P.; Isidoro, N.; Conti, C.; Zarantoniello, M.; et al. Fatty acids profile of black soldier fly (Hermetia illucens): Influence of feeding substrate based on coffee-waste silverskin enriched with microalgae. Anim. Feed Sci. Technol. 2020, 259, 114309. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Zimbelli, A.; Randazzo, B.; Compagni, M.D.; Truzzi, C.; Antonucci, M.; Riolo, P.; Loreto, N.; Osimani, A.; Milanović, V.; et al. Black Soldier Fly (Hermetia illucens) reared on roasted coffee by-product and Schizochytrium sp. as a sustainable terrestrial ingredient for aquafeeds production. Aquaculture 2020, 518, 734659. [Google Scholar] [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Gioacchini, G.; Cardinaletti, G.; Belloni, A.; Giorgini, E.; Faccenda, F.; Cerri, R.; Tibaldi, E.; Olivotto, I. Physiological response of rainbow trout (Oncorhynchus mykiss) to graded levels of Hermetia illucens or poultry by-product meals as single or combined substitute ingredients to dietary plant proteins. Aquaculture 2021, 538, 736550. [Google Scholar] [CrossRef]
- Bruni, L.; Randazzo, B.; Cardinaletti, G.; Zarantoniello, M.; Mina, F.; Secci, G.; Tulli, F.; Olivotto, I.; Parisi, G. Dietary inclusion of full-fat Hermetia illucens prepupae meal in practical diets for rainbow trout (Oncorhynchus mykiss): Lipid metabolism and fillet quality investigations. Aquaculture 2020, 529, 735678. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Gjesdal, P.; Biancarosa, I.; Menchetti, E.; Li, Y.; Waagbø, R.; Krogdahl, Å.; Lock, E.J. Black soldier fly larvae meal can replace fish meal in diets of sea-water phase Atlantic salmon (Salmo salar). Aquaculture 2019, 503, 609–619. [Google Scholar] [CrossRef]
- Li, Y.; Kortner, T.M.; Chikwati, E.M.; Belghit, I.; Lock, E.J.; Krogdahl, Å. Total replacement of fish meal with black soldier fly (Hermetia illucens) larvae meal does not compromise the gut health of Atlantic salmon (Salmo salar). Aquaculture 2020, 520, 734967. [Google Scholar] [CrossRef]
- Caimi, C.; Renna, M.; Lussiana, C.; Bonaldo, A.; Gariglio, M.; Meneguz, M.; Dabbou, S.; Schiavone, A.; Gai, F.; Elia, A.C.; et al. First insights on Black Soldier Fly (Hermetia illucens L.) larvae meal dietary administration in Siberian sturgeon (Acipenser baerii Brandt) juveniles. Aquaculture 2020, 515, 734539. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Nozzi, V.; Truzzi, C.; Giorgini, E.; Cardinaletti, G.; Freddi, L.; Ratti, S.; Girolametti, F.; Osimani, A.; et al. Physiological responses of Siberian sturgeon (Acipenser baerii) juveniles fed on full-fat insect-based diet in an aquaponic system. Sci. Rep. 2021, 11, 1057. [Google Scholar] [CrossRef] [PubMed]
- Zarantoniello, M.; Randazzo, B.; Truzzi, C.; Giorgini, E.; Marcellucci, C.; Vargas-Abúndez, J.A.; Zimbelli, A.; Annibaldi, A.; Parisi, G.; Tulli, F.; et al. A six-months study on Black Soldier Fly (Hermetia illucens) based diets in zebrafish. Sci. Rep. 2019, 9, 8598. [Google Scholar] [CrossRef] [PubMed]
- Zarantoniello, M.; Randazzo, B.; Gioacchini, G.; Truzzi, C.; Giorgini, E.; Riolo, P.; Gioia, G.; Bertolucci, C.; Osimani, A.; Cardinaletti, G.; et al. Zebrafish (Danio rerio) physiological and behavioural responses to insect-based diets: A multidisciplinary approach. Sci. Rep. 2020, 10, 10648. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Secci, G.; Notarstefano, V.; Giorgini, E.; Lock, E.J.; Parisi, G.; Olivotto, I. Application of laboratory methods for understanding fish responses to Black Soldier Fly (Hermetia illucens) based diets. J. Insects Food Feed 2021, in press. [Google Scholar]
