The Effects of Dietary Insect Meal from Hermetia illucens Prepupae on Autochthonous Gut Microbiota of Rainbow Trout (Oncorhynchus mykiss)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Diets
2.3. Feeding Trial and Sampling
2.4. Bacterial DNA Extraction
2.5. Illumina 16S Metagenomic Sequencing Library Construction
2.6. Metagenome Data Analysis
2.7. Statistical Analysis
3. Results
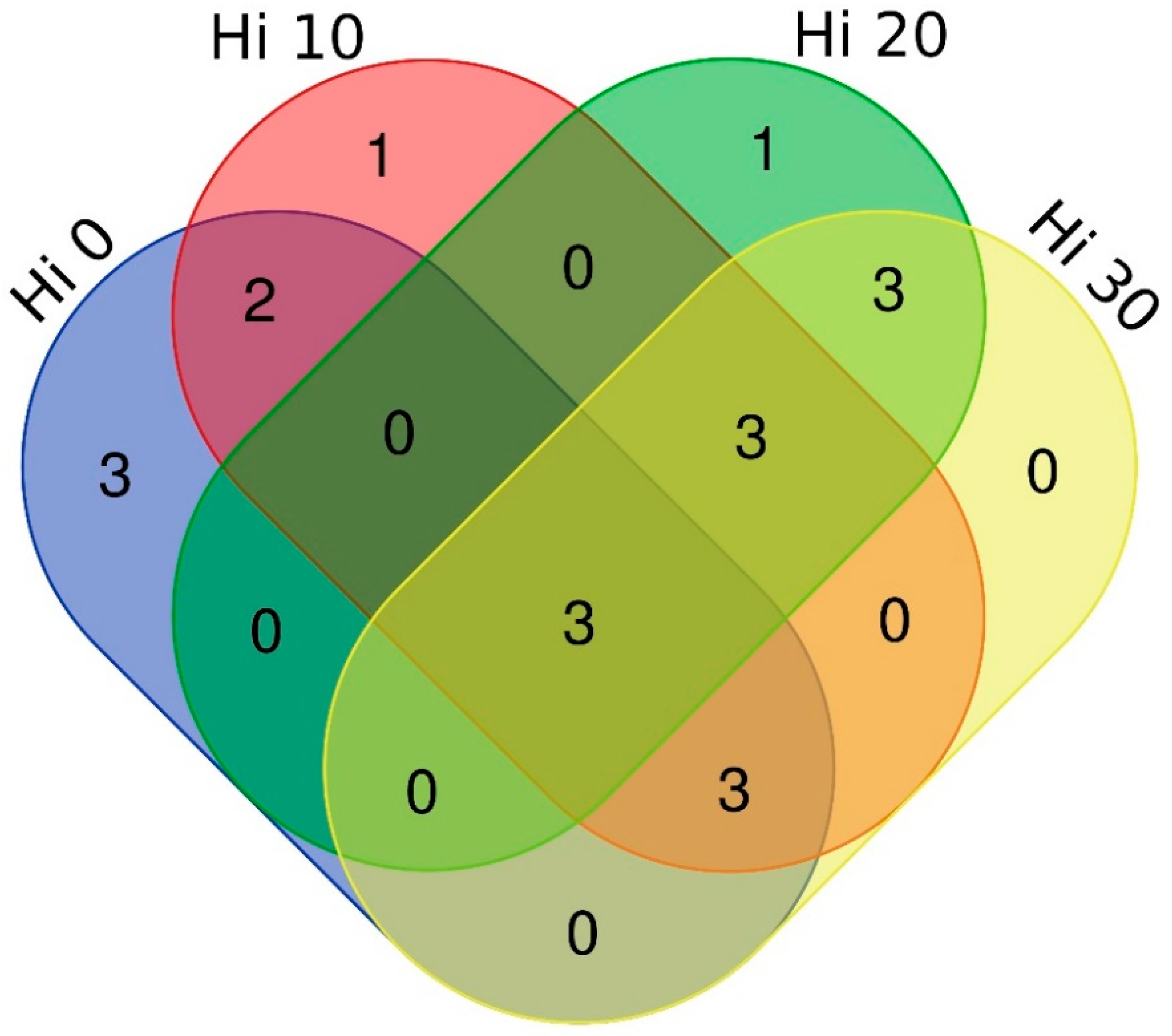
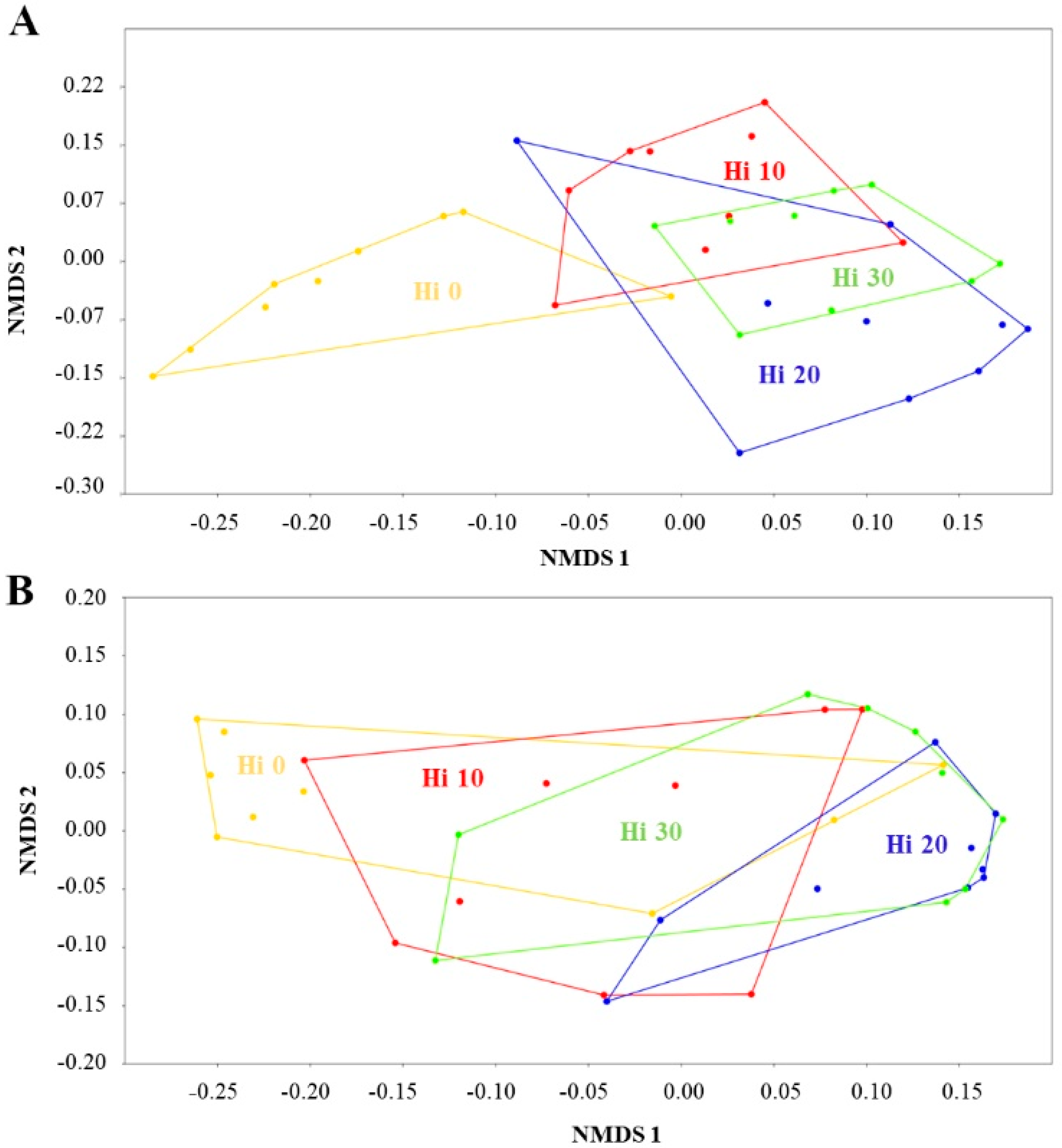
3.1. Structure of Autochthonous Intestinal Bacterial Communities
3.2. Dietary Modulation of Autochthonous Intestinal Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. FAO Edible Insects: Future Prospects for Food and Feed Security; FAO: Rome, Italy, 2013. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture. Contributing to Food Security and Nutrition for All; FAO: Rome, Italy, 2016; p. 200. [Google Scholar]
- Hardy, R.W. Collaborative opportunities between fish nutrition and other disciplines in aquaculture: An overview. Aquaculture 1999, 177, 217–230. [Google Scholar] [CrossRef]
- Naylor, L.R.; Goldburg, J.; Primavera, H.J.; Kautsky, N.; Beveridge, M.C.M.; Clay, J.; Folke, C.; Lubchenco, J.; Mooney, H.; Troell, M. Effect of aquaculture on world fish supplies. Nature 2000, 405, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Tacon, A.G.J.; Metian, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Naylor, L.R.; Hardy, R.W.; Bureau, D.P.; Chiu, A.; Elliot, M.; Farrel, A.P.; Forster, I.; Gatlin, D.M.; Goldburg, R.J.; Hua, K.; et al. Feeding aquaculture in an era of finite resources. Proc. Natl. Acad. Sci. USA 2009, 106, 15103–15110. