
The Intestinal Carriage of Plasmid-Mediated Colistin-Resistant Enterobacteriaceae in Tertiary Care Settings

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. The Prevalence of the mcr-1 Gene Was Low, and the mcr-2 to -8 Genes Were Not Detected
2.2. Whole-Genome Analysis Data Revealed a High Diversity of mcr-1 Positive E. coli Isolates and the Predominance of the IncX4 Plasmid Type as a mcr-1 Gene Vector
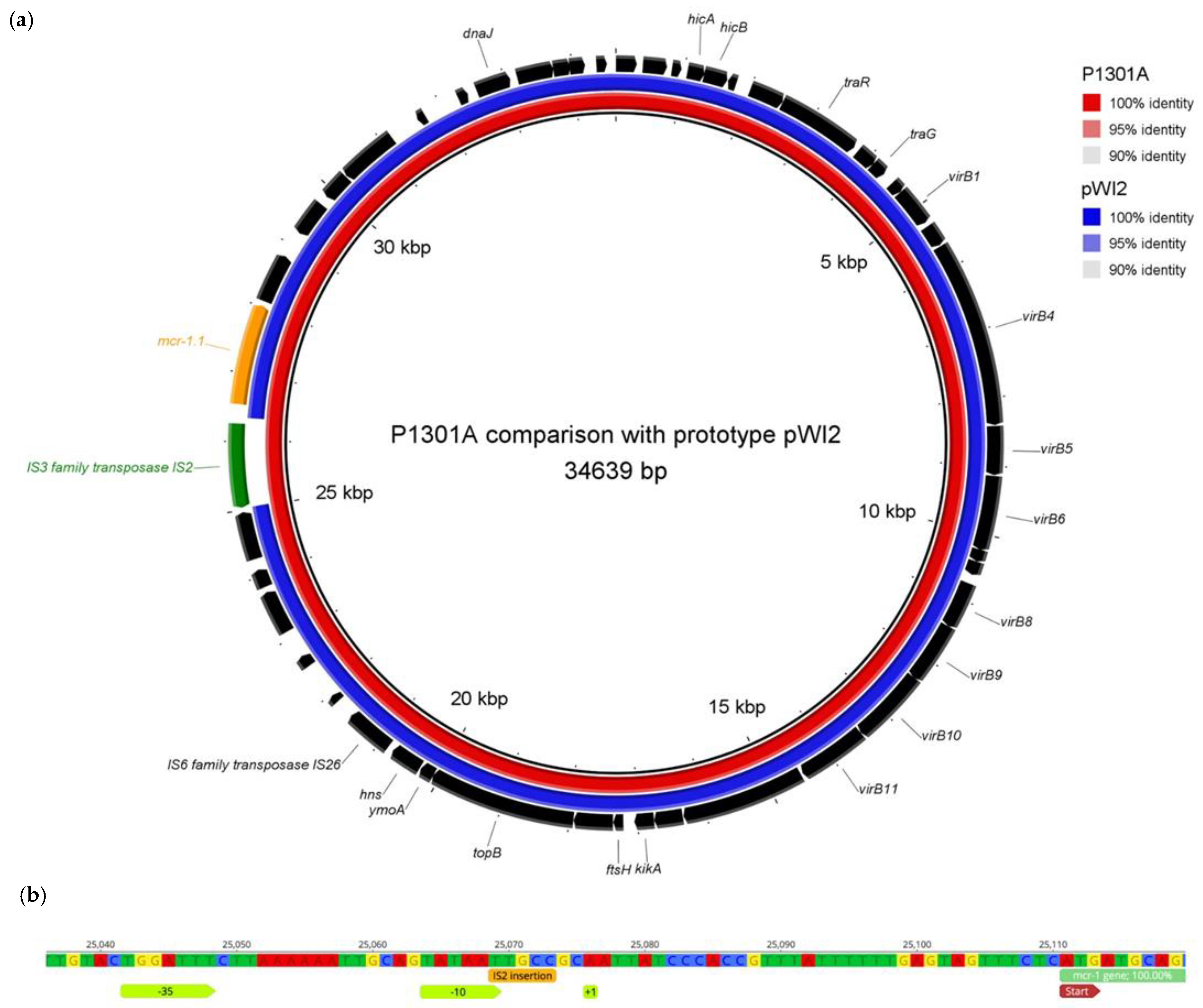
2.3. Colistin Resistance in the mcr-1 Positive/Colistin-Susceptible Isolate Was Inducible
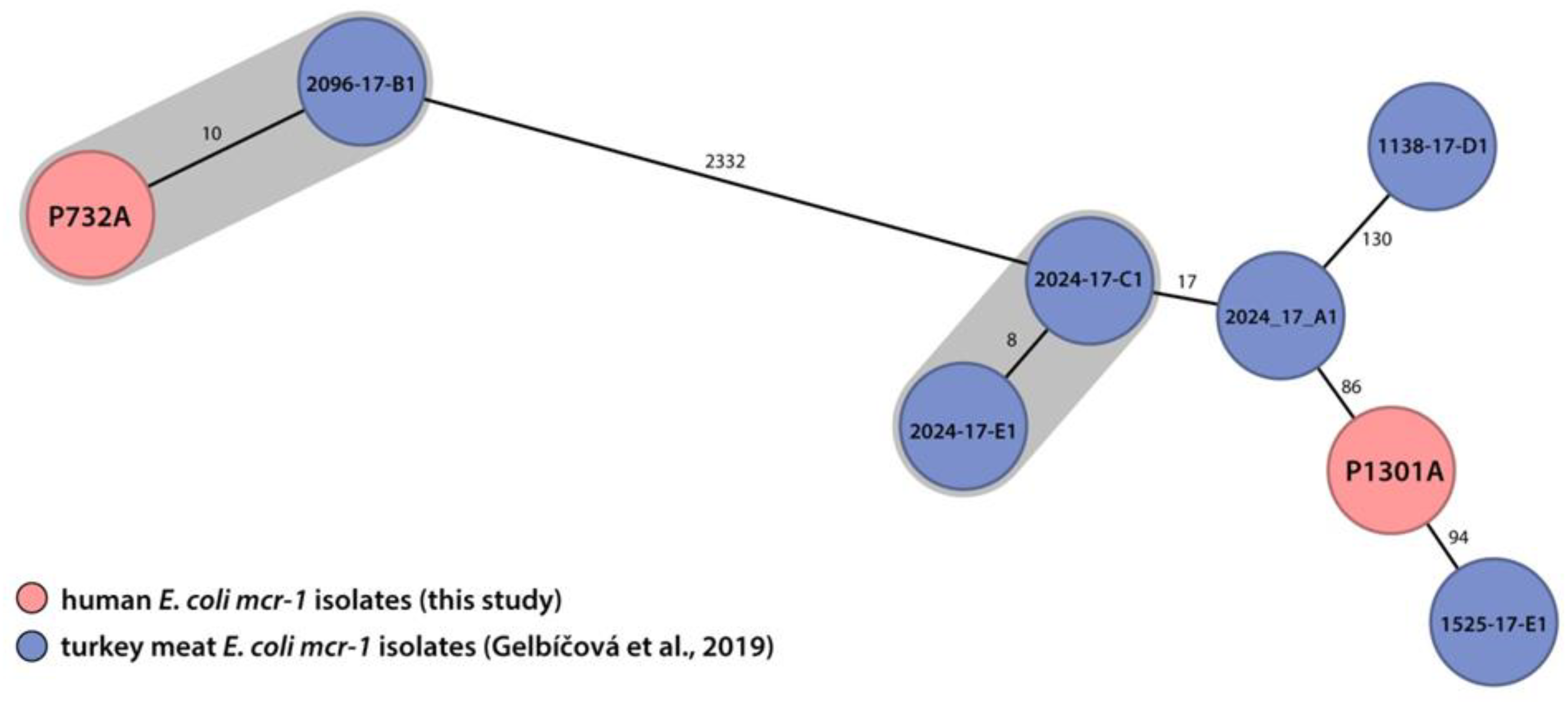
2.4. The Human mcr-1 E. coli Isolate Was Genetically Related to the mcr-1 E. coli Isolate from Turkey Meat Produced Domestically
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Screening for the mcr-Mediated Colistin Resistance
4.3. Whole-Genome Sequence Data Analysis of mcr-Positive Isolates
4.4. Colistin Resistance Induction Assay in a mcr-Positive Colistin-Susceptible Isolate
4.5. Confirmation of MCR-1 Expression
4.6. Antimicrobial Resistance Profile Determination
4.7. Comparative Analysis of Czech mcr-1 E. coli Isolates from Humans and Food of Animal Origin
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef]
- Skov, R.L.; Monnet, D.L. Plasmid-mediated colistin resistance (mcr-1 gene): Three months later, the story unfolds. Eurosurveillance 2016, 21, 30155. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- El-Sayed Ahmed, M.A.E.G.; Zhong, L.L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.B. Colistin and its role in the Era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of novel mobile colistin resistance gene mcr-10. Emerg. Microbes Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef]
- Nang, S.C.; Li, J.; Velkov, T. The rise and spread of mcr plasmid-mediated polymyxin resistance. Crit. Rev. Microbiol. 2019, 45, 131–161. [Google Scholar] [CrossRef]
- Shen, Y.; Zhou, H.; Xu, J.; Wang, Y.; Zhang, Q.; Walsh, T.R.; Shao, B.; Wu, C.; Hu, Y.; Yang, L.; et al. Anthropogenic and environmental factors associated with high incidence of mcr-1 carriage in humans across China. Nat. Microbiol. 2018, 3, 1054–1062. [Google Scholar] [CrossRef]