- Ewald, N.; Vidakovic, A.; Langeland, M.; Kiessling, A.; Sampels, S.; Lalander, C. Fatty acid composition of black soldier fly larvae (Hermetia illucens)—Possibilities and limitations for modification through diet. Waste Manag. 2020, 102, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Randazzo, B.; Zarantoniello, M.; Gioacchini, G.; Giorgini, E.; Truzzi, C.; Notarstefano, V.; Cardinaletti, G.; Huyen, K.T.; Carnevali, O.; Olivotto, I. Can insect-based diets affect zebrafish (Danio rerio) reproduction? A multidisciplinary study. Zebrafish 2020, 17, 287–304. [Google Scholar] [CrossRef]
- Wakefield, S.L.; Lane, M.; Schulz, S.J.; Hebart, M.L.; Thompson, J.G.; Mitchell, M. Maternal supply of omega-3 polyunsaturated fatty acids alter mechanisms involved in oocyte and early embryo development in the mouse. Am. J. Physiol.-Endocrinol. Metab. 2008, 294, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Wonnacott, K.E.; Kwong, W.Y.; Hughes, J.; Salter, A.M.; Lea, R.G.; Garnsworthy, P.C.; Sinclair, K.D. Dietary omega-3 and -6 polyunsaturated fatty acids affect the composition and development of sheep granulosa cells, oocytes and embryos. Reproduction 2010, 139, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Fuiman, L.A.; Ojanguren, A.F. Fatty acid content of eggs determines antipredator performance of fish larvae. J. Exp. Mar. Biol. Ecol. 2011, 407, 155–165. [Google Scholar] [CrossRef]
- Perez, K.O.; Fuiman, L.A. Maternal diet and larval diet influence survival skills of larval red drum Sciaenops ocellatus. J. Fish Biol. 2015, 86, 1286–1304. [Google Scholar] [CrossRef] [PubMed]
- Asil, S.M.; Kenari, A.A.; Miyanji, G.R.; Van Der Kraak, G. The influence of dietary arachidonic acid on growth, reproductive performance, and fatty acid composition of ovary, egg and larvae in an anabantid model fish, Blue gourami (Trichopodus trichopterus; Pallas, 1770). Aquaculture 2017, 476, 8–18. [Google Scholar] [CrossRef]
- Duque-Guimarães, D.E.; Ozanne, S.E. Nutritional programming of insulin resistance: Causes and consequences. Trends Endocrinol. Metab. 2013, 24, 525–535. [Google Scholar] [CrossRef]
- Xu, H.; Turkmen, S.; Rimoldi, S.; Terova, G.; Zamorano, M.J.; Afonso, J.M.; Sarih, S.; Fernández-Palacios, H.; Izquierdo, M. Nutritional intervention through dietary vegetable proteins and lipids to gilthead sea bream (Sparus aurata) broodstock affects the offspring utilization of a low fishmeal/fish oil diet. Aquaculture 2019, 513, 734402. [Google Scholar] [CrossRef]
- Hou, Z.; Fuiman, L.A. Nutritional programming in fishes: Insights from mammalian studies. Rev. Fish Biol. Fish. 2020, 30, 67–92. [Google Scholar] [CrossRef]
- Fernández-Palacios, H.; Norberg, B.; Izquierdo, M.; Hamre, K. Effects of Broodstock Diet on Eggs and Larvae. In Larval Fish Nutrition; Holt, G.J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 153–181. [Google Scholar] [CrossRef]
- Izquierdo, M.S.; Socorro, J.; Arantzamendi, L.; Hernández-Cruz, C.M. Recent advances in lipid nutrition in fish larvae. Fish Physiol. Biochem. 2000, 22, 97–107. [Google Scholar] [CrossRef]
