
First Description of SARS-CoV-2 Infection in Two Feral American Mink (Neovison vison) Caught in the Wild

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
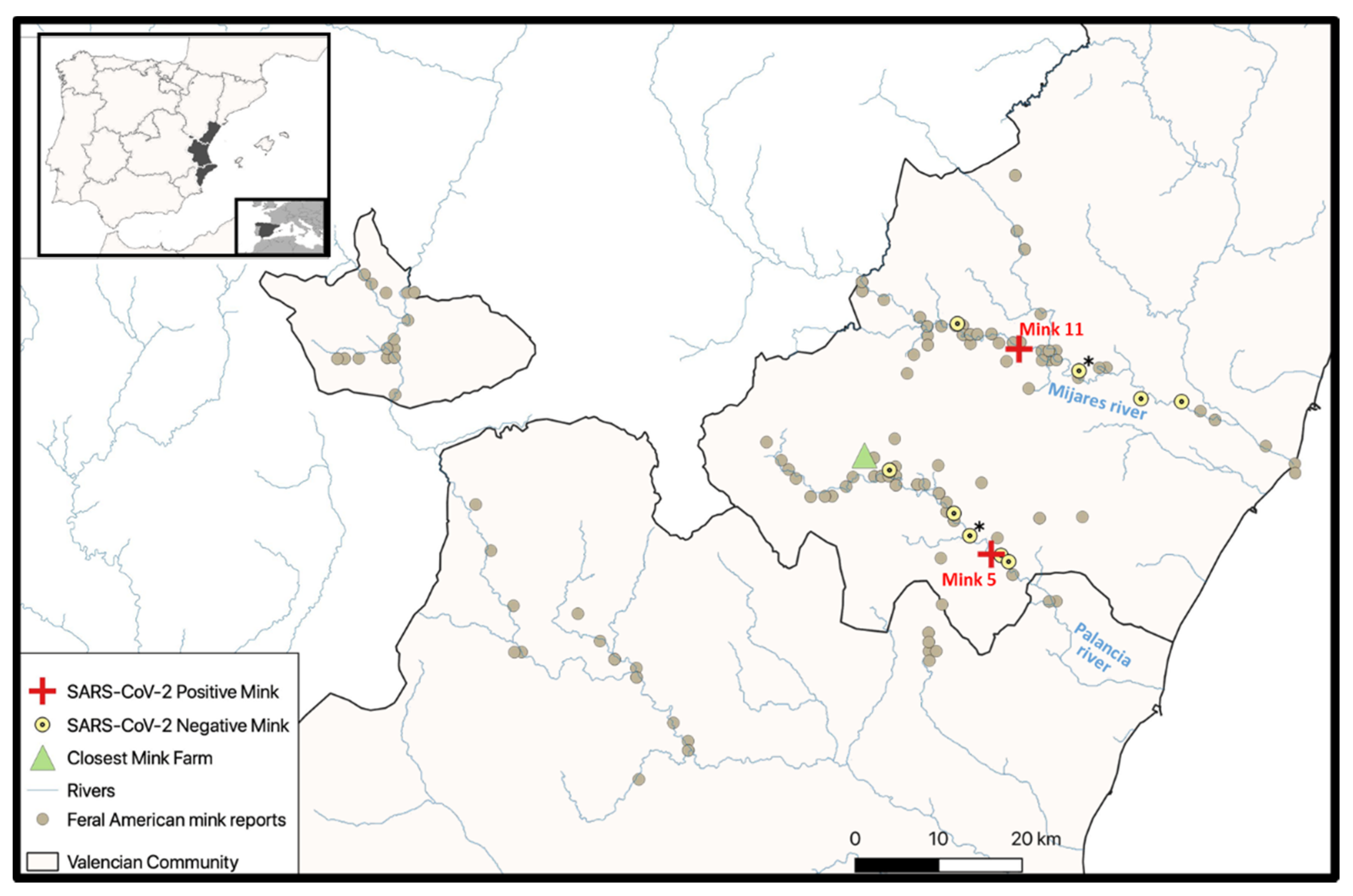
2.1. Study Area
2.2. Sample Collection
2.3. Molecular Analysis
2.4. Sequencing and Phylogenetic Analysis
3. Results
3.1. Animals
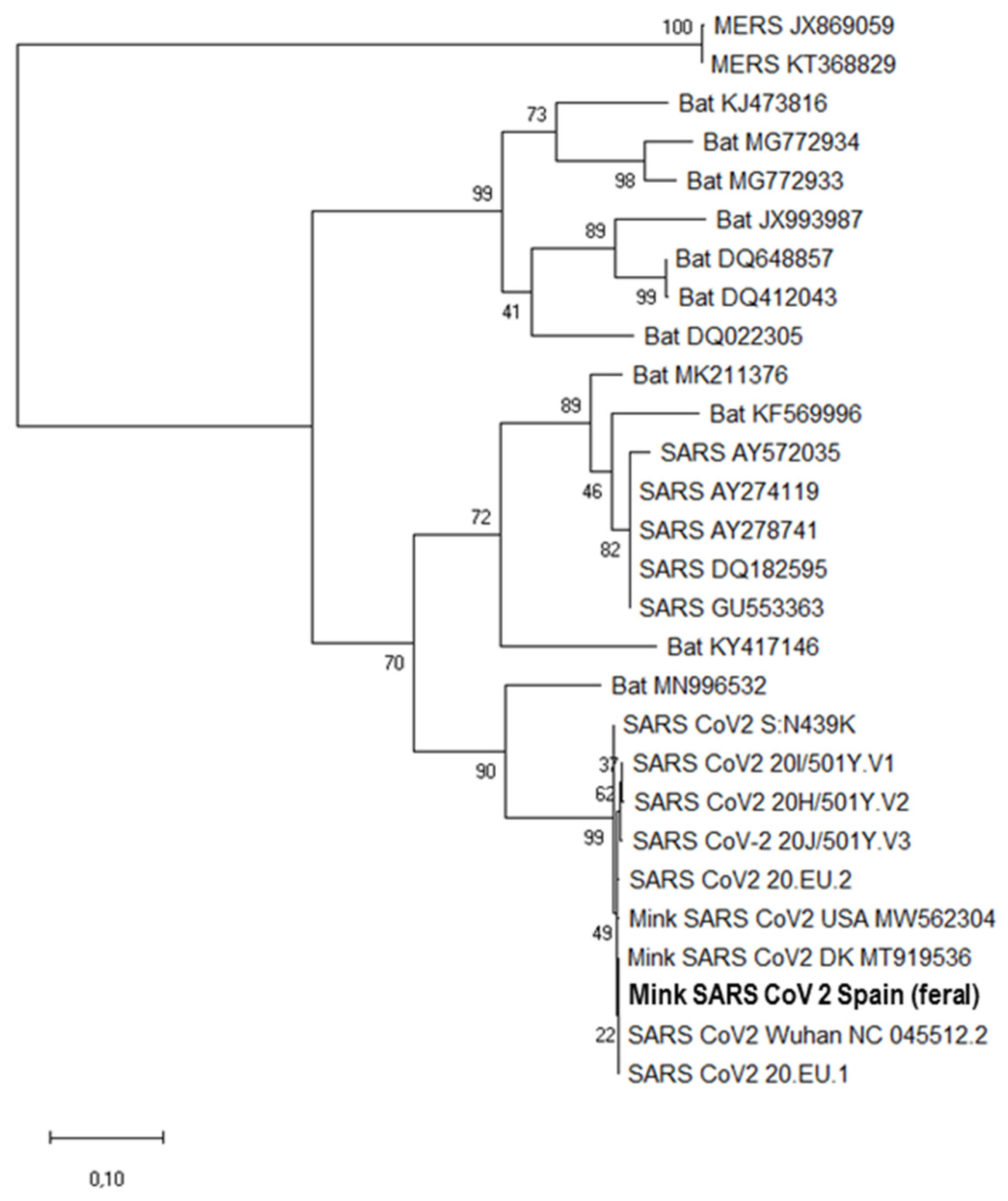
3.2. Molecular, Sequencing, and Phylogenetic Results
4. Discussion
5. Conclusions and Further Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Banerjee, A.; Doxey, A.C.; Mossman, K.; Irving, A.T. Unraveling the zoonotic origin and transmission of SARS-CoV-2. Trends Ecol. Evol. 2021, 36, 180–184. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, C.; Pearce, R.; Omenn, G.S.; Zhang, Y. Identifying the zoonotic origin of SARS-CoV-2 by modeling the binding affinity between the spike receptor-binding domain and host ACE2. J. Proteome Res. 2020, 19, 4844–4856. [Google Scholar] [CrossRef]
- Shereen, M.A.; Khan, S.; Kazmi, A.; Bashir, N.; Siddique, R. COVID-19 infection: Origin, transmission, and characteristics of human coronaviruses. J. Adv. Res. 2020, 24, 91–98. [Google Scholar] [CrossRef]
- Rodriguez-Morales, A.J.; MacGregor, K.; Kanagarajah, S.; Patel, D.; Schlagenhauf, P. Going global—Travel and the 2019 novel coronavirus. Travel Med. Infect. Dis. 2020, 33, 101578. [Google Scholar] [CrossRef]
