Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Effects of Azolla pinnata Ethanolic Extract against Lead-Induced Hepatotoxicity in Rats

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemicals
2.3. Preparation of Plant Extract
2.4. NMR Analysis
2.5. UPLC–MS-QToF Analysis and Molecular Networking for a Screening of Secondary Metabolites
2.6. Experimental Design
2.6.1. Control Group
2.6.2. Lead Acetate Group
2.6.3. A. pinnata Ethanolic Extract (APE) Group
2.6.4. Lead Acetate and A. pinnata Ethanolic Extract Group
2.6.5. A. pinnata Ethanolic Extract then Lead Acetate Group
2.6.6. Lead Acetate then A. pinnata Ethanolic Extract Group
2.7. Recording the Initial and Final Rats’ Weights
2.8. Blood and Tissue Sampling
2.9. Biochemical Assays
2.10. Histopathological Examination
2.11. Immunohistochemical Analysis
2.12. Statistical Analysis
3. Results
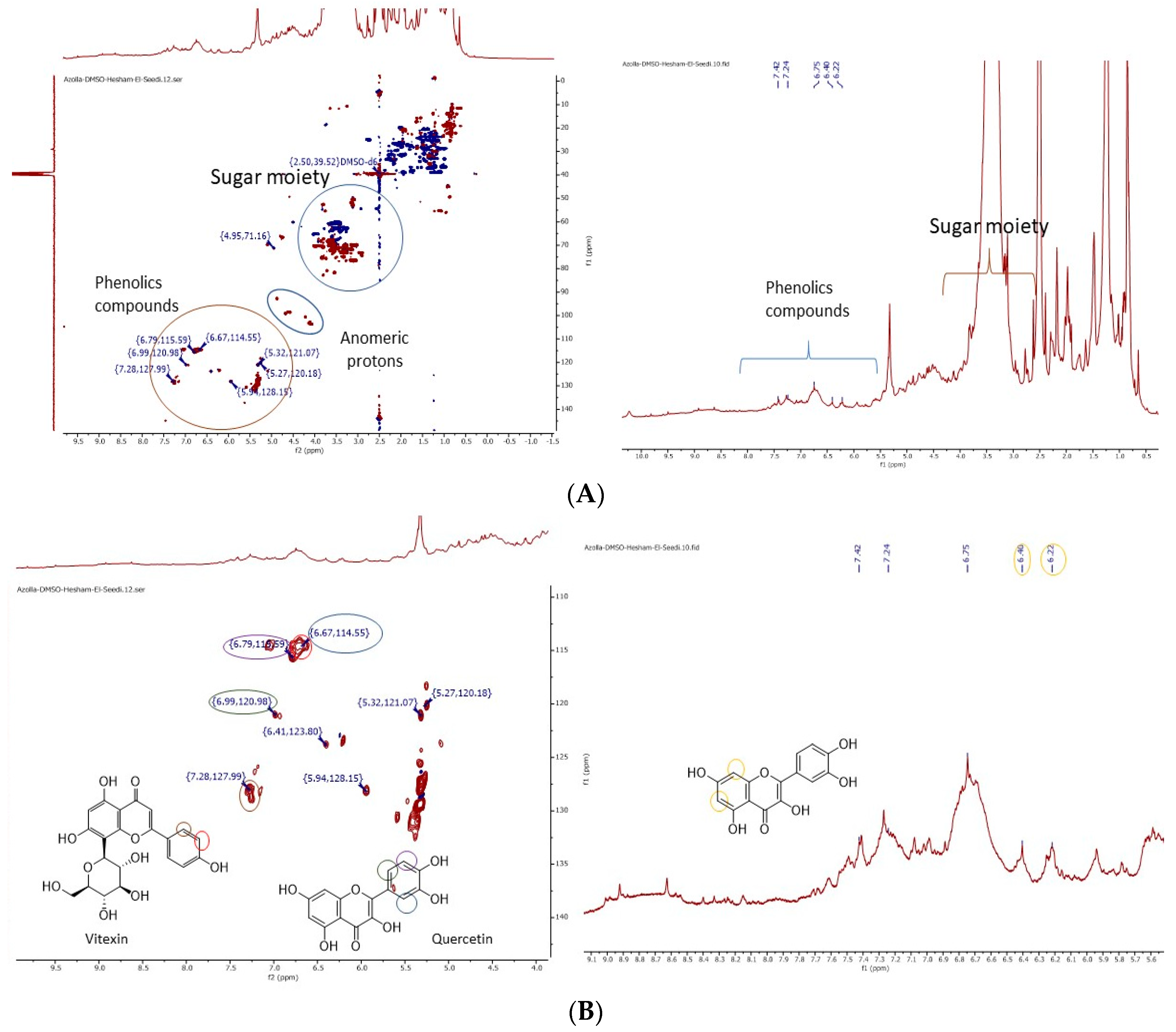
3.1. Nuclear Magnetic Resonanse (NMR) Analysis
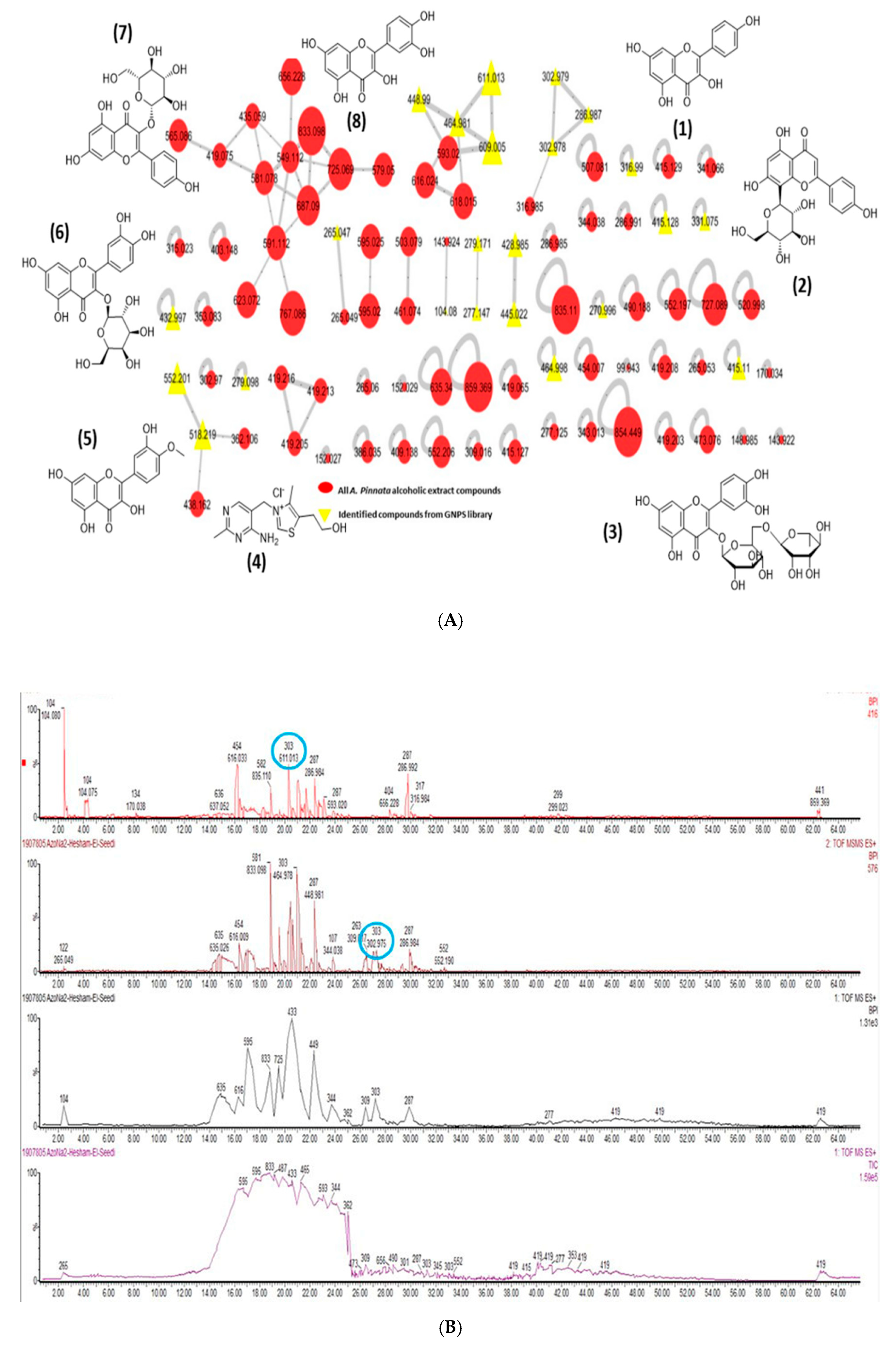
3.2. UPLC–MS-QToF and Molecular Networking Analysis
3.3. A. pinnata Extract Normalized Lead Acetate Reduced Final Body Weight of Rtas
3.4. A. pinnata Extract Modulated the Toxic Effects of Lead Acetate on Liver and Kidney Functions Biomarkers
3.5. A. pinnata Extract Reversed the Effects of Lead Acetate on Serum Levels of Inflammatory and Anti-Inflammatory Cytokines
3.6. A. pinnata Extract Ameliorated the Deleterious Effects of Lead Acetate on Oxidative/Antioxidant Status in Hepatic Tissues of Rats
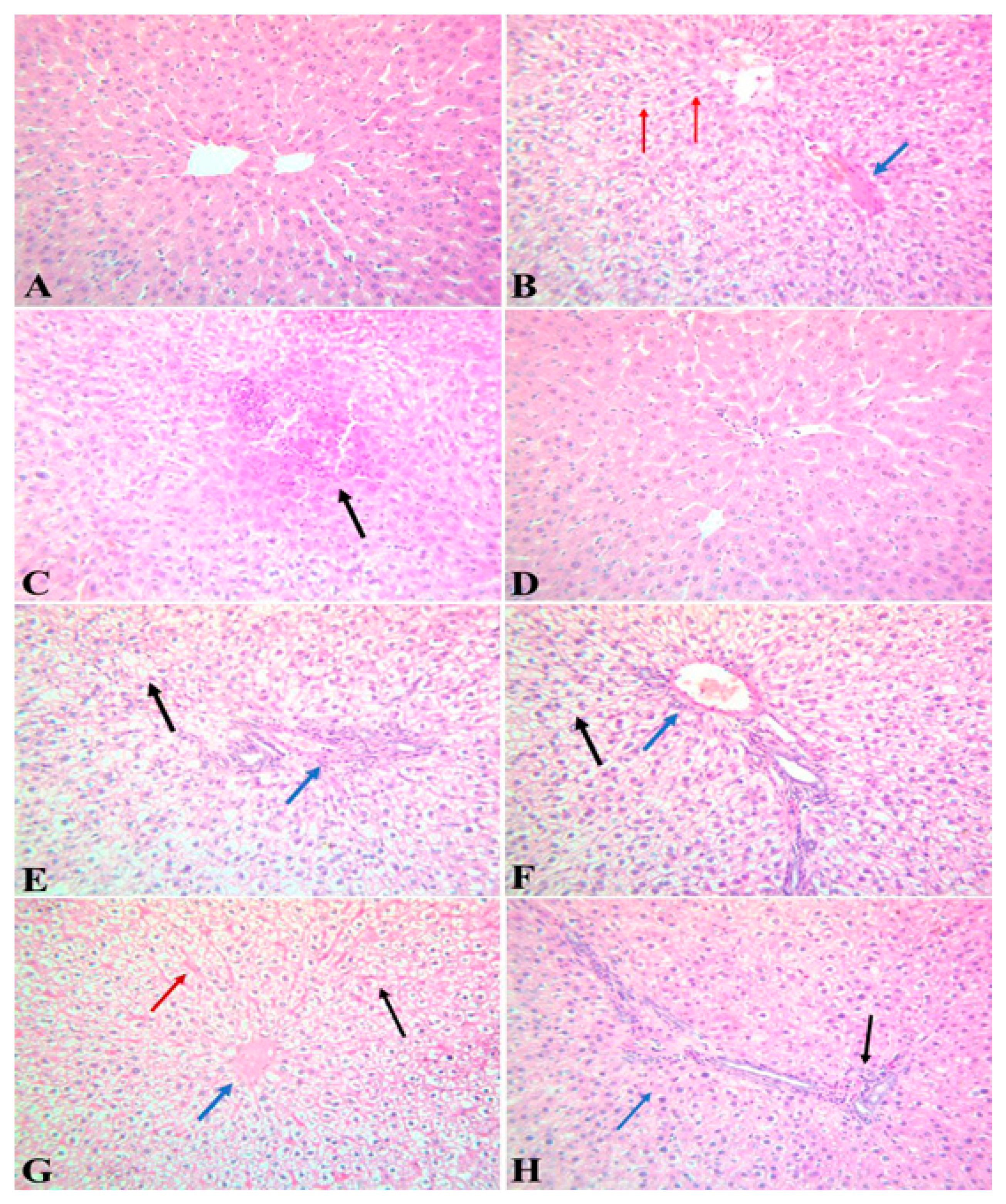
3.7. A. pinnata Extract Ameliorated the Degenerative Effect of Lead Acetate on Hepatic Tissues of Rats
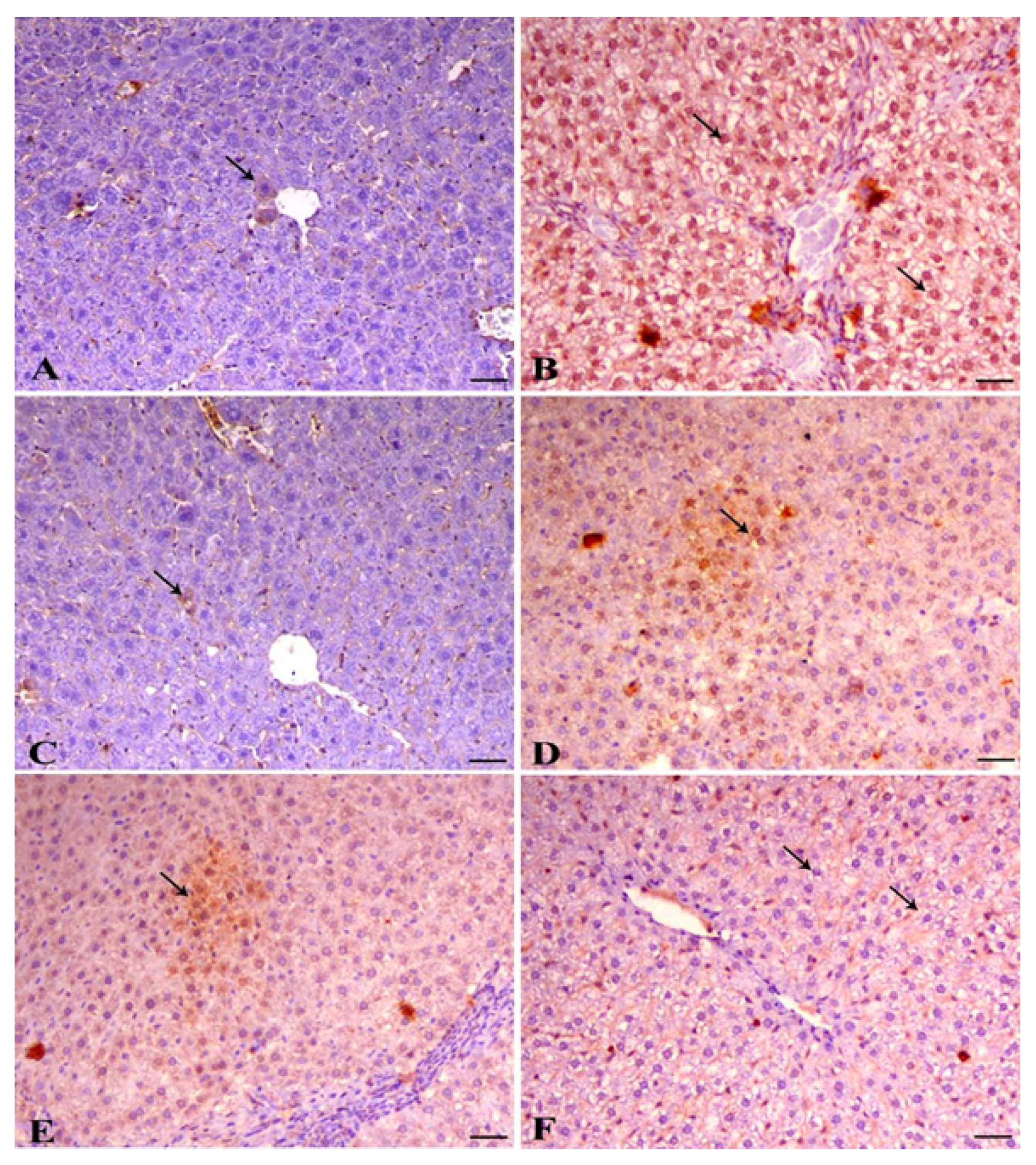
3.8. A. pinnata Extract Reduced Lead Acetate-Induced Caspase 3 Protein Expression in Hepatic Tissues of Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pal, A.; Banerjee, B.; Banerjee, T.; Masih, M.; Pal, K. Hepatoprotective activity of Chenopodium album Linn. plant against paracetamol induced hepatic injury in rats. Int. J. Pharm. Pharm. Sci. 2011, 3, 55–57. [Google Scholar]
- Gurer, H.; Ercal, N. Can antioxidants be beneficial in the treatment of lead poisoning? Free Radic. Biol. Med. 2000, 29, 927–945. [Google Scholar] [CrossRef]
- Lightfoot, T.L.; Yeager, J.M. Pet bird toxicity and related environmental concerns. Vet. Clin. N. Am. Exot. Anim. Pract. 2008, 11, 229–259. [Google Scholar] [CrossRef] [PubMed]
- Ferreyra, H.; Romano, M.; Uhart, M. Recent and chronic exposure of wild ducks to lead in human-modified wetlands in Santa Fe Province, Argentina. J. Wildl. Dis. 2009, 45, 823–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, J.R.; Moss, M.E.; Raubertas, R.F. The association between caries and childhood lead exposure. Environ. Health Perspect. 2000, 108, 1099–1102. [Google Scholar] [CrossRef]
- Flora, S.J.S.; Saxena, G.; Mehta, A. Reversal of lead-induced neuronal apoptosis by chelation treatment in rats: Role of reactive oxygen species and intracellular Ca2+. J. Pharmacol. Exp. Ther. 2007, 322, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Alhusaini, A.M.; Faddah, L.M.; Hasan, I.H.; Jarallah, S.J.; Alghamdi, S.H.; Alhadab, N.M.; Badr, A.; Elorabi, N.; Zakaria, E.; Al-Anazi, A. Vitamin C and Turmeric Attenuate Bax and Bcl-2 Proteins’ Expressions and DNA Damage in Lead Acetate-Induced Liver Injury. Dose-Response 2019, 17, 1559325819885782. [Google Scholar] [CrossRef]