- Fuiman, L.A.; Perez, K.O. Metabolic programming mediated by an essential fatty acid alters body composition and survival skills of a marine fish. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151414. [Google Scholar] [CrossRef]
- Adam, A.-C.; Skjærven, K.H.; Whatmore, P.; Moren, M.; Lie, K.K. Parental high dietary arachidonic acid levels modulated the hepatic transcriptome of adult zebrafish (Danio rerio) progeny. PLoS ONE 2018, 13, e0201278. [Google Scholar] [CrossRef] [Green Version]
- Fontagné-Dicharry, S.; Alami-Durante, H.; Aragão, C.; Kaushik, S.J.; Geurden, I. Parental and early-feeding effects of dietary methionine in rainbow trout (Oncorhynchus mykiss). Aquaculture 2017, 469, 16–27. [Google Scholar] [CrossRef]
- Izquierdo, M.; Koven, W. Lipids. In Larval Fish Nutrition; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 47–81. [Google Scholar] [CrossRef]
- Monaghan, P. Early growth conditions, phenotypic development and environmental change. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 1635–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotoh, T. Potential of the application of epigenetics in animal production. Anim. Prod. Sci. 2015, 55, 145–158. [Google Scholar] [CrossRef]
- Izquierdo, M.S.; Turkmen, S.; Montero, D.; Zamorano, M.J.; Afonso, J.M.; Karalazos, V.; Fernández-Palacios, H. Nutritional programming through broodstock diets to improve utilization of very low fishmeal and fish oil diets in gilthead sea bream. Aquaculture 2015, 449, 18–26. [Google Scholar] [CrossRef]
- Turkmen, S.; Zamorano, M.J.; Fernández-Palacios, H.; Hernández-Cruz, C.M.; Montero, D.; Robaina, L.; Izquierdo, M. Parental nutritional programming and a reminder during juvenile stage affect growth, lipid metabolism and utilisation in later developmental stages of a marine teleost, the gilthead sea bream (Sparus aurata). Br. J. Nutr. 2017, 118, 500–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turkmen, S.; Hernández-Cruz, C.M.; Zamorano, M.J.; Fernández-Palacios, H.; Montero, D.; Afonso, J.M.; Izquierdo, M. Long-chain PUFA profiles in parental diets induce long-term effects on growth, fatty acid profiles, expression of fatty acid desaturase 2 and selected immune system-related genes in the offspring of gilthead seabream. Br. J. Nutr. 2019, 122, 25–38. [Google Scholar] [CrossRef]
- Connor, W.E. Importance of n-3 fatty acids in health and disease. Am. J. Clin. Nutr. 2000, 71, 171S–175S. [Google Scholar] [CrossRef] [PubMed]
- FAO. State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals. 2018. Available online: http://www.fao.org/3/i9540en/i9540en.pdf (accessed on 9 July 2018).
- Lawrence, C. The husbandry of zebrafish (Danio rerio): A review. Aquaculture 2007, 269, 1–20. [Google Scholar] [CrossRef]
- Dutta, S. Food and feeding habits of Danio rerio (Ham. Buch) inhabiting Gadigarh Stream, Jammu. J. Freshw. Biol. 1993, 5, 165–168. [Google Scholar]
- Spence, R.; Fatema, M.K.; Ellis, S.; Ahmed, Z.F.; Smith, C. Diet, growth and recruitment of wild zebrafish in Bangladesh. J. Fish Biol. 2007, 71, 304–309. [Google Scholar] [CrossRef]
- McClure, M.M.; McIntyre, P.B.; McCune, A.R. Notes on the natural diet and habitat of eight danionin fishes, including the zebrafish Danio rerio. J. Fish Biol. 2006, 69, 553–570. [Google Scholar] [CrossRef]