- European Medicines Agency. Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2018: Trends from 2010 to 2018; Tenth European Surveillance of Veterinary Antimicrobial Consumption (ESVAC) Report; European Medicines Agency: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Shen, C.; Zhong, L.-L.; Yang, Y.; Doi, Y.; Paterson, D.L.; Stoesser, N.; Ma, F.; El-Sayed Ahmed, M.A.E.-G.; Feng, S.; Huang, S.; et al. Dynamics of mcr-1 prevalence and mcr-1-positive Escherichia coli after the cessation of colistin use as a feed additive for animals in China: A prospective cross-sectional and whole genome sequencing-based molecular epidemiological study. Lancet Microbe 2020, 1, e34–e43. [Google Scholar] [CrossRef]
- Giani, T.; Sennati, S.; Antonelli, A.; Di Pilato, V.; Di Maggio, T.; Mantella, A.; Niccolai, C.; Spinicci, M.; Monasterio, J.; Castellanos, P.; et al. High prevalence of carriage of mcr-1-positive enteric bacteria among healthy children from rural communities in the Chaco region, Bolivia, september to october 2016. Eurosurveillance 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Trung, N.V.; Matamoros, S.; Carrique-Mas, J.J.; Nghia, N.H.; Nhung, N.T.; Chieu, T.T.B.; Mai, H.H.; Van Rooijen, W.; Campbell, J.; Wagenaar, J.A.; et al. Zoonotic transmission of mcr-1 colistin resistance gene from small-scale poultry farms, Vietnam. Emerg. Infect. Dis. 2017, 23, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kawahara, R.; Fujiya, Y.; Sasaki, T.; Hirai, I.; Khong, D.T.; Nguyen, T.N.; Nguyen, B.X. Wide dissemination of colistin-resistant Escherichia coli with the mobile resistance gene mcr in healthy residents in Vietnam. J. Antimicrob. Chemother. 2019, 74, 523–524. [Google Scholar] [CrossRef]
- Germ, J.; Cerar Kišek, T.; Kokošar Ulčar, B.; Lejko Zupanc, T.; Mrvič, T.; Kerin Povšič, M.; Seme, K.; Pirs, M. Surveillance cultures for detection of rectal and lower respiratory tract carriage of colistin-resistant Gram-negative bacilli in intensive care unit patients: Comparison of direct plating and pre-enrichment step. J. Med. Microbiol. 2019, 68, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Saly, M.; Jayol, A.; Poirel, L.; Megraud, F.; Nordmann, P.; Dubois, V. Prevalence of faecal carriage of colistin-resistant gram-negative rods in a university hospital in Western France, 2016. J. Med. Microbiol. 2017, 66, 842–843. [Google Scholar] [CrossRef]
- Fernández-Verdugo, A.; Forcelledo, L.; Rodríguez-Lozano, J.; Rodríguez-Lucas, C.; Barreiro-Hurlé, L.; Canut, A.; de la Iglesia, P.; Escudero, D.; Calvo, J.; Boga, J.A.; et al. Prospective multicentre study of rectal carriage of multidrug-resistant Enterobacteriaceae among health-care workers in Spain. Clin. Microbiol. Infect. 2020, 26, 649.e1–649.e4. [Google Scholar] [CrossRef]