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y. Replacement of fishmeal with poultry by product meal and hydrolyzed feather meal in feeds for finfish. In Alternative Protein Sources in Aquaculture Diets; Lim, C., Webster, C.D., Lee, C.S., Eds.; The Haworth Press: New York, NY, USA, 2008; pp. 51–93. [Google Scholar]
- Hartviksen, M.; Vecino, J.L.G.; Ringø, E.; Bakke, A.M.; Wadsworth, S.; Krogdahl, Å.; Ruohonen, K.; Kettunen, A. Alternative dietary protein sources for Atlantic salmon (Salmo salar L.) effect on intestinal microbiota, intestinal and liver histology and growth. Aquac. Nut. 2014, 20, 381–398. [Google Scholar] [CrossRef]
- Rimoldi, S.; Terova, G.; Ascione, C.; Giannico, R.; Brambilla, F. Next generation sequencing for intestinal microbiota characterization in rainbow trout (Oncorhynchus mykiss) fed animal by-product meals as an alternative to fishmeal protein sources. PLoS ONE 2018, 13, e0193652. [Google Scholar] [CrossRef]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Ann. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed Sci. Technol. 2015, 203, 1–22. [Google Scholar] [CrossRef]
- Gasco, L.; Gai, F.; Maricchiolo, G.; Genovese, L.; Ragonese, S.; Bottari, T.; Caruso, G. Fishmeal alternative protein sources for aquaculture feeds. In Feeds for the Aquaculture Sector—Current Situation and Alternative Sources. SpringerBriefs in Molecular Science; Springer: Cham, Switzerland, 2018; pp. 1–20. Available online: http://link.springer.com/chapter/10.1007%2F978-3-319-77941-6_1 (accessed on 15 February 2019).