- Garigliany, M.; Van Laere, A.S.; Clercx, C.; Giet, D.; Escriou, N.; Huon, C.; van der Werf, S.; Eloit, M.; Desmecht, D. SARS-CoV-2 natural transmission from human to cat, Belgium, March 2020. Emerg. Infect. Dis. 2020, 26, 3069–3071. [Google Scholar] [CrossRef]
- Suarez, D.L.; Pantin-Jackwood, M.J.; Swayne, D.E.; Lee, S.A.; DeBlois, S.M.; Spackman, E. Lack of susceptibility to SARS-CoV-2 and MERS-CoV in poultry. Emerg. Infect. Dis. 2020, 26, 3074–3076. [Google Scholar] [CrossRef]
- Pickering, B.S.; Smith, G.; Pinette, M.M.; Embury-Hyatt, C.; Moffat, E.; Marszal, P.; Lewis, C.E. Susceptibility of Domestic Swine to Experimental Infection with Severe Acute Respiratory Syndrome Coronavirus 2. Emerg. Infect. Dis. 2021, 27, 104–112. [Google Scholar] [CrossRef]
- Ulrich, L.; Wernike, K.; Hoffmann, D.; Mettenleiter, T.C.; Beer, M. Experimental infection of cattle with SARS-CoV-2. Emerg. Infect. Dis. 2020, 26, 2979–2981. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Moneim, A.S.; Abdelwhab, E.M. Evidence for SARS-CoV-2 infection of animal hosts. Pathogens 2020, 9, 529. [Google Scholar] [CrossRef]
- McAloose, D.; Laverack, M.; Wang, L.; Killian, M.L.; Caserta, L.C.; Yuan, F.; Mitchell, P.K.; Queen, K.; Mauldin, M.R.; Cronk, B.D.; et al. From people to panthera: Natural SARS-CoV-2 infection in tigers and lions at the Bronx Zoo. mBio 2020, 11, e02220. [Google Scholar] [CrossRef]
- Gortazar, C.; Barroso-Arevalo, S.; Ferreras, E.; Isla, J.; de la Fuente, G.; Rivera-Arroyo, B.; Domínguez, L.; de la Fuente, J.; Sánchez-Vizcaíno, J.M. Natural SARS-CoV-2 infection in kept ferrets, Spain. BioRxiv 2021. [Google Scholar] [CrossRef]