- Bourrel, A.S.; Poirel, L.; Royer, G.; Darty, M.; Vuillemin, X.; Kieffer, N.; Clermont, O.; Denamur, E.; Nordmann, P.; Decousser, J.W. Colistin resistance in Parisian inpatient faecal Escherichia coli as the result of two distinct evolutionary pathways. J. Antimicrob. Chemother. 2019, 74, 1521–1530. [Google Scholar] [CrossRef]
- Terveer, E.M.; Nijhuis, R.H.T.; Crobach, M.J.T.; Knetsch, C.W.; Veldkamp, K.E.; Gooskens, J.; Kuijper, E.J.; Claas, E.C.J. Prevalence of colistin resistance gene (mcr-1) containing Enterobacteriaceae in feces of patients attending a tertiary care hospital and detection of a mcr-1 containing, colistin susceptible E. coli. PLoS ONE 2017, 12, e0178598. [Google Scholar] [CrossRef]
- von Wintersdorff, C.J.H.; Wolffs, P.F.G.; van Niekerk, J.M.; Beuken, E.; van Alphen, L.B.; Stobberingh, E.E.; Lashof, A.M.L.O.; Hoebe, C.J.P.A.; Savelkoul, P.H.M.; Penders, J. Detection of the plasmid-mediated colistin-resistance gene mcr-1 in faecal metagenomes of Dutch travellers. J. Antimicrob. Chemother. 2016, 71, 3416–3419. [Google Scholar] [CrossRef]
- Schaumburg, F.; Sertic, S.M.; Correa-Martinez, C.; Mellmann, A.; Köck, R.; Becker, K. Acquisition and colonization dynamics of antimicrobial-resistant bacteria during international travel: A prospective cohort study. Clin. Microbiol. Infect. 2019, 25, 1287.e1–1287.e7. [Google Scholar] [CrossRef] [PubMed]
- Gelbicova, T.; Kolackova, I.; Krutova, M.; Karpiskova, R. The emergence of mcr-1-mediated colistin-resistant Escherichia coli and Klebsiella pneumoniae in domestic and imported turkey meat in the Czech Republic 2017–2018. Folia Microbiol. 2020, 65, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Jayol, A.; Poirel, L. A universal culture medium for screening polymyxin-resistant gram-negative isolates. J. Clin. Microbiol. 2016, 54, 1395–1399. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Beyrouthy, R.; Robin, F.; Lessene, A.; Lacombat, I.; Dortet, L.; Naas, T.; Ponties, V.; Bonnet, R. MCR-1 and OXA-48 in vivo acquisition in KPC-producing Escherichia coli after colistin treatment. Antimicrob. Agents Chemother. 2017, 61, 8–11. [Google Scholar] [CrossRef]
- Poirel, L.; Kieffer, N.; Brink, A.; Coetze, J.; Jayol, A.; Nordmann, P. Genetic features of MCR-1-producing colistin-resistant Escherichia coli isolates in South Africa. Antimicrob. Agents Chemother. 2016, 60, 4394–4397. [Google Scholar] [CrossRef]