- Terova, G.; Rimoldi, S.; Ascione, C.; Ceccotti, C.; Gini, E.; Gasco, L. Rainbow trout (Oncorhynchus mykiss) gut microbiota is modulated by insect meal from Hermetia illucens prepupae in the diet. Rev. Fish. Biol Fish. 2019, in press. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; van Itterbeeck, J.; Heetkamp, M.J.W.; van den Brand, H.; van Loon, J.J.A.; van Huis, A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE 2010, 5, el14445. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.A.B.; de Boer, I.J.M. Environmental impact of the production of mealworms as a protein source for humans – A life cycle assessment. PLoS ONE 2012, 7, e51145. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Sheppard, C.; Tomberlin, J.K.; Irving, S.; Newton, L.; McGuire, M.A.; Mosley, E.E.; Hardy, R.W.; Sealey, W. Fly prepupae as a feedstuff for rainbow trout, Oncorhynchus mykiss. J. World Aquac. Soc. 2007, 38, 59–67. [Google Scholar] [CrossRef]
- Lock, E.; Arsiwalla, T.; Waagbo, R. Insect larvae meal as an alternative source of nutrients in the diet of Atlantic salmon (Salmo salar) postsmolt. Aquac. Nut. 2016, 22, 1202–1213. [Google Scholar] [CrossRef]
- Magalhaes, R.; Sanchez-Lopez, A.; Leal, R. Black soldier fly (Hermetia illucens L.) prepupae meal as a fish meal replacement in diets for European seabass (Dicentrarchus labrax L.). Aquaculture 2017, 476, 79–85. [Google Scholar] [CrossRef]
- Xiao, X.; Jin, P.; Zheng, L.; Cai, M.; Ziniu, Y.; Yu, J.; Zhang, J. Effects of black soldier fly (Hermetia illucens) larvae meal protein as a fishmeal replacement on the growth and immune index of yellow catfish (Pelteobagrus fulvidraco). Aquac. Res. 2018, 49, 1569–1577. [Google Scholar] [CrossRef]
- Kroeckel, S.; Harjes, A.G.E.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the Black Soldier Fly (Hermetia illucens) as fish meal substitute-growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364, 345–352. [Google Scholar] [CrossRef]
- Renna, M.; Schiavone, A.; Gai, F.; Dabbou, S.; Lussiana, C.; Malfatto, V.; Prearo, M.; Capucchio, M.T.; Biasato, I.; Biasibetti, E. Evaluation of the suitability of a partially defatted black soldier fly (Hermetia illucens L.) larvae meal as ingredient for rainbow trout (Oncorhynchus mykiss) diets. J. Anim. Sci. Biotech. 2017, 8, 57. [Google Scholar] [CrossRef]
- Stadtlander, T.; Stamer, A.; Buser, A.; Wohlfahrt, J.; Leiber, F.; Sandrock, C. Hermetia illucens meal as fish meal replacement for rainbow trout on farm. J. Insects Food Feed 2017, 3, 165–175. [Google Scholar] [CrossRef]
- Bruni, L.; Pastorelli, R.; Viti, C.; Gasco, L.; Parisi, G. Characterization of the intestinal microbial communities of rainbow trout (Oncorhynchus mykiss) fed with Hermetia illucens (black soldier fly) partially defatted larva meal as partial dietary protein source. Aquaculture 2018, 487, 56–63. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Mackay, C.R. Diet, gut microbiota and immune responses. Nat. Immunol. 2010, 12, 5–9. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost microbiomes: The state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.R.; Links, M.G.; Collins, S.A.; Mansfield, G.S.; Drew, M.D.; Van Kessel, A.G.; Hill, J.E. Effects of plant-based diets on the distal gut microbiome of rainbow trout (Oncorhynchus mykiss). Aquaculture 2012, 350–353, 134–142. [Google Scholar] [CrossRef]
