
Regulation of Cytochrome c Oxidase by Natural Compounds Resveratrol, (–)-Epicatechin, and Betaine

College of Medicine, Dankook University, Cheonan-si 31116, Chungcheongnam-do, Korea
Cells 2021, 10(6), 1346; https://doi.org/10.3390/cells10061346
Submission received: 17 March 2021
/
Revised: 15 May 2021
/
Accepted: 17 May 2021
/
Published: 29 May 2021
(This article belongs to the Collection Regulation of Eukaryotic Cytochrome c Oxidase)
Abstract
:Numerous naturally occurring molecules have been studied for their beneficial health effects. Many compounds have received considerable attention for their potential medical uses. Among them, several substances have been found to improve mitochondrial function. This review focuses on resveratrol, (–)-epicatechin, and betaine and summarizes the published data pertaining to their effects on cytochrome c oxidase (COX) which is the terminal enzyme of the mitochondrial electron transport chain and is considered to play an important role in the regulation of mitochondrial respiration. In a variety of experimental model systems, these compounds have been shown to improve mitochondrial biogenesis in addition to increased COX amount and/or its enzymatic activity. Given that they are inexpensive, safe in a wide range of concentrations, and effectively improve mitochondrial and COX function, these compounds could be attractive enough for possible therapeutic or health improvement strategies.
1. Introduction
Mitochondria produce cellular energy, intermediates for biosynthesis, and reactive oxygen species (ROS) and play important roles in the regulation of metabolic homeostasis and cell death. Disruption of their function, dynamics, or biogenesis has been associated with a wide range of human diseases including cancer, diabetes, obesity, and aging [1].
The majority of cellular energy in the form of ATP is produced by mitochondrial oxidative phosphorylation (OXPHOS). This is carried out by the electron transport chain (ETC) complexes, which consists of respiratory chain complexes (complex I, II, III, and IV) and ATP synthase (complex V), which reside in the inner mitochondrial membrane. During electron transfer, complexes I, III, and IV (also known as cytochrome c oxidase; COX) pump protons across the inner mitochondrial membrane and create the mitochondrial membrane potential which is utilized by ATP synthase to produce ATP from ADP and Pi. The OXPHOS complexes are also proposed to play a crucial role in maintaining cristae morphology, folded structures of the inner mitochondrial membrane [2]. In many species and tissues, the ETC complexes form supercomplexes with various stoichiometries, with respirasome (CI1CIII2CIV1) being the most well-known [2]. The significance of these supramolecular structures has been suggested to be functionally relevant to ETC activity by increasing the efficiency of NAD-linked respiration and by reducing ROS production [3]. Furthermore, disruptions in supercomplex organization have been associated with human diseases including congenital mitochondrial disorders, senescence, and other age-related diseases [4].
The efficiency of OXPHOS is controlled by mitochondrial dynamics as well. Although there are tissue and cell type-specific differences, mitochondrial size and shape continuously change by fusion and fission processes dependent on cellular bioenergetic requirements. These dynamic events also facilitate the maintenance of mitochondrial homeostasis and quality control [5]. Fused mitochondrial networks found in energetically demanding conditions allow mitochondria to mix their components, including mitochondrial DNA and OXPHOS complexes, to maintain efficient bioenergetic capacity [6,7]. In contrast, the fission process induces fragmentation of mitochondria that are often present in resting cells with low energetic demand and participate in the removal of dysfunctional ones.
Cytochrome c oxidase (COX, complex IV), the last step of the ETC receives electrons from cytochrome c and irreversibly reduces oxygen to water. Mammalian COX is dimeric and contains 13 tightly bound subunits as confirmed by X-ray crystallography [8]. Subunits, I, II, and III are encoded by the mitochondrial genome and among them the biggest two, subunit I and II fulfill the catalytic reaction of COX. The remaining 10 subunits, IV, Va, Vb, VIa, VIb, VIc, VIIa, VIIb, VIIc, and VIII are encoded by the nuclear genome and regulate the enzyme [9]. Therefore, the biogenesis of COX is indeed a finely concerted and synchronized process of both genomes [10]. Recently, NDUFA4, which had originally been considered as a subunit of complex I, was suggested to be a loosely bound subunit of COX [11]. Previously, this subunit had not been detected in a holoenzyme by X-ray crystallography and SDS-PAGE, since it is lost during the traditional COX purification process.
Considering the significance of the crucial role that COX plays in aerobic energy metabolism by controlling mitochondrial respiration, COX must be tightly regulated. Indeed, several regulatory mechanisms are known, such as an allosteric regulation, expression of tissue-/species-/development-specific isoforms, reversible phosphorylation modification via cell signaling, protein-protein interactions, and supercomplex formation [9,12]. These regulatory mechanisms affect the enzymatic activity of COX, which in turn alters mitochondrial membrane potential, and therefore ATP and ROS production. It has been reported that COX dysfunction is associated with many diseases where energy and ROS production are found to be dysregulated [13].
Many natural compounds that plants synthesize to reduce damage and increase their survival in response to environmental stresses, such as UV light, malnutrition, dehydration, pathogenic infection, and predators, are known to have anti-oxidative and anti-inflammatory properties [14]. Their biological activities are found to be beneficial for human health by diminishing or preventing pathological conditions that are associated with oxidative stress. Interestingly, a good number of these compounds affect mitochondrial function, including OXPHOS, by regulating mitochondrial biogenesis, via allosteric control, and possibly cell signaling [15,16,17,18]. Among them several compounds, such as resveratrol (RSV), (–)-epicatechin (EPI), betaine (BET), curcumin [19,20,21,22,23,24], and quercetin [25,26,27,28,29], have been shown to have an effect on COX. It has been reported that they increase protein amount and enzymatic activity of COX in various cell types and more pronounced effects were obtained under challenged or stressed conditions. Targeting and modulating COX by these compounds may therefore serve as a potential therapeutic approach since it could counteract COX dysfunction, which is observed in many pathological conditions. In this report, the most in-depth studied compounds, RSV, EPI, and BET, that affect COX are reviewed.
2. Resveratrol
Resveratrol (3,4’,5-trihydroxystilbene, RSV, Figure 1A) is a polyphenolic phytoalexin and synthesized in plants as a defensive mechanism responsive to UV light, microbial infection, pesticides, and others [30]. Its synthesis starts from a phenylpropanoid pathway, in which phenylalanine or tyrosine are converted into p-coumaroyl CoA, which is in turn condensed with malonyl CoA by stilbene synthase, producing RSV [31]. It was first isolated from Veratrum album, a type of grandiflorum, and characterized in 1939 [32]. It is found in various edible plants, such as peanuts, grape skins, berries, cacao, and others [33]. When consumed orally, it is converted into resveratrol sulfate and resveratrol glucuronide conjugates [34,35].
Initially, RSV drew attention for its possible role in the “French paradox” phenomenon, the epidemiological observation that French people have a lower incidence of coronary heart disease despite a diet rich in saturated fats [36], and its beneficial effects have been demonstrated to include anti-oxidative, anti-inflammatory, anti-apoptotic, and anti-cancer properties [37,38,39,40,41]. Due to the nature of its structure, RSV efficiently scavenges ROS and suppresses lipid peroxidation, and in addition, it exerts the anti-oxidative action indirectly by increasing expression of ROS scavenging enzymes [15]. There have been a number of studies showing that RSV enhances muscle performance [42,43,44,45] and alleviates or even prevents cardiovascular disease, neurodegenerative disease, cancer, and metabolic defects [46,47,48,49]. The compound is generally considered non-toxic, excluding mild adverse effects such as nausea, stomach pain, bloating, and diarrhea at high dosages (>1000 mg/day) [50]. RSV has been applied widely as a dietary supplement and active ingredient in cosmetic and dermatological products [51].
One of the molecular targets of RSV responsible for its protective properties is sirtuin 1 (SIRT1), a class III NAD+-dependent histone deacetylase in the nucleus. SIRT1 regulates several transcription factors and co-regulators for numerous physiological processes including glucose and fat metabolism, insulin production, and cell survival [52,53]. The intimate relationship between SIRT1 and mitochondria is possible through the action of a transcription coactivator, peroxisome proliferator activated receptor gamma coactivator 1 alpha (PGC1α) [54]. PGC1α is known to play a crucial role in mitochondrial energy metabolism and biogenesis by regulating transcription factors, such as NRF1/2, ERRα/β/γ, PPARα/γ/δ, FOXO1, and others [55,56]. It has been shown that the effects of RSV administration [57,58] are similar to those of caloric restriction in that both regimens share the same upstream regulating mechanism of SIRT1 activation and induction of PGC1α [59]. Another important regulator of PGC1α is AMP activated protein kinase (AMPK), a key metabolic energy sensor and regulator [60]. Interestingly, AMPK and SIRT1 are mutual regulators of each other, and they both play pivotal roles in regulating glucose homeostasis and mitochondrial biogenesis in response to stress and nutrient status [61,62,63]. The mediators of mitochondrial biogenesis induced by RSV are reviewed in detail in [15].
Enhanced mitochondrial biogenesis by RSV through SIRT1/PGC1α was consistently and reproducibly observed in liver, skeletal muscle, brown adipose tissue, brain, and other tissues [53,57,64,65,66]. Lagouge et al. showed that RSV (400 mg/kg per day for 15 weeks) induced increased gene expression of components that are involved in mitochondrial biogenesis [57]. The increase in mRNA levels of COX subunits (COX IV-1, COX Va, COX Vb, COX VIIaL) were confirmed along with those of other OXPHOS complex genes, such as NDUFB8 and ATP5G3 (Table 1). This enhancement could account for the increased muscle function seen in running endurance tests of high fat diet-fed mice with RSV administration.
A moderate dose of RSV in high fat diet-fed mouse skeletal muscle and hepatocytes (25–30 mg/kg per day for 8 months) and in C2C12 cells (25 μM for 24 h) increased AMPK activation, mitochondrial biogenesis, and mitochondrial function in a SIRT1 dependent manner [62]. In this study, mRNA levels of PGC1α, NRF, and subunits of OXPHOS complexes (NDUFS8, SDHB, UQCRC1, COX Vb, ATP5A1), ATP levels, mitochondrial mass, mitochondrial respiration, and membrane potential were all improved by RSV administration. The enhanced mitochondrial biogenesis by RSV was observed in nerve cells as well. A study of neurobehaviorally deficient rats found that intragastric administration of RSV (30 mg/kg per day) ameliorated disrupted mitochondrial features, such as reduced mitochondrial biogenesis, membrane potential, and ATP content, and elevated ROS level in hippocampal cells [71]. In a study of rats with status epilepticus, 100 μmol RSV microinjection into the hippocampus augmented mitochondrial content and protected the cells from apoptosis [67]. The increase in mitochondria after RSV injection was mediated by PGC1α-NRF1-mitochondrial transcription factor TFAM, and the levels of COX subunit I and mitochondrial DNA increased as a result of enhanced mitochondrial biogenesis. Another study with aged rats showed the neuroprotective effect of RSV alone and in combination with soy isoflavones [68]. RSV (80 mg/kg per day for 12 weeks) relieved the aberrant features of hippocampal mitochondria, such as swelling and vacuolization, and decreased apoptosis. Both protein levels of COX subunit I and antioxidant enzyme activity were restored following RSV administration.
The upregulation of COX might be explained through the regulation of COX assembly factors. Indeed, the expression of COX18, which is required for the translocation of the C-terminus of COX subunit II across the mitochondrial inner membrane [72], was reported to be increased by RSV [73]. RSV administration (500 mg per day) together with a 12-week exercise regimen produced a synergistic improvement of muscle capacity (i.e., improved oxygen uptake and muscle fatigue resistance) and mitochondrial density in 65–80-year-old men and women. In this group, gene expression levels of COX18 were increased three-fold, along with other mitochondrial metabolic genes following RSV treatment combined with exercise.
While the majority of literature demonstrates RSV’s effect on OXPHOS via mitochondrial biogenesis, another regulatory mechanism of RSV has been proposed. It has been suggested that RSV binds directly to complex I [74], III [75], and V [76,77,78] and alters their activities, yet no study has thus far shown that COX is a target of RSV. One to five μM RSV stimulated the activity of isolated complex I when co-incubated, which in turn increased the NAD+/NADH ratio [74]. In this study, enzymatic activities of complex I, II, and III of HepG2 cells were enhanced by RSV even though overall amounts of mitochondrial proteins did not change. In rat brain mitochondria, high concentrations of RSV (100 μM) competed with coenzyme Q for the substrate binding site on complex III and consequently suppressed the enzymatic activity by ~20% [75]. Another target affected by RSV is F0F1-ATPase (complex V). Incubation of pico to nano molar concentrations of RSV slightly activated F0F1-ATPase activity by ~10% in rat liver mitochondria but not in heart mitochondria [77], while RSV at higher concentrations (>1 μM) inhibited its enzymatic activity in rat brain and liver [77,78].
Free radical scavenging and anti-oxidative activities are well-known properties of RSV [79] and suggested to protect mitochondrial ETC complexes from oxidative stress. Incubation of rat brain homogenates with metabolites citrulline or ammonia, which were employed to mimic a urea cycle disorder, citrullinemia type 1, induced the deterioration of mitochondrial energy metabolism by downregulating complex II and COX [69]. The suppressed activities of these two enzymes were recovered by co-incubation with sub-millimolar concentrations of RSV for 1 h. Regarding the underlying mechanism, the authors proposed that anti-oxidative action of RSV protected the ETC complexes from ROS produced by the citrulline or ammonia treatment.