- Bray, T.M.; Bettger, W.J. The physiological role of zinc as an antioxidant. Free Radic. Biol. Med. 1990, 8, 281–291. [Google Scholar] [CrossRef]
- Atawodi, S.E. Evaluation of the Hypoglycemic, Hypolipidemic and Antioxidant Effects of Methanolic Extract of” Ata-Ofa” Polyherbal Tea (APolyherbal) in Alloxan-Induced Diabetic Rats. Drug Invent. Today 2011, 3, 11. [Google Scholar]
- White, L.D.; Cory-Slechta, D.A.; Gilbert, M.E.; Tiffany-Castiglioni, E.; Zawia, N.H.; Virgolini, M.; Rossi-George, A.; Lasley, S.M.; Qian, Y.C.; Basha, M.R. New and evolving concepts in the neurotoxicology of lead. Toxicol. Appl. Pharmacol. 2007, 225, 1–27. [Google Scholar] [CrossRef]
- Billings, R.J.; Berkowitz, R.J.; Watson, G. Teeth. Pediatrics 2004, 113, 1120–1127. [Google Scholar] [PubMed]
- Lanphear, B.P.; Hornung, R.; Khoury, J.; Yolton, K.; Baghurst, P.; Bellinger, D.C.; Canfield, R.L.; Dietrich, K.N.; Bornschein, R.; Greene, T. Low-level environmental lead exposure and children’s intellectual function: An international pooled analysis. Environ. Health Perspect. 2005, 113, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Pokras, M.A.; Kneeland, M.R. Lead poisoning: Using transdisciplinary approaches to solve an ancient problem. EcoHealth 2008, 5, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Merill, J.C.; Morton, J.J.P.; Soileau, S.D. Metals. In Principles and Methods of Toxicology, 5th ed.; Hayes, A.W., Ed.; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Henretig, F.M. Lead. In Goldfrank’s Toxicoliogic Emergencies, 8th ed.; Golgfrank, L.R., Ed.; McGraw Hill Professional: New York, NY, USA, 2006. [Google Scholar]
- Patrick, L. Lead toxicity, a review of the literature. Part 1: Exposure, evaluation, and treatment. Altern. Med. Rev. 2006, 11, 2–22. [Google Scholar] [PubMed]
- El Safoury, O.S.; Abd El Fatah, D.S.; Ibrahim, M. Treatment of periocular hyperpigmentation due to lead of kohl (surma) by penicillamine: A single group non-randomized clinical trial. Indian J. Dermatol. 2009, 54, 361. [Google Scholar] [CrossRef] [PubMed]
- Shamsi-Baghbanan, H.; Sharifian, A.; Esmaeili, S.; Minaei, B. Hepatoprotective herbs, avicenna viewpoint. Iran. Red Crescent Med. J. 2014, 16, e12313. [Google Scholar] [CrossRef] [Green Version]
- Boonstra, A.; van der Laan, L.J.W.; Vanwolleghem, T.; Janssen, H.L.A. Experimental models for hepatitis C viral infection. Hepatology 2009, 50, 1646–1655. [Google Scholar] [CrossRef]
- Lal, S.S.; Nayak, P.L. Green synthesis of gold nanoparticles using various extract of plants and spices. Int. J. Sci. Innov. Discov. 2012, 2, 325–350. [Google Scholar]
- Roy, D.; Kumar, V.; Kumar, M.; Sirohi, R.; Singh, Y.; Singh, J.K. Effect of feeding Azolla pinnata on growth performance, feed intake, nutrient digestibility and blood biochemical’s of Hariana heifers fed on roughage based diet. Indian J. Dairy Sci. 2016, 69, 190–196. [Google Scholar]
