
Gastric Microbiota beyond H. pylori: An Emerging Critical Character in Gastric Carcinogenesis



1
State Key Laboratory of Digestive Disease, Department of Medicine and Therapeutics, Institute of Digestive Disease, Li Ka Shing Institute of Health Sciences, CUHK Shenzhen Research Institute, The Chinese University of Hong Kong, Hong Kong
2
Department of Gastroenterology and Hepatology, Erasmus University Medical Center Rotterdam, Postbus 2040, 3000 CA Rotterdam, The Netherlands
3
Institute of Digestive Disease, Department of Medicine and Therapeutics, Prince of Wales Hospital, The Chinese University of Hong Kong, Shatin, NT, Hong Kong
*
Author to whom correspondence should be addressed.
Biomedicines 2021, 9(11), 1680; https://doi.org/10.3390/biomedicines9111680
Submission received: 15 October 2021
/
Revised: 8 November 2021
/
Accepted: 9 November 2021
/
Published: 12 November 2021
(This article belongs to the Special Issue Gut Dysbiosis: Molecular Mechanisms and Therapies)
Abstract
:Gastric cancer (GC) is one of the global leading causes of cancer death. The association between Helicobacter pylori, which is a predominant risk factor for GC, with GC development has been well-studied. Recently, accumulating evidence has demonstrated the presence of a large population of microorganisms other than H. pylori in the human stomach. Existing sequencing studies have revealed microbial compositional and functional alterations in patients with GC and highlighted a progressive shift in the gastric microbiota in gastric carcinogenesis with marked enrichments of oral or intestinal commensals. Moreover, using a combination of gastric bacterial signatures, GC patients could be significantly distinguished from patients with gastritis. These findings, therefore, emphasize the importance of a collective microbial community in gastric carcinogenesis. Here, we provide an overview of non-H. pylori gastric microbes in gastric carcinogenesis. The molecular mechanisms of gastric microbes-related carcinogenesis and potential clinical applications of gastric microbiota as biomarkers of GC are also explored.
1. Introduction
Despite the decreased incidence in recent years, gastric cancer (GC) remains one of the leading causes of cancer-associated death worldwide [1]. GC is a heterogeneous cancer that can be histologically classified into two types, namely an intestinal type (more frequent) and diffuse type (less frequent). Intestinal-type GC has been well-described as a cascade of histological changes in the gastric mucosa from inflammation and atrophy, then intestinal metaplasia (IM), followed by dysplasia, and eventually, to in situ gastric carcinoma and invasive carcinoma [2].
As with many other types of cancer, tumor formation in the stomach is a multifactorial process, and various genetic and environmental factors contribute to the development of GC. Among environmental factors, the role of microorganisms in gastric carcinogenesis has been increasingly recognized over the past decades. Historically, due to the hostile conditions of the gastric ecological niche, the human stomach was considered to be essentially sterile until the discovery of Helicobacter pylori in the 1980s [3]. The global attributable fraction of H. pylori in non-cardia GC was estimated at 89% in 2008 [4]. In general, H. pylori infection has been universally acknowledged as the main risk factor for GC and other gastroduodenal diseases, and its pathogenic mechanisms have been studied in great detail [5,6,7]. In comparison, little is known about the contribution of other microbes that are resident in the stomach to gastric carcinogenesis. Nevertheless, the collective microbial community in the stomach and its role in nontumorous gastric diseases and GC have attracted widespread attention over the past few years [8,9,10,11,12]. In this review, the roles of gastric microbiota in the development of GC and precancerous lesions are summarized with discussions on the mechanisms of microbiota-related carcinogenesis. The potential of utilizing specific microbial features as diagnostic biomarkers of GC is also explored.
1.1. Gastric Microbiota in Healthy Individuals
Back in 2000, Monstein et al. were the first to profile the gastric bacterial community other than H. pylori by temporal temperature gradient gel electrophoresis of 16S ribosomal DNA (rDNA) amplicons [13]. Enterococcus, Pseudomonas, Streptococcus, Staphylococcus, and Stomatococcus were identified as the most abundant genera in gastric biopsy specimens from both H. pylori-positive patients with gastritis and healthy individuals. Most of these genera are inhabitants of the oral cavity and respiratory tract, thus suggesting the existence of a microbial community indigenous to the gastric microenvironment.
Early evidence from cultivation studies reported that the most abundant bacteria isolated from healthy human stomachs belonged to Veillonella, Lactobacillus, Clostridium [14], Propionibacterium, Streptococcus, and Staphylococcus [15]. In the last decade, the advent of microarray, high-throughput sequencing, and bioinformatic processing have allowed the detection of a wide variety of uncultivated taxa to gain a more comprehensive picture of the gastric microbiota in humans. However, research on the gastric microbiota of healthy populations is still limited, primarily due to the ethical inappropriateness of collecting gastric specimens from healthy individuals. In brief, there is significant inter-subject variability in the gastric microbiota of healthy individuals, with the potential contributing factors including birth mode, geography, diet, lifestyle, antibiotic exposures, use of proton pump inhibitor, and presence of H. pylori [16,17,18,19]. In addition, the data available from the literature also varies greatly. Some studies profiled all identified taxa at different taxonomic levels, while other studies only focused on microbes that could discriminate one particular gastric disease from controls, with considerable differences in the definition of “healthy controls”. Nonetheless, representatives of five major phyla were consistently observed in healthy populations, namely Proteobacteria, Firmicutes, Bacteroidetes, Actinobacteria, and Fusobacteria [18,20,21,22,23], with Streptococcus, Prevotella, Veillonella, and Rothia found as the most prevalent genera [24].
1.2. Gastric Dysbiosis across Gastric Carcinogenesis
Significant differences in the composition, function, and interaction network of gastric microbiota exist among the different stages of gastric carcinogenesis, including premalignant lesions (atrophic gastritis (AG), intestinal metaplasia (IM), and dysplasia), and GC [25,26,27,28]. In this section, evidence on the dynamic and gradual alterations in the gastric microbiota, as well as their correlations with GC progression, are discussed and summarized in Figure 1.
1.2.1. Gastric Microbiota in Premalignant Status
Prolonged AG and IM have been well-recognized as premalignant lesions and risk factors of GC [29]. Chronic AG, a condition characterized by the reduced capacity of gastric acid secretion (mostly caused by a persistent infection of H. pylori), could promote the survival and colonization of nonindigenous microbes, including opportunistic pathogens from the oral cavity [18,30]. In contrast, Parsons et al. [31] reported that H. pylori-induced AG showed a significantly reduced microbial diversity, probably due to the dominance of H. pylori lowering the evenness and richness of the gastric microbiota. At the genus level, increased Streptococcus and decreased Prevotella were observed in patients with H. pylori-induced AG [24]. Yet again, inconsistent findings were reported, as another study identified that Streptococcus was negatively correlated with Helicobacter in patients with H. pylori-induced AG [31]. Nevertheless, several bacterial genera, including Prevotella, Tannerella, and Treponema, were consistently found to be depleted in patients with H. pylori-induced AG from multiple studies, with Helicobacter as the only increased taxon. Moreover, the microbial metabolic pathways could also be altered. For example, an increase in fumarate reductase and reduction in succinate dehydrogenase and glutamate synthase were detected in patients with H. pylori-induced AG [31]. Intriguingly, decreased succinate dehydrogenase and succinate accumulation have already been identified in GC, as well as other tumor types, such as colorectal cancer and hepatocellular carcinoma [32].
Autoimmune AG is another etiology of hypochlorhydria, though less prevalent. It is also associated with an increased risk of GC [33,34], particularly linked to the development of type I gastric carcinoid tumors [35]. Patients with autoimmune AG showed similar changes in their metabolic pathways as patients with H. pylori-induced AG, including increased abundances of alcohol dehydrogenase, (R-R)-butanediol dehydrogenase, glycerol dehydrogenase, D−arabinitol 4−dehydrogenase, L−iditol 2−dehydrogenase, and myo−inositol 2−dehydrogenase [31]. Whereas the overall composition of the gastric microbiota between patients with autoimmune AG and H. pylori-induced AG was found to be distinct. Individuals with autoimmune AG harbored a more diverse bacterial community and significantly higher proportion of Streptococcus than patients with H. pylori-induced AG and even healthy controls. These findings thus implicate the potential involvement of gastric microbes in gastric carcinogenesis even under the absence of H. pylori.
In comparison, much fewer studies have profiled the gastric microbiota in IM, even though IM is well-acknowledged to be the precancerous stage of GC. The gastric microenvironment of patients with IM was once again dominated by H. pylori [36]. After the removal of Helicobacteraceae-related operative taxonomic units (OTUs), Actinomyces, Prevotella, and Streptococcus were identified in relation to the histopathological diagnosis of multifocal AG with IM [37]. In contrast, the prevalence and abundance of H. pylori were significantly lower in GC patients. Moreover, a principal coordinate analysis performed in one study containing subjects with H. pylori-associated gastritis, IM, GC, and H. pylori-negative controls showed a clear separation in the gastric microbial profiles between GC and other noncancerous groups, as well as a large similarity between H. pylori-associated gastritis and IM [38]. A clustering analysis based on UniFrac distance also revealed that the chronic gastritis patients and GC patients were significantly separated, while the IM patients were distributed between the two groups [27]. On the other hand, while there was no significant difference in beta diversity among the patients with superficial gastritis (SG), AG, and IM, the microbial richness estimated by Chao-1 and the alpha diversity was significantly reduced in IM compared to SG or normal controls [26,38]. Additionally, in comparison with SG, seven taxa were significantly enriched in IM, including bacterial species of Pseudomonas and Dyella [26].