Conversely, some literature suggests that RSV may induce ROS production [80]. RSV has been proposed to inhibit cell proliferation, growth, angiogenesis, and cancer metastasis of several cancers, such as breast, lung, glioma, and prostate [81,82,83,84] and to potentiate the efficacy of chemotherapeutic agents [85,86,87,88]. The cell damage and eventual cell death by ROS-induced damage via modulating ETC complexes might be an anti-cancer mechanism of RSV. RSV-induced cell death mediated by elevated superoxide production was observed in CEM, a lymphoblastic leukemia cell line [70]. The cytotoxic effect of RSV in the leukemia cells was blunted by overexpression of Bcl-2 (CEM/Bcl-2), accompanied by increased COX activity and mitochondrial respiration. The elevated COX activity and mitochondrial respiration in the CEM/Bcl-2 declined after RSV incubation for 6 h in a dose-dependent manner up to 50 μM and was not due to the changes in COX amounts. However, no effect of RSV on COX was observed in control CEM cells. The apoptotic effect of RSV on SW620 colon cancer cells seemed to be triggered by increased ROS production due to elevated mitochondrial respiration [41]. Along with elevated oxygen consumption and mitochondrial metabolic activity, COX activity was increased by ~45% and ATP content was increased by ~35% after 10 μM RSV treatment for 48 h compared to control. This enhanced mitochondrial function might be due to the increased mitochondrial content, and mitochondrial mass was indeed increased. Moreover, protein levels of all OXPHOS complexes except complex III were increased, and among them, COX was the most affected by RSV treatment. However, unlike the studies discussed above, not SIRT1, but SIRT3, which resides inside the mitochondria, was associated with the RSV effect, accompanied by elevation of transcription factors for mitochondrial biogenesis, such as PGC1α, NRF1, TFAM [41]. Intriguingly, a recent study suggested that COX is a novel target of SIRT3, which led to COX activation in rat brain and K13, K264, K319, and K481 on COX subunit I were identified as deacetylation sites for SIRT3 [89].
The initial idea of using RSV to extend lifespan originated from studies with Saccharomyces cerevisiae, mediated by stimulating silent information regulator (Sir) [90]. Since then, RSV has been extensively investigated for the use in higher organisms including humans. RSV successfully prolonged life spans of unicellular organisms and some animals such as Caenorhabditis elegans [91,92,93] and Drosophila melanogaster [91,94]. However, its efficacy in mammals turned out to be virtually insignificant [95,96,97]. Rather than being a longevity wonder drug, RSV has been explored for its beneficial effects on human disease models [95]. Based on its functions regulating mitochondria and ROS, RSV has been proposed to be a possible treatment regimen for OXPHOS deficiency alone or in combination with other therapeutic substances. Papepe and Coster listed the studies evaluating RSV treatment in cultured skin fibroblast from patients with OXPHOS defects that are associated with genetic mutations [98]. The remedial values of RSV on COX deficiency caused by the mutation of COX assembly factors such as SURF1 and COX10 varied from positive to no effect [99,100,101]. It seems that RSV may be useful to treat certain defects and conditions but not others.
3. (–)-Epicatechin
(–)-epicatechin ((2R,3R)-2-(3,4-dihydroxyphenyl)-3,4-dihydro-2H-1-benzopyran-3,5,7-triol, EPI), a monomeric flavan-3-ol is a non-glycosylated flavonoid, and is found in rich quantities in cacao, tea, peanuts, apples, grapes, and berries [102]. It is suggested that EPI provides plants protection from microbial pathogens and predators by causing toxicity and interference in digestive enzymes. EPI is synthesized by phenylpropanoid and flavonoid biosynthetic pathways. It is produced by anthocyanin synthase and anthocyanin reductase from leucoanthocyanidin, which is derived from flavanone through a series of enzyme-catalyzed reactions [31]. EPI is one of four diastereoisomers of catechin that contains one dihydropyran and two benzene rings with five hydroxyl groups (Figure 1B). Due to its phenolic hydroxyl groups, EPI can efficiently scavenge free radicals [103]. It has the highest bioavailability in plasma and urine among other catechins [104], and is metabolized by conjugation with glucuronides, sulfates, and O-methyl sulfates once ingested [105].
EPI has been reported to guard cells from oxidative stress by several means [106]. Anti-oxidative efficacy of EPI is mediated by direct interaction with and removal of ROS and reactive metal ions. As an indirect anti-oxidant, EPI upregulates anti-oxidant enzymes, downregulates pro-oxidant enzymes, and produces phase II detoxifying/anti-oxidant enzymes. In addition, it suppresses inflammation by modulating oxidative stress-related cell signaling pathways [106]. Because of its anti-oxidative properties, EPI has been proposed to prevent or even rescue from diseases which are caused by or associated with oxidative stress, such as cardiovascular disease, neurodegenerative disease, diabetes, obesity, cancer, and others [107,108,109,110,111].
The fact that Kuna Indians in the San Blas Islands of Panama, who consume lots of cocoa drinks, have low incidence rates of hypertension and cardiovascular disease [112] evoked interest in the protective effect of EPI, since it is the most abundant polyphenolic monomer in cacao [113]. The administration of EPI rich cocoa has been suggested to improve the gastrointestinal system, nervous system, muscle performance, plasma lipid profiles, glucose homeostasis, obesity, and insulin sensitivity [114,115,116,117,118,119,120,121,122,123,124]. Many of these advantageous effects have been validated by the administration of pure EPI compound in humans and rodents [125,126,127,128,129,130,131,132,133,134,135,136,137,138].
Mounting evidence suggests that these beneficial effects of EPI might be facilitated by enhanced mitochondrial mass, integrity, and function (extensively reviewed in [125]). Several studies have reported the enhanced mitochondrial biogenesis in response to EPI was mediated by SIRT1-PGC1α [138,139,140] and nitric oxide [138,140,141], and considerable changes in protein levels and function of OXPHOS complexes have been reported in animals and cultured cells [125]. Direct modulation of mitochondria by EPI has been suggested to increase their function and integrity. EPI treatment of isolated rat heart mitochondria stimulated mitochondrial state 2 respiration in a dose-dependent manner and suppressed cytochrome c release, indicating stabilized integrity of mitochondrial membranes [142]. Its positive effect on membranes was further supported by an increase in cristae formation in skeletal muscle of patients with type 2 diabetes and heart failure [140] and with Becker muscular dystrophy [143], and in mouse heart and muscle [132].
EPI effects on OXPHOS complexes, particularly with regard to COX, have been investigated most extensively in muscle cells [144,145,146,147,148], along with endothelial cells [129], β cells [149], fat cells [150], and some cancer cells [151] (Table 2).
Moreno-Ulloa et al. showed increased mitochondrial biogenesis by 10 μM EPI treatment for 48 h with increased TFAM, NRF2, and porin levels, and citrate synthase activity in differentiated C2C12 cells [144]. In accordance with the increased mitochondrial mass, COX subunit I levels increased as well, while no significant change in complex II was reported. A possible contribution of a G-protein coupled estrogen receptor as an EPI receptor on the plasma membrane in skeletal muscle was proposed for mitochondrial biogenesis since the effect of EPI was blocked by inhibition of the receptor. Several rodent studies revealed that EPI has a positive effect on exercise performance, mitochondrial biogenesis, and function in skeletal muscles similar to the results of an exercise regimen. EPI administration (1.0 mg/kg, twice a day) stimulated mitochondrial biogenesis in mouse skeletal muscle, and its combination with 8 weeks of running exercise brought out cumulative results [148]. The enhanced exercise performance by EPI might be explained by the increased angiogenesis and mitochondrial biogenesis within the muscle. Increased capillary development was accompanied by the upregulation of pro-angiogenic factor VEGFR2 and downregulation of anti-angiogenic factors, such as ADAMTS1 and TSP1, along with suppressed FOXO1 levels. Expression levels of mitochondrial biogenesis regulators PGC1β and TFAM were upregulated and concurrently citrate synthase activity, which is often used as an indicator for mitochondrial content, was increased by EPI administration. Not only does EPI reinforce exercise performance by increased mitochondrial biogenesis and angiogenesis, it can also attenuate the loss of exercise benefits due to detraining [145]. Enhanced features in mouse hind muscles from 5 weeks of endurance training included increased running distance and time, capillarity, amount of complex V and COX, and COX activity. These adaptations reverted to the pre-exercise state 14 days post exercise cessation; however, EPI intake (1.0 mg/kg, twice a day) attenuated the regression and retained the adaptations to an almost similar extent as the exercised state. Among OXPHOS complexes, COX was the one most influenced by EPI with increases in the amount and activity of COX by training (~35% and ~144%, respectively) and those maintained by EPI administration during detraining (~45% and ~108%, respectively) compared to control. A similar beneficial effect of EPI on skeletal muscles against exercise cessation occurred in muscle atrophy induced by hindlimb suspension [146]. EPI administration (1 mg/kg, twice a day for 14 days) diminished the muscle degeneration caused by hindlimb suspension and improved the angiogenesis and mitochondrial biogenesis along with COX amount.
In contrast to the above studies, a clinical trial assessing the effect of EPI on the aerobic training adaptation in heathy human subjects turned out to be disappointing [133]. EPI supplementation (100 mg, twice a day) during 4 weeks of cycle training decreased the protein levels of ETC complex II, while those of other mitochondrial proteins, such as citrate synthase and cytochrome c were not altered in the skeletal muscle. Similar findings were obtained with EPI-rich cocoa products, where the supplementation did not improve exercise performance [152,153,154]. It seems that protective effects of EPI might be more pronounced under stressed conditions, as shown in H2O2 injured C2C12 cells [149]. Considering its properties, which improve exercise capacity mediated by upregulated muscular vascularization and mitochondrial biogenesis, EPI might be useful as a complement for medical conditions causing muscle weakness [147]. Upregulated capillarity by EPI administration (1 mg/kg, twice a day for 30 days) was accompanied by increased VEGF-A and decreased TSP1 levels in skeletal muscle of rats with congenital low running capacity. Along with the enhanced angiogenesis, myogenesis was also improved by EPI. The P38 MAPK signaling pathway, an important player of skeletal muscle differentiation, and MEF2A, a transcription factor for regulating muscle gene expression, were upregulated by EPI treatment. Increased mitochondrial biogenesis, mitochondrial volume, cristae abundancy, and COX were found together with increased expression levels of PGC1α, PGC1β, and TFAM. Interestingly, these beneficial effects lasted 15 days after EPI was discontinued.
Mitochondrial dysfunction has been proposed as a causative factor for the dysfunction of β cells and insulin resistance in type 2 diabetes [155]. Other evidence for the regulation of mitochondrial function by EPI was verified by the study with INS-1 832/13 rat β cells [149]. Glucose-stimulated insulin secretion from β cells was accompanied by upregulated protein levels of OXPHOS complexes including COX, and the consequently increased mitochondrial respiration following 10 μM EPI treatment for 24 h. The increased amounts of OXPHOS complexes by EPI were observed in adipocytes as well. The levels of complex II subunit A, complex V (subunit not specified in the paper), and COX subunit I were remarkably increased by 100 nM EPI treatment for 72 h. Enhanced mitochondrial biogenesis by EPI was confirmed by increased mitochondrial volume and its related regulators, such as SIRT1/3, PGC1α, NRF1/2, and TFAM [150]. Furthermore, EPI was able to reverse aging related changes in endothelial cells. Increased activity and amount of senescent biomarker β-galactosidase and decreased vasodilator nitric oxide and SIRT1 in aged cow endothelial cells were reversed by EPI treatment [129]. Declined mitochondrial biogenesis, along with downregulated ETC complex II, complex V, and COX along with TFAM and mitofilin in aged cells, were recovered to close to the levels in young control cells following incubation with 1 μM EPI for 48 h. Not only molecular profiles, but also physiological function of the endothelium was improved by EPI, since systolic blood pressure decreased due to increased vasodilation in aged rat aorta after EPI administration (1 mg/kg per day for 15 days).
The beneficial effects of EPI on mitochondrial function have also been used to sensitize cancer cells to radiotherapy using cultured cells [151]. After a 20 μM EPI treatment for 1 h, human pancreatic cancer cell lines (Panc-1, MIA PaCa-2) and a glioblastoma cell line (U87) were more sensitive to radiation and cell death. These changes were not observed in a normal fibroblast cell line (HNF). The cytotoxic effect on cancer cells was further confirmed by increased p21 production and Chk2 phosphorylation in Panc-1, but not in HNF; therefore, the sensitization by EPI was selective for cancer cells. This sensitization may be the result of enhanced mitochondrial function, since COX activity in Panc-1 but not in HNF cells was increased by EPI in a dose-dependent manner.
Notably, EPI modulates COX activity not only by increasing its biogenesis, but also by a more rapid process, likely through cell signaling and post-translational modifications, which has not been yet studied. For example, COX activity of healthy rat plantaris muscle was increased by incubation with 20 μM EPI for 25 min, which is insufficient time for the biosynthesis of COX, therefore another regulatory mechanism for COX and triggered by EPI must exist [147]. Interestingly, COX activity in rats with congenitally low running capacity did not respond to the short EPI treatment, which might be due to a faulty signaling pathway system in these animals. Increased EPI-mediated COX activity by mechanisms other than biogenesis was reproducibly shown in healthy mouse quadriceps under the same EPI treatment conditions [148]. A short-term effect on COX was also observed with a different type of catechin, epigallocatechin-3-gallate (EGCG) [156]. EGCG treatment increased COX activity, the mitochondrial membrane potential, mitochondrial respiration, and ATP content in primary human astrocyte and neuronal cell cultures. The effect of EGCG was not mediated by increased mitochondrial biogenesis since no change in the mRNA levels of mitofusin 2, TFAM, and PGC1α was observed. Further studies are needed to confirm the mechanism of short-term regulation by catechins on COX.
4. Betaine
Betaine (glycine betaine, N,N,N-trimethylglycine, BET, Figure 1C), a trimethyl derivative of the amino acid glycine, is a natural compound that is found ubiquitously in animals, plants, and microorganisms [157]. It is known that accumulation of BET improves the growth and survival of plants by protecting cellular components from abiotic stress such as drought, high salinity, and cold. BET is synthesized by betaine aldehyde dehydrogenase from betaine aldehyde, which is a derivative of choline [158]. It was first discovered in the nineteenth century as a byproduct of sucrose production from sugar beets (Beta vulgaris) [159]. The pool of BET in the body is mediated by synthesis through choline oxidation in hepatic and renal mitochondria as well as dietary intake [160], and is present mainly in the kidneys, liver, and brain [161]. Several studies have shown that the dietary intake of BET ranges from 100 to 400 mg/day, and the resting plasma BET concentration reaches 0.02–0.07 mmol/L in humans [162,163]. This compound is chemically stable and nontoxic, even at high concentrations, which broadens the potential for therapeutic uses.
