The Impact of Oxidative Stress on Ribosomes: From Injury to Regulation


Department of Cell Biology and Neuroscience, Rowan University School of Osteopathic Medicine, Stratford, NJ 08084, USA
*
Authors to whom correspondence should be addressed.
Cells 2019, 8(11), 1379; https://doi.org/10.3390/cells8111379
Submission received: 27 September 2019
/
Revised: 23 October 2019
/
Accepted: 30 October 2019
/
Published: 2 November 2019
(This article belongs to the Section Intracellular and Plasma Membranes)
Abstract
:The ribosome is a complex ribonucleoprotein-based molecular machine that orchestrates protein synthesis in the cell. Both ribosomal RNA and ribosomal proteins can be chemically modified by reactive oxygen species, which may alter the ribosome′s functions or cause a complete loss of functionality. The oxidative damage that ribosomes accumulate during their lifespan in a cell may lead to reduced or faulty translation and contribute to various pathologies. However, remarkably little is known about the biological consequences of oxidative damage to the ribosome. Here, we provide a concise summary of the known types of changes induced by reactive oxygen species in rRNA and ribosomal proteins and discuss the existing experimental evidence of how these modifications may affect ribosome dynamics and function. We emphasize the special role that redox-active transition metals, such as iron, play in ribosome homeostasis and stability. We also discuss the hypothesis that redox-mediated ribosome modifications may contribute to adaptive cellular responses to stress.
1. Introduction
Life in an oxygen-containing environment involves dealing with the omnipresent products of incomplete reduction of oxygen, known as reactive oxygen species (ROS) [1,2]. Depending on context and concentration, ROS can either play essential biochemical and signaling roles, or inflict damage on cellular components [3]. To minimize ROS-mediated damage, cells developed a variety of defensive enzymatic and nonenzymatic mechanisms that keep ROS levels within the optimal or tolerable range. When the load of ROS exceeds the capacity of the cellular antioxidant systems, a multitude of cellular functions may be affected, a state referred to as oxidative stress [4].
How oxidative stress affects the process of protein synthesis is a question of considerable interest for biomedical research. Excessive ROS generation is a recurrent molecular feature in the pathology and treatment of human disease, including neurodegeneration, cancer, and vascular disorders [5,6,7]. Many studies have shown that oxidative stress inhibits protein synthesis in cells [8,9]. However, a severe reduction of translation could be detrimental during stress as it is precisely the time when cells require new protein synthesis in order to repair damage and adapt to the new environment [10]. Recently developed technologies, such as ribosome profiling and quantitative proteomics, have revealed large-scale alterations in the translational landscape during oxidative stress, often preceding transcriptional changes [11,12]. Quick changes in translation may allow cells to more effectively react to adverse conditions, thereby reducing the extent of ROS-inflicted damage [13].
Due to the fact that the protein biosynthetic machinery is complex, it presents numerous control points for stress response regulation. However, complex machinery can malfunction when its critical parts are damaged. Studies using model organisms from every domain of life have uncovered ROS-induced chemical alterations in aminoacyl-tRNA-synthetases and protein factors involved in the initiation, elongation, and termination of translation [8,14,15,16,17,18,19]. Exposure to oxidants affects posttranslational modifications and the stability of tRNAs [20,21,22]. In some cases, ROS-mediated cleavages of mature tRNAs can result in the production of tRNA fragments [23,24,25] that either reprogram or inhibit translation [26,27,28,29]. Other studies demonstrated that damage to mRNAs by ROS may both stall translation and induce translational errors [30,31,32].