- Giner, J.; Villanueva-Saz, S.; Tobajas, A.P.; Pérez, M.D.; González, A.; Verde, M.; Yzuel, A.; García-García, A.; Taleb, V.; Lira-Navarrete, E.; et al. SARS-CoV-2 seroprevalence in household domestic ferrets (Mustela putorius furo). Animals 2021, 11, 667. [Google Scholar] [CrossRef]
- Enserink, M. Coronavirus rips through Dutch mink farms, triggering culls. Science 2020, 368, 1169. [Google Scholar] [CrossRef]
- Oreshkova, N.; Molenaar, R.J.; Vreman, S.; Harders, F.; Oude Munnink, B.B.; Hakze-van der Honing, R.W.; Gerhards, N.; Tolsma, P.; Bouwstra, R.; Sikkema, R.S.; et al. SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. Eurosurveillance 2020, 25. [Google Scholar] [CrossRef]
- Boklund, A.; Hammer, A.S.; Quaade, M.L.; Rasmussen, T.B.; Lohse, L.; Strandbygaard, B.; Jorgensen, C.S.; Olesen, A.S.; Hjerpe, F.B.; Petersen, H.H.; et al. SARS-CoV-2 in Danish mink farms: Course of the epidemic and a descriptive analysis of the outbreaks in 2020. Animals 2021, 11, 164. [Google Scholar] [CrossRef]
- Hammer, A.S.; Quaade, M.L.; Rasmussen, T.B.; Fonager, J.; Rasmussen, M.; Mundbjerg, K.; Lohse, L.; Strandbygaard, B.; Jorgensen, C.S.; Alfaro-Nunez, A.; et al. SARS-CoV-2 transmission between mink (Neovison vison) and humans, Denmark. Emerg. Infect. Dis. 2021, 27, 547–551. [Google Scholar] [CrossRef]
- ProMED-Mail. Coronavirus Disease 2019 Update (503): Animal, France, Mink, First Report. 2020. Available online: https://promedmail.org/promed-posts/?id=7965554 (accessed on 24 April 2021).