The zwitterionic quaternary ammonium [(CH3)3N+ CH2COO–] form allows BET to exert several physiological functions. The zwitterionic characteristics and high solubility in water allow BET to function as an osmolyte. Concentrations can go as high as the millimolar ranges with no detrimental effect on the cells [159]. Additionally, it functions as a methyl group donor in several metabolic pathways. For example, in the methionine cycle BET donates a methyl group to homocysteine [160]. During such processes, BET converts into N,N-dimethylglycine and is then transformed into the amino acid glycine through a series of biochemical steps. Unlike polyphenols such as RSV and EPI, which can interact and neutralize free radicals directly and also modulate anti-/pro-oxidative enzymes, the anti-oxidative property of BET results from metabolites of the methionine cycle, such as methionine and S-adenosylmethionine (SAM), which play roles as anti-oxidants [164]. Additionally, it functions as a chaperone to protect proteins and DNA from denaturation under high concentrations of urea and NaCl in the medulla of the kidney [159]. In cells, BET does not bind to but is excluded from the hydration shell of a protein. This causes a thermodynamic force to fold the protein more compactly in order to decrease the amount of excluded water, due to the so-called osmophobic effect; therefore, BET is able to stabilize the protein’s native form.
The remarkable ability of BET to protect cells from exogenous stress factors, such as drought, high salinity, and high temperature has been extensively studied from microbial cells to mammals. Because of its low cost, safety, and effectiveness over a wide range of doses, BET has been frequently applied in animal husbandry on pigs, poultry, and lambs [165]. Supplementation of BET has positive effects on growth and feed efficiency in pigs and on egg production and body gain in chickens and turkeys [166,167,168,169,170], and these beneficial effects have been further demonstrated under heat stress conditions [171,172]. In addition, it has been fed to young salmon in fish farms to maintain osmotic balance and stimulate growth when they migrate to the sea [173]. As with animals, BET is beneficial for plant life. It improves growth, photosynthesis, nutrient uptake, and protects cells from chemical toxicity by reducing oxidative stress damage and excessive heavy metal uptake [174]. A positive role for BET in cold acclimation in plants has also been reported [175,176]. Not only multicellular organisms, but also unicellular bacteria benefit from BET. Microbial strains used for fermentation of lactate, ethanol, lysine, pyruvate, and vitamin B12 have been reported to derive benefit from BET, which not only provides a methyl group during biosynthetic processes, but also protects the cells against abiotic stress [177,178,179,180]. Unlike the studies with livestock, the benefits of BET in human subjects seem to be limited (see review on clinical trials in [181]). BET supplementation did not alter either BMI or body weight, but reduced total body fat mass and body fat percentage.
BET has been applied for therapeutic use in humans. Currently, it is used to treat homocystinuria, a systemic disorder with vascular complications and neurodegeneration [182,183] that is caused by a rare genetic disorder of methionine metabolism or deficiency of vitamin B6, B12, or folate [184]. Another possible application for BET was suggested for the treatment of nonalcoholic fatty liver diseases (hepatic steatosis, nonalcoholic hepatitis, and liver fibrosis) and alcoholic liver disease based on studies with rodents [185,186]. In animal models, these conditions share several histological and functional disturbances such as triglyceride accumulation, hyper homocysteinemia, dysregulation of lipid metabolism, insulin resistance, ER stress, mitochondrial dysfunction, and others [187,188] and often co-exist in clinical practice [189]. Multiple studies indicate that BET administration not only arrests but also in some cases reverses the progression to liver dysfunction [159,190,191,192]. Indeed, fruits of Lycium barbarum, a shrub native to China, which contain high amounts of BET has been traditionally used for the treatment of liver diseases in Asian countries [193]. However, the benefit of BET on nonalcoholic steatohepatitis in humans is inconclusive due to limitations with the clinical trial study design (see reviews on clinical trials in [194,195]). Other studies evaluating the effect of BET on cancer, diabetes, obesity, and Alzheimer’s disease revealed that BET consumption could lower the risk of those diseases [157,196].
Other useful applications of BET are its use as an additive in polymerase chain reaction (PCR) [197], cryopreservation [198], and personal care products such as toothpaste and moisturizers [182].
BET has been suggested to have effects on mitochondrial function. Endurance performance and resistance-based exercises such as running, squat repetitions, bench press throw, and vertical jumps, which are metabolically demanding tasks, were improved after BET administration [199,200,201,202]. Although the exact underlying mechanism was not investigated in those studies, one can speculate that the improved performance may be the result of enhanced mitochondrial energy metabolism.
Not much is known about a molecular target of BET for mitogenesis. Although PGC1α was upregulated by BET in db/db mice and rats with alcohol treatment, its upstream regulator and effect on mitogenesis were not evaluated in the studies [186,203,204]. Recently it was suggested that enhanced mitogenesis by BET is mediated by the SIRT1-PGC1α pathway [205]. Increased mitochondrial protein amount and ATP content by BET treatment were accompanied by increased mRNA and protein levels of SIRT1, PGC1α, NRF1, TFAM, and phosphorylated AMPK in C2C12 cells.
Several studies indicate that BET increases activity of mitochondrial ETC complexes, especially under heat stress. As discussed above, BET feeding was proven to be useful in animal husbandry, especially at high ambient temperatures, which normally have a deleterious impact on the activities of ETC complex I, III, and COX [206,207,208]. Chickens housed at over 32 °C presented with inhibited activities of complexes I + III and COX in liver [206] and those of complex I, III, and COX in skeletal muscle [207,208]. BET supplementation rescued the declined complex I activity caused by thermal stress [208]. In this study, BET also upregulated complex I and III activity in the non-thermal stress control group; however, complex II and COX were not affected. Hepatoprotection from chronic alcoholic damage by BET administration (1% w/v added to the diet for 4–5 weeks) in rats was accompanied with increased amounts of OXPHOS complexes [209]. Significantly increased steatosis and loss of intact complexes I, III, V, and COX, but not complex II, caused by ethanol treatment was rescued by BET. However, BET supplementation in control animals showed no effect (Table 3). Increased amounts of COX holoenzyme and of COX subunit I by BET were confirmed by one- and two-dimensional blue native gels and Western blot analysis, respectively. These results suggest that the protective effect of BET on liver from deleterious alcohol exposure may operate through maintaining the ability of mitochondrial OXPHOS.
BET has been proposed to protect cells from other stressed conditions as well. Ganesan et al. showed heart ETC complexes could be beneficiaries of BET treatment when the cells were under stress [210]. Isoprenaline-induced myocardial infarction in rats presented with decreased energy levels and activities of complexes I, II, and COX. Interestingly, pre-administration of BET (250 mg/kg per day for 30 days) was able to prevent the deleterious action of isoprenaline and rescued the ATP levels and the activities of complexes I, II, and COX to near control levels. BET treatment in control animals also showed a tendency towards increased ATP levels and ETC complexes activities, although they were not statistically significant. Reduced OXPHOS gene expression, mitochondrial DNA copy number, and TFAM amount in chicken liver with corticosterone treatment were improved by in ovo injection of BET [211]. Downregulated mRNA levels of COX I, II, III along with subunits of complexes I, III, V, which are encoded by the mitochondrial genome, were improved by BET, as well as COX activity. However, the expression level of the COX IV gene, which is encoded by the nuclear genome was not affected by either corticosterone or EPI.
More evidence for COX as a target of BET was suggested in mouse hepatocyte cell line H2.35 [212]. COX activity was increased by BET treatment for 30 min in a dose-dependent manner up to 2 mM, which resulted in elevated mitochondrial respiration, mitochondrial membrane potential, and cellular energy ATP. However, BET is not an allosteric regulator of COX, since BET treatment of purified COX did not change the enzyme activity. The beneficial effects of BET on mitochondrial function and energy metabolism were proposed to be through a mechanism similar to that mediating its cytoprotective properties. Based on the finding that the AKT signaling pathway was activated by BET in human muscle [213] and that activated AKT in mitochondria induced an increase in COX activity in hypoxic cardiomyocytes [214], one could speculate that the AKT signaling pathway targets COX and increases COX activity, possibly through alterations in post-translational modifications.
5. Conclusions
This review summarized various studies validating the regulatory effects of the natural compounds derived from plants, RSV, EPI, and BET on COX. In conclusion, these three small molecules promote COX biosynthesis and activity.
Although not all compounds share commonalities in their structures—RSV and EPI belong to the polyphenols, while BET is a methyl derivative of glycine—they have similar bioactivity, such as anti-oxidative, anti-inflammatory, and anti-cancer properties. Many studies with cultured cells and lab animals have suggested that the administration of these compounds improves the physiological features and function of cells, organs, and organisms. However, more pronounced effects were obtained under challenged or stressed conditions. In many experimental systems, they were capable of mitigating, preventing, or even rescuing pathological conditions including cardiovascular disease, neurodegenerative disease, metabolic disease, cancer, and aging. They could ameliorate the impaired conditions by alleviating oxidative stress due to direct and indirect antioxidant effects and in addition, by enhancing mitochondrial function and biogenesis, which are often disrupted in pathological conditions.
It has been shown that the increased mitochondrial biogenesis was accompanied by upregulation of biogenesis-related regulators, such as SIRT1, APMK, PGC1α, NRF1/2, and TFAM. In most cases, they increased the amount of COX and/or its enzymatic activity that result in improved mitochondrial function, as seen in increased mitochondrial respiration, mitochondrial membrane potential, and ATP levels (Figure 2). Although the studies did not investigate how crucial the changes in COX were for ameliorating pathological conditions, it is assumed that the improved mitochondrial function via enhanced COX levels and activity by the compounds might contribute to improving those conditions.
Interestingly, their stimulating effect on COX was not limited to protection from oxidative stress and the biogenesis. At least EPI and BET evidently increased COX activity in response to short term treatments. This enhancement was not facilitated by the increased protein levels but possibly by changes in post-translational modification, such as (de)phosphorylation of COX. In fact, several phosphorylation sites of COX have been identified and some of their effects on COX enzymatic activity have been verified. Although it is found that COX is regulated by the compounds, little is known about their regulatory mechanism on COX besides biogenesis and possibly post-translational modifications, and therefore further studies are necessary to investigate whether other regulatory mechanisms may exist.
Funding
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2014R1A1A2058722).