Taken together, these results suggest that H. pylori plays a crucial role in the development of chronic inflammation, atrophy, and hypochlorhydria, which creates a favorable growth niche for other opportunistic pathogens to contribute to GC development and progression.
1.2.2. Gastric Microbiota in Gastric Cancer
Similar to the precancerous stages, accumulating evidence has demonstrated the global dysbiosis of gastric microbiota in GC. A gradual alteration in gastric microbial profiles from non-atrophic gastritis (NAG), IM, and intestinal-type GC was reported by Aviles-Jimenez et al., with a steady reduction in the microbial diversity [28]. Likewise, another recent study showed that the gastric microbial richness reduced progressively from healthy controls, NAG, and IM to GC [39]. In contrast, there have been several studies reporting increased microbial richness and diversity in GC tissues as compared to controls [27,40,41,42]. These controversial findings are possibly attributed to the methodological disparity among these studies, as they employed different sequencing platforms, including microarray, pyrosequencing, and next-generation sequencing. Moreover, their study cohorts, sample sizes, and analytic methods also varied widely, which may also account for the discrepant findings.
More recently, an increasing number of studies based on 16S ribosomal RNA (rRNA) gene or shotgun metagenomic sequencing have been performed to depict the gastric microbiota in gastric carcinogenesis [25,26,31,36,37,40,41,43,44,45,46] (Table 1). Despite heterogeneities in ethnic/geographic populations and profiling pipelines, some consistent results among studies have been shown. The enrichment of microbes that originated from the oral cavity or intestines was constantly detected in the gastric microbiota of GC patients [25,26,36,40,41,43,45,47], and these results are consistent with previous datasets using non-sequencing techniques such as terminal restriction fragment length polymorphism analysis and microarray [28,48]. Intriguingly, H. pylori has been frequently observed as significantly depleted or even completely absent in the later stages of gastric tumorigenesis, particularly in patients with dysplasia [25,27,39,41,43,46]. Additionally, epidemiological studies have shown the association of H. pylori abundance with different GC subtypes. For instance, the incidence of intestinal-type GC exhibits a markedly declining trend, whereas the diffuse type shows an increasing trend in both high- and low-risk areas for GC [49]. The decreased incidence of intestinal-type GC has been principally attributed to improved food preservation and declined H. pylori infection rates [1,4]. However, these factors could not explain well the changes in incidence of the diffuse-type GC. Thus, it is of interest to identify whether there is a potential association of non-H. pylori microbes with diffuse-type GC, which may possibly explain the changes of its incidence. All these findings imply the involvement of non-H. pylori components in the development and progression of GC.
Ferreira et al. investigated the gastric microbiota at the genus level in 135 cases with GC or chronic gastritis by 16S rRNA gene profiling [25]. They identified that GC patients have a dysbiotic microbial community with a significant decrease in microbial diversity compared to patients with chronic gastritis. The enrichment of bacterial genera, which are mainly intestinal commensals, including Phyllobacterium, Achromobacter, Citrobacter, Lactobacillus, Clostridium, and Rhodococcus, together with the depletion of Helicobacter, were observed in GC patients. By contrast, Helicobacter, Prevotella, and Streptococcus are the major components in the gastric microbiota of patients with chronic gastritis. Importantly, the results of this study were obtained from a Portuguese discovery cohort with validation by an independent Chinese cohort with 79 GC cases, thus confirming the accuracy of these findings across different geographical populations.
To fully elucidate the alterations in gastric microbiota across GC development, our previous study assessed the microbial composition in gastric mucosal biopsies from 81 cases comprising different histological stages of gastric carcinogenesis, including SG, AG, IM, and GC [26]. Dysbiosis occurred in the gastric microbiota of IM and GC patients with significantly reduced microbial richness in comparison to SG subjects. Lower species richness in GC than in SG was also observed in another metagenomic study [45]. At the genus level, the significant enrichment of 21 bacteria and depletion of 10 bacterial taxa were detected in GC [26]. In agreement with a previous study that showed increased oral-originated bacteria in GC [43], a higher abundance of oral bacteria was observed in GC than in the other three precancerous stages. The enrichment of oral bacterial genera such as Fusobacterium, Haemophilus, Veillonella, Leptotrichia, Dialister, and Lactococcus in GC patients was also highlighted in another study using cDNA from 16S rRNA transcripts [41]. Similarly, Hu et al. [45] reported that the most abundant taxa in GC are commensals or opportunistic pathogens originally colonizing the oral cavity, including Neisseria, Alloprevotella, and Aggregatibacter. Another study published by Chen et al. [40] also illustrated the enrichment of oral bacteria (Peptostreptococcus, Streptococcus, and Fusobacterium) in GC tissues as compared to nontumorous tissues.
In terms of microbial interactions, both cooccurrence and co-exclusive interactions among GC-enriched and GC-depleted bacteria progressively increase from SG, precancerous lesions (AG and IM), and to GC [26]. Microbial interaction networks are also denser, with enhanced complexities in GC patients than in functional dyspepsia controls [41], which could be attributed to the decreased dominance of H. pylori and subsequently elevated abundances of non-gastric commensal microbes (e.g., oral taxa) in GC. Collectively, the current findings have provided solid evidence of the predominant presence of oral bacteria in the gastric microbiota of GC patients.
On the other hand, although the widespread application of next-generation sequencing platforms has greatly facilitated microbiome research, it also has its inherent defects. One critical limitation is that almost all recent studies based on 16S rRNA or metagenomic sequencing have reported the composition and relative abundance of gastric microbes but barely considered their absolute abundances or concentrations of relevant microbial products, which are both of great importance in gastric diseases and tumorigenesis. In fact, culture-based research published in the 1970s has already revealed that the number of bacteria detected in a hypochlorhydric stomach was above 1000 times greater than that in normal groups [50]. Hence, it is reasonable to speculate that the increased absolute abundances of some crucial pathogenic microbes are not noticed in modern studies that only analyze the relative abundance. While other less critical bacteria, though with overgrowth, may overshadow the identifications of species that are actually important to GC development.
1.3. Interplay of H. pylori with Gastric Microbes in Gastric Carcinogenesis
H. pylori infection is known to trigger the initiation and development of GC [6,57,58,59,60]. Persistent H. pylori infection leads to a reduced secretion of gastric acid and chronic inflammation and consequently increases gastric pH, which are common features of patients with AG. Given that gastric acid acts as a formidable barrier against the invasion of oral-ingested microorganisms into the gastrointestinal tract, elevated pH may subsequently contribute to the overgrowth of non-H. pylori microbes in the gastric ecological niche, whereas reduced gastric acidity weakens its own survival; thus H. pylori undergoes a suicidal journey in pathogenesis [61]. Moreover, H. pylori can induce the formation of bacterial biofilms to facilitate the colonization of oral bacteria in the stomach [62].
With the rapid development of high-throughput next-generation sequencing technology and bioinformatic analysis methodology [63,64], the research on gastric microbiota has been massively accelerated. Most studies have focused on the characterization of the structure and composition of the gastric microbial community regarding the H. pylori status, as well as alterations of gastric microbiota in subjects after receiving H. pylori eradication [10,31,38,46,65,66,67,68,69,70]. An early study in 2011 performed a high-density 16S rRNA gene microarray on gastric biopsies of subjects with or without H. pylori infection [65]. The study reported that the gastric microbiota in H. pylori-positive and -negative subjects were both dominated by four phyla (Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes), whereas their compositions were remarkably different. The presence of H. pylori was significantly associated with higher abundances of Proteobacteria, Acidobacteria, and Spirochetes and lower abundances of Actinobacteria, Firmicutes, and Bacteroidetes. Similarly, gastric microbiota in chronic gastritis patients with or without H. pylori infection were distinct at the family level, of which relative abundances of Bradyhizobiaceae, Caulobacteraceae, Lactobacillaceae, and Burkholderiaceae were significantly increased in H. pylori-negative patients [27]. In general, these studies revealed a marked influence of H. pylori colonization on the gastric microbial community, hence providing new insights into research on the roles of gastric microbiota other than H. pylori in gastric diseases and tumorigenesis.