- Vargas, A.; Randazzo, B.; Riolo, P.; Truzzi, C.; Gioacchini, G.; Giorgini, E.; Loreto, N.; Ruschioni, S.; Zarantoniello, M.; Antonucci, M.; et al. Rearing zebrafish on Black Soldier Fly (Hermetia illucens): Biometric, histological, spectroscopic, biochemical, and molecular implications. Zebrafish 2018, 15, 404–419. [Google Scholar] [CrossRef]
- Ribas, L.; Piferrer, F. The zebrafish (Danio rerio) as a model organism, with emphasis on applications for finfish aquaculture research. Rev. Aquac. 2014, 6, 209–240. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef] [PubMed]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Influence of larval density and dietary nutrient concentration on performance, body protein, and fat contents of black soldier fly larvae (Hermetia illucens). Entomol. Exp. Appl. 2018, 166, 761–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diener, S.; Zurbrügg, C.; Tockner, K. Conversion of organic material by black soldier fly larvae: Establishing optimal feeding rates. Waste Manag. Res. 2009, 27, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Osimani, A.; Ferrocino, I.; Corvaglia, M.R.; Roncolini, A.; Milanović, V.; Garofalo, C.; Aquilanti, L.; Riolo, P.; Ruschioni, S.; Jamshidi, E.; et al. Microbial dynamics in rearing trials of Hermetia illucens larvae fed cofee silverskin and microalgae. Food Res. Int. 2021, 140, 110028. [Google Scholar] [CrossRef]
- Olivotto, I.; Yasumasu, S.; Gioacchini, G.; Maradonna, F.; Cionna, C.; Carnevali, O. Cloning and expression of high choriolytic enzyme, a component of the hatching enzyme system, during embryonic development of the marine ornamental fish Chrysiptera parasema. Mar. Biol. 2004, 145, 1235–1241. [Google Scholar] [CrossRef]
- Truzzi, C.; Illuminati, S.; Antonucci, M.; Scarponi, G.; Annibaldi, A. Heat shock influences the fatty acid composition of the muscle of the Antarctic fish Trematomus bernacchii. Mar. Environ. Res. 2018, 139, 122–128. [Google Scholar] [CrossRef]
- Canonico, L.; Ashoor, S.; Taccari, M.; Comitini, F.; Antonucci, M.; Truzzi, C.; Scarponi, G.; Ciani, M. Conversion of raw glycerol to microbial lipids by new Metschnikowia and Yarrowia lipolytica strains. Ann. Microbiol. 2016, 66, 1409–1418. [Google Scholar] [CrossRef]
- Truzzi, C.; Annibaldi, A.; Antonucci, M.; Scarponi, G.; Illuminati, S. Gas chromatography-mass spectrometry analysis on effects of thermal shock on the fatty acid composition of the gills of the Antarctic teleost, Trematomus bernacchii. Environ. Chem. 2018, 15, 424–435. [Google Scholar] [CrossRef]
- Panettieri, V.; Chatzifotis, S.; Messina, C.M.; Olivotto, I.; Manuguerra, S.; Randazzo, B.; Ariano, A.; Bovera, F.; Santulli, A.; Severino, L.; et al. Honey bee pollen in meagre (Argyrosomus regius) juvenile diets: Effects on growth, diet digestibility, intestinal traits, and biochemical markers related to health and stress. Animals 2020, 10, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, A. Role of nutritional programming in determining adult morbidity. Arch. Dis. Child. 1994, 71, 288–290. [Google Scholar] [CrossRef] [Green Version]
- Burdge, G.C.; Lillycrop, K.A. Nutrition, epigenetics, and developmental plasticity: Implications for understanding human disease. Annu. Rev. Nutr. 2010, 30, 315–339. [Google Scholar] [CrossRef] [PubMed]
- Pittman, K.; Yúfera, M.; Pavlidis, M.; Geffen, A.J.; Koven, W.; Ribeiro, L.; Zambonino-Infante, J.L.; Tandler, A. Fantastically plastic: Fish larvae equipped for a new world. Rev. Aquac. 2013, 5, S224–S267. [Google Scholar] [CrossRef] [Green Version]