- Gajardo, K.; Jaramillo-Torres, A.; Kortner, T.M.; Merrifield, D.L.; Tinsley, J.; Bakke, A.M.; Krogdahl, Å. Alternative protein sources in the diet modulate microbiota and functionality in the distal intestine of Atlantic salmon (Salmo salar). Appl. Environ. Microbiol. 2017, 83, e02615-16. [Google Scholar] [CrossRef] [PubMed]
- Huyben, D.; Nyman, A.; Vidaković, A.; Passoth, V.; Moccia, R.; Kiessling, A.; Dicksved, J.; Lundh, T. Effects of dietary inclusion of the yeasts Saccharomyces cerevisiae and Wickerhamomyces anomalus on gut microbiota of rainbow trout. Aquaculture 2017, 473, 528–537. [Google Scholar] [CrossRef]
- Huyben, D.; Sun, L.; Moccia, R.; Kiessling, A.; Dicksved, J.; Lundh, T. Dietary live yeast and increased water temperature influence the gut microbiota of rainbow trout. J. Appl. Microbiol. 2018, 124, 1377–1392. [Google Scholar] [CrossRef] [PubMed]
- Ingerslev, H.C.; von Gersdorff Jørgensen, L.; Strube, M.L.; Larsen, N.; Dalsgaard, I.; Boye, M.; Madsen, L. The development of the gut microbiota in rainbow trout (Oncorhynchus mykiss) is affected by first feeding and diet type. Aquaculture 2014, 424–425, 24–34. [Google Scholar] [CrossRef]
- Goycoolea, F.; Arguelles-Monal, W.; Peniche, C.; Higuera-Ciapara, I. Chitin and chitosan. In Novel Macromolecules in Food Systems. Developments in Food Science; Doxastakis, G., Kiosseoglou, V., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; Volume 41, pp. 265–308. [Google Scholar]
- Ghanbari, M.; Kneifel, W.; Domig, K.J. A new view of the fish gut microbiome: Advances from next generation sequencing. Aquaculture 2015, 448, 464–475. [Google Scholar] [CrossRef]
- Nawaz, A.; Javaid, A.B.; Irshad, S.; Hoseinifar, S.H.; Xionga, H. The functionality of prebiotics as immunostimulant: Evidences from trials on terrestrial and aquatic animals. Fish. Shellfish Immunol. 2018, 76, 272–278. [Google Scholar] [CrossRef]
- Zhou, Z.; Karlsen, Ø.; He, S.; Olsen, R.E.; Yao, B.; Ringø, E. The effect of dietary chitin on the autochthonous gut bacteria of Atlantic cod (Gadus morhua L.). Aquac. Res. 2013, 44, 1889–1900. [Google Scholar] [CrossRef]
- Qin, C.; Zhang, Y.; Liu, W.; Xu, L.; Yang, Y.; Zhou, Z. Effects of chito-oligosaccharides supplementation on growth performance, intestinal cytokine expression, autochthonous gut bacteria and disease resistance in hybrid tilapia Oreochromis niloticus ♀ X Oreochromis aureus ♂. Fish. Shell Immunol. 2014, 40, 267–274. [Google Scholar] [CrossRef]
- Skrivanova, E.; Marounek, M.; Dlouha, G.; Kanka, J. Susceptibility of Clostridium perfringens to C-C fatty acids. Lett. Appl. Microbiol. 2005, 41, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Skrivanova, E.; Marounek, M.V.B.; Brezina, P. Susceptibility of Escherichia coli, Salmonella sp and Clostridium perfringens to organic acids and monolaurin. Vet. Med. 2006, 51, 81–88. [Google Scholar] [CrossRef]
