Period of Boar Ejaculate Collection Contributes to the Yearly Intra-Male Variability of Seminal Plasma Cytokines

 ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals, Ejaculates, and Seminal Plasma Collection
2.2. Cytokine Measurement
2.3. Experimental Design
2.4. Statistical Analysis
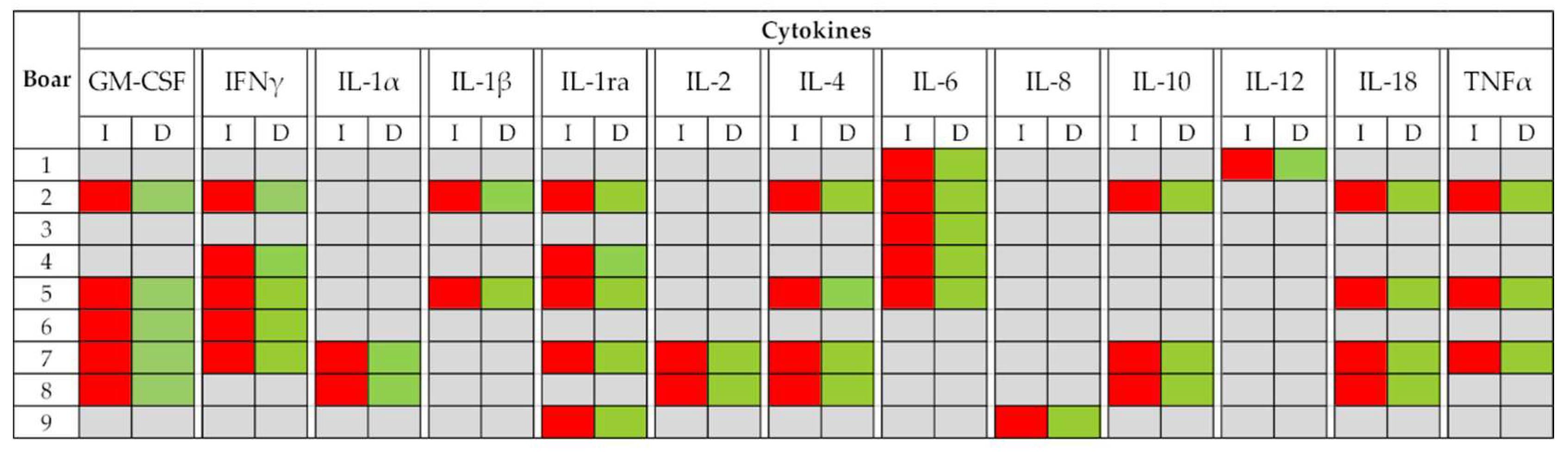
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Caballero, I.; Parrilla, I.; Almiñana, C.; del Olmo, D.; Roca, J.; Martínez, E.A.; Vázquez, J.M. Seminal Plasma Proteins as Modulators of the Sperm Function and Their Application in Sperm Biotechnologies. Reprod. Domest. Anim. 2012, 47, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martínez, H.; Kvist, U.; Saravia, F.; Wallgren, M.; Johannisson, A.; Sanz, L.; Peña, F.J.; Martínez, E.A.; Roca, J.; Vázquez, J.M.; et al. The physiological roles of the boar ejaculate. Soc. Reprod. Fertil. Suppl. 2009, 66, 1–21. [Google Scholar] [PubMed]
- Kumar, A.; Sridharn, T.B.; Rao, K.A. Role of Seminal Plasma Proteins in Effective Zygote Formation- A Success Road to Pregnancy. Protein Pept. Lett. 2019, 26, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Juyena, N.S.; Stelletta, C. Seminal plasma: An essential attribute to spermatozoa. J. Androl. 2012, 33, 536–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, S.A.; Sharkey, D.J. Seminal fluid and fertility in women. Fertil. Steril. 2016, 106, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Bromfield, J.J. Seminal fluid and reproduction: Much more than previously thought. J. Assist. Reprod. Genet. 2014, 31, 627–636. [Google Scholar] [CrossRef] [Green Version]
- Szczykutowicz, J.; Kałuża, A.; Kaźmierowska-Niemczuk, M.; Ferens-Sieczkowska, M. The Potential Role of Seminal Plasma in the Fertilization Outcomes. Biomed Res. Int. 2019, 2019, 5397804. [Google Scholar] [CrossRef]
- Druart, X.; Rickard, J.P.; Tsikis, G.; de Graaf, S.P. Seminal plasma proteins as markers of sperm fertility. Theriogenology 2019, 137, 30–35. [Google Scholar] [CrossRef]
- Pérez-Patiño, C.; Parrilla, I.; Barranco, I.; Vergara-Barberán, M.; Simó-Alfonso, E.F.; Herrero-Martínez, J.M.; Rodriguez-Martínez, H.; Martínez, E.A.; Roca, J. New In-Depth Analytical Approach of the Porcine Seminal Plasma Proteome Reveals Potential Fertility Biomarkers. J. Proteome Res. 2018, 17, 1065–1076. [Google Scholar] [CrossRef] [Green Version]
- Samanta, L.; Parida, R.; Dias, T.R.; Agarwal, A. The enigmatic seminal plasma: A proteomics insight from ejaculation to fertilization. Reprod. Biol. Endocrinol. 2018, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Schjenken, J.E.; Robertson, S.A. Seminal fluid and immune adaptation for pregnancy - comparative biology in mammalian species. Reprod. Domest. Anim. 2014, 49, 27–36. [Google Scholar] [CrossRef]
- Chyra-Jach, D.; Kaletka, Z.; Dobrakowski, M.; Machoń-Grecka, A.; Kasperczyk, S.; Birkner, E.; Kasperczyk, A. The Associations between Infertility and Antioxidants, Proinflammatory Cytokines, and Chemokines. Oxid. Med. Cell. Longev. 2018, 2018, 8354747. [Google Scholar] [CrossRef]
- Fraczek, M.; Kurpisz, M. Cytokines in the male reproductive tract and their role in infertility disorders. J. Reprod. Immunol. 2015, 108, 98–104. [Google Scholar] [CrossRef]