- Turkmen, S.; Zamorano, M.J.; Xu, H.; Fernández-Palacios, H.; Robaina, L.; Kaushik, S.; Izquierdo, M. Parental LC-PUFA biosynthesis capacity and nutritional intervention with alpha-linolenic acid affect performance of Sparus aurata progeny. J. Exp. Biol. 2020, 223, jeb214999. [Google Scholar] [CrossRef]
- Xu, H.; Ferosekhan, S.; Turkmen, S.; Afonso, J.M.; Zamorano, M.J.; Izquierdo, M. High broodstock fads2 expression combined with nutritional programing through broodstock diet improves the use of low fishmeal and low fish oil diets in gilthead seabream (Sparus aurata) progeny. Aquaculture 2021, 535, 736321. [Google Scholar] [CrossRef]
- Ferosekhan, S.; Xu, H.; Turkmen, S.; Gómez, A.; Afonso, J.M.; Fontanillas, R.; Rosenlund, G.; Kaushik, S.; Izquierdo, M. Reproductive performance of gilthead seabream (Sparus aurata) broodstock showing different expression of fatty acyl desaturase 2 and fed two dietary fatty acid profiles. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Cardinaletti, G.; Randazzo, B.; Messina, M.; Zarantoniello, M.; Giorgini, E.; Zimbelli, A.; Bruni, L.; Parisi, G.; Olivotto, I.; Tulli, F. Effects of graded dietary inclusion level of full-fat Hermetia illucens prepupae meal in practical diets for rainbow trout (Oncorhynchus mykiss). Animals 2019, 9, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Research Council. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Dhanasiri, A.K.S.; Johny, A.; Xue, X.; Berge, G.M.; Bogevik, A.S.; Rise, M.L.; Fæste, C.K.; Fernandes, J.M.O. Plant-based diets induce transcriptomic changes in muscle of zebrafish and Atlantic salmon. Front. Genet. 2020, 11, 1288. [Google Scholar] [CrossRef]
- Turkmen, S.; Perera, E.; Zamorano, M.J.; Simó-Mirabet, P.; Xu, H.; Pérez-Sánchez, J.; Izquierdo, M. Effects of dietary lipid composition and fatty acid desaturase 2 expression in broodstock gilthead sea bream on lipid metabolism-related genes and methylation of the fads2 gene promoter in their offspring. Int. J. Mol. Sci. 2019, 20, 6250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Benatti, P.; Nicolai, R.; Calvani, M.; Peluso, G. Polyunsaturated fatty acids: Biochemical, nutritional and epigenetic properties. J. Am. Coll. Nutr. 2004, 23, 281–302. [Google Scholar] [CrossRef] [PubMed]
- Feil, R. Environmental and nutritional effects on the epigenetic regulation of genes. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2006, 600, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Morais, S.; Mendes, A.C.; Castanheira, M.F.; Coutinho, J.; Bandarra, N.; Dias, J.; Conceição, L.E.C.; Pousão-Ferreira, P. New formulated diets for Solea senegalensis broodstock: Effects of parental nutrition on biosynthesis of long-chain polyunsaturated fatty acids and performance of early larval stages and juvenile fish. Aquaculture 2014, 432, 374–382. [Google Scholar] [CrossRef]
- Bell, M.V.; Tocher, D.R. Biosynthesis of polyunsaturated fatty acids in aquatic ecosystems: General pathways and new directions. In Lipids in Aquatic Ecosystems; Springer: New York, NY, USA, 2009; pp. 211–236. [Google Scholar] [CrossRef]
- Izquierdo, M.S.; Robaina, L.; Juárez-Carrillo, E.; Oliva, V.; Hernández-Cruz, C.M.; Afonso, J.M. Regulation of growth, fatty acid composition and delta 6 desaturase expression by dietary lipids in gilthead seabream larvae (Sparus aurata). Fish Physiol. Biochem. 2008, 34, 117–127. [Google Scholar] [CrossRef]