- Spranghers, T.; Michiels, J.; Vrancx, J.; Ovyn, A.; Eeckhoutc, M.; De Clercq, P.; De Smeta, S. Gut antimicrobial effects and nutritional value of black soldier fly (Hermetia illucens L.) prepupae for weaned piglets. Anim. Feed Sci. Tech. 2018, 235, 33–42. [Google Scholar] [CrossRef]
- Huyben, D.; Vidaković, A.; Werner Hallgren, S.; Langeland, M. High-throughput sequencing of gut microbiota in rainbow trout (Oncorhynchus mykiss) fed larval and pre-pupae stages of black soldier fly (Hermetia illucens). Aquaculture 2019, 500, 485–491. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a Prokaryotic Universal Primer for Simultaneous Analysis of Bacteria and Archaea Using Next-Generation Sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Paleontological Statistics Software Package for education and data analysis. Palaeontol. Electronica 2001, 4, 9–18. [Google Scholar]
- Van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed—A review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Gasco, L.; Finke, M.; van Huis, A. Can diets containing insects promote animal health? J. Insects Food Feed 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Lock, E.J.; Biancarosa, I.; Gasco, L. Insects as Raw Materials in Compound Feed for Aquaculture. In Edible insects in Sustainable Food System; Halloran, A., Flore, R., Vantomme, P., Roos, N., Eds.; Springer International Publishing AG: Basel, Switzerland, 2018; pp. 263–276. [Google Scholar]
- Lyons, P.P.; Turnbull, J.F.; Dawson, K.A.; Crumlish, M. Phylogenetic and functional characterization of the distal intestinal microbiome of rainbow trout Oncorhynchus mykiss from both farm and aquarium settings. J. Appl. Microbiol. 2016, 122, 347–363. [Google Scholar] [CrossRef]
- Lyons, P.P.; Turnbull, J.F.; Dawson, K.A.; Crumlish, M. Effects of low-level dietary microalgae supplementation on the distal intestinal microbiome of farmed rainbow trout Oncorhynchus mykiss (Walbaum). Aquac. Res. 2017, 48, 2438–2452. [Google Scholar] [CrossRef]
- Kim, D.H.; Brunt, J.; Austin, B. Microbial diversity of intestinal contents and mucus in rainbow trout (Oncorhynchus mykiss). J. Appl. Microbiol. 2007, 102, 1654–1664. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Dimitroglou, A.; Bradley, G.; Baker, T.M.; Davies, S.J. Soybean meal alters autochthonous microbial populations, microvilli morphology and compromises intestinal enterocyte integrity of rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish. Diseases 2009, 32, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Etyemez, M.; Balcázar, J.L. Bacterial community structure in the intestinal ecosystem of rainbow trout (Oncorhynchus mykiss) as revealed by pyrosequencing-based analysis of 16S rRNA genes. Res. Vet. Sci. 2015, 100, 8–11. [Google Scholar] [CrossRef]
- Askarian, F.; Zhou, Z.; Olsen, R.E.; Sperstad, S.; Ringo, E. Culturable autochthonous gut bacteria in Atlantic salmon (Salmo salar L.) fed diets with or without chitin. Characterization by 16S rRNA gene sequencing, ability to produce enzymes and in vitro growth inhibition of four fish pathogens. Acquaculture 2012, 326, 1–8. [Google Scholar] [CrossRef]