- Havrylyuk, A.; Chopyak, V.; Boyko, Y.; Kril, I.; Kurpisz, M. Cytokines in the blood and semen of infertile patients. Cent. J. Immunol. 2015, 40, 337–344. [Google Scholar] [CrossRef]
- Chyra-Jach, D.; Kaletka, Z.; Dobrakowski, M.; Machoń-Grecka, A.; Kasperczyk, S.; Bellanti, F.; Birkner, E.; Kasperczyk, A. Levels of Macro- and Trace Elements and Select Cytokines in the Semen of Infertile Men. Biol. Trace Elem. Res. 2020. [Google Scholar] [CrossRef]
- Ghandehari-Alavijeh, R.; Zohrabi, D.; Tavalaee, M.; Nasr-Esfahani, M.H. Association between expression of TNF-α, P53 and HIF1α with asthenozoospermia. Hum. Fertil. (Camb) 2019, 22, 145–151. [Google Scholar] [CrossRef]
- Grande, G.; Milardi, D.; Baroni, S.; Luca, G.; Pontecorvi, A. Identification of seminal markers of male accessory gland inflammation: From molecules to proteome. Am. J. Reprod. Immunol. 2018, 80, e12992. [Google Scholar] [CrossRef]
- Sharkey, D.J.; Tremellen, K.P.; Briggs, N.E.; Dekker, G.A.; Robertson, S.A. Seminal plasma pro-inflammatory cytokines interferon-γ (IFNG) and C-X-C motif chemokine ligand 8 (CXCL8) fluctuate over time within men. Hum. Reprod. 2017, 32, 1373–1381. [Google Scholar] [CrossRef]
- Barranco, I.; Rubér, M.; Perez-Patiño, C.; Atikuzzaman, M.; Martinez, E.A.; Roca, J.; Rodriguez-Martinez, H. The Seminal Plasma of the Boar is Rich in Cytokines, with Significant Individual and Intra-Ejaculate Variation. Am. J. Reprod. Immunol. 2015, 74, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Schirmer, M.; Kumar, V.; Netea, M.G.; Xavier, R.J. The causes and consequences of variation in human cytokine production in health. Curr. Opin. Immunol. 2018, 54, 50–58. [Google Scholar] [CrossRef]
- Ter Horst, R.; Jaeger, M.; Smeekens, S.P.; Oosting, M.; Swertz, M.A.; Li, Y.; Kumar, V.; Diavatopoulos, D.A.; Jansen, A.F.M.; Lemmers, H.; et al. Host and Environmental Factors Influencing Individual Human Cytokine Responses. Cell 2016, 167, 1111–1124.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kastelic, J.P.; Wilde, R.E.; Bielli, A.; Genovese, P.; Rizzoto, G.; Thundathil, J. Hyperthermia is more important than hypoxia as a cause of disrupted spermatogenesis and abnormal sperm. Theriogenology 2019, 131, 177–181. [Google Scholar] [CrossRef]
- Rivera, M.M.; Quintero-Moreno, A.; Barrera, X.; Palomo, M.J.; Rigau, T.; Rodríguez-Gil, J.E. Natural Mediterranean photoperiod does not affect the main parameters of boar-semen quality analysis. Theriogenology 2005, 64, 934–946. [Google Scholar] [CrossRef] [PubMed]
- Roca, J.; Broekhuijse, M.L.W.J.; Parrilla, I.; Rodriguez-Martinez, H.; Martinez, E.A.; Bolarin, A. Boar Differences In Artificial Insemination Outcomes: Can They Be Minimized? Reprod. Domest. Anim. 2015, 50, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suriyasomboon, A.; Lundeheim, N.; Kunavongkrit, A.; Einarsson, S. Effect of temperature and humidity on sperm morphology in duroc boars under different housing systems in Thailand. J. Vet. Med. Sci. 2005, 67, 777–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrout, P.E.; Fleiss, J.L. Intraclass correlations: Uses in assessing rater reliability. Psychol. Bull. 1979, 86, 420–428. [Google Scholar] [CrossRef]