The abundance of H. pylori in later stages of carcinogenesis is consistently reduced in multiple profiling studies, with the least or even undetectable H. pylori at the GC stage. Likewise, H. pylori seems to be more abundant in the early stages of gastric tumorigenesis (i.e., AG and IM). These findings implicate potential interactions among H. pylori, gastric commensal microbes, and translocated oral bacteria. The effects of H. pylori on the richness, diversity, and microbial interactions of the gastric microhabitat have therefore been investigated. Li et al. [38] found that H. pylori-positive patients showed significant alterations in the structure of gastric mucosal microbiota and reduction in microbial diversity. The negative association of the abundance of H. pylori with microbial diversity and interaction networks was also reported in another two studies [31,68]. Similarly, our previous publication revealed that interactions among gastric microbes were drastically weaker in H. pylori-positive than H. pylori-negative specimens across all stages of gastric tumorigenesis (SG, AG, and IM) [26]. In particular, distinct representative taxa could interact with H. pylori at different stages of gastric tumorigenesis, further indicating the critical role of H. pylori in altering microbial interactions. Notably, Liu et al. [46] performed 16S rRNA gene sequencing on normal, peri-tumoral, and intra-tumoral stomach tissues from 276 GC patients and identified that higher H. pylori colonization significantly impacted the whole structure of gastric microbiota in both normal and peri-tumoral microenvironments [40,46].
The association of H. pylori and other microbes in GC development was also elucidated by restored gastric microbiota after H. pylori eradication. Whilst well-established evidence supports the preventive effect of H. pylori eradication against GC prevention, Li et al. [38] indicated that H. pylori eradication could play a role in preventing GC, possibly by restoring the dysbiotic gastric microbiota. In our previous study, 404 gastric biopsy samples from H. pylori-positive patients before and after one-year H. pylori eradication were analyzed [71]. After one year of H. pylori eradication, the patients had a significant increase in bacterial diversity with distinct composition of gastric microbiota compared to those without eradication treatment, which could be attributed to the absence of H. pylori, the dominant taxa before treatment. A significant decrease in cooccurrence interactions in the microbial ecological network was also observed after H. pylori eradication, whereas H. pylori had predominant co-exclusive correlations with other gastric microbes in the placebo group, indicating the inverse interplay between H. pylori and other gastric microbes. In the meanwhile, the depletion of probiotic Faecalibacterium was observed in subjects developing AG after H. pylori eradication. Further metagenomic functional predictions suggested an increase in energy generation and stress adaptation in the inflammation-related microbiota, as well as an enrichment of KEGG ontologies encoding bacterial virulent factors. This study elucidated the contribution of non-H. pylori microbes in the persistence and progression of the precancerous status after H. pylori eradication and rendered new clues for utilizing the microbiota as potential interventional targets for the prevention and therapeutic modulation of gastric carcinogenesis.
1.4. Mechanisms of Gastric Microbes in Gastric Carcinogenesis
Solid evidence has revealed the link between gastric commensal microbes other than H. pylori and the development of GC. However, much fewer studies have deciphered the mechanisms of non-H. pylori in promoting carcinogenesis. In general, from the limited amount of animal studies, gastric microbes could induce DNA damage, inflammation, and immunosuppression to promote gastric carcinogenesis.
1.4.1. The Involvement of Non-H. pylori Commensals in GC Development
Hypergastrinemic insulin-gastrin (INS-GAS) mice are one of the most commonly used mouse models of gastric carcinoma that can develop spontaneous AG and gastrointestinal intraepithelial neoplasia (GIN) 6 months after H. pylori infection. Using this mouse model, an early study showed that H. pylori-monocolonized mice exhibited a delayed development of GIN compared to H. pylori-infected specific pathogen-free mice [72]. These findings suggested the involvement of a complex gastric microbial community other than H. pylori in promoting gastric carcinogenesis. Another study by Lertpiriyapong et al. [73] found that colonizing INS-GAS mice with a combination of microbes limited to only three species of commensal bacteria, including Clostridium species, Bacteroides species, and Lactobacillus murinus, had equivalent gastritis, atrophy, and dysplasia to those colonized with complex intestinal microbiota, and both groups were independent from H. pylori infection. Further H. pylori colonization promoted invasive GIN to a similar extent in mice co-colonized with restricted commensals or diverse microbiota. In these H. pylori-infected mice, the enrichment of Lactobacillus species was both observed in mice with restricted colonization of commensals and colonization of complex bacteria, which were both linked to the depleted abundance of H. pylori. Consistent with previous studies, an inverse association of the abundance of H. pylori and microbial diversity was also observed in this study. These results altogether indicated that, in the setting of H. pylori-induced gastritis and atrophy, restricted commensal bacteria are sufficient for progression to GIN comparable with complex microbiota.
1.4.2. Non-H. pylori Commensals Produce Carcinogens to Contribute GC Development
Another mechanism of non-H. pylori gastric microbes in gastric carcinogenesis is that nitrate-reducing bacteria could contribute to gastric malignancy by increasing the concentrations of nitrite and N-nitroso compounds in the stomach. Ferreira et al. reported that, when compared to chronic gastritis, the functional composition of gastric microbiota in GC displayed enhanced nitrate reductase and nitrite reductase functions, which could promote the generation of nitrite (precursor of carcinogenic N-nitroso compounds) and nitric oxide (a DNA damage inducer), respectively [25]. These data suggest the contribution of a gastric microbial community to genotoxicity in GC. Consistently, another study revealed that proportions of nitrosating and nitrate-reducing non-H. pylori gastric bacteria were two times higher in the GC group than in the controls [51]. Besides, Escherichia–Shigella, Lactobacillus, and Nitrospirae, which are all known to contribute to the metabolism of nitrate or nitrite, were also reported to be enriched in GC [67]. Therefore, these bacteria could play a role in gastric carcinogenesis by promoting the production of carcinogenic N-nitroso compounds. Additionally, several more pathways, including carbohydrate metabolism, amino acid metabolism, nucleotide metabolism, energy metabolism, and DNA replication and repair, were reported to be significantly enriched in GC [26,41,43].
1.4.3. Non-H. pylori Commensals Induce Inflammatory Response and Immunosuppressive Microenvironment to Contribute GC Development
There are a few concordant findings on the role of specific species in gastric carcinogenesis. Propionibacterium acnes was significantly enriched in the GC microenvironment in two independent studies and identified as a strong risk factor for GC development [46,52]. P. acnes was previously detected as a trigger for corpus-dominant lymphocytic gastritis by activating the natural killer group 2 member D (NKG2D) system and upregulating the secretion of proinflammatory cytokine interleukin (IL)-15 [74]. Inducing the expression of NKG2D ligands on stressed cells was shown to be a consequence of the DNA damage response. The current evidence also illustrates that microbial dysbiosis can trigger a DNA damage response in host cells such as epithelial cells, which may result in prolonged inflammatory responses, as well as tumorigenesis [75]. On the other hand, some controversial findings remain to be addressed. For instance, patients with high levels of Prevotella copri showed a significantly increased risk for GC in a case–control study consisting of 268 GC patients and 288 healthy controls [52], while its abundance was significantly reduced in tumor tissues in another cohort of 276 GC patients [46]. P. copri could promote chronic inflammatory conditions via enhancing the resistance to host-derived reactive oxygen species and generating redox proteins, whereas it is also able to ameliorate glucose metabolism in a beneficial manner [76,77]. Such heterogeneous results are possibly due to the high strain variabilities across individuals. Thus, further mechanistic studies are required to verify the role of P. copri in gastric carcinogenesis.
The infamous Fusobacterium nucleatum has been well-studied in the carcinogenesis of colorectal cancer, while a recent study also showed its diagnostic potential in GC [36]. The interaction between F. nucleatum adhesins (e.g., FadA and Fap2) and host cells can induce inflammatory responses and immune suppression [78,79]. The adhesion from FadA of F. nucleatum to E-cadherin of intestinal epithelial cells drives the activation of the Wnt/β-catenin pathway to promote the proliferation of tumor cells [80]. F. nucleatum can also activate the NF-κB pathway to stimulate the production and release of inflammatory cytokines such as IL-1β, IL-6, IL-8, and TNF, thereby creating a proinflammatory microenvironment that favors tumor development [81]. Additionally, F. nucleatum can modulate the tumor microenvironment by suppressing antitumor immune responses. The binding of F. nucleatum adhesin Fap2 to the TIGIT receptor expressed on natural killer cells and other tumor-infiltrating T cells inhibits the cytotoxicity of these immune cells [82]. Although direct mechanistic evidence of how F. nucleatum contributes to gastric carcinogenesis is lacking, F. nucleatum may possibly promote the development of GC through inducing inflammation and suppressing the host antitumor immunity.
Additionally, Rolig et al. [83] treated C57BL/6 mice with antibiotics prior to H. pylori infection, and significantly reduced H. pylori-induced inflammation occurred in these microbiota-depleted mice. Notably, the infiltration of CD4+ T cells and levels of IFN-γ transcript in the stomach were decreased in H. pylori-infected mice with an antibiotic pretreatment. Given the declined abundance of H. pylori during progression from gastritis and IM to GC [36], it is reasonable to hypothesize that the enrichment of some bacteria such as Lactobacillus species may play an essential role in outcompeting and inhibiting H. pylori colonization, thereby promoting the persistent inflammation and progression of GC. Furthermore, our recent study revealed that the enrichment of Acinetobacter, Ralstonia, Actinobacillus, and Erwinia was correlated with persistent inflammation after one year of H. pylori eradication, and oral microbes, including Granulicatella, Peptostreptococcus, Streptococcus, Rothia, Parvimonas, and Prevotella, were also positively correlated with persistent or emerged atrophy or IM after treatment [71].