- ProMED-Mail. Coronavirus Disease 2019 Update (490): Animal, Greece (em) Mink, First Report, OIE, Assessment. 2020. Available online: https://promedmail.org/promed-posts/?id=7944705 (accessed on 24 April 2021).
- ProMED-Mail. Coronavirus Disease 2019 Update (458): Animal, Italy, Mink, Request for Information. 2020. Available online: https://promedmail.org/promed-posts/?id=7897986 (accessed on 24 April 2021).
- ProMED-Mail. Coronavirus Disease 2019 Update (510): Animal, Mink, Lithuania, Poland, First Reports, France, OIE. 2020. Available online: https://promedmail.org/promed-posts/?id=7976927 (accessed on 24 April 2021).
- ProMED-Mail. Coronavirus Disease 2019 Update (319): Spain (Aragon) Animal, Farmed Mink, First Report. 2020. Available online: https://promedmail.org/promed-posts/?id=7584560 (accessed on 24 April 2021).
- ProMED-Mail. Coronavirus Disease 2019 Update (468): Animal, Sweden, Mink, First Report, OIE. 2020. Available online: https://promedmail.org/promed-posts/?id=7912846 (accessed on 24 April 2021).
- ProMED-Mail. Coronavirus Disease 2019 Update (526): Canada (British Columbia) Animal, Mink, Human. 2020. Available online: https://promedmail.org/promed-posts/?id=8002622 (accessed on 24 April 2021).
- ProMED-Mail. Coronavirus Disease 2019 Update (366): Animal, USA (Utah) Mink. 2020. Available online: https://promedmail.org/promed-posts/?id=7692815 (accessed on 24 April 2021).
- Shriner, S.A.; Ellis, J.W.; Root, J.J.; Roug, A.; Stopak, S.R.; Wiscomb, G.W.; Zierenberg, J.R.; Ip, H.S.; Torchetti, M.K.; DeLiberto, T.J. SARS-CoV-2 exposure in escaped mink, Utah, USA. Emerg. Infect. Dis. 2021, 27, 988–990. [Google Scholar] [CrossRef]
- Events in Animals. COVID-19 Portal. OIE. Available online: https://www.oie.int/en/scientific-expertise/specific-information-and-recommendations/questions-and-answers-on-2019novel-coronavirus/events-in-animals/ (accessed on 13 April 2021).
- ProMED-Mail. Coronavirus Disease 2019 Update (536): Animal, USA (UT) Wild Mink, 1st Case. 2020. Available online: https://promedmail.org/promed-posts/?id=8015608 (accessed on 24 April 2021).
- Põdra, M.; Gómez, A. Rapid expansion of the American mink poses a serious threat to the European mink in Spain. Mammalia 2018, 82, 580–588. [Google Scholar] [CrossRef]
- Quaglietta, L.; Aulagnier, S. Galemys pyrenaicus. The IUCN Red List of Threatened Species. 2020. e.T8826A114145177. Available online: https://www.iucn.org/resources/conservation-tools/iucn-red-list-threatened-species (accessed on 24 April 2021).