- Koo, T.K.; Li, M.Y. A Guideline of Selecting and Reporting Intraclass Correlation Coefficients for Reliability Research. J. Chiropr. Med. 2016, 15, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Petrie, A.; Watson, P. Linear Correlation and Regression. In Statistics for Veterinary and Animal Science, 3rd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 126–142. ISBN 978-0-470-67075-0. [Google Scholar]
- Eldamnhoury, E.M.; Elatrash, G.A.; Rashwan, H.M.; El-Sakka, A.I. Association between leukocytospermia and semen interleukin-6 and tumor necrosis factor-alpha in infertile men. Andrology 2018, 6, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Loveland, K.L.; Klein, B.; Pueschl, D.; Indumathy, S.; Bergmann, M.; Loveland, B.E.; Hedger, M.P.; Schuppe, H.C. Cytokines in male fertility and reproductive pathologies: Immunoregulation and beyond. Front. Endocrinol. 2017, 8, 1–16. [Google Scholar] [CrossRef]
- Zeinali, M.; Hadian Amree, A.; Khorramdelazad, H.; Karami, H.; Abedinzadeh, M. Inflammatory and anti-inflammatory cytokines in the seminal plasma of infertile men suffering from varicocele. Andrologia 2017, 49. [Google Scholar] [CrossRef]
- Micheli, L.; Collodel, G.; Cerretani, D.; Menchiari, A.; Noto, D.; Signorini, C.; Moretti, E. Relationships between Ghrelin and Obestatin with MDA, Proinflammatory Cytokines, GSH/GSSG Ratio, Catalase Activity, and Semen Parameters in Infertile Patients with Leukocytospermia and Varicocele. Oxid. Med. Cell. Longev. 2019, 2019, 7261842. [Google Scholar] [CrossRef] [PubMed]
- Barranco, I.; Padilla, L.; Pérez-Patiño, C.; Vazquez, J.M.; Martínez, E.A.; Rodríguez-Martínez, H.; Roca, J.; Parrilla, I. Seminal Plasma Cytokines Are Predictive of the Outcome of Boar Sperm Preservation. Front. Vet. Sci. 2019, 6, 436. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.; Strzeżek, J.; Filipowicz, K.; Mogielnicka-Brzozowska, M.; Zasiadczyk, L. Age and seasonal-dependent variations in the biochemical composition of boar semen. Theriogenology 2016, 86, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Barranco, I.; Tvarijonaviciute, A.; Perez-Patiño, C.; Vicente-Carrillo, A.; Parrilla, I.; Ceron, J.J.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. Glutathione Peroxidase 5 Is Expressed by the Entire Pig Male Genital Tract and Once in the Seminal Plasma Contributes to Sperm Survival and In Vivo Fertility. PLoS ONE 2016, 11, e0162958. [Google Scholar] [CrossRef] [PubMed]
- Barranco, I.; Casao, A.; Perez-Patiño, C.; Parrilla, I.; Muiño-Blanco, T.; Martinez, E.A.; Cebrian-Perez, J.A.; Roca, J. Profile and reproductive roles of seminal plasma melatonin of boar ejaculates used in artificial insemination programs. J. Anim. Sci. 2017, 95, 1660–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharkey, D.J.; Tremellen, K.P.; Briggs, N.E.; Dekker, G.A.; Robertson, S.A. Seminal plasma transforming growth factor-β, activin A and follistatin fluctuate within men over time. Hum. Reprod. 2016, 31, 2183–2191. [Google Scholar] [CrossRef]