In 2020, a considerable impact of the gut microbiota on the human immune system was reported [84]. Likewise, a preliminary study explored the association between gastric mucosal microbiota and immunosuppressive cells, the critical components of the tumor microenvironment [53]. The enrichment of regulatory T cells (Tregs) and BDCA2+ plasmacytoid dendritic cells (pDCs), which can both contribute to the immunosuppressive microenvironment in GC [85,86], were observed in intra-tumoral and peri-tumoral tissues. Meanwhile, the composition, diversity, and function of the gastric microbiota were also altered significantly in intra-tumoral tissues. A correlation analysis revealed that an abundance of Stenotrophomonas and Selenomonas is positively correlated with pDCs and Tregs, respectively, whereas abundances of Comamonas and Gaiella are negatively correlated with pDCs and Tregs, respectively. These findings indicate that alterations in the gastric microbiota could participate in the modulation of immune cell populations, contributing to an immunosuppressive microenvironment of GC. Nevertheless, further research is still needed to elucidate the detailed mechanism by which these certain microbes modulate the tumor-immune microenvironment.
1.5. Clinical Application of Gastric Microbes for Diagnosis of Gastric Cancer
Given the identification of specific gastric microbes that are enriched/depleted in patients with GC, these microbes could be employed as potential biomarkers for GC screening. A gastric microbiota-based diagnostic measurement known as the microbial dysbiosis index (MDI) was developed, which utilizes and combines GC-enriched and -depleted taxa for discriminating GC patients from patients with chronic gastritis [25]. MDI exhibited excellent sensitivity and specificity in identifying GC, achieving areas under the receiver operating characteristic curves (AUROC) of 0.89–0.91.
Our previous study identified five GC-enriched bacteria with significant centralities among the microbial interaction network of GC [26]. This core set of bacteria includes Peptostreptococcus stomatis, Streptococcus anginosus, Parvimonas micra, Slackia exigua, and Dialister pneumosintes, all of which are commensals in the human oral microbiome. This set of bacterial markers was employed to distinguish GC patients from patients with SG, showing a confident AUROC of 0.82, while, in another study comprising cases of gastritis, IM, and GC, a combination of five GC-enriched bacteria, including Clostridium colicanis, Fusobacterium canifelinum, F. nucleatum, Lactobacillus gasseri, and Lactobacillus reuteri, also showed sufficient power to discriminate GC patients from patients with noncancerous gastric diseases with excellent specificity (100%) and good sensitivity (70%) [36]. The diagnostic performance of using only two of the above five bacteria (C. colicanis and F. nucleatum) exhibited 100% sensitivity for the detection of GC. Strikingly, both studies had large sample sizes, hence providing firm evidence for utilizing these bacterial markers to accurately identify GC patients. Overall, with the advances in microbial profiling technology, recent publications have emphasized the importance of a collective microbial community rather than single bacterial species in gastric carcinogenesis, as well as its clinical potential in diagnosing GC.
Although the above-mentioned microbes may serve as potential diagnostic markers for GC, it does not seem feasible to replace traditional screening or diagnostic methods with a gastric microbial examination. The main reason is the difficulty in collecting gastric microbiota, as gastric biopsy specimens are needed to obtain a thorough invasive gastroscopic examination. In comparison, a microbiota-based diagnosis is much more suitable for screening colorectal cancer, since this can be done without any invasive approach simply by examining the fecal samples of patients. On the other hand, if a gastroscopy with the collection of gastric mucosal biopsies has already been performed, it appears to be unreasonable and unnecessary to use any additional diagnostic approaches apart from histopathological tests. Hence, the therapeutic potential of targeting gastric microbiota to prevent or alleviate GC development could possibly be an alternative clinical application. Nevertheless, it is essential to elucidate the role and detailed mechanism of GC-associated bacterial species identified in previous sequencing studies. If any microbe is found to be an oncogenic pathogen and plays a key role in the development of GC, the elimination of this microbe, just like H. pylori eradication, may protect patients, especially those with gastric premalignant lesions, from progressing to GC.
1.6. Further Perspectives
There are now increasing studies with high-throughput sequencing to depict the roles of gastric microbiota in GC. This research direction has become more extensive, with increasingly detailed discussions. However, due to the wide variations among individuals with diverse genetic or environmental backgrounds, approaches for sampling, sample storage, DNA extraction methods, sequencing platform and depth, primer design, and data analytic pipelines, the results of many microbiota sequencing studies lack comparability [87,88]. Moreover, these studies mostly focus on profiling the gastric microbiota, while the mechanism of how the altered microbiota contributes to carcinogenesis is much less acknowledged. Additionally, given that most publications on gastric microbiota are based on 16S rRNA gene sequencing, relevant research on other components in the gastric microbiota, including fungi and viruses, is scarce. For instance, we previously reported the importance of dysbiosis among gastric fungi in gastric carcinogenesis [26]. Taken together, as compared to the intestinal microbiota, many areas of gastric microbe-associated carcinogenesis require extensive investigation, and the potential clinical applications of targeting gastric microbiota for the diagnosis, prevention, and treatment of GC still require further investigation.
Author Contributions
J.W. collected the data and drafted the manuscript; H.C.-H.L. discussed the content and revised the manuscript; and M.P. and J.Y. supervised and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Key R&D Program of China (No. 2020YFA0509200/2020YFA0509203), RGC Theme-based Res Scheme Hong Kong (T21-705/20-N), RGC Collaborative Research Fund (C4039-19GF and C7065-18GF), RGC-GRF Hong Kong (14163817), and Vice-Chancellor’s Discretionary Fund Chinese University of Hong Kong.
Data Availability Statement
The data is available within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Correa, P.; Haenszel, W.; Cuello, C.; Tannenbaum, S.; Archer, M. A model for gastric cancer epidemiology. Lancet 1975, 2, 58–60. [Google Scholar] [CrossRef]
- Warren, J.R.; Marshall, B. Unidentified curved bacilli on gastric epithelium in active chronic gastritis. Lancet 1983, 1, 1273–1275. [Google Scholar] [PubMed]
- Plummer, M.; Franceschi, S.; Vignat, J.; Forman, D.; de Martel, C. Global burden of gastric cancer attributable to Helicobacter pylori. Int. J. Cancer 2015, 136, 487–490. [Google Scholar] [CrossRef]
- Yang, I.; Nell, S.; Suerbaum, S. Survival in hostile territory: The microbiota of the stomach. FEMS Microbiol. Rev. 2013, 37, 736–761. [Google Scholar] [CrossRef]