- Rigaux, P.; Vaslin, M.; Noblet, J.F.; Amori, G.; Palomo, L.J. Arvicola sapidus. The IUCN Red List of Threatened Species. ISSN 2307-8235. 2008. e.T2150A9290712. Available online: https://www.iucn.org/resources/conservation-tools/iucn-red-list-threatened-species (accessed on 24 April 2021).
- MAPAMA; Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente. Estrategia de gestión, control y erradicación del visón americano (Neovison vison) en España. Boletín Oficial del Estado. 2014. Available online: https://www.miteco.gob.es/es/biodiversidad/publicaciones/pbl-fauna-flora-estrategias-eei-vison-americano.aspx (accessed on 24 April 2021).
- Pagh, S.; Pertoldi, C.; Chriel, M.; Petersen, H.H.; Jensen, T.H.; Madsen, S.; Kraft, D.C.E.; Schou, T.M.; Hansen, M.S. Estimation of the age and reproductive performance of wild-born and escaped mink (Neovison vison) caught in the wild in Denmark. Animals 2021, 11, 162. [Google Scholar] [CrossRef]
- GBIF.org. GBIF Occurrence Download 11 March 2021. Available online: https://doi.org/10.15468/dl.88uc2j (accessed on 18 March 2021).
- Gaudreault, N.N.; Trujillo, J.D.; Carossino, M.; Meekins, D.A.; Morozov, I.; Madden, D.W.; Indran, S.V.; Bold, D.; Balaraman, V.; Kwon, T.; et al. SARS-CoV-2 infection, disease and transmission in domestic cats. Emerg. Microbes Infect. 2020, 9, 2322–2332. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.L.; Medrano, J.F. Real-time PCR for mRNA quantitation. Biotechniques 2005, 39, 75–85. [Google Scholar] [CrossRef]
- Eftekhari, A.; Alipour, M.; Chodari, L.; Maleki Dizaj, S.; Ardalan, M.; Samiei, M.; Sharifi, S.; Zununi Vahed, S.; Huseynova, I.; Khalilov, R.; et al. Comprehensive Review of Detection Methods for SARS-CoV-2. Microorganisms 2021, 9, 232. [Google Scholar] [CrossRef]
- Freire-Paspuel, B.; Vega-Marino, P.; Velez, A.; Cruz, M.; Perez, F.; Garcia-Bereguiain, M.A. Analytical and clinical comparison of Viasure (CerTest Biotec) and 2019-nCoV CDC (IDT) RT-qPCR kits for SARS-CoV-2 diagnosis. Virology 2021, 553, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Noer, C.L.; Balsby, T.J.S.; Anistoroaei, R.; Stelvig, M.; Dabelsteen, T. Mate choice screening in captive solitary carnivores: The role of male behavior and cues on mate preference and paternity in females of a model species, American mink (Neovison vison). Zoo Biol. 2017, 36, 367–381. [Google Scholar] [CrossRef] [Green Version]
- Melero, Y.; Palazón, S.; Revilla, E.; Martelo, J.; Gosàlbez, J. Space use and habitat preferences of the invasive American mink (Mustela vison) in a Mediterranean area. Eur. J. Wild. Res. 2008, 54, 609–617. [Google Scholar] [CrossRef]
- Tran, H.N.; Le, G.T.; Nguyen, D.T.; Juang, R.S.; Rinklebe, J.; Bhatnagar, A.; Lima, E.C.; Iqbal, H.M.N.; Sarmah, A.K.; Chao, H.P. SARS-CoV-2 coronavirus in water and wastewater: A critical review about presence and concern. Environ. Res. 2021, 193, 110265. [Google Scholar] [CrossRef]
- Dhama, K.; Patel, S.K.; Yatoo, M.I.; Tiwari, R.; Sharun, K.; Dhama, J.; Natesan, S.; Malik, Y.S.; Singh, K.P.; Harapan, H. SARS-CoV-2 existence in sewage and wastewater: A global public health concern? J. Environ. Manag. 2021, 280, 111825. [Google Scholar] [CrossRef] [PubMed]
- Ajuntament de València. Resultados del Análisis de Restos de Coronavirus en Aguas Residuales de València. 2021. Available online: https://www.valencia.es/documents/20142/9932765/0223+Covid+en+aig%C3%BCes+residuals+23+febrer.pdf/4cb8bb49-3e66-9b3e-f805-cfd592c213db?t=1614088620250 (accessed on 24 April 2021).