Acknowledgments
The author thanks Maik Hüttemann and Paul T. Morse for comments on the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
| RSV | resveratrol |
| EPI | (–)-epicatechin |
| BET | betaine |
| COX | cytochrome c oxidase |
| ETC | electron transport chain |
| OXPHOS | oxidative phosphorylation |
| ROS | reactive oxygen species |
References
- Cheng, Z.; Ristow, M. Mitochondria and metabolic homeostasis. Antioxid. Redox Signal. 2013, 19, 240–242. [Google Scholar] [CrossRef]
- Wu, M.; Gu, J.; Zong, S.; Guo, R.; Liu, T.; Yang, M. Research journey of respirasome. Protein Cell 2020, 11, 318–338. [Google Scholar] [CrossRef] [Green Version]
- Genova, M.L.; Lenaz, G. Functional role of mitochondrial respiratory supercomplexes. Biochim. Biophys. Acta 2014, 1837, 427–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milenkovic, D.; Blaza, J.N.; Larsson, N.G.; Hirst, J. The enigma of the respiratory chain supercomplex. Cell Metab. 2017, 25, 765–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, H.M.; Williams, J.A.; Ding, W.X. Mitochondrial dynamics and mitochondrial quality control. Redox Biol. 2015, 4, 6–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westermann, B. Bioenergetic role of mitochondrial fusion and fission. Biochim. Biophys. Acta 2012, 1817, 1833–1838. [Google Scholar] [CrossRef] [Green Version]
- Silva Ramos, E.; Larsson, N.G.; Mourier, A. Bioenergetic roles of mitochondrial fusion. Biochim. Biophys. Acta 2016, 1857, 1277–1283. [Google Scholar] [CrossRef]
- Tsukihara, T.; Aoyama, H.; Yamashita, E.; Tomizaki, T.; Yamaguchi, H.; Shinzawa-Itoh, K.; Nakashima, R.; Yaono, R.; Yoshikawa, S. The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8. Science 1996, 272, 1136–1144. [Google Scholar] [CrossRef]
- Kadenbachand, B.; Hüttemann, M. The subunit composition and function of mammalian cytochrome c oxidase. Mitochondrion 2015, 24, 64–76. [Google Scholar] [CrossRef]
- Watson, S.A.; McStay, G.P. Functions of cytochrome c oxidase assembly factors. Int. J. Mol. Sci. 2020, 21, 7254. [Google Scholar] [CrossRef] [PubMed]
- Zong, S.; Wu, M.; Gu, J.; Liu, T.; Guo, R.; Yang, M. Structure of the intact 14-subunit human cytochrome c oxidase. Cell Res. 2018, 28, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Hüttemann, M.; Helling, S.; Sanderson, T.H.; Sinkler, C.; Samavati, L.; Mahapatra, G.; Varughese, A.; Lu, G.; Liu, J.; Ramzan, R.; et al. Regulation of mitochondrial respiration and apoptosis through cell signaling: Cytochrome c oxidase and cytochrome c in ischemia/reperfusion injury and inflammation. Biochim. Biophys. Acta 2012, 1817, 598–609. [Google Scholar] [CrossRef]
- Sinkler, C.A.; Kalpage, H.; Shay, J.; Lee, I.; Malek, M.H.; Grossman, L.I.; Hüttemann, M. Tissue- and condition-specific isoforms of mammalian cytochrome c oxidase subunits: From function to human disease. Oxid. Med. Cell. Longev. 2017, 2017, 1534056. [Google Scholar] [CrossRef] [Green Version]
- Howitzand, K.T.; Sinclair, D.A. Xenohormesis: Sensing the chemical cues of other species. Cell 2008, 133, 387–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibellini, L.; Bianchini, E.; de Biasi, S.; Nasi, M.; Cossarizza, A.; Pinti, M. Natural compounds modulating mitochondrial functions. Evid. Based Complement. Alternat. Med. 2015, 2015, 527209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval-Acuna, C.; Ferreira, J.; Speisky, H. Polyphenols and mitochondria: An update on their increasingly emerging ros-scavenging independent actions. Arch. Biochem. Biophys. 2014, 559, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.F.; Revel, J.S.; Maier, C.S. Mitochondria-centric review of polyphenol bioactivity in cancer models. Antioxid. Redox Signal. 2018, 29, 1589–1611. [Google Scholar] [CrossRef]
- Kicinska, A.; Jarmuszkiewicz, W. Flavonoids and mitochondria: Activation of cytoprotective pathways? Molecules 2020, 25, 3060. [Google Scholar] [CrossRef]
- Raza, H.; John, A.; Brown, E.M.; Benedict, S.; Kambal, A. Alterations in mitochondrial respiratory functions, redox metabolism and apoptosis by oxidant 4-hydroxynonenal and antioxidants curcumin and melatonin in pc12 cells. Toxicol. Appl. Pharmacol. 2008, 226, 161–168. [Google Scholar] [CrossRef]
- Rastogi, M.; Ojha, R.P.; Sagar, C.; Agrawal, A.; Dubey, G.P. Protective effect of curcuminoids on age-related mitochondrial impairment in female wistar rat brain. Biogerontology 2014, 15, 21–31. [Google Scholar] [CrossRef]
- Ray Hamidie, R.D.; Yamada, T.; Ishizawa, R.; Saito, Y.; Masuda, K. Curcumin treatment enhances the effect of exercise on mitochondrial biogenesis in skeletal muscle by increasing camp levels. Metabolism 2015, 64, 1334–1347. [Google Scholar] [CrossRef] [PubMed]
- Jia, N.; Sun, Q.; Su, Q.; Chen, G. Sirt1-mediated deacetylation of pgc1alpha attributes to the protection of curcumin against glutamate excitotoxicity in cortical neurons. Biochem. Biophys. Res. Commun. 2016, 478, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Tang, J.; Li, Y.; Xie, Y.; Shan, H.; Chen, M.; Zhang, J.; Yang, X.; Zhang, Q.; Yang, X. Curcumin attenuates skeletal muscle mitochondrial impairment in copd rats: Pgc-1alpha/sirt3 pathway involved. Chem. Biol. Interact. 2017, 277, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, A.; Plumitallo, C.; de Nuccio, C.; Visentin, S.; Minghetti, L. Curcumin promotes oligodendrocyte differentiation and their protection against tnf-alpha through the activation of the nuclear receptor ppar-gamma. Sci. Rep. 2021, 11, 4952. [Google Scholar] [CrossRef] [PubMed]
- Rayamajhi, N.; Kim, S.K.; Go, H.; Joe, Y.; Callaway, Z.; Kang, J.G.; Ryter, S.W.; Chung, H.T. Quercetin induces mitochondrial biogenesis through activation of ho-1 in hepg2 cells. Oxid. Med. Cell Longev. 2013, 2013, 154279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballmann, C.; Denney, T.S.; Beyers, R.J.; Quindry, T.; Romero, M.; Amin, R.; Selsby, J.T.; Quindry, J.C. Lifelong quercetin enrichment and cardioprotection in mdx/utrn+/- mice. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H128–H140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dave, A.; Shukla, F.; Wala, H.; Pillai, P. Mitochondrial electron transport chain complex dysfunction in mecp2 knock-down astrocytes: Protective effects of quercetin hydrate. J. Mol. Neurosci. 2019, 67, 16–27. [Google Scholar] [CrossRef]
- Sharma, S.; Raj, K.; Singh, S. Neuroprotective effect of quercetin in combination with piperine against rotenone- and iron supplement-induced parkinson’s disease in experimental rats. Neurotox. Res. 2020, 37, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xu, G.; Dong, Y.; Li, M.; Yang, L.; Lu, W. Quercetin protects against lipopolysaccharide-induced intestinal oxidative stress in broiler chickens through activation of nrf2 pathway. Molecules 2020, 25, 1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, M.; Bae, H. An overview of stress-induced resveratrol synthesis in grapes: Perspectives for resveratrol-enriched grape products. Molecules 2017, 22, 294. [Google Scholar] [CrossRef]
- Yu, O.J.; Jez, M. Nature’s assembly line: Biosynthesis of simple phenylpropanoids and polyketides. Plant J. 2008, 54, 750–762. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, M. Resveratrol, a new phenolic compound, from veratrum grandiflorum. J. Chem. Soc. Jpn. 1939, 60, 1090–1100. [Google Scholar]
- Jasinski, M.; Jasinska, L.; Ogrodowczyk, M. Resveratrol in prostate diseases—A short review. Cent. Eur. J. Urol. 2013, 66, 144–149. [Google Scholar] [CrossRef] [Green Version]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef]
- Burkon, A.; Somoza, V. Quantification of free and protein-bound trans-resveratrol metabolites and identification of trans-resveratrol-c/o-conjugated diglucuronides—Two novel resveratrol metabolites in human plasma. Mol. Nutr. Food Res. 2008, 52, 549–557. [Google Scholar] [CrossRef]
- Kopp, P. Resveratrol, a phytoestrogen found in red wine. A possible explanation for the conundrum of the ‘french paradox’? Eur. J. Endocrinol. 1998, 138, 619–620. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.C.; Wu, J.M. Resveratrol: Biological and pharmaceutical properties as anticancer molecule. Biofactors 2010, 36, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Kalantari, H.; Das, D.K. Physiological effects of resveratrol. Biofactors 2010, 36, 401–406. [Google Scholar] [CrossRef]
- Schnekenburger, M.; Dicato, M.; Diederich, M. Plant-derived epigenetic modulators for cancer treatment and prevention. Biotechnol. Adv. 2014, 32, 1123–1132. [Google Scholar] [CrossRef]
- Chan, S.; Kantham, S.; Rao, V.M.; Palanivelu, M.K.; Pham, H.L.; Shaw, P.N.; McGeary, R.P.; Ross, B.P. Metal chelation, radical scavenging and inhibition of abeta(4)(2) fibrillation by food constituents in relation to alzheimer’s disease. Food Chem. 2016, 199, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Blanquer-Rossello, M.D.; Hernandez-Lopez, R.; Roca, P.; Oliver, J.; Valle, A. Resveratrol induces mitochondrial respiration and apoptosis in sw620 colon cancer cells. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 431–440. [Google Scholar] [CrossRef]
- Singh, A.P.; Singh, R.; Verma, S.S.; Rai, V.; Kaschula, C.H.; Maiti, P.; Gupta, S.C. Health benefits of resveratrol: Evidence from clinical studies. Med. Res. Rev. 2019, 39, 1851–1891. [Google Scholar] [CrossRef]
- Zhou, J.; Liao, Z.; Jia, J.; Chen, J.L.; Xiao, Q. The effects of resveratrol feeding and exercise training on the skeletal muscle function and transcriptome of aged rats. PeerJ 2019, 7, e7199. [Google Scholar] [CrossRef]
- Sung, M.M.; Byrne, N.J.; Robertson, I.M.; Kim, T.T.; Samokhvalov, V.; Levasseur, J.; Soltys, C.L.; Fung, D.; Tyreman, N.; Denou, E.; et al. Resveratrol improves exercise performance and skeletal muscle oxidative capacity in heart failure. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H842–H853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamura, K.; Fukumura, S.; Nikaido, K.; Tachi, N.; Kozuka, N.; Seino, T.; Hatakeyama, K.; Mori, M.; Ito, Y.M.; Takami, A.; et al. Resveratrol improves motor function in patients with muscular dystrophies: An open-label, single-arm, phase iia study. Sci. Rep. 2020, 10, 20585. [Google Scholar] [CrossRef]
- Bonnefont-Rousselot, D. Resveratrol and cardiovascular diseases. Nutrients 2016, 8, 250. [Google Scholar] [CrossRef] [PubMed]
- Jardim, F.R.; de Rossi, F.T.; Nascimento, M.X.; da Silva Barros, R.G.; Borges, P.A.; Prescilio, I.C.; de Oliveira, M.R. Resveratrol and brain mitochondria: A review. Mol. Neurobiol. 2018, 55, 2085–2101. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Butt, M.S.; Nadeem, M.; Peters, D.G.; Mubarak, M.S. Resveratrol as an anti-cancer agent: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1428–1447. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.Y.; Tain, Y.L.; Yu, H.R.; Huang, L.T. The effects of resveratrol in the treatment of metabolic syndrome. Int. J. Mol. Sci. 2019, 20, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, V.A.; Patel, K.R.; Viskaduraki, M.; Crowell, J.A.; Perloff, M.; Booth, T.D.; Vasilinin, G.; Sen, A.; Schinas, A.M.; Piccirilli, G.; et al. Repeat dose study of the cancer chemopreventive agent resveratrol in healthy volunteers: Safety, pharmacokinetics, and effect on the insulin-like growth factor axis. Cancer Res. 2010, 70, 9003–9011. [Google Scholar] [CrossRef] [Green Version]
- Ratz-Lyko, A.; Arct, J. Resveratrol as an active ingredient for cosmetic and dermatological applications: A review. J. Cosmet. Laser Ther. 2019, 21, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Schoonjans, K.; Auwerx, J. Sirtuin functions in health and disease. Mol. Endocrinol. 2007, 21, 1745–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.C. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: A dawn for evolutionary medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Ward, W.F. Pgc-1alpha: A key regulator of energy metabolism. Adv. Physiol. Educ. 2006, 30, 145–151. [Google Scholar] [CrossRef]
- Rowe, G.C.; Jiang, A.; Arany, Z. Pgc-1 coactivators in cardiac development and disease. Circ. Res. 2010, 107, 825–838. [Google Scholar] [CrossRef] [Green Version]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating sirt1 and pgc-1alpha. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Pearson, K.J.; Baur, J.A.; Lewis, K.N.; Peshkin, L.; Price, N.L.; Labinskyy, N.; Swindell, W.R.; Kamara, D.; Minor, R.K.; Perez, E.; et al. Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell Metab. 2008, 8, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Higashida, K.; Kim, S.H.; Jung, S.R.; Asaka, M.; Holloszy, J.O.; Han, D.H. Effects of resveratrol and sirt1 on pgc-1alpha activity and mitochondrial biogenesis: A reevaluation. PLoS Biol. 2013, 11, e1001603. [Google Scholar] [CrossRef] [Green Version]