Among translation components, the impact of oxidative stress on the central player in this process, the ribosome, remains the least studied. Ribosomes are large ribonucleoprotein-based molecular machines composed of two subunits; the large subunits (LSU) in cytoplasmic ribosomes are referred to as 60S in eukaryotes and 50S in prokaryotes, while the small subunits (SSU) are called 40S and 30S, respectively. Each ribosomal subunit is built around intricately folded RNA molecules (rRNAs), which are bound by several dozen ribosomal proteins (r-proteins) [33]. In addition to cytoplasmic ribosomes, distinct types of ribosomes are present in mitochondria and plastids. As these energy-generating organelles produce large quantities of ROS, protecting their ribosomes against oxidative damage may require additional safeguard mechanisms, details of which are only beginning to come to light. In this review, we focus primarily on cytoplasmic ribosomes and first provide a concise summary of the known types of chemical changes in ribosomal components, induced by increased levels of ROS. We then discuss the impact of oxidative damage on ribosome function and outline areas for future exploration.
2. Oxidative Modification of rRNA
rRNA constitutes the structural and functional core of the ribosome. ROS can affect RNA in a multitude of ways [34,35,36,37], including chemical modification of the base and sugar moieties [38], generation of abasic sites [39], and strand breaks [40]. Guanine is the most easily oxidized nucleobase [41] and best studied in this respect. An oxidized form of guanine, 8-oxo-7,8-dihydroguanine (8-hydroxyguanine; 8-oxo-G), is a ubiquitous oxidative lesion, readily detectable in cellular nucleic acids [34,42]. When present in mRNA, 8-oxo-G interferes with decoding, possibly through the formation of a Hoogsteen pair with adenine when the base rotates around the N-glycosidic bond [43]. The altered base-pairing capacity of 8-oxo-G can also perturb RNA folding. A recent study revealed a variety of outcomes when 8-oxo-G was incorporated into model RNA substrates, from stabilization of existing structural motifs to their destabilization and rearrangements into new structures [44]. Since ribosome activities in translation depend on many precisely tuned conformational changes and movements within its rRNA framework [45], oxidation of the bases that are critical for maintaining the correct rRNA structure may impair ribosome functions. It seems therefore likely that the need to withstand a certain degree of oxidation was one of the forces in shaping the evolution of the ribosome. A striking disappearance of many guanines from mitochondrial rRNA [46] and reduced overall RNA content [47] may be two such evolutionary adaptations that “hardened” mitoribosomes against damage from the abundant ROS generated inside mitochondria.
The extent of cellular RNA oxidation was reported to exceed that of DNA when cells were directly treated with H2O2 [40,48] or subjected to conditions that induce secondary oxidative damage, such as UVA irradiation [49] and ammonia overload [50]. In one study [40], exposure of E. coli cells to H2O2 was shown to induce a dosage-dependent increase of 8-oxo-G in all tested RNA types, including rRNA. By comparing 8-oxo-G levels after in vitro H2O2 treatment of RNAs in their native and denatured forms, the same study concluded that folding of rRNA did not protect it against oxidative damage [40]. Similar conclusions were made by Willi and colleagues, who used an 8-oxo-G antibody to immunoprecipitate oxidized RNA from H2O2-treated E. coli cells [51].
A large body of observations points to the association of oxidized RNA with disease [52,53,54]. High 8-oxo-G content in cytoplasmic and nucleolar RNA was found in vulnerable neurons in Alzheimer′s disease (AD) [55], Parkinson’s disease [56], and neurons of the hippocampal region and temporal neocortex in patients with Lewy body dementia [57]. Elevated levels of 8-oxo-G in RNA were also detected in neuropsychiatric disorders, including schizophrenia, depressive, and bipolar disorders [58]. Furthermore, RNA extracted from atherosclerotic plaques was heavily modified with 8-oxo-G [59]. Although links between 8-oxo-G and human pathologies have been based largely on the analysis of total RNA, rRNA is the most abundant cellular RNA type, which typically accounts for about 80% of the total RNA [60], from which one can reasonably assume that oxidative damage to ribosomes in all these pathological conditions must be widespread. Studies that focused specifically on rRNA confirmed it to be a major target of the 8-oxo-G modification in patients with AD and mild cognitive impairment, a condition that often precedes AD [61,62].