- Kunavongkrit, A.; Suriyasomboon, A.; Lundeheim, N.; Heard, T.W.; Einarsson, S. Management and sperm production of boars under differing environmental conditions. Theriogenology 2005, 63, 657–667. [Google Scholar] [CrossRef]
- Claus, R.; Weiler, U.; Wagner, H. Photoperiodic influences on reproduction of domestic boars. II. Light influences on semen characteristics and libido. J. Vet. Med. Ser. A 1985, 32, 99–109. [Google Scholar]
- Gòdia, M.; Estill, M.; Castelló, A.; Balasch, S.; Rodríguez-gil, J.E.; Krawetz, S.A.; Sánchez, A.; Clop, A. A RNA-Seq Analysis to Describe the Boar Sperm Transcriptome and Its Seasonal Changes. Front. Genet. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sancho, S.; Pinart, E.; Briz, M.; Garcia-Gil, N.; Badia, E.; Bassols, J.; Kádár, E.; Pruneda, A.; Bussalleu, E.; Yeste, M.; et al. Semen quality of postpubertal boars during increasing and decreasing natural photoperiods. Theriogenology 2004, 62, 1271–1282. [Google Scholar] [CrossRef]
- Rempel, L.A.; Krautkramer, M.M.; Loether, T.M.; Parrish, J.J.; Miles, J.R. Season of collection and sperm head shape impacts expression of CARHSP and FTL from motile-rich boar sperm. Agri Gene 2018, 7, 1–6. [Google Scholar] [CrossRef]
- Peña, S.T.J.; Stone, F.; Gummow, B.; Parker, A.J.; Paris, D.B.B.P. Tropical summer induces DNA fragmentation in boar spermatozoa: Implications for evaluating seasonal infertility. Reprod. Fertil. Dev. 2019, 31, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Peltoniemi, O.A.T.; Virolainen, J.V. Seasonality of reproduction in gilts and sows. Soc. Reprod. Fertil. Suppl. 2006, 62, 205–218. [Google Scholar] [PubMed]
- Claus, R.; Schopper, D.; Wagner, H.G. Seasonal effect on steroids in blood plasma and seminal plasma of boars. J. Steroid Biochem. 1983, 19, 725–729. [Google Scholar] [CrossRef]
- Wang, C.; Guo, L.-L.; Wei, H.-K.; Zhou, Y.-F.; Tan, J.-J.; Sun, H.-Q.; Jiang, S.-W.; Peng, J. Logistic regression analysis of the related factors in discarded semen of boars in Southern China. Theriogenology 2019, 131, 47–51. [Google Scholar] [CrossRef]
- Wolf, J.; Smital, J. Quantification of factors affecting semen traits in artificial insemination boars from animal model analyses. J. Anim. Sci. 2009, 87, 1620–1627. [Google Scholar] [CrossRef] [Green Version]
- Kuhlgatz, D.A.; Kuhlgatz, C.; Aepli, M.; Schumann, B.; Grossfeld, R.; Bortfeldt, R.; Jakop, U.; Jung, M.; Schulze, M. Development of predictive models for boar semen quality. Theriogenology 2019, 134, 129–140. [Google Scholar] [CrossRef]
- Frydrychová, S.; Lustykova, A.; Cerovsky, J.; Lipensky, J.; Rozkot, M. Seasonal changes of boars semen production. Res. Pig Breed. 2007, 1, 31–33. [Google Scholar]
- Knecht, D.; Jankowska-Mąkosa, A.; Duziński, K. The effect of age, interval collection and season on selected semen parameters and prediction of AI boars productivity. Livest. Sci. 2017, 201, 13–21. [Google Scholar] [CrossRef]
- Zasiadczyk, L.; Fraser, L.; Kordan, W.; Wasilewska, K. Individual and seasonal variations in the quality of fractionated boar ejaculates. Theriogenology 2015, 83, 1287–1303. [Google Scholar] [CrossRef]
- Koziorowka-Gilun, M.; Koziorowski, M.; Strzezek, J.; Fraser, L. Seasonal changes in antioxidant defence systems in seminal plasma and fluids of the boar reproductive tract. Reprod. Biol. 2011, 11, 37–47. [Google Scholar] [CrossRef]