- Jager, S.; Handschin, C.; St-Pierre, J.; Spiegelman, B.M. Amp-activated protein kinase (ampk) action in skeletal muscle via direct phosphorylation of pgc-1alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 12017–12022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulco, M.; Sartorelli, V. Comparing and contrasting the roles of ampk and sirt1 in metabolic tissues. Cell Cycle 2008, 7, 3669–3679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, N.L.; Gomes, A.P.; Ling, A.J.; Duarte, F.V.; Martin-Montalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. Sirt1 is required for ampk activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab. 2012, 15, 675–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.J.; Xu, C.; Liu, M.Y.; Wang, B.K.; Liu, W.B.; Chen, D.H.; Zhang, L.; Xu, C.Y.; Li, X.F. Resveratrol improves the energy sensing and glycolipid metabolism of blunt snout bream megalobrama amblycephala fed high-carbohydrate diets by activating the ampk-sirt1-pgc-1alpha network. Front. Physiol. 2018, 9, 1258. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Wang, L.; Kon, N.; Zhao, W.; Lee, S.; Zhang, Y.; Rosenbaum, M.; Zhao, Y.; Gu, W.; Farmer, S.R.; et al. Brown remodeling of white adipose tissue by sirt1-dependent deacetylation of ppargamma. Cell 2012, 150, 620–632. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Tian, Z.; Zhou, G.; Niu, Q.; Chen, J.; Li, P.; Dong, L.; Xia, T.; Zhang, S.; Wang, A. Sirt1-dependent mitochondrial biogenesis supports therapeutic effects of resveratrol against neurodevelopment damage by fluoride. Theranostics 2020, 10, 4822–4838. [Google Scholar] [CrossRef]
- Csiszar, A.; Labinskyy, N.; Pinto, J.T.; Ballabh, P.; Zhang, H.; Losonczy, G.; Pearson, K.; de Cabo, R.; Pacher, P.; Zhang, C.; et al. Resveratrol induces mitochondrial biogenesis in endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H13–H20. [Google Scholar] [CrossRef] [Green Version]
- Chuang, Y.C.; Chen, S.D.; Hsu, C.Y.; Chen, S.F.; Chen, N.C.; Jou, S.B. Resveratrol promotes mitochondrial biogenesis and protects against seizure-induced neuronal cell damage in the hippocampus following status epilepticus by activation of the pgc-1alpha signaling pathway. Int. J. Mol. Sci. 2019, 20, 998. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Zhang, Y.; Cheng, L.; Zhang, L.; Li, X.; Shi, M.; Zhao, H. Effects of resveratrol combined with soy isoflavones on apoptosis induced by oxidative stress in hippocampus of aging model rats. Wei Sheng Yan Jiu 2020, 49, 932–937. [Google Scholar] [CrossRef]
- Vincenzi, K.L.; Maia, T.P.; Delmonego, L.; Lima, A.B.; Pscheidt, L.C.; Delwing-Dal Magro, D.; Delwing-de Lima, D. Effects of resveratrol on alterations in cerebrum energy metabolism caused by metabolites accumulated in type i citrullinemia in rats. Naunyn. Schmiedebergs Arch. Pharmacol. 2020. [Google Scholar] [CrossRef]
- Low, I.C.; Chen, Z.X.; Pervaiz, S. Bcl-2 modulates resveratrol-induced ros production by regulating mitochondrial respiration in tumor cells. Antioxid. Redox Signal. 2010, 13, 807–819. [Google Scholar] [CrossRef]
- Chen, K.G.; Kang, R.R.; Sun, Q.; Liu, C.; Ma, Z.; Liu, K.; Deng, Y.; Liu, W.; Xu, B. Resveratrol ameliorates disorders of mitochondrial biogenesis and mitophagy in rats continuously exposed to benzo(a)pyrene from embryonic development through adolescence. Toxicology 2020, 442, 152532. [Google Scholar] [CrossRef] [PubMed]
- Saracco, S.A.; Fox, T.D. Cox18p is required for export of the mitochondrially encoded saccharomyces cerevisiae cox2p c-tail and interacts with pnt1p and mss2p in the inner membrane. Mol. Biol. Cell 2002, 13, 1122–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alway, S.E.; McCrory, J.L.; Kearcher, K.; Vickers, A.; Frear, B.; Gilleland, D.L.; Bonner, D.E.; Thomas, J.M.; Donley, D.A.; Lively, M.W.; et al. Resveratrol enhances exercise-induced cellular and functional adaptations of skeletal muscle in older men and women. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 1595–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desquiret-Dumas, V.; Gueguen, N.; Leman, G.; Baron, S.; Nivet-Antoine, V.; Chupin, S.; Chevrollier, A.; Vessieres, E.; Ayer, A.; Ferre, M.; et al. Resveratrol induces a mitochondrial complex i-dependent increase in nadh oxidation responsible for sirtuin activation in liver cells. J. Biol. Chem. 2013, 288, 36662–36675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zini, R.; Morin, C.; Bertelli, A.; Bertelli, A.A.; Tillement, J.P. Effects of resveratrol on the rat brain respiratory chain. Drugs Exp. Clin. Res. 1999, 25, 87–97. [Google Scholar]
- Gledhill, J.R.; Montgomery, M.G.; Leslie, A.G.; Walker, J.E. Mechanism of inhibition of bovine f1-atpase by resveratrol and related polyphenols. Proc. Natl. Acad. Sci. USA 2007, 104, 13632–13637. [Google Scholar] [CrossRef] [Green Version]
- Kipp, J.L.; Ramirez, V.D. Effect of estradiol, diethylstilbestrol, and resveratrol on f0f1-atpase activity from mitochondrial preparations of rat heart, liver, and brain. Endocrine 2001, 15, 165–175. [Google Scholar] [CrossRef]
- Zheng, J.; Ramirez, V.D. Inhibition of mitochondrial proton f0f1-atpase/atp synthase by polyphenolic phytochemicals. Br. J. Pharmacol. 2000, 130, 1115–1123. [Google Scholar] [CrossRef]
- Khanduja, K.L.; Bhardwaj, A. Stable free radical scavenging and antiperoxidative properties of resveratrol compared in vitro with some other bioflavonoids. Indian J. Biochem. Biophys. 2003, 40, 416–422. [Google Scholar]
- Juan, M.E.; Wenzel, U.; Daniel, H.; Planas, J.M. Resveratrol induces apoptosis through ros-dependent mitochondria pathway in ht-29 human colorectal carcinoma cells. J. Agric. Food Chem. 2008, 56, 4813–4818. [Google Scholar] [CrossRef]
- Rai, G.; Suman, S.; Mishra, S.; Shukla, Y. Evaluation of growth inhibitory response of resveratrol and salinomycin combinations against triple negative breast cancer cells. Biomed. Pharmacother. 2017, 89, 1142–1151. [Google Scholar] [CrossRef]
- Luo, H.; Yang, A.; Schulte, B.A.; Wargovich, M.J.; Wang, G.Y. Resveratrol induces premature senescence in lung cancer cells via ros-mediated DNA damage. PLoS ONE 2013, 8, e60065. [Google Scholar] [CrossRef] [Green Version]
- Tseng, S.H.; Lin, S.M.; Chen, J.C.; Su, Y.H.; Huang, H.Y.; Chen, C.K.; Lin, P.Y.; Chen, Y. Resveratrol suppresses the angiogenesis and tumor growth of gliomas in rats. Clin. Cancer Res. 2004, 10, 2190–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stokes, J., 3rd; Singh, U.P.; Scissum-Gunn, K.; Singh, R.; Manne, U.; Mishra, M.K. Prolonged exposure of resveratrol induces reactive superoxide species-independent apoptosis in murine prostate cells. Tumour Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Chavez, J.; Fonseca-Sanchez, M.A.; Arechaga-Ocampo, E.; Flores-Perez, A.; Palacios-Rodriguez, Y.; Dominguez-Gomez, G.; Marchat, L.A.; Fuentes-Mera, L.; Mendoza-Hernandez, G.; Gariglio, P.; et al. Proteomic profiling reveals that resveratrol inhibits hsp27 expression and sensitizes breast cancer cells to doxorubicin therapy. PLoS ONE 2013, 8, e64378. [Google Scholar] [CrossRef] [Green Version]
- Osman, A.M.; Al-Harthi, S.E.; AlArabi, O.M.; Elshal, M.F.; Ramadan, W.S.; Alaama, M.N.; Al-Kreathy, H.M.; Damanhouri, Z.A.; Osman, O.H. Chemosensetizing and cardioprotective effects of resveratrol in doxorubicin- treated animals. Cancer Cell Int. 2013, 13, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, A.M.; Telity, S.A.; Telity, S.A.; Damanhouri, Z.A.; Al-Harthy, S.E.; Al-Kreathy, H.M.; Ramadan, W.S.; Elshal, M.F.; Khan, L.M.; Kamel, F. Chemosensitizing and nephroprotective effect of resveratrol in cisplatin -treated animals. Cancer Cell Int. 2015, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Sprouse, A.A.; Herbert, B.S. Resveratrol augments paclitaxel treatment in mda-mb-231 and paclitaxel-resistant mda-mb-231 breast cancer cells. Anticancer. Res. 2014, 34, 5363–5374. [Google Scholar]
- Tu, L.F.; Cao, L.F.; Zhang, Y.H.; Guo, Y.L.; Zhou, Y.F.; Lu, W.Q.; Zhang, T.Z.; Zhang, T.; Zhang, G.X.; Kurihara, H.; et al. Sirt3-dependent deacetylation of cox-1 counteracts oxidative stress-induced cell apoptosis. FASEB J. 2019, 33, 14118–14128. [Google Scholar] [CrossRef] [PubMed]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.L.; et al. Small molecule activators of sirtuins extend saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef]
- Wood, J.G.; Rogina, B.; Lavu, S.; Howitz, K.; Helfand, S.L.; Tatar, M.; Sinclair, D. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nature 2004, 430, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Rezaizadehnajafi, L.; Wink, M. Influence of resveratrol on oxidative stress resistance and life span in caenorhabditis elegans. J. Pharm. Pharmacol. 2013, 65, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, M.; Kim, S.K.; Berdichevsky, A.; Guarente, L. A role for sir-2.1 regulation of er stress response genes in determining c. Elegans life span. Dev. Cell 2005, 9, 605–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, J.H.; Goupil, S.; Garber, G.B.; Helfand, S.L. An accelerated assay for the identification of lifespan-extending interventions in drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2004, 101, 12980–12985. [Google Scholar] [CrossRef] [Green Version]
- Bhullar, K.S.; Hubbard, B.P. Lifespan and healthspan extension by resveratrol. Biochim. Biophys. Acta 2015, 1852, 1209–1218. [Google Scholar] [CrossRef] [Green Version]
- Hector, K.L.; Lagisz, M.; Nakagawa, S. The effect of resveratrol on longevity across species: A meta-analysis. Biol. Lett. 2012, 8, 790–793. [Google Scholar] [CrossRef]
- Semba, R.D.; Ferrucci, L.; Bartali, B.; Urpi-Sarda, M.; Zamora-Ros, R.; Sun, K.; Cherubini, A.; Bandinelli, S.; Andres-Lacueva, C. Resveratrol levels and all-cause mortality in older community-dwelling adults. JAMA Intern. Med. 2014, 174, 1077–1084. [Google Scholar] [CrossRef] [Green Version]
- De Paepe, B.; Van Coster, R. A critical assessment of the therapeutic potential of resveratrol supplements for treating mitochondrial disorders. Nutrients 2017, 9, 1017. [Google Scholar] [CrossRef] [Green Version]
- Lopes Costa, A.; le Bachelier, C.; Mathieu, L.; Rotig, A.; Boneh, A.; de Lonlay, P.; Tarnopolsky, M.A.; Thorburn, D.R.; Bastin, J.; Djouadi, F. Beneficial effects of resveratrol on respiratory chain defects in patients’ fibroblasts involve estrogen receptor and estrogen-related receptor alpha signaling. Hum. Mol. Genet. 2014, 23, 2106–2119. [Google Scholar] [CrossRef] [Green Version]
- Douiev, L.; Soiferman, D.; Alban, C.; Saada, A. The effects of ascorbate, n-acetylcysteine, and resveratrol on fibroblasts from patients with mitochondrial disorders. J. Clin. Med. 2016, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- De Paepe, B.; Vandemeulebroecke, K.; Smet, J.; Vanlander, A.; Seneca, S.; Lissens, W.; van Hove, J.L.; Deschepper, E.; Briones, P.; van Coster, R. Effect of resveratrol on cultured skin fibroblasts from patients with oxidative phosphorylation defects. Phytother Res. 2014, 28, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, C.; Artacho, R.; Gimenez, R. Beneficial effects of green tea—A review. J. Am. Coll. Nutr. 2006, 25, 79–99. [Google Scholar] [CrossRef]
- Fraga, C.G.; Galleano, M.; Verstraeten, S.V.; Oteiza, P.I. Basic biochemical mechanisms behind the health benefits of polyphenols. Mol. Asp. Med. 2010, 31, 435–445. [Google Scholar] [CrossRef]
- Ottaviani, J.I.; Momma, T.Y.; Heiss, C.; Kwik-Uribe, C.; Schroeter, H.; Keen, C.L. The stereochemical configuration of flavanols influences the level and metabolism of flavanols in humans and their biological activity in vivo. Free Radic. Biol. Med. 2011, 50, 237–244. [Google Scholar] [CrossRef]
- Actis-Goretta, L.; Leveques, A.; Giuffrida, F.; Romanov-Michailidis, F.; Viton, F.; Barron, D.; Duenas-Paton, M.; Gonzalez-Manzano, S.; Santos-Buelga, C.; Williamson, G.; et al. Elucidation of (–)-epicatechin metabolites after ingestion of chocolate by healthy humans. Free Radic. Biol. Med. 2012, 53, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Bernatonieneand, J.D.; Kopustinskiene, M. The role of catechins in cellular responses to oxidative stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [Green Version]
- Mangels, D.R.; Mohler, E.R., 3rd. Catechins as potential mediators of cardiovascular health. Arter. Thromb. Vasc. Biol. 2017, 37, 757–763. [Google Scholar] [CrossRef]
- Farkhondeh, T.; Yazdi, H.S.; Samarghandian, S. The protective effects of green tea catechins in the management of neurodegenerative diseases: A review. Curr. Drug Discov. Technol. 2019, 16, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.; Elbaz, H.A.; Lee, I.; Zielske, S.P.; Malek, M.H.; Hüttemann, M. Molecular mechanisms and therapeutic effects of (–)-epicatechin and other polyphenols in cancer, inflammation, diabetes, and neurodegeneration. Oxid. Med. Cell Longev. 2015, 2015, 181260. [Google Scholar] [CrossRef] [Green Version]
- Cremonini, E.; Iglesias, D.E.; Kang, J.; Lombardo, G.E.; Mostofinejad, Z.; Wang, Z.; Zhu, W.; Oteiza, P.I. (–)-epicatechin and the comorbidities of obesity. Arch. Biochem. Biophys. 2020, 690, 108505. [Google Scholar] [CrossRef]
- Rahman, T.; Hosen, I.; Islam, M.M.T.; Shekhar, H.U. Oxidative stress and human health. Adv. Biosci. Biotechnol. 2012, 3, 997–1019. [Google Scholar] [CrossRef] [Green Version]