- Ringø, E.; Zhou, Z.; Olsen, R.E.; Song, S.K. Use of chitin and krill in aquaculture—The effect on gut microbiota and the immune system: A review. Aquac. Nut. 2012, 18, 117–131. [Google Scholar] [CrossRef]
- Cerezuela, R.; Fumanal, M.; Tapia-Paniagua, S.T.; Meseguer, J.; Moriñigo, M.Á.; Esteban, M.Á. Changes in intestinal morphology and microbiota caused by dietary administration of inulin and Bacillus subtilis in gilthead sea bream (Sparus aurata L.) specimens. Fish. Shellfish Immunol. 2013, 34, 1063–1070. [Google Scholar] [CrossRef]
- Apper, E.; Weissman, D.; Respondek, F.; Guyonvarch, A.; Baron, F.; Boisot, P.; Rodiles, A.; Merrifield, D.L. Hydrolysed wheat gluten as part of a diet based on animal and plant proteins supports good growth performance of Asian seabass (Lates calcarifer), without impairing intestinal morphology or microbiota. Aquaculture 2016, 453, 40–48. [Google Scholar] [CrossRef]
- Dailey, F.E.; McGraw, J.E.; Jensen, B.J.; Bishop, S.S.; Lokken, J.P.; Dorff, K.J.; Ripley, M.P.; Munro, J.B. The microbiota of freshwater fish and freshwater niches contain omega-3 fatty acid-producing Shewanella species. Appl. Environ. Microbiol. 2016, 82, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Satomi, M.; Oikawa, H.; Yano, Y. Shewanella marinintestina sp. nov., Shewanella schlegeliana sp. nov. and Shewanella sairae sp. nov., novel eicosapentaenoic-acid-producing marine bacteria isolated from sea-animal intestines. Int. J. Syst. Evol. Microbiol. 2003, 53, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.; Müller, A.; Heckel, D.G.; Gutzeit, H.; Vilcinskas, A. Nutritional immunology: Diversification and diet-dependent expression of antimicrobial peptides in the black soldier fly Hermetia illucens. Dev. Comp. Immunol. 2018, 78, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Freundt, E.A.; Razin, S. Genus Mycoplasma. In Bergey’s Manual of Systematic Bacteriology; Krieg, N.R., Holt, J.G., Eds.; Williams & Wilkins: Baltimore, Maryland, 1958; Volume 1, pp. 742–770. [Google Scholar]
- Brown, R.M.; Wiens, G.D.; Salinas, I. Analysis of the gut and gill microbiome of resistant and susceptible lines of rainbow trout (Onchorynchus mykiss). Fish Shellfish Immunol. 2019, 86, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Ciric, M.; Waite, D.; Draper, J.; Jones, J.B. Characterisation of gut microbiota of farmed Chinook salmon using metabarcoding. BioRxiv 2018, 288761. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H. illucens Meal | DIET | ||||
|---|---|---|---|---|---|
| Hi 0 | Hi 10 | Hi 20 | Hi 30 | ||
| Ingredients (% as it) | |||||
| Fishmeal a | 60 | 54 | 48 | 42 | |
| Hermetia illucens meal | 0 | 10 | 20 | 30 | |
| Fish oil | 7 | 7 | 7 | 7 | |
| Soybean oil | 5 | 4 | 3 | 2 | |
| Wheat bran | 10 | 7 | 4 | 1 | |
| Wheat meal | 4 | 4 | 4 | 4 | |
| Starch gelatinized D500 | 11 | 11 | 11 | 11 | |
| Vitamin premix b | 1.5 | 1.5 | 1.5 | 1.5 | |
| Mineral premix c | 1.5 | 1.5 | 1.5 | 1.5 | |
| Proximate composition (g/100g DM) | |||||
| DM | 89.82 | 89.03 | 88.62 | 87.70 | 86.74 |