- Mithraja, M.J.; Marimuthu, J.; Mahesh, M.; Paul, Z.M.; Jeeva, S. Phytochemical studies on Azolla pinnata R. Br., Marsilea minuta L. and Salvinia molesta Mitch. Asian Pac. J. Trop. Biomed. 2011, 1, S26–S29. [Google Scholar] [CrossRef]
- Kumar, G.; Chander, H. Study on the Potential of Azolla pinnata as livestock Feed Supplement for climate Change adaptation and Mitigation. Asian J. Adv. Basic Sci. 2017, 5, 65–68. [Google Scholar]
- Mousa, A.A.; El-Gansh, H.A.I.; Abd Eldaim, M.A.; Mohamed, M.A.E.-G.; Morsi, A.H.; El Sabagh, H.S. Protective effect of Moringa oleifera leaves ethanolic extract against thioacetamide-induced hepatotoxicity in rats via modulation of cellular antioxidant, apoptotic and inflammatory markers. Environ. Sci. Pollut. Res. 2019, 26, 32488–32504. [Google Scholar] [CrossRef]
- Kunjiappan, S.; Chowdhury, R.; Bhattacharjee, C. A green chemistry approach for the synthesis and characterization of bioactive gold nanoparticles using Azolla microphylla methanol extract. Front. Mater. Sci. 2014, 8, 123–135. [Google Scholar] [CrossRef]
- Gurer-Orhan, H.; Sabır, H.U.; Özgüneş, H. Correlation between clinical indicators of lead poisoning and oxidative stress parameters in controls and lead-exposed workers. Toxicology 2004, 195, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumari, J.; Kumar, H.; Nath, A.; Singh, J.K.; Ali, M.; Kumar, R. Hepatoprotective and antioxidant effect of Azolla filiculoides on profenofos induced hepatotoxicity in swiss albino mice. Carib. J. Sci. Tech. 2014, 2, 372–377. [Google Scholar]
- Fawcett, J.K.; Scott, J.E. A rapid and precise method for the determination of urea. J. Clin. Pathol. 1960, 13, 156–159. [Google Scholar] [CrossRef] [Green Version]
- Bartels, H.; Böhmer, M.; Heierli, C. Serum creatinine determination without protein precipitation. Clin. Chim. Acta Int. J. Clin. Chem. 1972, 37, 193. [Google Scholar] [CrossRef]
- Larsen, K. Creatinine assay by a reaction-kinetic principle. Clin. Chim. Acta 1972, 41, 209–217. [Google Scholar] [CrossRef]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Kei, S. Serum lipid peroxide in cerebrovascular disorders determined by a new colorimetric method. Clin. Chim. Acta 1978, 90, 37–43. [Google Scholar] [CrossRef]
- Beutler, E. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Fossati, P.; Prencipe, L.; Berti, G. Use of 3, 5-dichloro-2-hydroxybenzenesulfonic acid/4-aminophenazone chromogenic system in direct enzymic assay of uric acid in serum and urine. Clin. Chem. 1980, 26, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Vidal-Vanaclocha, F.; Fantuzzi, G.; Mendoza, L.; Fuentes, A.M.; Anasagasti, M.J.; Martín, J.; Carrascal, T.; Walsh, P.; Reznikov, L.L.; Kim, S.-H. IL-18 regulates IL-1β-dependent hepatic melanoma metastasis via vascular cell adhesion molecule-1. Proc. Natl. Acad. Sci. USA 2000, 97, 734–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brynskov, J.; Foegh, P.; Pedersen, G.; Ellervik, C.; Kirkegaard, T.; Bingham, A.; Saermark, T. Tumour necrosis factor α converting enzyme (TACE) activity in the colonic mucosa of patients with inflammatory bowel disease. Gut 2002, 51, 37–43. [Google Scholar] [CrossRef]