- Gelbicova, T.; Barakova, A.; Florianova, M.; Jamborova, I.; Zelendova, M.; Pospisilova, L.; Kolackova, I.; Karpiskova, R. Dissemination and comparison of genetic determinants of mcr-mediated colistin resistance in Enterobacteriaceae via retailed raw meat products. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Zhang, R.; Chen, Y.; Shen, Y.; Hu, F.; Liu, D.; Lu, J.; Guo, Y.; Xia, X.; et al. Changes in colistin resistance and mcr-1 abundance in Escherichia coli of animal and human origins following the ban of colistin-positive additives in China: An epidemiological comparative study. Lancet Infect. Dis. 2020, 20, 1161–1171. [Google Scholar] [CrossRef]
- Zurfluh, K.; Stephan, R.; Widmer, A.; Poirel, L.; Nordmann, P.; Nüesch, H.J.; Hächler, H.; Nüesch-Inderbinen, M. Screening for fecal carriage of MCR-producing Enterobacteriaceae in healthy humans and primary care patients. Antimicrob. Resist. Infect. Control. 2017, 6, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Kusumoto, M.; Nishiya, Y.; Kawamura, Y. Reactivation of insertionally inactivated Shiga toxin 2 genes of Escherichia coli O157:H7 caused by nonreplicative transposition of the insertion sequence. Appl. Environ. Microbiol. 2000, 66, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, N.; Royer, G.; Decousser, J.W.; Bourrel, A.S.; Palmieri, M.; De La Rosa, J.M.O.; Jacquier, H.; Denamur, E.; Nordmann, P.; Poirel, L. mcr-9, an inducible gene encoding an acquired phosphoethanolamine transferase in Escherichia coli, and its origin. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [PubMed]
- Pham Thanh, D.; Thanh Tuyen, H.; Nguyen Thi Nguyen, T.; Chung The, H.; Wick, R.R.; Thwaites, G.E.; Baker, S.; Holt, K.E. Inducible colistin resistance via a disrupted plasmid-borne mcr-1 gene in a 2008 Vietnamese Shigella sonnei isolate. J. Antimicrob. Chemother. 2016, 71, 2314–2317. [Google Scholar] [CrossRef]
- Zhou, K.; Luo, Q.; Wang, Q.; Huang, C.; Lu, H.; Rossen, J.W.A.; Xiao, Y.; Li, L. Silent transmission of an IS1294b-deactivated mcr-1 gene with inducible colistin resistance. Int. J. Antimicrob. Agents 2018, 51, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Liassine, N.; Assouvie, L.; Descombes, M.C.; Tendon, V.D.; Kieffer, N.; Poirel, L.; Nordmann, P. Very low prevalence of MCR-1/MCR-2 plasmid-mediated colistin resistance in urinary tract Enterobacteriaceae in Switzerland. Int. J. Infect. Dis. 2016, 51, 4–5. [Google Scholar] [CrossRef]