- Padilla, L.; Pallarés, F.; Molina, M.; Li, J.; Martinez, E.; Parrilla, I.; Roca, J. Immunohistochemical localization of interleukin 10 in genital tract of fertile boars. In Proceedings of the 21st Annual Conference of the European Society for Domestic Animal Reproduction, Bern, Switzerland, 24–26 August 2017; p. 63. [Google Scholar]
- Padilla, L.; Martínez-Hernández, J.; Pallarés, F.; Perez-Patiño, C.; Barranco, I.; Martinez, E.; Roca, J. Localization of Granulocyte-macrophage colony-stimulating factor (GM-CSF) in the genital tract of fertile boars. In Proceedings of the 22nd Annual Conference of the European Society for Domestic Animal Reproduction, Córdoba, Spain, 27–29 September 2018; p. 177. [Google Scholar]
- Barranco, I.; Perez-Patino, C.; Padilla, L.; Alvarez-Rodriguez, M.; Parrilla, I.; Martinez, E.; Roca, J.; Rodriguez-Martinez, H. The Transforming growth-factor (TGF)-β1,-β2 and -β3 is sythetized by most of the boar internal genital organs. In Proceedings of the 22nd Annual Conference of the European Society for Domestic Animal Reproduction, Córdoba, Spain, 27–29 September 2018; p. 109. [Google Scholar]
- Park, C.S.; Yi, Y.J. Comparison of semen characteristics, sperm freezability and testosterone concentration between Duroc and Yorkshire boars during seasons. Anim. Reprod. Sci. 2002, 73, 53–61. [Google Scholar] [CrossRef]
- Vanpouille, C.; Introini, A.; Morris, S.R.; Margolis, L.; Daar, E.S.; Dube, M.P.; Little, S.J.; Smith, D.M.; Lisco, A.; Gianella, S. Distinct cytokine/chemokine network in semen and blood characterize different stages of HIV infection. AIDS 2016, 30, 193–201. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ejaculate Variables | Overall | Daylength/Temperature Period of the Year 1 | p Value 2 | |
|---|---|---|---|---|
| Increasing | Decreasing | |||
| Volume (mL) | 212.41 ± 4.98 | 190.33 ± 5.05 | 234.99 ± 7.98 | <0.001 |
| Sperm concentration (×106 sperm/mL) | 259.92 ± 2.15 | 227.86 ± 3.09 | 294.10 ± 3.40 | <0.001 |
| Total sperm number (×109 spermatozoa) | 56.35 ± 1.96 | 43.77 ± 1.71 | 68.27 ± 2.62 | <0.001 |
| Spermatozoa with normal morphology (%) | 87.35 ± 0.93 | 92.80 ± 1.88 | 82.11 ± 2.19 | 0.017 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padilla, L.; Lucas, X.; Parrilla, I.; Perez-Patiño, C.; Rodriguez-Martinez, H.; Roca, J.; Barranco, I. Period of Boar Ejaculate Collection Contributes to the Yearly Intra-Male Variability of Seminal Plasma Cytokines. Biology 2020, 9, 105. https://doi.org/10.3390/biology9050105
Padilla L, Lucas X, Parrilla I, Perez-Patiño C, Rodriguez-Martinez H, Roca J, Barranco I. Period of Boar Ejaculate Collection Contributes to the Yearly Intra-Male Variability of Seminal Plasma Cytokines. Biology. 2020; 9(5):105. https://doi.org/10.3390/biology9050105
Chicago/Turabian StylePadilla, Lorena, Xiomara Lucas, Inmaculada Parrilla, Cristina Perez-Patiño, Heriberto Rodriguez-Martinez, Jordi Roca, and Isabel Barranco. 2020. "Period of Boar Ejaculate Collection Contributes to the Yearly Intra-Male Variability of Seminal Plasma Cytokines" Biology 9, no. 5: 105. https://doi.org/10.3390/biology9050105