- Mégraud, F.; Bessède, E.; Varon, C. Helicobacter pylori infection and gastric carcinoma. Clin. Microbiol. Infect. 2015, 21, 984–990. [Google Scholar] [CrossRef] [Green Version]
- Plummer, M.; de Martel, C.; Vignat, J.; Ferlay, J.; Bray, F.; Franceschi, S. Global burden of cancers attributable to infections in 2012: A synthetic analysis. Lancet Glob. Health 2016, 4, e609–e616. [Google Scholar] [CrossRef] [Green Version]
- Sheh, A.; Fox, J.G. The role of the gastrointestinal microbiome in Helicobacter pylori pathogenesis. Gut Microbes 2013, 4, 505–531. [Google Scholar] [CrossRef] [Green Version]
- Nardone, G.; Compare, D. The human gastric microbiota: Is it time to rethink the pathogenesis of stomach diseases? United Eur. Gastroenterol. J. 2015, 3, 255–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, C.; Schütte, K.; Koch, N.; Vilchez-Vargas, R.; Wos-Oxley, M.L.; Oxley, A.P.A.; Vital, M.; Malfertheiner, P.; Pieper, D.H. The active bacterial assemblages of the upper GI tract in individuals with and without Helicobacter infection. Gut 2018, 67, 216–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Gao, X.; Guo, J.; Yu, D.; Xiao, Y.; Wang, H.; Li, Y. Helicobacter pylori infection alters gastric and tongue coating microbial communities. Helicobacter 2019, 24, e12567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajilic-Stojanovic, M.; Figueiredo, C.; Smet, A.; Hansen, R.; Kupcinskas, J.; Rokkas, T.; Andersen, L.; Machado, J.C.; Ianiro, G.; Gasbarrini, A.; et al. Systematic review: Gastric microbiota in health and disease. Aliment. Pharmacol. Ther. 2020, 51, 582–602. [Google Scholar] [CrossRef]
- Monstein, H.J.; Tiveljung, A.; Kraft, C.H.; Borch, K.; Jonasson, J. Profiling of bacterial flora in gastric biopsies from patients with Helicobacter pylori-associated gastritis and histologically normal control individuals by temperature gradient gel electrophoresis and 16S rDNA sequence analysis. J. Med. Microbiol. 2000, 49, 817–822. [Google Scholar] [CrossRef] [Green Version]
- Zilberstein, B.; Quintanilha, A.G.; Santos, M.A.A.; Pajecki, D.; de Moura, E.; Alves, P.R.A.; Filho, F.M.; De Souza, J.A.U.; Gama-Rodrigues, J. Digestive tract microbiota in healthy volunteers. Clinics 2007, 62, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Delgado, S.; Cabrera-Rubio, R.; Mira, A.; Suárez, A.; Mayo, B. Microbiological survey of the human gastric ecosystem using culturing and pyrosequencing methods. Microb. Ecol. 2013, 65, 763–772. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The healthy human microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Chung, J.; Battaglia, T.; Henderson, N.; Jay, M.; Li, H.; Lieber, A.D.; Wu, F.; Perez, G.I.; Chen, Y.; et al. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci. Transl. Med. 2016, 8, 343ra82. [Google Scholar] [CrossRef] [Green Version]
- Nardone, G.; Compare, D.; Rocco, A. A microbiota-centric view of diseases of the upper gastrointestinal tract. Lancet Gastroenterol. Hepatol. 2017, 2, 298–312. [Google Scholar] [CrossRef]
- Tsuda, A.; Suda, W.; Morita, H.; Takanashi, K.; Takagi, A.; Koga, Y.; Hattori, M. Influence of Proton-Pump Inhibitors on the Luminal Microbiota in the Gastrointestinal Tract. Clin. Transl. Gastroenterol. 2015, 6, e89. [Google Scholar] [CrossRef]
- Bik, E.M.; Eckburg, P.B.; Gill, S.R.; Nelson, K.E.; Purdom, E.A.; Francois, F.; Perez, G.P.; Blaser, M.J.; Relman, D.A. Molecular analysis of the bacterial microbiota in the human stomach. Proc. Natl. Acad. Sci. USA 2006, 103, 732–737. [Google Scholar] [CrossRef] [Green Version]
- Andersson, A.F.; Lindberg, M.; Jakobsson, H.; Bäckhed, F.; Nyrén, P.; Engstrand, L. Comparative analysis of human gut microbiota by barcoded pyrosequencing. PLoS ONE 2008, 3, e2836. [Google Scholar] [CrossRef]
- Stearns, J.C.; Lynch, M.D.J.; Senadheera, D.B.; Tenenbaum, H.C.; Goldberg, M.B.; Cvitkovitch, D.G.; Croitoru, K.; Moreno-Hagelsieb, G.; Neufeld, J.D. Bacterial biogeography of the human digestive tract. Sci. Rep. 2011, 1, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunathilake, M.; Lee, J.; Choi, I.J.; Kim, Y.I.; Kim, J. Association between bacteria other than Helicobacter pylori and the risk of gastric cancer. Helicobacter 2021, 26, e12836. [Google Scholar] [CrossRef] [PubMed]
- Engstrand, L.; Lindberg, M. Helicobacter pylori and the gastric microbiota. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 39–45. [Google Scholar] [CrossRef]
- Ferreira, R.M.; Pereira-Marques, J.; Pinto-Ribeiro, I.; Costa, J.L.; Carneiro, F.; Machado, J.C.; Figueiredo, C. Gastric microbial community profiling reveals a dysbiotic cancer-associated microbiota. Gut 2018, 67, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Coker, O.O.; Dai, Z.; Nie, Y.; Zhao, G.; Cao, L.; Nakatsu, G.; Wu, W.K.; Wong, S.H.; Chen, Z.; Sung, J.J.Y.; et al. Mucosal microbiome dysbiosis in gastric carcinogenesis. Gut 2018, 67, 1024–1032. [Google Scholar] [CrossRef]
- Eun, C.S.; Kim, B.K.; Han, D.S.; Kim, S.Y.; Kim, K.M.; Choi, B.Y.; Song, K.S.; Kim, Y.S.; Kim, J.F. Differences in gastric mucosal microbiota profiling in patients with chronic gastritis, intestinal metaplasia, and gastric cancer using pyrosequencing methods. Helicobacter 2014, 19, 407–416. [Google Scholar] [CrossRef]
- Aviles-Jimenez, F.; Vazquez-Jimenez, F.; Medrano-Guzman, R.; Mantilla, A.; Torres, J. Stomach microbiota composition varies between patients with non-atrophic gastritis and patients with intestinal type of gastric cancer. Sci. Rep. 2014, 4, 4202. [Google Scholar] [CrossRef] [Green Version]
- Sugano, K.; Tack, J.; Kuipers, E.J.; Graham, D.Y.; El-Omar, E.; Miura, S.; Haruma, K.; Asaka, M.; Uemura, N.; Malfertheiner, P. Kyoto global consensus report on Helicobacter pylori gastritis. Gut 2015, 64, 1353–1367. [Google Scholar] [CrossRef] [Green Version]
- Salama, N.R.; Hartung, M.L.; Müller, A. Life in the human stomach: Persistence strategies of the bacterial pathogen Helicobacter pylori. Nat. Rev. Microbiol. 2013, 11, 385–399. [Google Scholar] [CrossRef]
- Parsons, B.N.; Ijaz, U.Z.; D’Amore, R.; Burkitt, M.D.; Eccles, R.; Lenzi, L.; Duckworth, C.A.; Moore, A.R.; Tiszlavicz, L.; Varro, A.; et al. Comparison of the human gastric microbiota in hypochlorhydric states arising as a result of Helicobacter pylori-induced atrophic gastritis, autoimmune atrophic gastritis and proton pump inhibitor use. PLoS Pathog. 2017, 13, e1006653. [Google Scholar] [CrossRef] [Green Version]
- Pozza, E.D.; Dando, I.; Pacchiana, R.; Liboi, E.; Scupoli, M.T.; Donadelli, M.; Palmieri, M. Regulation of succinate dehydrogenase and role of succinate in cancer. Semin. Cell Dev. Biol. 2020, 98, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Hemminki, K.; Liu, X.; Ji, J.; Sundquist, J.; Sundquist, K. Autoimmune disease and subsequent digestive tract cancer by histology. Ann. Oncol. 2012, 23, 927–933. [Google Scholar] [CrossRef]
- Vannella, L.; Lahner, E.; Osborn, J.; Annibale, B. Systematic review: Gastric cancer incidence in pernicious anaemia. Aliment. Pharmacol. Ther. 2013, 37, 375–382. [Google Scholar] [CrossRef]
- Burkitt, M.D.; Pritchard, D.M. Review article: Pathogenesis and management of gastric carcinoid tumours. Aliment. Pharmacol. Ther. 2006, 24, 1305–1320. [Google Scholar] [CrossRef]
- Hsieh, Y.-Y.; Tung, S.-Y.; Pan, H.-Y.; Yen, C.-W.; Xu, H.-W.; Lin, Y.-J.; Deng, Y.-F.; Hsu, W.-T.; Wu, C.-S.; Li, C. Increased Abundance of Clostridium and Fusobacterium in Gastric Microbiota of Patients with Gastric Cancer in Taiwan. Sci. Rep. 2018, 8, 158. [Google Scholar] [CrossRef]