- Pomorska, K.; Jakubu, V.; Zelendova, M.; Dolejska, M.; Zemlickova, H. Detection of plasmid-determined colistin resistance mediated by mcr genes in the Czech Republic. Zpravy Cent. Epidemiol. A Mikrobiol. 2018, 27, 219–222. [Google Scholar]
- Krutova, M.; Kalova, A.; Nycova, E.; Gelbicova, T.; Karpiskova, R.; Smelikova, E.; Nyc, O.; Drevinek, P.; Tkadlec, J. The colonisation of Czech travellers and expatriates living in the Czech Republic by colistin-resistant Enterobacteriaceae and whole genome characterisation of E. coli isolates harbouing the mcr-1 genes on plasmid or chromosome: A cross-sectional study. Travel Med. Infect. Dis. 2021, 39, 101914. [Google Scholar] [CrossRef] [PubMed]
- Xavier, B.B.; Lammens, C.; Ruhal, R.; Kumar-Singh, S.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Identification of a novel plasmid-mediated colistin- resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Eurosurveillance 2016, 21, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Li, H.; Shen, Y.; Liu, Z.; Wang, S.; Shen, Z.; Zhang, R.; Walsh, T.R.; Shen, J.; Wang, Y. Novel plasmid-mediated colistin resistance gene mcr-3 in Escherichia coli. MBio 2017, 8, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Eurosurveillance 2017, 22, 30589. [Google Scholar] [CrossRef] [PubMed]
- Borowiak, M.; Fischer, J.; Hammerl, J.A.; Hendriksen, R.S.; Szabo, I.; Malorny, B. Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5, conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. enterica serovar Paratyphi B. J. Antimicrob. Chemother. 2017, 72, 3317–3324. [Google Scholar] [CrossRef] [PubMed]
- AbuOun, M.; Stubberfield, E.J.; Duggett, N.A.; Kirchner, M.; Dormer, L.; Nunez-Garcia, J.; Randall, L.P.; Lemma, F.; Crook, D.W.; Teale, C.; et al. mcr-1 and mcr-2 variant genes identified in Moraxella species isolated from pigs in Great Britain from 2014 to 2015. J. Antimicrob. Chemother. 2017, 72, 2745–2749. [Google Scholar] [CrossRef]
- Yang, Y.Q.; Li, Y.X.; Lei, C.W.; Zhang, A.Y.; Wang, H.N. Novel plasmid-mediated colistin resistance gene mcr-7.1 in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2018, 73, 1791–1795. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Zhou, Y.; Li, J.; Yin, W.; Wang, S.; Zhang, S.; Shen, J.; Shen, Z.; Wang, Y. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg. Microbes Infect. 2018, 7, 122. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. Testing Breakpoint Tables for Interpretation of MICs and Zone Diameters. 2020. Available online: https://eucast.org/clinical_breakpoints/ (accessed on 10 November 2020).