- Li, S.; Mai, K.; Xu, W.; Yuan, Y.; Zhang, Y.; Ai, Q. Characterization, mRNA expression and regulation of δ6 fatty acyl desaturase (FADS2) by dietary n-3 long chain polyunsaturated fatty acid (LC-PUFA) levels in grouper larvae (Epinephelus coioides). Aquaculture 2014, 434, 212–219. [Google Scholar] [CrossRef]
- Vagner, M.; Santigosa, E. Characterization and modulation of gene expression and enzymatic activity of delta-6 desaturase in teleosts: A review. Aquaculture 2011, 315, 131–143. [Google Scholar] [CrossRef]
- Madison, B.N.; Tavakoli, S.; Kramer, S.; Bernier, N.J. Chronic cortisol and the regulation of food intake and the endocrine growth axis in rainbow trout. J. Endocrinol. 2015, 226, 103–119. [Google Scholar] [CrossRef] [Green Version]
- Deck, C.A.; Honeycutt, J.L.; Cheung, E.; Reynolds, H.M.; Borski, R.J. Assessing the functional role of leptin in energy homeostasis and the stress response in vertebrates. Front. Endocrinol. 2017, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Chisada, S.; Kurokawa, T.; Murashita, K.; Rønnestad, I.; Taniguchi, Y.; Toyoda, A.; Sakaki, Y.; Takeda, S.; Yoshiura, Y. Leptin receptor-deficient (knockout) medaka, Oryzias latipes, show chronical up-regulated levels of orexigenic neuropeptides, elevated food intake and stage specific effects on growth and fat allocation. Gen. Comp. Endocrinol. 2014, 195, 9–20. [Google Scholar] [CrossRef]
- Li, A.; Yuan, X.; Liang, X.-F.; Liu, L.; Li, J.; Li, B.; Fang, J.; Li, J.; He, S.; Xue, M.; et al. Adaptations of lipid metabolism and food intake in response to low and high fat diets in juvenile grass carp (Ctenopharyngodon idellus). Aquaculture 2016, 457, 43–49. [Google Scholar] [CrossRef]
- Kwasek, K.; Wojno, M.; Iannini, F.; McCracken, V.J.; Molinari, G.S.; Terova, G. Nutritional programming improves dietary plant protein utilization in zebrafish Danio rerio. PLoS ONE 2020, 15, e0225917. [Google Scholar] [CrossRef] [Green Version]
- Buddington, R.K.; Krogdahl, Å. Hormonal regulation of the fish gastrointestinal tract. Comp. Biochem. Physiol.-A Mol. Integr. Physiol. 2004, 139, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Baeverfjord, G.; Krogdahl, A. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: A comparison with the intestines of fasted fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Perera, E.; Yufera, M. Soybean Meal and Soy Protein Concentrate in Early Diet Elicit Different Nutritional Programming Effects on Juvenile Zebrafish. Zebrafish 2016, 13, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Randazzo, B.; Zarantoniello, M.; Cardinaletti, G.; Cerri, R.; Giorgini, E.; Belloni, A.; Tibaldi, E.; Olivotto, I. Hermetia illucens and poultry by-product meals as alternatives to plant protein sources in Gilthead seabream (Sparus aurata) diet. A multidisciplinary study on fish gut status. Animals 2021, 11, 677. [Google Scholar] [CrossRef]
- Rimoldi, S.; Gini, E.; Iannini, F.; Gasco, L.; Terova, G. The effects of dietary insect meal from Hermetia illucens prepupae on autochthonous gut microbiota of rainbow trout (Oncorhynchus mykiss). Animals 2019, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Ringø, E.; Zhou, Z.; Vecino, J.L.G.; Wadsworth, S.; Romero, J.; Krogdahl, A.A.; Olsen, R.E.; Dimitroglou, A.; Foey, A.; Davies, S.; et al. Effect of dietary components on the gut microbiota of aquatic animals. A never-ending story? Aquac. Nutr. 2016, 22, 219–282. [Google Scholar] [CrossRef] [Green Version]