- McCullough, M.L.; Chevaux, K.; Jackson, L.; Preston, M.; Martinez, G.; Schmitz, H.H.; Coletti, C.; Campos, H.; Hollenberg, N.K. Hypertension, the kuna, and the epidemiology of flavanols. J. Cardiovasc. Pharmacol. 2006, 47 (Suppl. 2), S103–S109. [Google Scholar] [CrossRef]
- Wollgast, J.; Anklam, E. Review on polyphenols in theobroma cacao: Changes in composition during the manufacture of chocolate and methodology for identification and quantification. Food Res. Int. 2000, 33, 423–447. [Google Scholar] [CrossRef]
- Engler, M.B.; Engler, M.M. The emerging role of flavonoid-rich cocoa and chocolate in cardiovascular health and disease. Nutr. Rev. 2006, 64, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Shrime, M.G.; Bauer, S.R.; McDonald, A.C.; Chowdhury, N.H.; Coltart, C.E.; Ding, E.L. Flavonoid-rich cocoa consumption affects multiple cardiovascular risk factors in a meta-analysis of short-term studies. J. Nutr. 2011, 141, 1982–1988. [Google Scholar] [CrossRef]
- Katz, D.L.; Doughty, K.; Ali, A. Cocoa and chocolate in human health and disease. Antioxid. Redox Signal. 2011, 15, 2779–2811. [Google Scholar] [CrossRef] [Green Version]
- Faridi, Z.; Njike, V.Y.; Dutta, S.; Ali, A.; Katz, D.L. Acute dark chocolate and cocoa ingestion and endothelial function: A randomized controlled crossover trial. Am. J. Clin. Nutr. 2008, 88, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Oteiza, P.I.; Fraga, C.G.; Mills, D.A.; Taft, D.H. Flavonoids and the gastrointestinal tract: Local and systemic effects. Mol. Asp. Med. 2018, 61, 41–49. [Google Scholar] [CrossRef]
- Nehlig, A. The neuroprotective effects of cocoa flavanol and its influence on cognitive performance. Br. J. Clin. Pharmacol. 2013, 75, 716–727. [Google Scholar] [CrossRef]
- Taub, P.R.; Ramirez-Sanchez, I.; Ciaraldi, T.P.; Gonzalez-Basurto, S.; Coral-Vazquez, R.; Perkins, G.; Hogan, M.; Maisel, A.S.; Henry, R.R.; Ceballos, G.; et al. Perturbations in skeletal muscle sarcomere structure in patients with heart failure and type 2 diabetes: Restorative effects of (–)-epicatechin-rich cocoa. Clin. Sci. 2013, 125, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Sanchez, I.; de los Santos, S.; Gonzalez-Basurto, S.; Canto, P.; Mendoza-Lorenzo, P.; Palma-Flores, C.; Ceballos-Reyes, G.; Villarreal, F.; Zentella-Dehesa, A.; Coral-Vazquez, R. (–)-epicatechin improves mitochondrial-related protein levels and ameliorates oxidative stress in dystrophic delta-sarcoglycan null mouse striated muscle. FEBS J. 2014, 281, 5567–5580. [Google Scholar] [CrossRef] [Green Version]
- Angel Garcia-Merino, J.; Moreno-Perezde, D.; Lucas, B.; Montalvo-Lominchar, M.G.; Munoz, E.; Sanchez, L.; Naclerio, F.; Herrera-Rocha, K.M.; Moreno-Jimenez, M.R.; Rocha-Guzman, N.E.; et al. Chronic flavanol-rich cocoa powder supplementation reduces body fat mass in endurance athletes by modifying the follistatin/myostatin ratio and leptin levels. Food Funct. 2020, 11, 3441–3450. [Google Scholar] [CrossRef]
- Cremonini, E.; Fraga, C.G.; Oteiza, P.I. Epicatechin in the control of glucose homeostasis: Involvement of redox-regulated mechanisms. Free Radic. Biol. Med. 2019, 130, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Buitrago-Lopez, A.; Sanderson, J.; Johnson, L.; Warnakula, S.; Wood, A.; di Angelantonio, E.; Franco, O.H. Chocolate consumption and cardiometabolic disorders: Systematic review and meta-analysis. BMJ 2011, 343, d4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daussin, F.N.; Heyman, E.; Burelle, Y. Effects of (–)-epicatechin on mitochondria. Nutr. Rev. 2021, 79, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Dower, J.I.; Geleijnse, J.M.; Gijsbers, L.; Zock, P.L.; Kromhout, D.; Hollman, P.C. Effects of the pure flavonoids epicatechin and quercetin on vascular function and cardiometabolic health: A randomized, double-blind, placebo-controlled, crossover trial. Am. J. Clin. Nutr. 2015, 101, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Salmean, G.; Ortiz-Vilchis, P.; Vacaseydel, C.M.; Garduno-Siciliano, L.; Chamorro-Cevallos, G.; Meaney, E.; Villafana, S.; Villarreal, F.; Ceballos, G.; Ramirez-Sanchez, I. Effects of (–)-epicatechin on a diet-induced rat model of cardiometabolic risk factors. Eur. J. Pharmacol. 2014, 728, 24–30. [Google Scholar] [CrossRef]
- Panneerselvam, M.; Tsutsumi, Y.M.; Bonds, J.A.; Horikawa, Y.T.; Saldana, M.; Dalton, N.D.; Head, B.P.; Patel, P.M.; Roth, D.M.; Patel, H.H. Dark chocolate receptors: Epicatechin-induced cardiac protection is dependent on delta-opioid receptor stimulation. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1604–H1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Sanchez, I.; Mansour, C.; Navarrete-Yanez, V.; Ayala-Hernandez, M.; Guevara, G.; Castillo, C.; Loredo, M.; Bustamante, M.; Ceballos, G.; Villarreal, F. Epicatechin induced reversal of endothelial cell aging and improved vascular function: Underlying mechanisms. Food Funct. 2018, 9, 4802–4813. [Google Scholar] [CrossRef]
- Rozza, A.L.; Hiruma-Lima, C.A.; Tanimoto, A.; Pellizzon, C.H. Morphologic and pharmacological investigations in the epicatechin gastroprotective effect. Evid. Based Complement. Alternat. Med. 2012, 2012, 708156. [Google Scholar] [CrossRef] [Green Version]
- Navarrete-Yanez, V.; Garate-Carrillo, A.; Rodriguez, A.; Mendoza-Lorenzo, P.; Ceballos, G.; Calzada-Mendoza, C.; Hogan, M.C.; Villarreal, F.; Ramirez-Sanchez, I. Effects of (–)-epicatechin on neuroinflammation and hyperphosphorylation of tau in the hippocampus of aged mice. Food Funct. 2020, 11, 10351–10361. [Google Scholar] [CrossRef]
- Nogueira, L.; Ramirez-Sanchez, I.; Perkins, G.A.; Murphy, A.; Taub, P.R.; Ceballos, G.; Villarreal, F.J.; Hogan, M.C.; Malek, M.H. Epicatechin enhances fatigue resistance and oxidative capacity in mouse muscle. J. Physiol. 2011, 589, 4615–4631. [Google Scholar] [CrossRef]
- Schwarz, N.A.; Blahnik, Z.J.; Prahadeeswaran, S.; McKinley-Barnard, S.K.; Holden, S.L.; Waldhelm, A. (–)-epicatechin supplementation inhibits aerobic adaptations to cycling exercise in humans. Front. Nutr. 2018, 5, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, H.; Wang, X.; Zhang, L.; Parnell, L.D.; Admed, B.; LeRoith, T.; Ansah, T.A.; Zhang, L.; Li, J.; Ordovas, J.M.; et al. Dietary epicatechin improves survival and delays skeletal muscle degeneration in aged mice. FASEB J. 2019, 33, 965–977. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Salmean, G.; Ortiz-Vilchis, P.; Vacaseydel, C.M.; Rubio-Gayosso, I.; Meaney, E.; Villarreal, F.; Ramirez-Sanchez, I.; Ceballos, G. Acute effects of an oral supplement of (–)-epicatechin on postprandial fat and carbohydrate metabolism in normal and overweight subjects. Food Funct. 2014, 5, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Cremonini, E.; Bettaieb, A.; Haj, F.G.; Fraga, C.G.; Oteiza, P.I. (–)-epicatechin improves insulin sensitivity in high fat diet-fed mice. Arch. Biochem. Biophys. 2016, 599, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, T.; Nagayasu, S.; Suzuki, S.; Iwashita, M.; Yamashita, A.; Shinjo, T.; Sanui, T.; Kushiyama, A.; Kanematsu, T.; Asano, V.; et al. Epicatechin downregulates adipose tissue ccl19 expression and thereby ameliorates diet-induced obesity and insulin resistance. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 249–259. [Google Scholar] [CrossRef]
- Ramirez-Sanchez, I.; Rodriguez, A.; Moreno-Ulloa, A.; Ceballos, G.; Villarreal, F. (–)-epicatechin-induced recovery of mitochondria from simulated diabetes: Potential role of endothelial nitric oxide synthase. Diab. Vasc. Dis. Res. 2016, 13, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Ulloa, A.; Nogueira, L.; Rodriguez, A.; Barboza, J.; Hogan, M.C.; Ceballos, G.; Villarreal, F.; Ramirez-Sanchez, I. Recovery of indicators of mitochondrial biogenesis, oxidative stress, and aging with (–)-epicatechin in senile mice. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 1370–1378. [Google Scholar] [CrossRef]
- Taub, P.R.; Ramirez-Sanchez, I.; Ciaraldi, T.P.; Perkins, G.; Murphy, A.N.; Naviaux, R.; Hogan, M.; Maisel, A.S.; Henry, R.R.; Ceballos, G.; et al. Alterations in skeletal muscle indicators of mitochondrial structure and biogenesis in patients with type 2 diabetes and heart failure: Effects of epicatechin rich cocoa. Clin. Transl. Sci. 2012, 5, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Ulloa, A.; Cid, A.; Rubio-Gayosso, I.; Ceballos, G.; Villarreal, F.; Ramirez-Sanchez, I. Effects of (–)-epicatechin and derivatives on nitric oxide mediated induction of mitochondrial proteins. Bioorg. Med. Chem. Lett. 2013, 23, 4441–4446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopustinskiene, D.M.; Savickas, A.; Vetchy, D.; Masteikova, R.; Bernatoniene, A.K.J. Direct effects of (–)-epicatechin and procyanidin b2 on the respiration of rat heart mitochondria. Biomed. Res. Int. 2015, 2015, 232836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, C.M.; Ramirez-Sanchez, I.; Oskarsson, B.; Joyce, N.; Aguilar, C.; Nicorici, A.; Dayan, J.; Goude, E.; Abresch, R.T.; Villarreal, F.; et al. (–)-epicatechin induces mitochondrial biogenesis and markers of muscle regeneration in adults with becker muscular dystrophy. Muscle Nerve 2021, 63, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Ulloa, A.; Miranda-Cervantes, A.; Licea-Navarro, A.; Mansour, C.; Beltran-Partida, E.; Donis-Maturano, L.; de la Herran, H.C.D.; Villarreal, F.; Alvarez-Delgado, C. (–)-epicatechin stimulates mitochondrial biogenesis and cell growth in c2c12 myotubes via the g-protein coupled estrogen receptor. Eur. J. Pharmacol. 2018, 822, 95–107. [Google Scholar] [CrossRef]
- Hüttemann, M.; Lee, I.; Malek, M.H. (–)-epicatechin maintains endurance training adaptation in mice after 14 days of detraining. FASEB J. 2012, 26, 1413–1422. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Hüttemann, M.; Malek, M.H. (–)-epicatechin attenuates degradation of mouse oxidative muscle following hindlimb suspension. J. Strength Cond Res. 2016, 30, 1–10. [Google Scholar] [CrossRef]
- Hüttemann, M.; Lee, I.; Perkins, G.A.; Britton, S.L.; Koch, L.G.; Malek, M.H. (–)-epicatechin is associated with increased angiogenic and mitochondrial signalling in the hindlimb of rats selectively bred for innate low running capacity. Clin. Sci. 2013, 124, 663–674. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Hüttemann, M.; Kruger, A.; Bollig-Fischer, A.; Malek, M.H. (–)-epicatechin combined with 8 weeks of treadmill exercise is associated with increased angiogenic and mitochondrial signaling in mice. Front. Pharmacol. 2015, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Bitner, B.F.; Ray, J.D.; Kener, K.B.; Herring, J.A.; Tueller, J.A.; Johnson, D.K.; Freitas, C.M.T.; Fausnacht, D.W.; Allen, M.E.; Thomson, A.H.; et al. Common gut microbial metabolites of dietary flavonoids exert potent protective activities in beta-cells and skeletal muscle cells. J. Nutr. Biochem. 2018, 62, 95–107. [Google Scholar] [CrossRef]
- Varela, C.E.; Rodriguez, A.; Romero-Valdovinos, M.; Mendoza-Lorenzo, P.; Mansour, C.; Ceballos, G.; Villarreal, F.; Ramirez-Sanchez, I. Browning effects of (–)-epicatechin on adipocytes and white adipose tissue. Eur. J. Pharmacol. 2017, 811, 48–59. [Google Scholar] [CrossRef]
- Elbaz, H.A.; Lee, I.; Antwih, D.A.; Liu, J.; Hüttemann, M.; Zielske, S.P. Epicatechin stimulates mitochondrial activity and selectively sensitizes cancer cells to radiation. PLoS ONE 2014, 9, e88322. [Google Scholar] [CrossRef] [Green Version]
- Allgrove, J.; Farrell, E.; Gleeson, M.; Williamson, G.; Cooper, K. Regular dark chocolate consumption’s reduction of oxidative stress and increase of free-fatty-acid mobilization in response to prolonged cycling. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.K.; Brouner, J.; Spendiff, O. Dark chocolate supplementation reduces the oxygen cost of moderate intensity cycling. J. Int. Soc. Sports Nutr. 2015, 12, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carvalho, F.G.; Fisher, M.G.; Thornley, T.T.; Roemer, K.; Pritchett, R.; Freitas, E.C.; Pritchett, K. Cocoa flavanol effects on markers of oxidative stress and recovery after muscle damage protocol in elite rugby players. Nutrition 2019, 62, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.H.; Park, K.S.; Lee, K.U.; Lee, H.K. Mitochondrial metabolism and diabetes. J. Diabetes Investig. 2010, 1, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Castellano-Gonzalez, G.; Pichaud, N.; Ballard, J.W.; Bessede, A.; Marcal, H.; Guillemin, G.J. Epigallocatechin-3-gallate induces oxidative phosphorylation by activating cytochrome c oxidase in human cultured neurons and astrocytes. Oncotarget 2016, 7, 7426–7440. [Google Scholar] [CrossRef]
- Zhao, G.; He, F.; Wu, C.; Li, P.; Li, N.; Deng, J.; Zhu, G.; Ren, W.; Peng, Y. Betaine in inflammation: Mechanistic aspects and applications. Front. Immunol. 2018, 9, 1070. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, A.; Murata, N. Genetic engineering of glycinebetaine synthesis in plants: Current status and implications for enhancement of stress tolerance. J. Exp. Bot. 2000, 51, 81–88. [Google Scholar] [CrossRef]