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Joensen, K.G.; Tetzschner, A.M.M.; Iguchi, A.; Aarestrup, F.M.; Scheutz, F. Rapid and easy in silico serotyping of Escherichia coli isolates by use of whole-genome sequencing data. J. Clin. Microbiol. 2015, 53, 2410–2426. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Beghain, J.; Bridier-Nahmias, A.; Le Nagard, H.; Denamur, E.; Clermont, O. ClermonTyping: An easy-to-use and accurate in silico method for Escherichia genus strain phylotyping. Microb. Genom. 2018, 4, e000192. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; Informational Supplement CLSI Document M100-S27; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Feng, Y.; Zou, S.; Chen, H.; Yu, Y.; Ruan, Z. BacWGSTdb 2.0: A one-stop repository for bacterial whole-genome sequence typing and source tracking. Nucleic Acids Res. 2021, 49, D644–D650. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Resistant > 2 mg/L (EUCAST) | ||||
|---|---|---|---|---|---|
| ≤2 mg/L E | 4 mg/L | 8 mg/L | ≥16 mg/L | Total Resistant | |
| Escherichia spp. (n = 84) A | 66 | 4 | 4 | 10 | 18 |
| Klebsiella spp. (n = 132) B | 53 | 7 | 11 | 61 | 79 |
| Enterobacter spp. (n = 47) C | 17 | − | 1 | 29 | 30 |
| Citrobacter spp. (n = 22) D | 20 | − | 2 | − | 2 |
| Salmonella spp. (n = 9) | 7 | − | 2 | − | 2 |
| Total (n = 294) | 163 | 11 | 20 | 100 | 131 |
| Isolate No. | COL (≥4) | AMP (≥16) | AMS (≥16/4) | CFZ (≥8) | CXM (≥16) | CPZ (≥64) | PIP (≥32) | TET (≥16) | TGC (≥1) | T/S (≥8/152) | CIP (≥1) | TOB (≥4) | CMP (≥16) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P642A | 4 | >128 | 16/8 | 16 | 16 | 8 | >128 | >32 | 1 | >4/76 | 0,5 | 1 | 8 |
| P732A | 8 | >128 | 128/64 | 16 | 8 | >64 | >128 | >32 | 0.25 | >4/76 | >8 | 4 | 4 |
| P1301A | 0.25 | 2 | 2/1 | 2 | 4 | <0.5 | <1 | >32 | 0.25 | 0.25/4.75 | 8 | 1 | >32 |
| P1519A | 8 | >128 | 64/32 | 8 | 4 | 2 | >128 | >32 | 0.5 | >4/76 | >8 | 1 | 8 |
| Isolate/Sample | Isolation (mm/yyyy) | Dept. | ST Achtman/Pasteur | cgMLST | Serotype | Phylogroup (Clermont Typing) |
|---|---|---|---|---|---|---|
| P224 * | 8/2018 | Children neurology | − | − | − | − |
| P642A | 10/2018 | Pneumology | 8778/76 | 81202 | O9:H10 | A |
| P732A | 11/2018 | Anesthesiology and ICM | 69/3 | 71872 | O unknown: H18 | D |
| P1301A | 5/2019 | Children cardiology | 744/2 | 58727 | O unknown: H9 | A |
| P1519A | 7/2019 | Children surgery | 1193/53 | 31972 | O18a:H5 | B2 |
| Isolate | Contig A | Size (bp) | Plasmids (PlasmidFinder 2.0) | AMR Genes (ResFinder 3.2) | AMR Mutations (ResFinder 3.2) | Virulence Genes (VirulenceFinder 2.0) |
|---|---|---|---|---|---|---|