- Ringø, E.; Zhou, Z.; Olsen, R.E.; Song, S.K. Use of chitin and krill in aquaculture—The effect on gut microbiota and the immune system: A review. Aquac. Nutr. 2012, 18, 117–131. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hi0 | Hi25 | Hi50 | Hi75 | Hi100 | |
|---|---|---|---|---|---|
| Ingredients (g/kg) | |||||
| Fish meal 1 | 470 | 400 | 250 | 110 | - |
| Vegetable protein mix 2 | 220 | 230 | 298 | 385 | 440 |
| BSF prepupae meal | - | 115 | 235 | 350 | 460 |
| Wheat flour 3 | 198 | 172 | 120 | 110 | 72 |
| Fish oil | 80 | 51 | 25 | 10 | - |
| Soy lecithin | 8 | 8 | 8 | 11 | 4 |
| Mineral and Vitamin supplements 4 | 14 | 14 | 14 | 14 | 14 |
| Binder | 10 | 10 | 10 | 10 | 10 |
| Proximate composition (%) | |||||
| Moisture | 2.9 ± 0.1 | 4.2 ± 0.1 | 5.1 ± 0.1 | 6.5 ± 0.1 | 7.3 ± 0.1 |
| Crude protein, CP | 51.6 ± 0.1 | 50.7 ± 2.6 | 50.4 ± 0.3 | 51.2 ± 1.5 | 50.5 ± 3.1 |
| Crude lipid, CL | 14.4 ± 0.6 | 13.1 ± 0.4 | 12.9 ± 0.4 | 13.2 ± 0.5 | 13.0 ± 0.5 |
| Nitrogen-free extract | 21.3 ± 0.3 | 20.8 ± 1.0 | 20.6 ± 0.5 | 19.0 ± 0.7 | 18.5 ± 1.3 |
| Ash | 9.8 ± 0.2 | 11.1± 0.01 | 11.0 ± 0.00 | 10.1 ± 0.1 | 10.7 ± 0.1 |
| Fatty acid content (as % of total FA) | |||||
| SFA | 27.8 ± 1.3 a | 40.9 ± 0.7 c | 40.0 ± 2.0 c | 35.9 ± 0.7 b | 37.6 ± 2.8 b |
| MUFA | 24.7 ± 0.6 d | 19.8 ± 0.3 b | 19.0 ± 0.9 a | 21.5 ± 0.2 c | 20.0 ± 1.0 b |
| PUFA | 47.4 ± 1.4 c | 39.3 ± 1.0 a | 41.0 ± 1.0 ab | 42.6 ± 0.3 b | 42.4 ± 3.2 b |
| n3 | 38.8 ± 1.4 e | 27.6 ± 0.9 d | 20.8 ± 0.9 c | 15.6 ± 0.3 b | 11.1 ± 3.1 a |
| n6 | 8.6 ± 0.1 a | 11.7 ± 0.3 b | 20.2 ± 0.4 c | 26.9 ± 0.1 d | 31.3 ± 0.9 e |
| n9 | 13.9 ± 0.3 c | 10.7 ± 0.2 a | 12.1 ± 0.7 b | 14.6 ± 0.2 d | 15.2 ± 0.7 e |
| n6/n3 | 0.22 ± 0.05 a | 0.42 ± 0.10 b | 1.00 ± 0.10 c | 1.70 ± 0.10 d | 2.80 ± 0.20 e |
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | References |
|---|---|---|---|
| arpc1a | CTGAACATCTCGCCCTTCTC | TAGCCGATCTGCAGACACAC | [8,16] |
| rpl13 | TCTGGAGGACTGTAAGAGGTATGC | AGACGCACAATCTTGAGAGCAG | [8,16] |
| igf1 | GGCAAATCTCCACGATCTCTAC | CGGTTTCTCTTGTCTCTCTCAG | [8,16,45] |
| igf2a | GAGTCCCATCCATTCTGTTG | GTGGATTGGGGTTTGATGTG | [8,16,45] |
| mstnb | GGACTGGACTGCGATGAG | GATGGGTGTGGGGATACTTC | [8,16,45] |
| nr3c1 | AGACCTTGGTCCCCTTCACT | CGCCTTTAATCATGGGAGAA | [8,16,45] |
| hsp70.1 | TGTTCAGTTCTCTGCCGTTG | AAAGCACTGAGGGACGCTAA | [8,16,45] |
| elovl2 | CACTGGACGAAGTTGGTGAA | GTTGAGGACACACCACCAGA | [8,16,45] |
| elovl5 | TGGATGGGACCGAAATACAT | GTCTCCTCCACTGTGGGTGT | [8,16,45] |
| fads2 | CATCACGCTAAACCCAACA | GGGAGGACCAATGAAGAAGA | [8,16] |
| ghrl | CAGCATGTTTCTGCTCCTGTG | TCTTCTGCCCACTCTTGGTG | [8,16] |
| npy | GTCTGCTTGGGGACTCTCAC | CGGGACTCTGTTTCACCAAT | [8] |
| cnr1 | AGCAAAAGGAGCAACAGGCA | GTTGGTCTGGTACTTTCACTTGAC | [8,16] |
| lepa | CTCCAGTGACGAAGGCAACTT | GGGAAGGAGCCGGAAATGT | [8,16] |
| il1b | GCTGGGGATGTGGACTTC | GTGGATTGGGGTTTGATGTG | [8,16] |
| il10 | ATTTGTGGAGGGCTTTCCTT | AGAGCTGTTGGCAGAATGGT | [8,16] |
| tnfa | TTGTGGTGGGGTTTGATG | TTGGGGCATTTTATTTTGTAAG | [8,16] |
| chia.2 | GGTGCTCTGCCACCTTGCCTT | GGCATGGTTGATCATGGCGAAAGC | [8,16,45] |
| chia.3 | TCGACCCTTACCTTTGCACACACCT’ | ACACCATGATGGAGAACTGTGCCGA | [8,16,45] |
| F1 Zebrafish Larvae | |||||
|---|---|---|---|---|---|