- Odewumi, C.; Latinwo, L.M.; Sinclair, A.; Badisa, V.L.D.; Abdullah, A.; Badisa, R.B. Effect of cadmium on the expression levels of interleukin-1α and interleukin-10 cytokines in human lung cells. Mol. Med. Rep. 2015, 12, 6422–6426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopwood, D.; Bamcroft, J.D.; Stevens, A. Theory and practice of histopathological techniques. In Fixation and Fixatives, 4th ed.; Churchill Livingstone: Hong Kong, 1996; pp. 23–46. [Google Scholar]
- Hsu, B.-G.; Lee, R.-P.; Yang, F.-L.; Harn, H.-J.; Chen, H.I. Post-treatment with N-acetylcysteine ameliorates endotoxin shock-induced organ damage in conscious rats. Life Sci. 2006, 79, 2010–2016. [Google Scholar] [CrossRef]
- Chen, T.M.; Subeq, Y.M.; Lee, R.P.; Chiou, T.W.; Hsu, B.G. Single dose intravenous thioacetamide administration as a model of acute liver damage in rats. Int. J. Exp. Pathol. 2008, 89, 223–231. [Google Scholar] [CrossRef]
- Orabi, S.H.; Al-Sabbagh, E.S.H.; Khalifa, H.K.; Mohamed, M.A.E.-G.; Elhamouly, M.; Gad-Allah, S.M.; Abdel-Daim, M.M.; Eldaim, M.A.A. Commiphora myrrha Resin Alcoholic Extract Ameliorates High Fat Diet Induced Obesity via Regulation of UCP1 and Adiponectin Proteins Expression in Rats. Nutrients 2020, 12, 803. [Google Scholar] [CrossRef] [Green Version]
- Aron, A.T.; Gentry, E.C.; McPhail, K.L.; Nothias, L.-F.; Nothias-Esposito, M.; Bouslimani, A.; Petras, D.; Gauglitz, J.M.; Sikora, N.; Vargas, F. Reproducible molecular networking of untargeted mass spectrometry data using GNPS. Nat. Protoc. 2020, 15, 1954–1991. [Google Scholar] [CrossRef]
- Amjad, Z.; Yasmin, T.; Ashraf, I.; Perveen, K.; Mirza, T.; Shoro, A.A. Lead-induced morphometric changes in the kidneys of albino rats ameliorated by ginkgo biloba extract (EGb 761). J. Pak. Med. Assoc. 2017, 67, 58–65. [Google Scholar] [PubMed]
- Rafique, M.; Perveen, K.; Khan, N.; Nigar, S. Lead intoxication causing loss of body weight and loss of absolute weight of testes in albino rats. Hamdard Med. 2008, 51, 123–128. [Google Scholar]
- Ercal, N.; Gurer-Orhan, H.; Aykin-Burns, N. Toxic metals and oxidative stress part I: Mechanisms involved in metal-induced oxidative damage. Curr. Top. Med. Chem. 2001, 1, 529–539. [Google Scholar] [CrossRef]
- Yeh, Y.C.; Liu, T.J.; Wang, L.C.; Lee, H.W.; Ting, C.T.; Lee, W.-L.; Hung, C.-J.; Wang, K.Y.; Lai, H.C. A standardized extract of Ginkgo biloba suppresses doxorubicin-induced oxidative stress and p53-mediated mitochondrial apoptosis in rat testes. Br. J. Pharmacol. 2009, 156, 48–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, R.C.; Rautray, A.K.; Swarup, D. Oxidative stress in lead and cadmium toxicity and its amelioration. Vet. Med. Int. 2011, 2011, 457327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.P.; Schwartz, R.J.; Waddell, I.D.; Holloway, B.R.; Reid, M.B. Skeletal muscle myocytes undergo protein loss and reactive oxygen-mediated NF-kappaB activation in response to tumor necrosis factor alpha. FASEB J. 1998, 12, 871–880. [Google Scholar] [PubMed]
- Singh, R.; Kumar, S.; Rana, A.C.; Sharma, N. Different models of hepatotoxicity and related liver diseases: A review. Int. Res. J. Pharm. 2012, 3, 86–95. [Google Scholar]