- Day, C.R.; Kempson, S.A. Betaine chemistry, roles, and potential use in liver disease. Biochim. Biophys. Acta 2016, 1860, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Craig, S.A. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Zeisel, S.H.; Mar, M.H.; Howe, J.C.; Holden, J.M. Concentrations of choline-containing compounds and betaine in common foods. J. Nutr. 2003, 133, 1302–1307. [Google Scholar] [CrossRef]
- Lever, M.; Sizeland, P.C.; Bason, L.M.; Hayman, C.M.; Chambers, S.T. Glycine betaine and proline betaine in human blood and urine. Biochim. Biophys. Acta 1994, 1200, 259–264. [Google Scholar] [CrossRef]
- Schwahn, B.C.; Hafner, D.; Hohlfeld, T.; Balkenhol, N.; Laryea, M.D.; Wendel, U. Pharmacokinetics of oral betaine in healthy subjects and patients with homocystinuria. Br. J. Clin. Pharmacol. 2003, 55, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhang, H.; Li, H.; Lai, F.; Li, X.; Tang, Y.; Min, T.; Wu, H. Antioxidant mechanism of betaine without free radical scavenging ability. J. Agric. Food Chem. 2016, 64, 7921–7930. [Google Scholar] [CrossRef]
- Fernández, C.; Gallego, L.; Lopez-Bote, C.J. Effect of betaine on fat content in growing lambs. Anim. Feed Sci. Technol. 1998, 73, 329–338. [Google Scholar] [CrossRef]
- Eklund, M.; Bauer, E.; Wamatu, J.; Mosenthin, R. Potential nutritional and physiological functions of betaine in livestock. Nutr. Res. Rev. 2005, 18, 31–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.Y.; Xu, Z.R.; Li, W.F. Effects of betaine on growth performance and carcass characteristics in growing pigs. Asian Australas. J. Anim. Sci. 2004, 17, 1700–1704. [Google Scholar] [CrossRef]
- Best, P. Designer nutrition for pigs: Feeding against backfat. Feed Int. 1994, 5, 38–39. [Google Scholar]
- Gudev, D.; Popova-Ralcheva, S.; Yanchev, I.; Moneva, P.; Petkov, E.; Ignatova, M. Effect of betaine on egg performance and some blood constituents in laying hens reared indoor under natural summer temperatures and varying levels of air ammonia. Bulg. J. Agric. Sci. 2011, 17, 859–866. [Google Scholar]
- Al-Shukri, A.Y.; Kaab, H.T.; Abdulwahab, H.M. Effect of betaine on some productive traits of turkey under heat stress. Kufa J. Vet. Med. Sci. 2012, 3, 12–20. [Google Scholar]
- Akhavan-Salamatand, H.H.; Ghasemi, A. Alleviation of chronic heat stress in broilers by dietary supplementation of betaine and turmeric rhizome powder: Dynamics of performance, leukocyte profile, humoral immunity, and antioxidant status. Trop Anim. Health Prod. 2016, 48, 181–188. [Google Scholar] [CrossRef]
- Mendoza, S.M.; Boyd, R.D.; Ferket, P.R.; van Heugten, E. Effects of dietary supplementation of the osmolyte betaine on growing pig performance and serological and hematological indices during thermoneutral and heat-stressed conditions. J. Anim. Sci. 2017, 95, 5040–5053. [Google Scholar] [CrossRef]
- Clarke, W.C.; Virtanen, E.; Blackburn, J.; Higgs, D.A. Effects of a dietary betaine/amino acid additive on growth and seawater adaptation in yearling chinook salmon. Aquaculture 1994, 121, 137–145. [Google Scholar] [CrossRef]
- Ali, S.; Abbas, Z.; Seleiman, M.F.; Rizwan, M.; Yava, S.I.; Alhammad, B.A.; Shami, A.; Hasanuzzaman, M.; Kalderis, D. Glycine betaine accumulation, significance and interests for heavy metal tolerance in plants. Plants 2020, 9, 896. [Google Scholar] [CrossRef]
- Allard, F.; Houde, M.; Krol, M.; Ivanov, A.; Huner, N.P.A.; Sarhan, F. Betaine improves freezing tolerance in wheat. Plant Cell Physiol. 1998, 39, 1194–1202. [Google Scholar] [CrossRef]
- Nomura, M.; Muramoto, Y.; Yasuda, S.; Takabe, T.; Kishitani, S. The accumulation of glycinebetaine during cold acclimation in early and late cultivars of barley. Euphytica 1995, 83, 247–250. [Google Scholar] [CrossRef]
- Zou, H.; Chen, N.; Shi, M.; Xian, M.; Song, Y.; Liu, J. The metabolism and biotechnological application of betaine in microorganism. Appl. Microbiol. Biotechnol. 2016, 100, 3865–3876. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Xu, P. Betaine and beet molasses enhance l-lactic acid production by bacillus coagulans. PLoS ONE 2014, 9, e100731. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.C.; Hynes, S.H.; Ingledew, W.M. Effects of particulate materials and osmoprotectants on very-high-gravity ethanolic fermentation by saccharomyces cerevisiae. Appl. Environ. Microbiol. 1994, 60, 1519–1524. [Google Scholar] [CrossRef] [Green Version]
- Holtmann, G.; Bremer, E. Thermoprotection of bacillus subtilis by exogenously provided glycine betaine and structurally related compatible solutes: Involvement of opu transporters. J. Bacteriol. 2004, 186, 1683–1693. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Zhang, H.; Guo, X.F.; Li, K.; Li, S.; Li, D. Effect of betaine on reducing body fat-a systematic review and meta-analysis of randomized controlled trials. Nutrients 2019, 11, 2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lever, M.; Slow, S. The clinical significance of betaine, an osmolyte with a key role in methyl group metabolism. Clin. Biochem. 2010, 43, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.S.; Kumar, T.; Dar, T.A.; Singh, L.R. Protein n-homocysteinylation: From cellular toxicity to neurodegeneration. Biochim. Biophys. Acta 2015, 1850, 2239–2245. [Google Scholar] [CrossRef] [PubMed]
- Wilcken, D.E.; Wilcken, B.; Dudman, N.P.; Tyrrell, P.A. Homocystinuria--the effects of betaine in the treatment of patients not responsive to pyridoxine. N. Engl. J. Med. 1983, 309, 448–453. [Google Scholar] [CrossRef]
- Da Deminice, R.; Silva, R.P.; Lamarre, S.G.; Kelly, K.B.; Jacobs, R.L.; Brosnan, M.E.; Brosnan, J.T. Betaine supplementation prevents fatty liver induced by a high-fat diet: Effects on one-carbon metabolism. Amino Acids 2015, 47, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Huang, L.; Gao, J.; Wen, S.; Tai, Y.; Chen, M.; Huang, Z.; Liu, R.; Tang, C.; Li, J. Betaine attenuates chronic alcoholinduced fatty liver by broadly regulating hepatic lipid metabolism. Mol. Med. Rep. 2017, 16, 5225–5234. [Google Scholar] [CrossRef] [Green Version]
- Arroyave-Ospina, J.C.; Wu, Z.; Geng, Y.; Moshage, H. Role of oxidative stress in the pathogenesis of non-alcoholic fatty liver disease: Implications for prevention and therapy. Antioxidants 2021, 10, 174. [Google Scholar] [CrossRef]
- You, M.; Crabb, D.W. Recent advances in alcoholic liver disease ii. Minireview: Molecular mechanisms of alcoholic fatty liver. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G1–G6. [Google Scholar] [CrossRef] [Green Version]
- Idalsoaga, F.; Kulkarni, A.V.; Mousa, O.Y.; Arrese, M.; Arab, J.P. Non-alcoholic fatty liver disease and alcohol-related liver disease: Two intertwined entities. Front. Med. 2020, 7, 448. [Google Scholar] [CrossRef]
- Kathirvel, E.; Morgan, K.; Nandgiri, G.; Sandoval, B.C.; Caudill, M.A.; Bottiglieri, T.; French, S.W.; Morgan, T.R. Betaine improves nonalcoholic fatty liver and associated hepatic insulin resistance: A potential mechanism for hepatoprotection by betaine. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G1068–G1077. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Deaciuc, I.; Zhou, Z.; Song, M.; Chen, T.; Hill, D.; McClain, C.J. Involvement of amp-activated protein kinase in beneficial effects of betaine on high-sucrose diet-induced hepatic steatosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G894–G902. [Google Scholar] [CrossRef]
- Ji, C.; Kaplowitz, N. Betaine decreases hyperhomocysteinemia, endoplasmic reticulum stress, and liver injury in alcohol-fed mice. Gastroenterology 2003, 124, 1488–1499. [Google Scholar] [CrossRef]
- Bucheli, P.; Gao, Q.; Redgwell, R.; Vidal, K.; Wang, J.; Zhang, W. Biomolecular and clinical aspects of chinese wolfberry. In Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; Benzie, I.F.F., Wachtel-Galor, S., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 289–314. [Google Scholar]
- Mukherjee, S. Betaine and nonalcoholic steatohepatitis: Back to the future? World J. Gastroenterol. 2011, 17, 3663–3664. [Google Scholar] [CrossRef]
- Mukherjee, S. Role of betaine in liver disease-worth revisiting or has the die been cast? World J. Gastroenterol. 2020, 26, 5745–5748. [Google Scholar] [CrossRef] [PubMed]
- Ueland, P.M. Choline and betaine in health and disease. J. Inherit. Metab. Dis. 2011, 34, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Henke, W.; Herdel, K.; Jung, K.; Schnorr, D.; Loening, S.A. Betaine improves the pcr amplification of gc-rich DNA sequences. Nucleic Acids Res. 1997, 25, 3957–3958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleland, D.; Krader, P.; McCree, C.; Tang, J.; Emerson, D. Glycine betaine as a cryoprotectant for prokaryotes. J. Microbiol. Methods 2004, 58, 31–38. [Google Scholar] [CrossRef]
- Armstrong, L.E.; Casa, D.J.; Roti, M.W.; Lee, E.C.; Craig, S.A.; Sutherland, J.W.; Fiala, K.A.; Maresh, C.M. Influence of betaine consumption on strenuous running and sprinting in a hot environment. J. Strength Cond Res. 2008, 22, 851–860. [Google Scholar] [CrossRef]
- Hoffman, J.R.; Ratamess, N.A.; Kang, J.; Gonzalez, A.M.; Beller, N.A.; Craig, S.A. Effect of 15 days of betaine ingestion on concentric and eccentric force outputs during isokinetic exercise. J. Strength Cond Res. 2011, 25, 2235–2241. [Google Scholar] [CrossRef]
- Trepanowski, J.F.; Farney, T.M.; McCarthy, C.G.; Schilling, B.K.; Craig, S.A.; Bloomer, R.J. The effects of chronic betaine supplementation on exercise performance, skeletal muscle oxygen saturation and associated biochemical parameters in resistance trained men. J. Strength Cond Res. 2011, 25, 3461–3471. [Google Scholar] [CrossRef]
- Cholewa, J.M.; Wyszczelska-Rokiel, M.; Glowacki, R.; Jakubowski, H.; Matthews, T.; Wood, R.; Craig, S.A.; Paolone, V. Effects of betaine on body composition, performance, and homocysteine thiolactone. J. Int. Soc. Sports Nutr. 2013, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, G.Y.; Won, S.B.; Kim, J.; Jeon, S.; Han, A.; Kwon, Y.H. Betaine alleviates hypertriglycemia and tau hyperphosphorylation in db/db mice. Toxicol. Res. 2013, 29, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varatharajalu, R.; Garige, M.; Leckey, L.C.; Arellanes-Robledo, J.; Reyes-Gordillo, K.; Shah, R.; Lakshman, M.R. Adverse signaling of scavenger receptor class b1 and pgc1s in alcoholic hepatosteatosis and steatohepatitis and protection by betaine in rat. Am. J. Pathol. 2014, 184, 2035–2044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Meng, X.; Kang, S.Y.; Zhang, J.; Jung, H.W.; Park, Y.K. Regulatory effects of the fruit extract of lycium chinense and its active compound, betaine, on muscle differentiation and mitochondrial biogenesis in c2c12 cells. Biomed. Pharm. 2019, 118, 109297. [Google Scholar] [CrossRef]
- Tan, G.Y.; Yang, L.; Fu, Y.Q.; Feng, J.H.; Zhang, M.H. Effects of different acute high ambient temperatures on function of hepatic mitochondrial respiration, antioxidative enzymes, and oxidative injury in broiler chickens. Poult. Sci. 2010, 89, 115–122. [Google Scholar] [CrossRef]
- Huang, C.; Jiao, H.; Song, Z.; Zhao, J.; Wang, X.; Lin, H. Heat stress impairs mitochondria functions and induces oxidative injury in broiler chickens. J. Anim. Sci. 2015, 93, 2144–2153. [Google Scholar] [CrossRef]
- Ala, F.S.; Ahmad, H.; Golian, A. Effects of dietary supplemental methionine source and betaine replacement on the growth performance and activity of mitochondrial respiratory chain enzymes in normal and heatstressed broiler chickens. J. Anim. Physiol. Anim. Nutr. 2018, 103, 87–99. [Google Scholar]
- Kharbanda, K.K.; Todero, S.L.; King, A.L.; Osna, N.A.; McVicker, B.L.; Tuma, D.J.; Wisecarver, J.L.; Bailey, S.M. Betaine treatment attenuates chronic ethanol-induced hepatic steatosis and alterations to the mitochondrial respiratory chain proteome. Int. J. Hepatol. 2012, 2012, 962183. [Google Scholar] [CrossRef]
- Ganesan, B.; Rajesh, R.; Anandan, R.; Dhandapani, N. Biochemical studies on the protective effect of betaine on mitochondrial function in experimentally induced myocardial infarction in rats. J. Health Sci. 2007, 53, 671–681. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Sun, Q.; Liu, J.; Jia, Y.; Cai, D.; Idriss, A.A.; Omer, N.A.; Zhao, R. In ovo injection of betaine alleviates corticosterone-induced fatty liver in chickens through epigenetic modifications. Sci. Rep. 2017, 7, 40251. [Google Scholar] [CrossRef]
- Lee, I. Betaine is a positive regulator of mitochondrial respiration. Biochem. Biophys. Res. Commun. 2015, 456, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Apicella, J.M.; Lee, E.C.; Bailey, B.L.; Saenz, C.; Anderson, J.M.; Craig, S.A.; Kraemer, W.J.; Volek, J.S.; Maresh, C.M. Betaine supplementation enhances anabolic endocrine and akt signaling in response to acute bouts of exercise. Eur. J. Appl. Physiol. 2013, 113, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Song, H.P.; Chu, Z.G.; Zhang, D.X.; Dang, Y.M.; Zhang, Q. Pi3k-akt pathway protects cardiomyocytes against hypoxia-induced apoptosis by mitokatp-mediated mitochondrial translocation of pakt. Cell Physiol. Biochem. 2018, 49, 717–727. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structures of resveratrol (A), (–)-epicatechin (B), and betaine (C).