| CP | 48.62 | 49.07 | 49.88 | 49.48 | 49.03 |
| EE | 20.58 | 17.73 | 17.98 | 17.95 | 17.56 |
| Ash | 8.74 | 13.71 | 14.16 | 14.48 | 14.26 |
| CHI | 4.96 | 0.00 | 0.50 | 0.99 | 1.51 |
| NFE | 17.10 | 19.48 | 17.49 | 17.10 | 17.64 |
| Items | Hi 0 | Hi 10 | Hi 20 | Hi 30 |
|---|---|---|---|---|
| Reads | 43,654 ± 18,004 | 46,206 ± 14,511 | 37,146 ± 6815 | 41,233 ± 9479 |
| Observed OTUs | 24.3 ± 9.4 | 24.7 ± 3.9 | 23.9 ± 6.9 | 30.0 ± 10.9 |
| Chao 1 | 27.1 ± 9.9 | 26.8 ± 5.1 | 30.6 ± 12.0 | 36.5 ± 10.6 |
| Shannon_e | 1.0 ± 0.4 a | 0.7 ± 0.4 a | 0.2 ± 0.3 b | 0.3 ± 0.3 b |
| Simpson | 0.5 ± 0.2 b | 0.6 ± 0.2 b | 0.9 ± 0.2 a | 0.9 ± 0.2 a |
| Berger_parker (1/d) | 0.6 ± 0.2 b | 0.7 ± 0.2 b | 0.9 ± 0.1 a | 0.9 ± 0.1 a |
| Statistics | Bray–Curtis | Binary Bray–Curtis | ||
|---|---|---|---|---|
| PERMANOVA | ||||
| Permutation N | 999 | 999 | ||
| Total sum of squares | 1.383 | 0.448 | ||
| Within-group sum of squares | 0.929 | 0.274 | ||
| F | 5.202 | 6.785 | ||
| p (same) | 0.001 | 0.001 | ||
| Pairwise comparisons | p-value | F-value | p-value | F-value |
| Hi 10 vs. Hi 0 | 0.708 | 2.144 | 0.006 | 9.156 |
| Hi 20 vs. Hi 0 | 0.012 | 12.350 | 0.012 | 9.849 |
| Hi 30 vs. Hi 0 | 0.078 | 7.191 | 0.006 | 12.450 |
| Hi 10 vs. Hi 20 | 0.042 | 5.824 | 0.030 | 5.189 |
| Hi 10 vs. Hi 30 | 0.486 | 2.691 | 0.156 | 2.490 |
| Hi 20 vs. Hi 30 | 1.000 | 1.052 | 0.564 | 1.934 |
| ANOSIM | ||||
| Permutation N | 999 | 999 | ||
| R | 0.252 | 0.480 | ||
| p (same) | 0.002 | 0.001 | ||
| Pairwise comparisons | p-value | R | p-value | R |
| Hi 10 vs. Hi 0 | 0.468 | 0.133 | 0.006 | 0.585 |
| Hi 20 vs. Hi 0 | 0.018 | 0.515 | 0.006 | 0.703 |
| Hi 30 vs. Hi 0 | 0.036 | 0.386 | 0.006 | 0.765 |
| Hi 10 vs. Hi 20 | 0.042 | 0.344 | 0.030 | 0.425 |
| Hi 10 vs. Hi 30 | 0.534 | 0.130 | 0.348 | 0.159 |
| Hi 20 vs. Hi 30 | 1.000 | 0.008 | 0.180 | 0.190 |
| SIMPER | ||||
| Hi 10 vs. Hi 0 | 26.16 | 16.77 | ||
| Hi 20 vs. Hi 0 | 32.75 | 19.23 | ||
| Hi 30 vs. Hi 0 | 31.27 | 18.33 | ||
| Hi 10 vs. Hi 20 | 25.81 | 16.09 | ||
| Hi 10 vs. Hi 30 | 24.79 | 12.98 | ||
| Hi 20 vs. Hi 30 | 19.24 | 13.85 | ||
| Taxa | Hi 0 | Hi 10 | Hi 20 | Hi 30 |
|---|---|---|---|---|
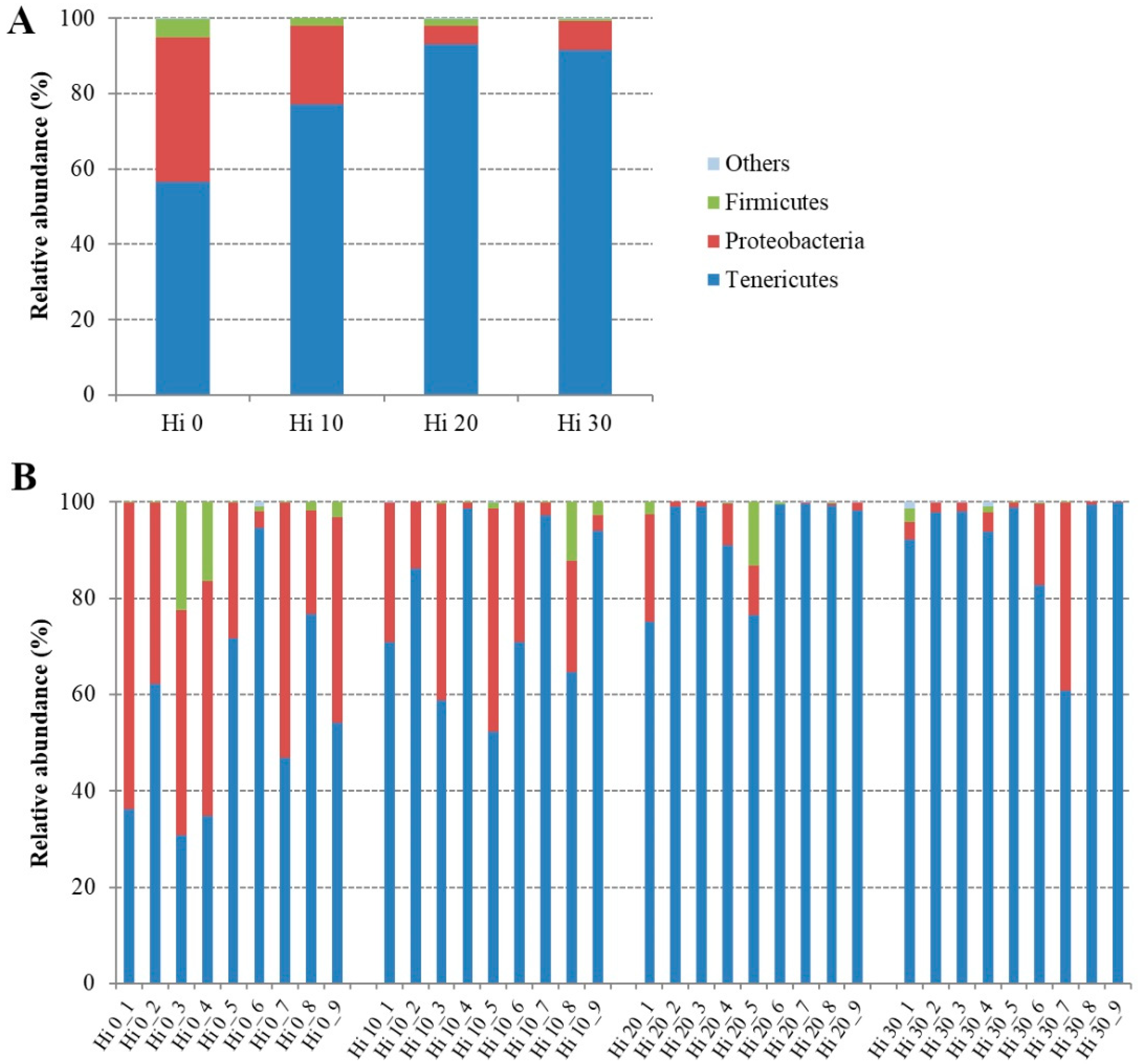
| Phylum | ||||
| Tenericutes | 56.48 ± 21.74 b | 77.06 ± 17.44 ab | 93.03 ± 10.12 a | 91.52 ± 12.68 a |
| Proteobacteria | 38.36 ± 18.26 a | 21.07 ± 16.85 ab | 5.03 ± 7.51 b | 7.69 ± 12.82 b |
| Firmicutes | 5.05 ± 8.37 | 1.84 ± 4.00 | 1.86 ± 4.34 | 0.49 ± 0.96 |
| Class | ||||
| Mollicutes | 56.48 ± 21.74 b | 77.06 ± 17.44 ab | 93.03 ± 10.12 a | 91.52 ± 12.68 a |
| Gammaproteobacteria | 36.20 ± 18.58 a | 20.65 ± 16.55 ab | 4.98 ± 7.46 b | 7.57 ± 12.78 b |
| Betaproteobacteria | 5.01 ± 8.39 a | 1.82 ± 4.00 a | 1.81 ± 4.34 b | 0.37 ± 0.78 ab |
| Clostridia | 2.15 ± 2.85 | 0.41 ± 0.39 | 0.03 ± 0.07 | 0.10 ± 0.20 |
| Order | ||||
| Mycoplasmatales | 56.48 ± 21.74 b | 77.06 ± 17.44 ab | 93.03 ± 10.12 a | 91.52 ± 12.68 a |
| Alteromonadales | 28.69 ± 20.77 | 15.85 ± 13.38 | 4.74 ± 7.46 | 5.32 ± 12.69 |
| Aeromonadales | 2.30 ± 2.87 a | 1.67 ± 3.10 ab | 0.12 ± 0.30 ab | 0.11 ± 0.30 b |