- Suganthi, V.; Gowri, S.; Gurusamy, K. Hepatoprotective activity of Cayratia carnosa on liver damage caused by lead acetate in rats. Sch. Res. Lib. 2013, 3, 76–79. [Google Scholar]
- Haouem, S.; Chargui, I.; Najjar, M.F.; Sriha, B.; El Hani, A. Liver Function and Structure in Rats Treated Simultaneously with Cadmium and Mercury. Open J. Pathol. 2013, 3, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Drill, V.A. Pharmacology of hepatotoxic agents. NYASA 1963, 104, 858–874. [Google Scholar] [CrossRef]
- Gao, B.; Seki, E.; Brenner, D.A.; Friedman, S.; Cohen, J.I.; Nagy, L.; Szabo, G.; Zakhari, S. Innate immunity in alcoholic liver disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G516–G525. [Google Scholar] [CrossRef] [PubMed]
- Núñez, M. Hepatotoxicity of antiretrovirals: Incidence, mechanisms and management. J. Hepatol. 2006, 44, S132–S139. [Google Scholar] [CrossRef] [PubMed]
- Malekirad, A.A.; Oryan, S.; Fani, A.; Babapor, V.; Hashemi, M.; Baeeri, M.; Bayrami, Z.; Abdollahi, M. Study on clinical and biochemical toxicity biomarkers in a zinc-lead mine workers. Toxicol. Ind. Health 2010, 26, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Sirivarasai, J.; Wananukul, W.; Kaojarern, S.; Chanprasertyothin, S.; Thongmung, N.; Ratanachaiwong, W.; Sura, T.; Sritara, P. Association between inflammatory marker, environmental lead exposure, and glutathione S-transferase gene. Biomed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Valentino, M.; Rapisarda, V.; Santarelli, L.; Bracci, M.; Scorcelletti, M.; Di Lorenzo, L.; Cassano, F.; Soleo, L. Effect of lead on the levels of some immunoregulatory cytokines in occupationally exposed workers. Hum. Exp. Toxicol. 2007, 26, 551–556. [Google Scholar] [CrossRef]
- Machoń-Grecka, A.; Dobrakowski, M.; Boroń, M.; Lisowska, G.; Kasperczyk, A.; Kasperczyk, S. The influence of occupational chronic lead exposure on the levels of selected pro-inflammatory cytokines and angiogenic factors. Hum. Exp. Toxicol. 2017, 36, 467–473. [Google Scholar] [CrossRef]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Kasten-Jolly, J.; Pabello, N.; Bolivar, V.J.; Lawrence, D.A. Developmental lead effects on behavior and brain gene expression in male and female BALB/cAnNTac mice. Neurotoxicology 2012, 33, 1005–1020. [Google Scholar] [CrossRef] [Green Version]
- Chibowska, K.; Baranowska-Bosiacka, I.; Falkowska, A.; Gutowska, I.; Goschorska, M.; Chlubek, D. Effect of lead (Pb) on inflammatory processes in the brain. Int. J. Mol. Sci. 2016, 17, 2140. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.-L.; Bongiorno, P.B.; Rettori, V.; McCann, S.M.; Licinio, J. Int.erleukin (IL) 1β, IL-1 receptor antagonist, IL-10, and IL-13 gene expression in the central nervous system and anterior pituitary during systemic inflammation: Pathophysiological implications. Proc. Natl. Acad. Sci. USA 1997, 94, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Schuchmann, M.; Varfolomeev, E.E.; Hermann, F.; Rueckert, F.; Strand, D.; Koehler, H.; Strand, S.; Lohse, A.W.; Wallach, D.; Galle, P.R. Dominant negative MORT1/FADD rescues mice from CD95 and TNF-induced liver failure. Hepatology 2003, 37, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Salvesen, G.S. Program in apoptosis and cell death research, The Burnham Institute. Chem. Rev. 2002, 102, 4489–4500. [Google Scholar]