- Yang, I.; Woltemate, S.; Piazuelo, M.B.; Bravo, L.E.; Yepez, M.C.; Romero-Gallo, J.; Delgado, A.G.; Wilson, K.; Peek, R.M.; Correa, P.; et al. Different gastric microbiota compositions in two human populations with high and low gastric cancer risk in Colombia. Sci. Rep. 2016, 6, 18594. [Google Scholar] [CrossRef] [PubMed]
- Li, T.H.; Qin, Y.; Sham, P.C.; Lau, K.S.; Chu, K.M.; Leung, W.K. Alterations in Gastric Microbiota after H. Pylori Eradication and in Different Histological Stages of Gastric Carcinogenesis. Sci. Rep. 2017, 7, 44935. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, X.; Zeng, R.; Wu, Q.; Sun, H.; Wu, W.; Zhang, X.; Sun, G.; Yan, B.; Wu, L.; et al. Changes of the Gastric Mucosal Microbiome Associated with Histological Stages of Gastric Carcinogenesis. Front. Microbiol. 2020, 11, 997. [Google Scholar] [CrossRef]
- Chen, X.H.; Wang, A.; Chu, A.N.; Gong, Y.H.; Yuan, Y. Mucosa-Associated Microbiota in Gastric Cancer Tissues Compared with Non-cancer Tissues. Front. Microbiol. 2019, 10, 1261. [Google Scholar] [CrossRef] [Green Version]
- Castaño-Rodríguez, N.; Goh, K.L.; Fock, K.M.; Mitchell, H.M.; Kaakoush, N.O. Dysbiosis of the microbiome in gastric carcinogenesis. Sci. Rep. 2017, 7, 15957. [Google Scholar] [CrossRef] [Green Version]
- Gantuya, B.; El Serag, H.B.; Matsumoto, T.; Ajami, N.J.; Uchida, T.; Oyuntsetseg, K.; Bolor, D.; Yamaoka, Y. Gastric mucosal microbiota in a Mongolian population with gastric cancer and precursor conditions. Aliment. Pharmacol. Ther. 2020, 51, 770–780. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Torres, J.; Hu, N.; Medrano-Guzman, R.; Herrera-Goepfert, R.; Humphrys, M.S.; Wang, L.; Wang, C.; Ding, T.; Ravel, J.; et al. Molecular Characterization of the Human Stomach Microbiota in Gastric Cancer Patients. Front. Cell. Infect. Microbiol. 2017, 7, 302. [Google Scholar] [CrossRef]
- Thorell, K.; Bengtsson-Palme, J.; Liu, O.H.-F.; Gonzales, R.V.P.; Nookaew, I.; Rabeneck, L.; Paszat, L.; Graham, D.Y.; Nielsen, J.; Lundin, S.B.; et al. In Vivo Analysis of the Viable Microbiota and Helicobacter pylori Transcriptome in Gastric Infection and Early Stages of Carcinogenesis. Infect. Immun. 2017, 85, e00031-17. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.L.; Pang, W.; Huang, Y.; Zhang, Y.; Zhang, C.J. The Gastric Microbiome Is Perturbed in Advanced Gastric Adenocarcinoma Identified through Shotgun Metagenomics. Front. Cell. Infect. Microbiol. 2018, 8, 433. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Shao, L.; Liu, X.; Ji, F.; Mei, Y.; Cheng, Y.; Liu, F.; Yan, C.; Li, L.; Ling, Z. Alterations of gastric mucosal microbiota across different stomach microhabitats in a cohort of 276 patients with gastric cancer. EBioMedicine 2019, 40, 336–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakhti, S.Z.; Latifi-Navid, S. Oral microbiota and Helicobacter pylori in gastric carcinogenesis: What do we know and where next? BMC Microbiol. 2021, 21, 71. [Google Scholar] [CrossRef]
- Dicksved, J.; Lindberg, M.; Rosenquist, M.; Enroth, H.; Jansson, J.K.; Engstrand, L. Molecular characterization of the stomach microbiota in patients with gastric cancer and in controls. J. Med. Microbiol. 2009, 58, 509–516. [Google Scholar] [CrossRef]
- Assumpção, P.P.; Barra, W.F.; Ishak, G.; Coelho, L.G.V.; Coimbra, F.J.F.; Freitas, H.C.; Dias-Neto, E.; Camargo, M.C.; Szklo, M. The diffuse-type gastric cancer epidemiology enigma. BMC Gastroenterol. 2020, 20, 223. [Google Scholar] [CrossRef]
- Giannella, R.A.; Broitman, S.A.; Zamcheck, N. Gastric acid barrier to ingested microorganisms in man: Studies in vivo and in vitro. Gut 1972, 13, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Jo, H.J.; Kim, J.; Kim, N.; Park, J.H.; Nam, R.H.; Seok, Y.; Kim, Y.-R.; Kim, J.S.; Kim, J.M.; Lee, D.H.; et al. Analysis of Gastric Microbiota by Pyrosequencing: Minor Role of Bacteria Other Than Helicobacter pylori in the Gastric Carcinogenesis. Helicobacter 2016, 21, 364–374. [Google Scholar] [CrossRef]
- Gunathilake, M.N.; Lee, J.; Choi, I.J.; Kim, Y.-I.; Ahn, Y.; Park, C.; Kim, J. Association between the relative abundance of gastric microbiota and the risk of gastric cancer: A case-control study. Sci. Rep. 2019, 9, 13589. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Shao, L.; Liu, X.; Cheng, Y.; Yan, C.; Mei, Y.; Ji, F.; Liu, X. Regulatory T Cells and Plasmacytoid Dendritic Cells Within the Tumor Microenvironment in Gastric Cancer Are Correlated with Gastric Microbiota Dysbiosis: A Preliminary Study. Front. Immunol. 2019, 10, 533. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Lee, A.R.; Lee, Y.R.; Eun, C.S.; Lee, S.K.; Han, D.S. Evaluation of gastric microbiome and metagenomic function in patients with intestinal metaplasia using 16S rRNA gene sequencing. Helicobacter 2019, 24, e12547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Xin, Y.; Zhou, J.; Tian, Z.; Liu, C.; Yu, X.; Meng, X.; Jiang, W.; Zhao, S.; Dong, Q. Gastric Mucosa-Associated Microbial Signatures of Early Gastric Cancer. Front. Microbiol. 2020, 11, 1548. [Google Scholar] [CrossRef]
- Ravegnini, G.; Fosso, B.; Di Saverio, V.; Sammarini, G.; Zanotti, F.; Rossi, G.; Ricci, M.; D’Amico, F.; Valori, G.; Ioli, A.; et al. Gastric Adenocarcinomas and Signet-Ring Cell Carcinoma: Unraveling Gastric Cancer Complexity through Microbiome Analysis-Deepening Heterogeneity for a Personalized Therapy. Int. J. Mol. Sci. 2020, 21, 9735. [Google Scholar] [CrossRef] [PubMed]
- Hartung, M.L.; Gruber, D.C.; Koch, K.N.; Grüter, L.; Rehrauer, H.; Tegtmeyer, N.; Backert, S.; Müller, A.H. Pylori-Induced DNA Strand Breaks Are Introduced by Nucleotide Excision Repair Endonucleases and Promote NF-κB Target Gene Expression. Cell Rep. 2015, 13, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Graham, D.Y. Helicobacter pylori update: Gastric cancer, reliable therapy, and possible benefits. Gastroenterology 2015, 148, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Muratakamiya, N.; Kurashima, Y.; Teishikata, Y.; Yamahashi, Y.; Saito, Y.; Higashi, H.; Aburatani, H.; Akiyama, T.; Peek, R.M.; Azuma, T.; et al. Helicobacter pylori CagA interacts with E-cadherin and deregulates the beta-catenin signal that promotes intestinal transdifferentiation in gastric epithelial cells. Oncogene 2007, 26, 4617–4626. [Google Scholar] [CrossRef] [Green Version]
- Touati, E.; Michel, V.; Thiberge, J.M.; Wuscher, N.; Huerre, M.; Labigne, A. Chronic Helicobacter pylori infections induce gastric mutations in mice. Gastroenterology 2003, 124, 1408–1419. [Google Scholar] [CrossRef]
- De Assumpção, P.P.; Araújo, T.M.T.; De Assumpção, P.B.; Barra, W.F.; Khayat, A.S.; Assumpção, C.B.; Ishak, G.; Nunes, D.N.; Dias-Neto, E.; Coelho, L.G.V. Suicide journey of H. pylori through gastric carcinogenesis: The role of non-H. pylori microbiome and potential consequences for clinical practice. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1591–1597. [Google Scholar] [CrossRef] [PubMed]
- Hathroubi, S.; Servetas, S.L.; Windham, I.; Merrell, D.S.; Ottemann, K.M. Helicobacter pylori Biofilm Formation and Its Potential Role in Pathogenesis. Microbiol. Mol. Biol. Rev. 2018, 82, e00001-18. [Google Scholar] [CrossRef] [Green Version]