- Hodcroft, E.B.; Zuber, M.; Nadeau, S.; Crawford, K.H.D.; Bloom, J.D.; Veesler, D.; Vaughan, T.G.; Comas, I.; Candelas, F.G.; Stadler, T.; et al. Emergence and spread of a SARS-CoV-2 variant through Europe in the summer of 2020. medRxiv 2020, 27. [Google Scholar] [CrossRef]
- Manes, C.; Gollakner, R.; Capua, I. Could Mustelids spur COVID-19 into a panzootic? Vet. Ital. 2020, 56, 65–66. [Google Scholar] [CrossRef]
- Gollakner, R.; Capua, I. Is COVID-19 the first pandemic that evolves into panzootic? Vet. Ital. 2020, 56, 11–12. [Google Scholar] [CrossRef]
- Costagliola, A.; Liguori, G.; d’Angelo, D.; Costa, C.; Ciani, F.; Giordano, A. Do animals play a role in the transmission of Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2)? A commentary. Animals 2021, 11, 16. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Animal | ID | Sex | Trapped on | Riverbed | Belongs to | X-Coordinate | Y-Coordinate |
|---|---|---|---|---|---|---|---|
| 1 | 130 | M | 1-Feb-2021 | Palancia | Segorbe | 716,595 | 4,413,436 |
| 2 | 129 | F | 21-Jan-2021 | Mijares | Fanzara | 729,691 | 4,433,165 |
| 3 | 125 | F | 20-Nov-2020 | Mijares | Arañuel | 715,106 | 4,438,807 |
| 4 | 134 | M | 28-Jan-2021 | Palancia | Soneja | 720,334 | 4,411,080 |
| 5 | 136 | M | 28-Jan-2021 | Palancia | Soneja | 719,168 | 4,411,224 |
| 6 | 135 | M | 29-Jan-2021 | Palancia | Segorbe | 716,595 | 4,413,436 |
| 7 | 131 | F | 23-Nov-2020 | Mijares | Onda | 737,126 | 4,429,832 |
| 8 | 127 | F | 20-Nov-2020 | Mijares | Fanzara | 729,691 | 4,433,165 |
| 9 | 132 | M | 19-Jan-2021 | Mijares | Onda | 741,997 | 4,429,482 |
| 10 | 126 | M | 18-Nov-2020 | Palancia | Jérica | 706,960 | 4,421,281 |
| 11 | 137 | M | 14-Jan-2021 | Mijares | Torrechiva | 722,497 | 4,435,770 |
| 12 | 133 | F | 14-Dec-2020 | Palancia | Segorbe | 714,680 | 4,416,113 |
| 13 | 128 | M | 1-Feb-2021 | Palancia | Soneja | 721,249 | 4,410,329 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguiló-Gisbert, J.; Padilla-Blanco, M.; Lizana, V.; Maiques, E.; Muñoz-Baquero, M.; Chillida-Martínez, E.; Cardells, J.; Rubio-Guerri, C. First Description of SARS-CoV-2 Infection in Two Feral American Mink (Neovison vison) Caught in the Wild. Animals 2021, 11, 1422. https://doi.org/10.3390/ani11051422
Aguiló-Gisbert J, Padilla-Blanco M, Lizana V, Maiques E, Muñoz-Baquero M, Chillida-Martínez E, Cardells J, Rubio-Guerri C. First Description of SARS-CoV-2 Infection in Two Feral American Mink (Neovison vison) Caught in the Wild. Animals. 2021; 11(5):1422. https://doi.org/10.3390/ani11051422
Chicago/Turabian StyleAguiló-Gisbert, Jordi, Miguel Padilla-Blanco, Victor Lizana, Elisa Maiques, Marta Muñoz-Baquero, Eva Chillida-Martínez, Jesús Cardells, and Consuelo Rubio-Guerri. 2021. "First Description of SARS-CoV-2 Infection in Two Feral American Mink (Neovison vison) Caught in the Wild" Animals 11, no. 5: 1422. https://doi.org/10.3390/ani11051422