- Alnemri, E.S.; Livingston, D.J.; Nicholson, D.W.; Salvesen, G.; Thornberry, N.A.; Wong, W.W.; Yuan, J. Human ICE/CED-3 protease nomenclature. Cell 1996, 87, 171. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.-L.; Fan, M.-X.; Wu, J.-L.; Li, N.; Guo, M.-Q. Antioxidant and anti-inflammatory properties of flavonoids from lotus plumule. Food Chem. 2019, 277, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Wu, F.; Zheng, T.; Wang, X.; Chen, Y.; Wu, X. Kaempferol attenuates retinal ganglion cell death by suppressing NLRP1/NLRP3 inflammasomes and caspase-8 via JNK and NF-κB pathways in acute glaucoma. Eye 2019, 33, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Dohaei, M.; Karimi, K.; Rahimmalek, M.; Satari, B. Integrated biorefinery of aquatic fern Azolla filiculoides for enhanced extraction of phenolics, protein, and lipid and methane production from the residues. J. Clean. Prod. 2020, 276, 123175. [Google Scholar] [CrossRef]
- Da Silva, A.B.; Coelho, P.L.C.; das Neves Oliveira, M.; Oliveira, J.L.; Amparo, J.A.O.; da Silva, K.C.; Soares, J.R.P.; Pitanga, B.P.S.; dos Santos Souza, C.; de Faria Lopes, G.P. The flavonoid rutin and its aglycone quercetin modulate the microglia inflammatory profile improving antiglioma activity. Brain Behav. Immun. 2020, 85, 170–185. [Google Scholar] [CrossRef]
- Borghi, S.M.; Carvalho, T.T.; Staurengo-Ferrari, L.; Hohmann, M.S.N.; Pinge-Filho, P.; Casagrande, R.; Verri, W.A., Jr. Vitexin inhibits inflammatory pain in mice by targeting TRPV1, oxidative stress, and cytokines. J. Nat. Prod. 2013, 76, 1141–1149. [Google Scholar] [CrossRef]
- Park, J.Y.; Han, X.; Piao, M.J.; Oh, M.C.; Fernando, P.M.D.J.; Kang, K.A.; Ryu, Y.S.; Jung, U.; Kim, I.G.; Hyun, J.W. Hyperoside induces endogenous antioxidant system to alleviate oxidative stress. J. Cancer Prev. 2016, 21, 41. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, K.-H.; Yoo, D.H.; Kang, D.; Cho, S.-H.; Hong, Y.-C. GSTM1 and TNF-α gene polymorphisms and relations between blood lead and inflammatory markers in a non-occupational population. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2007, 629, 32–39. [Google Scholar] [CrossRef]
- Mkhize, N.V.P.; Qulu, L.; Mabandla, M.V. The effect of quercetin on pro-and anti-inflammatory cytokines in a prenatally stressed rat model of febrile seizures. J. Exp. Neurosci. 2017, 11, 1179069517704668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, G.L.; Russo, M.; Spagnuolo, C.; Tedesco, I.; Bilotto, S.; Iannitti, R.; Palumbo, R. Quercetin: A pleiotropic kinase inhibitor against cancer. In Advances in Nutrition and Cancer; Springer: Berlin/Heidelberg, Germany, 2014; pp. 185–205. [Google Scholar]
- Murakami, A.; Ashida, H.; Terao, J. Multitargeted cancer prevention by quercetin. Cancer Lett. 2008, 269, 315–325. [Google Scholar] [CrossRef]