- Methé, B.A.; Nelson, K.E.; Pop, M.; Creasy, H.H.; Giglio, M.G.; Huttenhower, C.; Gevers, D.; Petrosino, J.F.; Abubucker, S.; Mannon, P.J.; et al. A framework for human microbiome research. Nature 2012, 486, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Maldonado-Contreras, A.; Goldfarb, K.C.; Godoy-Vitorino, F.; Karaoz, U.; Contreras, M.; Blaser, M.J.; Brodie, E.L.; Dominguez-Bello, M.G. Structure of the human gastric bacterial community in relation to Helicobacter pylori status. ISME J. 2011, 5, 574–579. [Google Scholar] [CrossRef]
- Kienesberger, S.; Cox, L.; Livanos, A.; Zhang, X.-S.; Chung, J.; Perez, G.P.; Gorkiewicz, G.; Zechner, E.L.; Blaser, M.J. Gastric Helicobacter pylori Infection Affects Local and Distant Microbial Populations and Host Responses. Cell Rep. 2016, 14, 1395–1407. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhou, J.; Xin, Y.; Geng, C.; Tian, Z.; Yu, X.; Dong, Q. Bacterial overgrowth and diversification of microbiota in gastric cancer. Eur. J. Gastroenterol. Hepatol. 2016, 28, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Pereira, V.; Saxena, S.; Ghosh, T.; Anbumani, D.; Bag, S.; Das, B.; Nair, G.B.; Abraham, P.; Mande, S.S. Gastric microbiome of Indian patients with Helicobacter pylori infection, and their interaction networks. Sci. Rep. 2017, 7, 15438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterbini, F.P.; Palladini, A.; Masucci, L.; Cannistraci, C.V.; Pastorino, R.; Ianiro, G.; Bugli, F.; Martini, C.; Ricciardi, W.; Gasbarrini, A.; et al. Effects of Proton Pump Inhibitors on the Gastric Mucosa-Associated Microbiota in Dyspeptic Patients. Appl. Environ. Microbiol. 2016, 82, 6633–6644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Zhang, Y.; Gerhard, M.; Gao, J.-J.; Mejias-Luque, R.; Zhang, L.; Vieth, M.; Ma, J.-L.; Bajbouj, M.; Suchanek, S.; et al. Effect of Helicobacter pylori on gastrointestinal microbiota: A population-based study in Linqu, a high-risk area of gastric cancer. Gut 2020, 69, 1598–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, J.J.Y.; Coker, O.O.; Chu, E.; Szeto, C.H.; Luk, S.T.Y.; Lau, H.; Yu, J. Gastric microbes associated with gastric inflammation, atrophy and intestinal metaplasia 1 year after Helicobacter pylori eradication. Gut 2020, 69, 1572–1580. [Google Scholar] [CrossRef] [Green Version]
- Lofgren, J.L.; Whary, M.T.; Ge, Z.; Muthupalani, S.; Taylor, N.S.; Mobley, M.; Potter, A.; Varro, A.; Eibach, D.; Suerbaum, S.; et al. Lack of commensal flora in Helicobacter pylori-infected INS-GAS mice reduces gastritis and delays intraepithelial neoplasia. Gastroenterology 2011, 140, 210–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lertpiriyapong, K.; Whary, M.T.; Muthupalani, S.; Lofgren, J.L.; Gamazon, E.; Feng, Y.; Ge, Z.; Wang, T.C.; Fox, J.G. Gastric colonisation with a restricted commensal microbiota replicates the promotion of neoplastic lesions by diverse intestinal microbiota in the Helicobacter pylori INS-GAS mouse model of gastric carcinogenesis. Gut 2014, 63, 54–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montalban-Arques, A.; Wurm, P.; Trajanoski, S.; Schauer, S.; Kienesberger, S.; Halwachs, B.; Högenauer, C.; Langner, C.; Gorkiewicz, G. Propionibacterium acnes overabundance and natural killer group 2 member D system activation in corpus-dominant lymphocytic gastritis. J. Pathol. 2016, 240, 425–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinoza, J.L.; Minami, M. Sensing Bacterial-Induced DNA Damaging Effects via Natural Killer Group 2 Member D Immune Receptor: From Dysbiosis to Autoimmunity and Carcinogenesis. Front. Immunol. 2018, 9, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E. Gut microbiota in 2015: Prevotella in the gut: Choose carefully. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 69–70. [Google Scholar] [CrossRef]
- Hofer, U. Microbiome: Pro-inflammatory Prevotella? Nat. Rev. Microbiol. 2014, 12, 5. [Google Scholar] [CrossRef]
- Wu, J.; Li, Q.; Fu, X. Fusobacterium nucleatum Contributes to the Carcinogenesis of Colorectal Cancer by Inducing Inflammation and Suppressing Host Immunity. Transl. Oncol. 2019, 12, 846–851. [Google Scholar] [CrossRef]
- Brennan, C.A.; Garrett, W.S. Fusobacterium nucleatum-symbiont, opportunist and oncobacterium. Nat. Rev. Microbiol. 2019, 17, 156–166. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gur, C.; Ibrahim, Y.; Isaacson, B.; Yamin, R.; Abed, J.; Gamliel, M.; Enk, J.; Bar-On, Y.; Stanietsky-Kaynan, N.; Coppenhagen-Glazer, S.; et al. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity 2015, 42, 344–355. [Google Scholar] [CrossRef] [Green Version]
- Rolig, A.S.; Cech, C.; Ahler, E.; Carter, J.E.; Ottemann, K.M. The degree of Helicobacter pylori-triggered inflammation is manipulated by preinfection host microbiota. Infect. Immun. 2013, 81, 1382–1389. [Google Scholar] [CrossRef] [Green Version]
- Schluter, J.; Peled, J.; Taylor, B.P.; Markey, K.A.; Smith, J.A.; Taur, Y.; Niehus, R.; Staffas, A.; Dai, A.; Fontana, E.; et al. The gut microbiota is associated with immune cell dynamics in humans. Nature 2020, 588, 303–307. [Google Scholar] [CrossRef]
- Huang, X.; Liu, X.; Lin, X.; Yu, H.; Sun, J.; Chen, C.; Jin, H.; Zhang, G.; Shi, X.; Zhang, Q.; et al. Role of plasmacytoid dendritic cells and inducible costimulator-positive regulatory T cells in the immunosuppression microenvironment of gastric cancer. Cancer Sci. 2014, 105, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yu, H.; Yan, C.; Mei, Y.; Lin, C.; Hong, Y.; Lin, X.; Zhang, Q.; Yu, J. Plasmacytoid Dendritic Cells and ICOS(+) Regulatory T Cells Predict Poor Prognosis in Gastric Cancer: A Pilot Study. J. Cancer 2019, 10, 6711–6715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurilshikov, A.; Medina-Gomez, C.; Bacigalupe, R.; Radjabzadeh, D.; Wang, J.; Demirkan, A.; Le Roy, C.I.; Garay, J.A.R.; Finnicum, C.T.; Liu, X.; et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 2021, 53, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
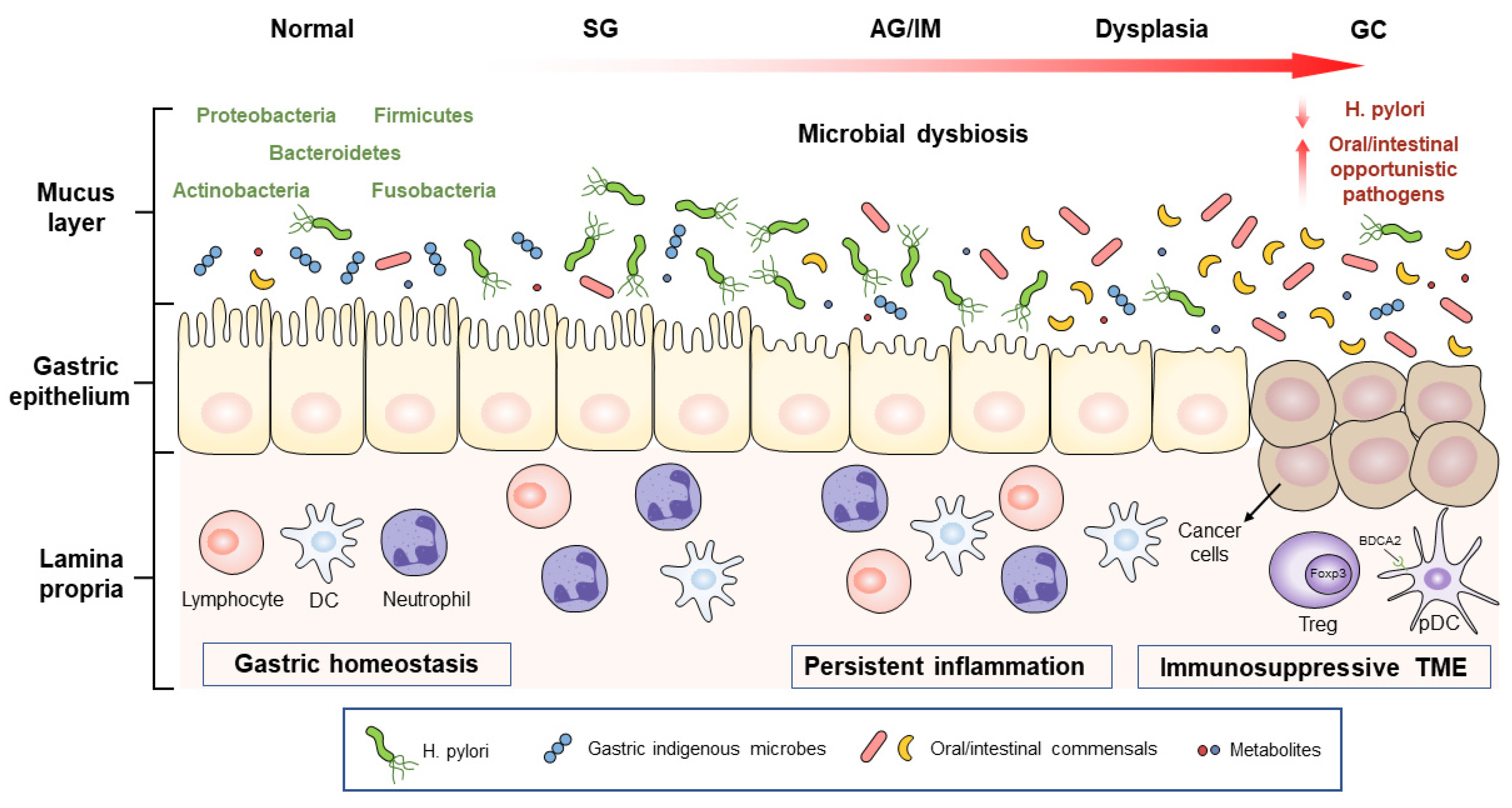
Figure 1.
Schematic representation of the gastric microbial alteration across different stages of gastric carcinogenesis. H. pylori can trigger the initiation of GC, while non-H. pylori microorganisms, especially oral or intestinal opportunistic pathogens, are also involved in GC development. These microbes and their metabolites promote gastric carcinogenesis by inducing persistent inflammatory responses and immunosuppressive microenvironments. SG, superficial gastritis; AG, atrophic gastritis; IM, intestinal metaplasia; GC, gastric cancer; DC, dendritic cell; Treg, regulatory T cell; pDC, plasmacytoid dendritic cell; Foxp3, forkhead box P3; BDCA2, blood DC antigen 2; TME, tumor-associated microenvironment.
Figure 1.
Schematic representation of the gastric microbial alteration across different stages of gastric carcinogenesis. H. pylori can trigger the initiation of GC, while non-H. pylori microorganisms, especially oral or intestinal opportunistic pathogens, are also involved in GC development. These microbes and their metabolites promote gastric carcinogenesis by inducing persistent inflammatory responses and immunosuppressive microenvironments. SG, superficial gastritis; AG, atrophic gastritis; IM, intestinal metaplasia; GC, gastric cancer; DC, dendritic cell; Treg, regulatory T cell; pDC, plasmacytoid dendritic cell; Foxp3, forkhead box P3; BDCA2, blood DC antigen 2; TME, tumor-associated microenvironment.