| Neisseriales | 2.15 ± 2.85 a | 0.41 ± 0.39 a | 0.03 ± 0.07 b | 0.10 ± 0.20 ab |
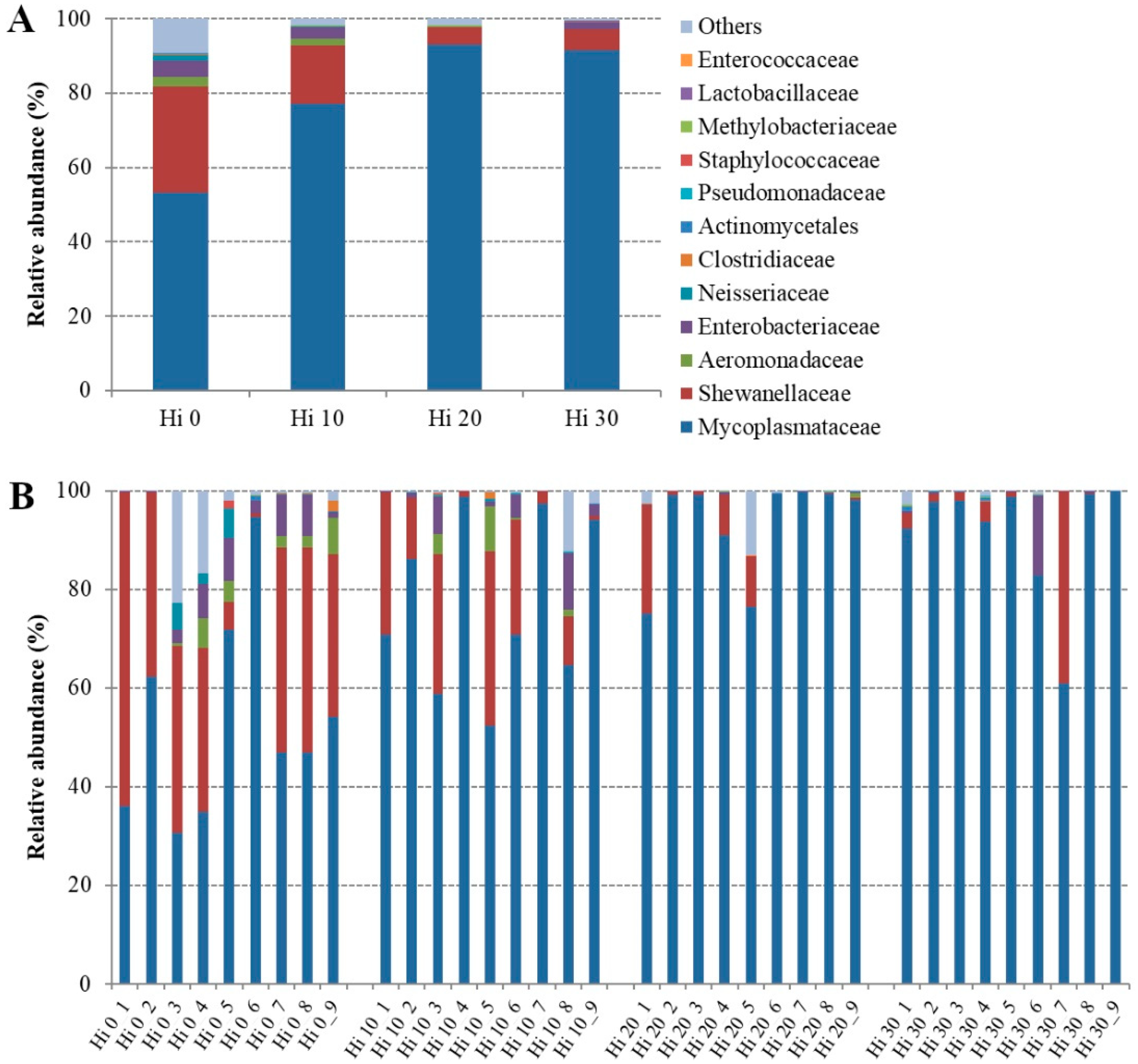
| Family | ||||
| Mycoplasmataceae | 53.16 ± 20.49 b | 77.06 ± 17.44 ab | 93.03 ± 10.12 a | 91.52 ± 12.68 a |
| Shewanellaceae | 28.69 ± 20.77 a | 15.85 ± 13.38 ab | 4.74 ± 7.46 b | 5.62 ± 12.59 ab |
| Aeromonadaceae | 2.53 ± 2.77 a | 1.67 ± 3.10 ab | 0.12 ± 0.30 b | 0.01 ± 0.02 b |
| Enterobacteriaceae | 4.31 ± 3.67 a | 3.13 ± 4.01ab | 0.13 ± 0.16 b | 1.95 ± 5.33 ab |
| Neisseriaceae | 1.60 ± 2.45 a | 0.22 ± 0.21 b | 0.02 ± 0.05 b | 0.01 ± 0.02 ab |
| Clostridiaceae | 0.31 ± 0.70 | 0.17 ± 0.42 | 0.01 ± 0.01 | 0.02 ± 0.03 |
| Actinomycetales | 0.11 ± 0.30 | 0.03 ± 0.02 | 0.06 ± 0.06 | 0.23 ± 0.35 |
| Pseudomonadaceae | 0.16 ± 0.47 | 0.00 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Staphylococcaceae | 0.03 ± 0.07 | 0.01 ± 0.01 | 0.03 ± 0.04 | 0.07 ± 0.12 |
| Methylobacteriaceae | 0.00 ± 0.01 | 0.01 ± 0.01 | 0.02 ± 0.02 | 0.01 ± 0.01 |
| Lactobacillaceae | 0.00 ± 0.00 | 0.00 ± 0.01 | 0.00 ± 0.01 | 0.02 ± 0.03 |
| Enterococcaceae | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.02 |
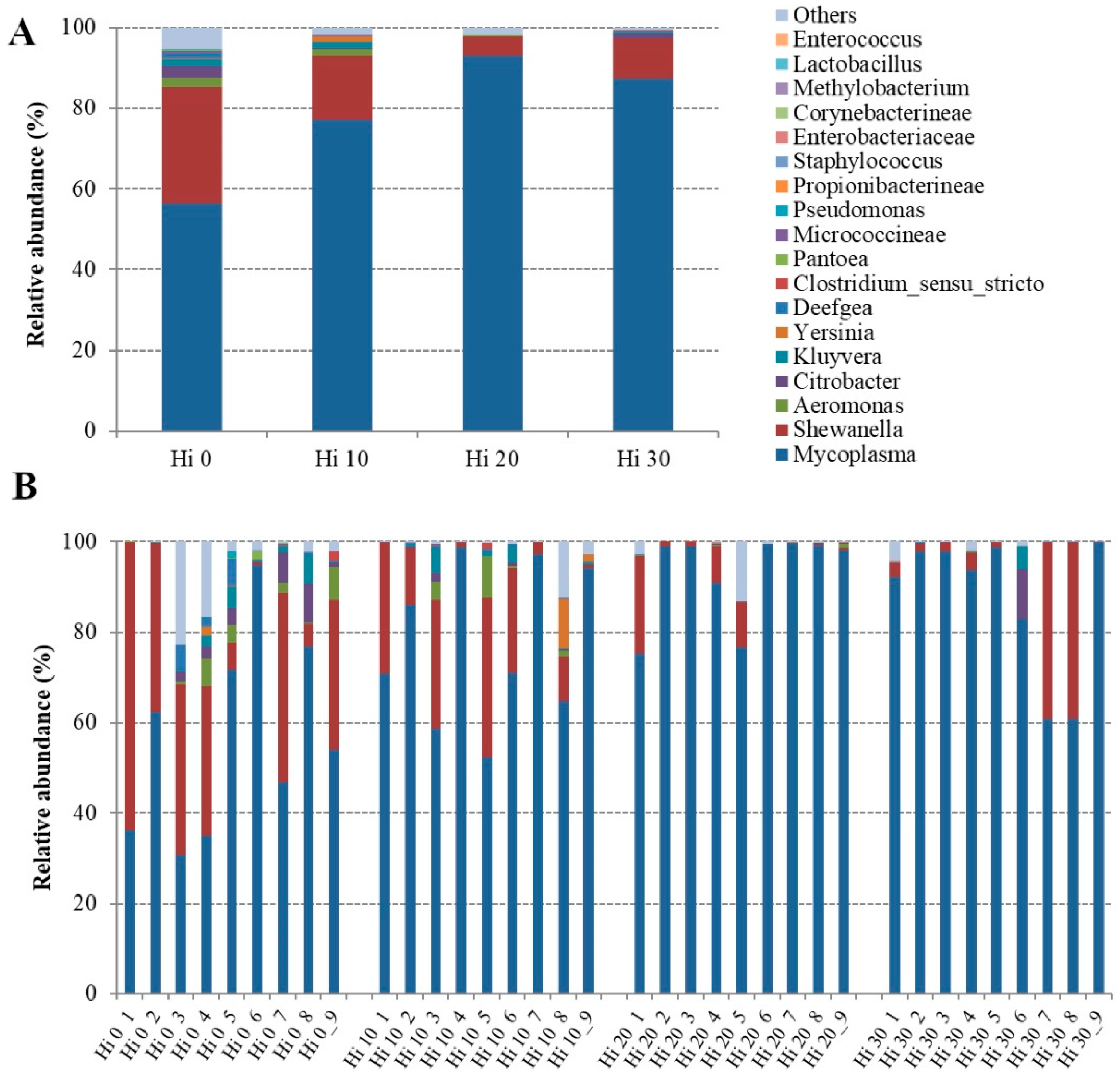
| Genus | ||||
| Mycoplasma | 56.48 ± 21.74 b | 77.06 ± 17.44 ab | 93.03 ± 10.12 a | 87.24 ± 15.81 ab |
| Shewanella | 28.69 ± 20.77 a | 15.85 ± 13.38 ab | 4.74 ± 7.46 b | 9.93 ± 16.53 ab |
| Aeromonas | 2.30 ± 2.87 a | 1.67 ± 3.10 ab | 0.12 ± 0.30 ab | 0.01 ± 0.02 b |
| Citrobacter | 2.83 ± 3.15 a | 0.41 ± 0.62 ab | 0.04 ± 0.06 b | 1.28 ± 3.69 ab |
| Kluyvera | 1.72 ± 2.23 a | 1.31 ± 2.02 ab | 0.01 ± 0.02 b | 0.57 ± 1.67 ab |
| Yersinia | 0.23 ± 0.58 | 1.40 ± 3.64 | 0.07 ± 0.11 | 0.00 ± 0.01 |
| Deefgea | 1.66 ± 2.42 a | 0.22 ± 0.21 a | 0.02 ± 0.05 b | 0.01 ± 0.02 ab |
| Clostridium_sensu_stricto | 0.28 ± 0.71 | 0.17 ± 0.42 | 0.01 ± 0.01 | 0.02 ± 0.03 |
| Pantoea | 0.27 ± 0.66 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.02 |
| Pseudomonas | 0.16 ± 0.47 | 0.00 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Staphylococcus | 0.03 ± 0.07 | 0.01 ± 0.01 | 0.03 ± 0.04 | 0.07 ± 0.11 |
| Methylobacterium | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.02 ± 0.02 | 0.01 ± 0.01 |
| Lactobacillus | 0.00 ± 0.00 | 0.00 ± 0.01 | 0.00 ± 0.01 | 0.02 ± 0.03 |
| Enterococcus | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rimoldi, S.; Gini, E.; Iannini, F.; Gasco, L.; Terova, G. The Effects of Dietary Insect Meal from Hermetia illucens Prepupae on Autochthonous Gut Microbiota of Rainbow Trout (Oncorhynchus mykiss). Animals 2019, 9, 143. https://doi.org/10.3390/ani9040143
Rimoldi S, Gini E, Iannini F, Gasco L, Terova G. The Effects of Dietary Insect Meal from Hermetia illucens Prepupae on Autochthonous Gut Microbiota of Rainbow Trout (Oncorhynchus mykiss). Animals. 2019; 9(4):143. https://doi.org/10.3390/ani9040143
Chicago/Turabian StyleRimoldi, Simona, Elisabetta Gini, Federica Iannini, Laura Gasco, and Genciana Terova. 2019. "The Effects of Dietary Insect Meal from Hermetia illucens Prepupae on Autochthonous Gut Microbiota of Rainbow Trout (Oncorhynchus mykiss)" Animals 9, no. 4: 143. https://doi.org/10.3390/ani9040143