- Portari, G.V.; Marchini, J.S.; Vannucchi, H.; Jordao, A.A. Antioxidant effect of thiamine on acutely alcoholized rats and lack of efficacy using thiamine or glucose to reduce blood alcohol content. Basic Clin. Pharmacol. Toxicol. 2008, 103, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-Q.; Xu, T.; Bu, X.-M.; Liu, B.-Y. Anti-inflammation effects of corn silk in a rat model of carrageenin-induced pleurisy. Inflammation 2012, 35, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Aihara, M.; Chen, Y.-N.; Araie, M.; Tomita-Yokotani, K.; Iwashina, T. Neuroprotective effects of flavonoids on hypoxia-, glutamate-, and oxidative stress–induced retinal ganglion cell death. Mol. Vis. 2011, 17, 1784. [Google Scholar]
- Janbaz, K.H.; Saeed, S.A.; Gilani, A.H. Protective effect of rutin on paracetamol-and CCl4-induced hepatotoxicity in rodents. Fitoterapia 2002, 73, 557–563. [Google Scholar] [CrossRef]
- Abbasi, E.; Nassiri-Asl, M.; Sheikhi, M.; Shafiee, M. Effects of vitexin on scopolamine-induced memory impairment in rats. Chin. J. Physiol. 2013, 56, 184–1889. [Google Scholar]
- Bai, Y.; Chang, J.; Xu, Y.; Cheng, D.; Liu, H.; Zhao, Y.; Yu, Z. Antioxidant and myocardial preservation activities of natural phytochemicals from mung bean (Vigna radiata L.) seeds. J. Agric. Food Chem. 2016, 64, 4648–4655. [Google Scholar] [CrossRef]
- Choi, J.S.; Islam, M.N.; Ali, M.Y.; Kim, E.J.; Kim, Y.M.; Jung, H.A. Effects of C-glycosylation on anti-diabetic, anti-Alzheimer’s disease and anti-inflammatory potential of apigenin. Food Chem. Toxicol. 2014, 64, 27–33. [Google Scholar] [CrossRef]
- Nikfarjam, B.A.; Hajiali, F.; Adineh, M.; Nassiri-Asl, M. Anti-inflammatory effects of quercetin and vitexin on activated human peripheral blood neutrophils-The effects of quercetin and vitexin on human neutrophils. J. Pharmacopunct. 2017, 20, 127–131. [Google Scholar]
- Praveena, R.; Sadasivam, K.; Kumaresan, R.; Deepha, V.; Sivakumar, R. Experimental and DFT studies on the antioxidant activity of a C-glycoside from Rhynchosia capitata. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2013, 103, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liang, D.; Yang, Z.; Wang, T.; Wang, W.; Song, X.; Guo, M.; Zhou, E.; Li, D.; Cao, Y. Astragalin suppresses inflammatory responses via down-regulation of NF-κB signaling pathway in lipopolysaccharide-induced mastitis in a murine model. Int. Immunopharmacol. 2013, 17, 478–482. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | M.F | M.W(Da) |
|---|---|---|
| Kaempferol (1) | [C15H10O6] + | 286.987 |
| Vitexin (2) | [C21H20O10] + | 432.997 |
| Rutin (3) | [C27H30O16] + | 611.013 |
| Thiamine (4) | [C12H17N4OS] + | 265.047 |
| Tamarixetin (5) | [C16H12O7] + | 316.99 |
| Hyperoside (6) | [C21H20O12] + | 464.981 |
| Astragalin (7) | [C21H20O11] + | 448.99 |
| Quercetin (8) | [C15H10O7] + | 302.979 |
| Parameters | Control | Lead | APE | Lead and APE | APE Then Lead | Lead Then APE |
|---|---|---|---|---|---|---|
| Initial BW (g) | 121.20 ± 1.28 | 120.20 ± 1.14 | 120.30 ± 1.03 | 120.00 ± 0.85 | 120.30 ± 1.03 | 120.40 ± 0.79 |
| Final BW (g) | 166.90 ± 2.91 a | 101.29 ± 3.39 b | 161.30 ± 2.54 a | 150.33 ± 0.88 a | 154.56 ± 4.19 a | 151.50 ± 3.33 a |
| Parameters | Control | Lead | APE | Lead and APE | APE Then Lead | Lead Then APE |
|---|---|---|---|---|---|---|
| ALT (U/L) | 18.00 ± 3.42 b | 52.00 ± 0.85 a | 19.00 ± 2.09 b | 22.75 ± 3.21 b | 45. 80 ± 0.88 a | 23.75 ± 5.61 b |