{kind=link}
Table 1.
Characteristics of sequencing-based studies on human gastric microbiota in gastric carcinogenesis.
Table 1.
Characteristics of sequencing-based studies on human gastric microbiota in gastric carcinogenesis.
| First Author | Country | Sample Size | Sample Type | Sequencing Method | Key Results |
|---|---|---|---|---|---|
| Dicksve [48] (2009) | Sweden | Dyspeptic controls, n = 5; GC, n = 10 | Gastric biopsies | 16S rRNA gene seq | Streptococcus, Lactobacillus, Veillonella, and Prevotella dominated GC microbiota, with relatively low abundance of H. pylori. |
| Aviles-Jimenez [28] (2014) | Mexico | NAG, n = 5; IM, n = 5; GC, n = 5 | Gastric biopsies | 16S rRNA gene microarray | Steady reduction in gastric bacterial diversity observed from NAG, IM, and to intestinal-type GC. |
| Eun [27] (2014) | South Korea | Chronic gastritis, n = 10; IM, n = 10; GC, n = 11 | Gastric biopsies | 16S rRNA gene seq | Depleted Helicobacteraceae and enriched Streptococcaceae in GC compared with chronic gastritis and IM. |
| Jo [51] (2016) | South Korea | H. pylori-negative controls, n = 13; H. pylori-negative GC, n = 19; H. pylori-positive controls, n = 16; H. pylori-positive GC, n = 15 | Gastric biopsies | 16S rRNA gene seq | Enriched nitrosating or nitrate-reducing bacteria other than H. pylori in GC. |
| Castaño-Rodríguez [41] (2017) | Singapore, Malaysia | Functional dyspepsia, n = 20; GC, n = 12 | Gastric biopsies | 16S rRNA gene seq | Increased richness and phylogenetic diversity in GC; Lactococcus, Veilonella, Fusobacterium, and Leptotrichia enriched in GC. |
| Yu [43] (2017) | China, Mexico | GC, n = 80 (China); GC, n = 80 (Mexico) | Surgical resection specimens | 16S rRNA gene seq | H. pylori as the most abundant microbes in nontumor tissue in GC, followed by oral-associated bacteria; Gastric microbiota composition did not differ by H. pylori status or gastric anatomic sites but did differ between paired tumor and nontumor tissues. |
| Coker [26] (2018) | China | Discovery cohort: SG, n = 21; AG, n = 23; IM, n = 17; GC, n = 20; Validation cohort: SG, n = 56; AG, n = 51; GC, n = 19 | Gastric biopsies | 16S rRNA gene seq | Significant dysbiosis in gastric microbiota of IM and GC compared with SG; Oral microbes including P. stomatis, S. anginosus, P. micra, S. exigua, and D. pneumosintes were top enriched in GC and could be used as biomarkers to distinguish GC from SG. |
| Ferreira [25] (2018) | Portugal | Discovery cohort: Chronic gastritis, n = 81; GC, n = 54; Validation cohort: Chronic gastritis, n = 15 GC, n = 23 (Portugal); GC, n = 79 (China); GC, n = 53 (Mexico) | Gastric biopsies | 16S rRNA gene seq | Decreased microbial diversity; depleted Helicobacter; and enriched intestinal commensals, including Phyllobacterium, Achromobacter, Citrobacter, Clostridium, Lactobacillus, and Rhodococcus in GC; Combining the above taxa could excellently discriminate patients with GC from those with chronic gastritis. |
| Hu [45] (2018) | China | SG, n = 5; GC, n = 6 | Gastric wash samples | Shotgun metagenomic seq | Decreased microbial richness in GC; Oral commensals or opportunistic pathogens including Neisseria, Alloprevotella, and Aggregatibacter were the most representative taxa in GC. |
| Hsieh [36] (2018) | Taiwan | Gastritis, n = 9; IM, n = 7; GC, n = 11 | Gastric biopsies | 16S rRNA gene seq | Significantly depleted H. pylori in GC; Clostridium, Fusobacterium, and Lactobacillus enriched in GC. |
| Chen [40] (2019) | China | 62 pairs of GC tumor and adjacent nontumor tissues | Gastric biopsies | 16S rRNA gene seq | Increased microbial richness and diversity in tumor tissues; Oral bacteria, including Peptostreptococcus, Streptococcus, and Fusobacterium enriched in tumor tissues; actic acid-producing bacteria (e.g., L. lactis and L. brevis) in nontumor tissues. |
| Gunathilake [52] (2019) | South Korea | Healthy controls, n = 288; Early GC, n = 268 | Gastric biopsies | 16S rRNA gene seq | Significantly higher risk for GC in patients with high levels of H. pylori, Propionibacterium acnes, and Prevotella copri; L. lactis as a protective factor for GC development. |
| Ling [53] (2019) | China | GC, n = 64 | Surgical resection specimens | 16S rRNA gene seq | Abundances of Stenotrophomonas and Selenomonas positively correlated with pDCs and Tregs, respectively; Abundances of Comamonas and Gaiella negatively correlated with pDCs and Tregs, respectively. |
| Liu [46] (2019) | China | GC, n = 276 | Surgical resection specimens | 16S rRNA gene seq | Bacterial richness reduced in peri-tumoral and tumoral microhabitats; Depleted H. pylori, P. copri, and B. uniformis but enriched P. melaninogenica, S. anginosus, and P. acnes in tumoral microhabitats. |
| Park [54] (2019) | Korea | SG, n = 62 IM, n = 21 GC, n = 55 | Gastric biopsies | 16S rRNA gene seq | Enriched Rhizobiales in patients with H. pylori-negative IM compared to H. pylori-negative SG or GC; Abundances of Rhizobiales and Neisseriaceae gradually increased as H. pylori decreased. |
| Wang [55] (2020) | China | Chronic gastritis, n = 60 Early GC, n = 30 Advanced GC, n = 30 | Gastric biopsies | 16S rRNA gene seq | Distinct microbial community structure and composition between early and advanced GC |
| Wang [39] (2020) | China | Healthy controls, n = 30; NAG, n = 21; IM, n = 27; GIN, n = 25; GC, n = 29 | Gastric biopsies | 16S rRNA gene seq | Progressive decrease in gastric microbial richness and alpha diversity from healthy controls, NAG, IM, intraepithelial neoplasia, and to GC; Nitrospirae, a nitrite-oxidizing phylum, depleted during neoplastic progression. |
| Ravegnini [56] (2020) | Italy | Signet-ring cell carcinoma, n = 10 Adenocarcinoma, n = 10 | Surgical specimens | 16S rRNA gene seq | Enriched Prevotella, Fusobacterium, Actinomyces, Stenotrophomonas, and Roseburia in signet-ring cell carcinoma; Increased Halomonas, Shewanella, Pantoea, Faecalibacterium, and Neoasaia in adenocarcinoma. |
| Gantuya [42] (2020) | Mongolia | Healthy controls, n = 20; Gastritis, n = 20; AG, n = 40; IM, n = 40; GC, n = 48 | Gastric biopsies | 16S rRNA gene seq | Highest bacterial alpha diversity in healthy controls, followed by IM and GC with the least in gastritis and AG; Carnobacterium, Glutamicibacter, Paeniglutamicibacter, Fusobacterium, and Parvimonas associated with GC, regardless of H. pylori infection status. |
| Gunathilake [23] (2021) | Korea | Healthy controls, n = 288; GC, n = 268 | Gastric biopsies | 16S rRNA gene seq | Bacterial richness and evenness negatively correlated with Helicobacter species abundance, while MDI positively correlated with Helicobacter species abundance in both GC and controls; Higher abundance of Actinobacteria species associated with increased risk of GC. |
Note: GC, gastric cancer; NAG, non-atrophic gastritis; AG, atrophic gastritis; IM, intestinal metaplasia; SG, superficial gastritis; pDCs, plasmacytoid dendritic cells; Tregs, regulatory T cells; MDI, microbial dysbiosis index; rRNA, ribosomal RNA.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wen, J.; Lau, H.C.-H.; Peppelenbosch, M.; Yu, J. Gastric Microbiota beyond H. pylori: An Emerging Critical Character in Gastric Carcinogenesis. Biomedicines 2021, 9, 1680. https://doi.org/10.3390/biomedicines9111680
AMA Style
Wen J, Lau HC-H, Peppelenbosch M, Yu J. Gastric Microbiota beyond H. pylori: An Emerging Critical Character in Gastric Carcinogenesis. Biomedicines. 2021; 9(11):1680. https://doi.org/10.3390/biomedicines9111680
Chicago/Turabian StyleWen, Jun, Harry Cheuk-Hay Lau, Maikel Peppelenbosch, and Jun Yu. 2021. "Gastric Microbiota beyond H. pylori: An Emerging Critical Character in Gastric Carcinogenesis" Biomedicines 9, no. 11: 1680. https://doi.org/10.3390/biomedicines9111680
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.