| F1Hi0 | F1Hi25 | F1Hi50 | F1Hi75 | F1Hi100 | |
| 10:0 | 0.010 ± 0.001 | 0.023 ± 0.001 | 0.047 ± 0.004 | 0.052 ± 0.003 | 0.080 ± 0.009 |
| 12:0 | 0.29 ± 0.01 a | 2.70 ± 0.10 b | 4.80 ± 0.30 c | 5.70 ± 0.30 d | 6.40 ± 0.50 e |
| 13:0 | 0.051 ± 0.002 | 0.067 ± 0.002 | 0.084 ± 0.005 | 0.088 ± 0.002 | 0.092 ± 0.008 |
| 14:0 | 4.5 ± 0.3 a | 5.2 ± 0.3 ab | 5.5 ± 0.5 b | 5.4 ± 0.5 b | 5.5 ± 0.5 b |
| 14:1n5 | 0.09 ± 0.01 | 0.17 ± 0.02 | 0.28 ± 0.03 | 0.29 ± 0.03 | 0.26 ± 0.03 |
| 15:0 | 0.78 ± 0.02 | 0.90 ± 0.03 | 0.98 ± 0.04 | 0.98 ± 0.06 | 0.99 ± 0.07 |
| 16:0 | 18.2 ± 0.8 a | 19.6 ± 1.0 ab | 20.3 ± 1.0 b | 20.2 ± 0.7 b | 19.9 ± 0.9 b |
| 16:1n9 | 0.90 ± 0.05 a | 1.10 ± 0.04 b | 1.40 ± 0.10 c | 1.30 ± 0.10 c | 1.30 ± 0.10 c |
| 16:1n7 | 7.0 ± 0.5 a | 7.5 ± 0.4 a | 7.5 ± 0.6 a | 7.2 ± 0.6 a | 7.2 ± 0.5 a |
| 16:2n7 | 0.19 ± 0.02 | 0.20 ± 0.01 | 0.22 ± 0.02 | 0.29 ± 0.03 | 0.28 ± 0.03 |
| 17:0 | 0.80 ± 0.04 a | 0.90 ± 0.05 a | 1.20 ± 0.05 b | 1.30 ± 0.07 b | 1.20 ± 0.09 b |
| 17:1n7 | 0.09 ± 0.01 | 0.29 ± 0.02 | 0.46 ± 0.04 | 0.54 ± 0.04 | 0.54 ± 0.04 |
| 18:0 | 6.6 ± 0.3 a | 6.2 ± 0.4 a | 6.3 ± 0.5 a | 6.1 ± 0.5 a | 6.1 ± 0.4 a |
| 18:1n9 | 12.4 ± 0.6 a | 14.4 ± 1.0 b | 14.9 ± 1.0 b | 14.7 ± 1.2 b | 14.6 ± 1.1 b |
| 18:1n7 | 3.1 ± 0.2 a | 3.6 ± 0.2 a | 4.4 ± 0.3 b | 4.7 ± 0.4 bc | 5.1 ± 0.4 c |
| 18:2n6 | 8.7 ± 0.6 a | 9.0 ± 0.5 a | 8.1 ± 0.6 a | 8.8 ± 0.7 a | 9.0 ± 0.6 a |
| 18:3n3 | 1.3 ± 0.1 a | 1.3 ± 0.1 a | 1.7 ± 0.1 b | 1.9 ± 0.2 b | 1.7 ± 0.2 b |
| 20:0 | 0.32 ± 0.02 | 0.30 ± 0.02 | 0.28 ± 0.02 | 0.29 ± 0.02 | 0.33 ± 0.03 |
| 20:1n9 | 1.06 ± 0.06 | 1.01 ± 0.07 | 0.68 ± 0.04 | 0.60 ± 0.04 | 0.40 ± 0.03 |
| 20:2n6 | 0.31 ± 0.03 | 0.30 ± 0.02 | 0.30 ± 0.03 | 0.37 ± 0.04 | 0.37 ± 0.03 |
| 20:3n6 | 0.42 ± 0.04 | 0.56 ± 0.05 | 0.82 ± 0.08 | 0.92 ± 0.07 | 1.00 ± 0.10 |
| 20:4n6 | 2.0 ± 0.1 a | 2.3 ± 0.1 a | 2.9 ± 0.2 b | 3.1 ± 0.2 bc | 3.3 ± 0.2 c |
| 20:3n3 | 0.11 ± 0.01 | 0.10 ± 0.01 | 0.11 ± 0.01 | 0.10 ± 0.01 | 0.11 ± 0.01 |
| 20:5n3 | 8.6 ± 0.5 c | 5.0 ± 0.4 b | 3.1 ± 0.3 a | 2.8 ± 0.3 a | 2.7 ± 0.2 a |
| 22:0 | 0.17 ± 0.02 c | 0.24 ± 0.03 b | 0.30 ± 0.03 a | 0.25 ± 0.03 a | 0.28 ± 0.03 a |
| 22:1n9 | 0.48 ± 0.05 | 0.47 ± 0.04 | 0.21 ± 0.03 | 0.15 ± 0.03 | 0.03 ± 0.01 |
| 22:6n3 | 21.5 ± 1.0 c | 16.6 ± 1.1 b | 13.1 ± 1.0 a | 12.0 ± 0.9 a | 11.3 ± 0.8 a |
| DHA/EPA | 2.5 ± 0.2 a | 3.3 ± 0.4 b | 4.2 ± 0.5 c | 4.3 ± 0.5 c | 4.2 ± 0.4 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarantoniello, M.; Randazzo, B.; Cardinaletti, G.; Truzzi, C.; Chemello, G.; Riolo, P.; Olivotto, I. Possible Dietary Effects of Insect-Based Diets across Zebrafish (Danio rerio) Generations: A Multidisciplinary Study on the Larval Phase. Animals 2021, 11, 751. https://doi.org/10.3390/ani11030751
Zarantoniello M, Randazzo B, Cardinaletti G, Truzzi C, Chemello G, Riolo P, Olivotto I. Possible Dietary Effects of Insect-Based Diets across Zebrafish (Danio rerio) Generations: A Multidisciplinary Study on the Larval Phase. Animals. 2021; 11(3):751. https://doi.org/10.3390/ani11030751
Chicago/Turabian StyleZarantoniello, Matteo, Basilio Randazzo, Gloriana Cardinaletti, Cristina Truzzi, Giulia Chemello, Paola Riolo, and Ike Olivotto. 2021. "Possible Dietary Effects of Insect-Based Diets across Zebrafish (Danio rerio) Generations: A Multidisciplinary Study on the Larval Phase" Animals 11, no. 3: 751. https://doi.org/10.3390/ani11030751