| P642A | 1 | 4,726,579 | none (chromosome) | mdf(A) | none | gad; ireA; iss |
| 2 | 129,386 | IncFIB(AP001918); IncFII; IncQ1 | aadA1; aph(3″)-Ib; aph(6)-Id; blaTEM-1B; sul1; sul2; tet(A); dfrA1 | none | iroN; iss | |
| 3 | 47,696 | IncX1 | blaTEM-1B; qnrS1 | none | none | |
| 4 | 33,308 | IncX4 | mcr-1.1 | none | none | |
| P732A | 1 | 5,262,918 | none (chromosome) | mdf(A) | parE p.S458A (FQ); parC p.S80I (FQ); gyrA p.S83L (FQ); gyrA p.D87N (FQ) | air; eilA; gad; iss; lpfA |
| 2 | 93,793 | not found | none | none | none | |
| 3 | 85,283 | IncFIA; IncFIB(AP001918); IncQ1 | aac(3)-IId; aph(3″)-Ib; aph(6)-Id; catA1; sul2; tet(B) | none | none | |
| 4 | 34,080 | IncX4 | mcr-1.1 | none | none | |
| 5 | 14,745 | not found | aadA5; blaTEM-1B; mph(A); sul1; dfrA17 | none | none | |
| 6 | 8431 | not found | aph(3′)-Ia | none | none | |
| P1301A | 1 | 4,778,337 | IncQ1 B (chromosome) | aph(3″)-Ib; aph(3′)-Ia; aph(6)-Id; mdf(A); catA1; sul2; tet(B) | parC p.A56T; parC p.S80I; gyrA p.S83L; gyrA p.D87N | gad |
| 2 | 89,356 | IncFIA;IncFIB(AP001918); IncFII(pCoo) | none | none | none | |
| 3 | 73,158 | IncFII(pCoo) | tet(B) | none | none | |
| 4 | 34,639 | IncX4 | mcr-1.1 | none | none | |
| P1519A | 1 | 5,082,995 | none (chromosome) | mdf(A) | gyrA p.S83L; gyrA p.D87N; parC p.S80I; parE p.L416F | gad; iha; sat; vat |
| 2 | 250,486 | IncHI2; IncHI2A | mcr-1.1; aadA1; aadA2b; aph(3′)-Ia; cmlA1; sul3; tet(A) | none | none | |
| 3 | 110,729 | Col156; IncFIA; IncFIB(AP001918); IncQ1 | aph(3″)-Ib; aph(6)-Id; blaTEM-1B; mph(A); sul2; tet(B); dfrA17 | none | senB | |
| 5 | 2113 | Col(BS512) | none | none | none |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tkadlec, J.; Kalova, A.; Brajerova, M.; Gelbicova, T.; Karpiskova, R.; Smelikova, E.; Nyc, O.; Drevinek, P.; Krutova, M. The Intestinal Carriage of Plasmid-Mediated Colistin-Resistant Enterobacteriaceae in Tertiary Care Settings. Antibiotics 2021, 10, 258. https://doi.org/10.3390/antibiotics10030258
Tkadlec J, Kalova A, Brajerova M, Gelbicova T, Karpiskova R, Smelikova E, Nyc O, Drevinek P, Krutova M. The Intestinal Carriage of Plasmid-Mediated Colistin-Resistant Enterobacteriaceae in Tertiary Care Settings. Antibiotics. 2021; 10(3):258. https://doi.org/10.3390/antibiotics10030258
Chicago/Turabian StyleTkadlec, Jan, Alzbeta Kalova, Marie Brajerova, Tereza Gelbicova, Renata Karpiskova, Eva Smelikova, Otakar Nyc, Pavel Drevinek, and Marcela Krutova. 2021. "The Intestinal Carriage of Plasmid-Mediated Colistin-Resistant Enterobacteriaceae in Tertiary Care Settings" Antibiotics 10, no. 3: 258. https://doi.org/10.3390/antibiotics10030258