Figure 2.
Regulatory mechanisms of resveratrol (RSV), (–)-epicatechin (EPI), and betaine (BET) on cytochrome c oxidase (COX): ROS, reactive oxygen species; SIRT1, sirtuin 1; AMPK, AMP activated protein kinase; PGC1α, peroxisome proliferator activated receptor gamma coactivator 1 alpha; NRFs, nuclear respiratory factors; TFAM, mitochondrial transcription factor A.
Figure 2.
Regulatory mechanisms of resveratrol (RSV), (–)-epicatechin (EPI), and betaine (BET) on cytochrome c oxidase (COX): ROS, reactive oxygen species; SIRT1, sirtuin 1; AMPK, AMP activated protein kinase; PGC1α, peroxisome proliferator activated receptor gamma coactivator 1 alpha; NRFs, nuclear respiratory factors; TFAM, mitochondrial transcription factor A.


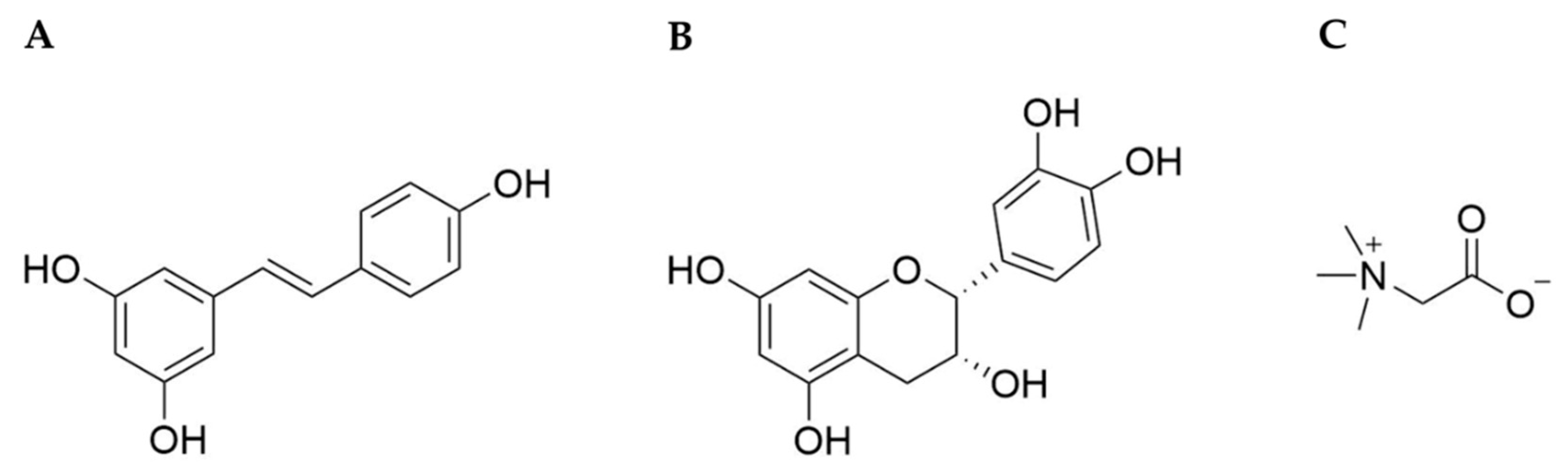{kind=link}
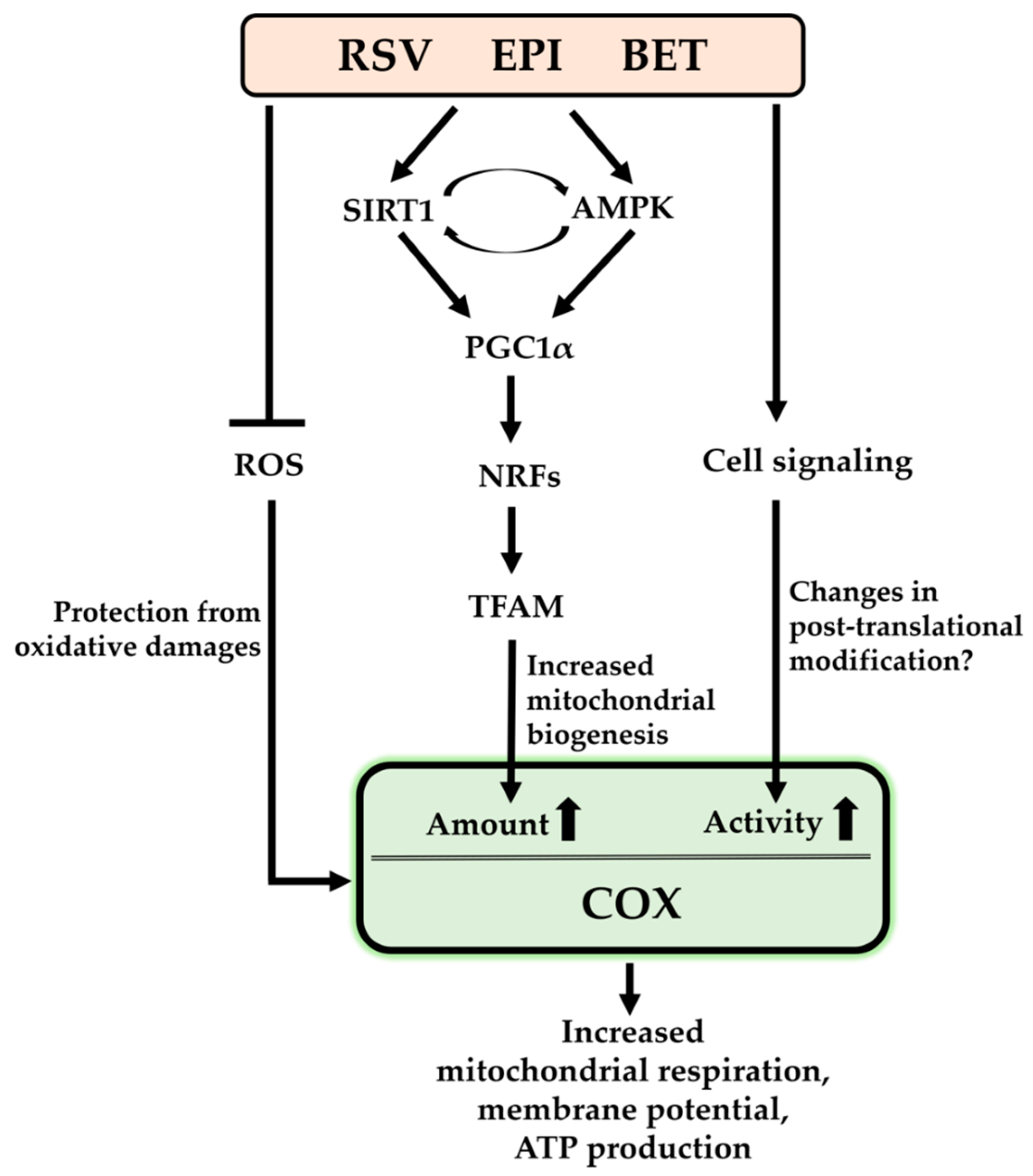{kind=link}
Table 1.
Effect of resveratrol (RSV) administration on cytochrome c oxidase (COX).
| Reference | Experimental Subject | Experimental Condition | Result |
|---|---|---|---|
| [57] | Gastrocnemius muscle from 8-week-old male C57BL/6J mice with high fat diet | Supplementation of 400 mg/kg/day RSV to high fat diet for 15 weeks | Increased mRNA levels of COX Va, COX IV-1, COX Vb, COX VIIaL |
| [62] | Cultured C2C12 mouse myoblast cells | Incubation with 25 μM RSV for 24 h | Increased mRNA level of COX Vb |
| [62] | Gastrocnemius muscle and hepatocytes from C57BL/6J mice with high fat diet; WT mouse for SIRT1-KO model | Supplementation of 25~30 mg/kg/day RSV with high fat diet for 8 months | Increased mRNA level of COX Vb |
| [67] | Hippocampus from male Sprague Dawley rats with status epilepticus induced by kainic acid | Microinjection of 100 μmol RSV into hippocampus prior to kainic acid treatment | Increased COX I amount |
| [68] | Hippocampus from aged female Sprague Dawley rats induced by bilateral ovariectomy combined with intraperitoneal injection of D-galactose | Intragastric administration of 80 mg/kg/day RSV alone and in combination with isoflavones for 12 weeks | Increased COX I amount by RSV alone and in combination with isoflavones |
| [69] | Hippocampus, cerebral cortex, and cerebellum from 60-day-old male Wistar rats | Incubation with 0.1, 0.5, 5 mM RSV for 1 h in combination with citrulline or ammonia in brain homogenates | Increased COX activity in a dose-dependent manner; increased COX activity in the presence of citrulline by 0.1, 0.5 mM RSV in cerebral cortex and 5 mM RSV in hippocampus & increased COX activity in the presence of ammonia by 0.1, 0.5 mM RSV in cerebral cortex and cerebellum |
| [70] | Cultured human lymphoblastic leukemia cells overexpressed with Bcl-2; CEM/Bcl-2 | Incubation with 10, 30, 50 μM RSV for 6 h | Decreased COX activity in a dose-dependent manner, while no change in COX I amount (50 μM RSV for 18 h) |
| [41] | Cultured human colon cancer cells; SW620 | Incubation with 10 μM RSV for 48 h | Increased COX activity (~45%) and COX I amount |
Table 2.
Effect of (–)-epicatechin (EPI) administration on cytochrome c oxidase (COX).
| Reference | Experimental Subject | Experimental Condition | Result |
|---|---|---|---|
| [144] | Cultured C2C12 mouse myoblast cells | Incubation with 10 μM EPI for 48 h | Increased COX I amount |
| [145] | Quadriceps femoris from detrained 5-month-old C57BL/6 male mice | Intragastric administration of 1 mg/kg of EPI twice a day during 14 days of detraining after 5 weeks of training | Increased COX activity (~108%) and COX II amount |
| [146] | Gastrocnemius muscle from hindlimb suspended 6-month-old male C57BL/6N mice | Intragastric administration of 1 mg/kg of EPI twice a day during 14 days of hindlimb suspension | Increased COX I amount |
| [147] | Plantaris muscle from 5-month-old male rats with congenital low running capacity | Intragastric administration of 1 mg/kg of EPI twice a day for 30 days | Increased COX II amount |
| [149] | Cultured rat β cells; INS-1 derived 832/13 cells | Incubation with 10 μM EPI for 24 h | Increased COX I amount |
| [150] | Cultured adipocytes excised from human subcutaneous adipose tissue | Incubation with 100 nM EPI for 72 h | Increased COX I amount |
| [129] | Cultured cow coronary artery endothelial cells with low passage number (young) and high passage number (aged) | Incubation with 1 μM EPI for 48 h | Increased COX I amount in both young and aged cells |
| [151] | Cultured Panc-1 pancreatic cancer cells | Incubation with 20, 50, 100, 200 μM EPI for 1 h | Increased COX activity in a dose-dependent manner (~59% by 200 μM EPI) |
| [147] | Plantaris muscle from healthy Sprague Dawley rats | Incubation with 20 μM EPI for 25 min | Increased COX activity |
| [148] | Quadriceps femoris muscle from 14-month-old male C57BL/6N mice | Incubation with 20 μM EPI for 25 min | Increased COX activity |
Table 3.
Effect of betaine (BET) administration on cytochrome c oxidase (COX).
| Reference | Experimental Subject | Experimental Condition | Result |
|---|---|---|---|
| [209] | Livers from 45–48-day-old male Wister rats | Supplementation of 1% (w/v) BET to ethanol diet for 4–5 weeks | Increased amount of both COX holoenzyme and subunit I by BET in alcoholic liver, while no changes in controls |
| [210] | Hearts from Wistar male rats with myocardial infarction induced by isoprenaline | Intragastric administration of 250 mg/kg/day BET for 30 days prior to isoprenaline injection | Increased COX activity by BET in myocardial infarction, while no changes in controls |
| [211] | Livers from 8-week-old chickens injected with corticosterone | In ovo injection of 2.5 mg BET | Increased mRNA levels of COX I, II, and III and COX activity in corticosterone treated liver, while no changes in controls |
| [212] | Cultured H.2.35 mouse hepatocytes | Incubation of 0.5, 1, 2, 5, 10 mM BET for 30 min | Increased COX activity in a dose-dependent manner up to 2mM BET, while no change in activity of purified COX by incubation with EPI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, I. Regulation of Cytochrome c Oxidase by Natural Compounds Resveratrol, (–)-Epicatechin, and Betaine. Cells 2021, 10, 1346. https://doi.org/10.3390/cells10061346
AMA Style
Lee I. Regulation of Cytochrome c Oxidase by Natural Compounds Resveratrol, (–)-Epicatechin, and Betaine. Cells. 2021; 10(6):1346. https://doi.org/10.3390/cells10061346
Chicago/Turabian StyleLee, Icksoo. 2021. "Regulation of Cytochrome c Oxidase by Natural Compounds Resveratrol, (–)-Epicatechin, and Betaine" Cells 10, no. 6: 1346. https://doi.org/10.3390/cells10061346
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.