| AST (U/L) | 110.40 ± 6.35 c | 157.20 ± 8.5 a | 110.00 ± 7.78 c | 135.00 ± 4.44 b | 132.40 ± 3.02 b | 137.00 ± 4.69 b |
| Creatinine (mg/dL) | 0.72 ± 0.02 b | 1.03 ± 0.05 a | 0.62 ± 0.02 b | 0.85 ± 0.03 b | 0.73 ± 0.04 b | 0.74 ± 0.02 b |
| Urea (mg /dL) | 19.41 ± 1.08 d | 38.03 ± 0.65 a | 21.60 ± 0.66 cd | 23.85 ± 1.29 c | 27.73 ± 0.42 b | 29.72 ± 1.78 b |
| Parameters | Control | Lead | APE | Lead and APE | APE Then Lead | Lead Then APE |
|---|---|---|---|---|---|---|
| TNF-α (pg/mL) | 85.80 ± 2.10 c | 117.75 ± 2.78 a | 86.60 ± 2.56 c | 97.50 ± 2.96 b | 95.0 ± 2.59 b | 104.20 ± 2.54 b |
| IL-1β (pg/mL) | 177.20 ± 3.34 b | 284.60 ± 3.30 a | 169.20 ± 1.93 b | 194.75 ± 3.07 b | 193.60 ± 5.27 b | 199.40 ± 4.23 b |
| IL-10 (pg/mL) | 5.82 ± 0.32 b | 1.73 ± 0.23 d | 8.92 ± 0.46 a | 5.70 ± 0.38 b | 3.73 ± 0.71 c | 3.725 ± 0.55 c |
| Parameters | Control | Lead | APE | Lead and APE | APE Then Lead | Lead Then APE |
|---|---|---|---|---|---|---|
| MDA (nmol/g tissue) | 8.85 ± 0.38 c | 17.64 ± 0.27 a | 10.32 ± 0.44 bc | 12.42 ± 0.73 b | 12.34 ± 0.16 b | 10.95 ± 0.63 b |
| GSH (mmol/g tissue) | 2.84 ± 0.14 a | 0.49 ± 0.0 c | 2.69 ± 0.11 a | 1.22 ± 0.09 b | 2.63 ± 0.21 a | 1.31 ± 0.30 b |
| CAT (U/ g tissue) | 0.66 ± 0.01 a | 0.22 ± 0.0 c | 0.63 ± 0.01 a | 0.35 ± 0.02 b | 0.35 ± 0.02 b | 0.33 ± 0.05 b |
| SOD (U/g tissue) | 3.43 ± 0.08 a | 1.67 ± 0.06 c | 3.57 ± 0.06 a | 2.84 ± 0.09 b | 3.16 ± 0.18 a | 2. 82 ± 0.16 b |
| Lesions | Control | Lead | APE | Lead and APE | APE Then Lead | Lead Then APE |
|---|---|---|---|---|---|---|
| Vascular and inflammatory changes | - | +++ | - | + | + | ++ |
| Degenerative and necrotic changes | - | +++ | - | + | + | ++ |
| Proliferative changes | - | ++ | - | - | - |
| Control | Lead | APE | Lead and APE | APE then Lea | Lead then APE | |
|---|---|---|---|---|---|---|
| Caspase 3 expression | 10.25 ± 0.67 e | 70.5 ± 3.73 a | 9.20 ± 0.61 e | 27.03 ± 1.97 c | 18.37 ± 1.01 d | 38.83 ± 1.22 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elrasoul, A.S.A.; Mousa, A.A.; Orabi, S.H.; Mohamed, M.A.E.-G.; Gad-Allah, S.M.; Almeer, R.; Abdel-Daim, M.M.; Khalifa, S.A.M.; El-Seedi, H.R.; Eldaim, M.A.A. Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Effects of Azolla pinnata Ethanolic Extract against Lead-Induced Hepatotoxicity in Rats. Antioxidants 2020, 9, 1014. https://doi.org/10.3390/antiox9101014
Elrasoul ASA, Mousa AA, Orabi SH, Mohamed MAE-G, Gad-Allah SM, Almeer R, Abdel-Daim MM, Khalifa SAM, El-Seedi HR, Eldaim MAA. Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Effects of Azolla pinnata Ethanolic Extract against Lead-Induced Hepatotoxicity in Rats. Antioxidants. 2020; 9(10):1014. https://doi.org/10.3390/antiox9101014
Chicago/Turabian StyleElrasoul, Ahmed Shaaban Abd, Ahmed Abdelmoniem Mousa, Sahar Hassan Orabi, Mostafa Abd El-Gaber Mohamed, Shaban M. Gad-Allah, Rafa Almeer, Mohamed M. Abdel-Daim, Shaden A. M. Khalifa, Hesham R. El-Seedi, and Mabrouk Attia Abd Eldaim. 2020. "Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Effects of Azolla pinnata Ethanolic Extract against Lead-Induced Hepatotoxicity in Rats" Antioxidants 9, no. 10: 1014. https://doi.org/10.3390/antiox9101014