3. Reversible Oxidation of Cysteine and Methionine in r-Proteins
The cyclic oxidation and reduction of cysteine (Cys) and methionine (Met) residues in a protein represents an important mechanism in the regulation of protein functions. The sulfur atom present in the fully reduced free thiol (-SH) side chain of Cys can be oxidized to the disulfide bond (Cys-S-S-Cys), sulfenic (-SOH), sulfinic (SO2H), and sulfonic acids (-SO3H), thereby making Cys an exquisite redox stress sensor [63]. Analysis of the data generated by semiquantitative mass spectrometry coupled with thiol trapping suggests that some r-proteins may be oxidized during normal growth in yeast cells [64]. Similar results were obtained in the human cell line HT-29 [65]. R-proteins represented a prominent cluster in the quantitative whole-cell redoxome analysis using oxidative isotope-coded affinity tags (OxICAT) in different organisms including E. coli [66], S. cerevisiae [9,67], C. elegans [68], and D. melanogaster [69]. In all cases, OxICAT identified r-proteins from both subunits, suggesting that oxidation of Cys residues may influence various stages and features of translation. Based on yeast studies, Topf and colleagues further proposed that r-proteins can act as redox sensors, mediating the attenuation of protein synthesis in response to oxidative stress [9]. Exactly which r-proteins might affect the translation inhibition remains to be determined. It is also currently unclear whether attenuation of protein synthesis represents the main outcome of thiol oxidation in r-proteins. Considering that OxICAT identified a number of r-proteins in which oxidative stress affects redox state, it is tempting to speculate that different patterns of oxidation in r-proteins could lead to variable translational remodeling as a means to adapt to oxidative stress, although more studies will be needed to test this hypothesis.
Similar to Cys, Met residues can be readily oxidized, leading to the formation of methionine sulfoxide [70]. Little is known about the potential contribution of oxidized Met residues to changes in ribosome function. Two independent studies published in 1978 reported oxidation of Met residues of the L7/L12 r-protein in bacterial ribosomes [71,72]. L7 and L12 are identical, except that L7 is acetylated at the N terminus; L7/L12 are normally present on E. coli 50S subunits as two dimers that constitute the L7/L12 stalk, important for interactions with translation factors [73,74]. It was found that oxidation of the Met residues in L7/L12 abolished the dimer formation, affecting stalk structure and altering the ability of the bacterial ribosome to interact with the elongation factor EF-G [71,72].
4. Oxidative Damage to r-Proteins
The effects of ROS on proteins are mediated to a large extent through initial modifications of cysteine, methionine, tyrosine, tryptophan, and histidine residues, which in turn may give rise to a complex set of secondary reactions that are damaging for the proteins and other biological molecules with which they interact [75,76,77]. One important type of modification is carbonylation, mostly affecting arginine, lysine, proline, threonine, and a few other amino acid side chains [78]. Whereas cells maintain enzymatic systems that can repair oxidized Cys and Met residues [79], carbonylation is not repairable and may irreversibly damage and/or destabilize proteins. Several excellent reviews discuss carbonylation mechanisms in detail [77,80,81,82].
Carbonylation presents technical challenges for detection because it generates a wide variety of adducts [83]. For example, oxidation of tryptophan alone results in the formation of seven potential carbonylated products [84,85,86]. Nevertheless, techniques have been developed to reliably identify carbonylated proteins, modification sites, and types of carbonylation by stabilizing carbonyl moieties followed by chromatographic enrichment and proteomics-based analysis [81,87,88]. A recent high-throughput screen to examine carbonyl-modified proteins detected components of the translation machinery, including several amino-acyl tRNA synthetases and two r-proteins (Rpl32 and Rpl35) in HeLa cells treated with low doses of H2O2 (0.05 mM–0.5 mM) [89]. In another study, Mirzaei and Regnier found that a major group of proteins carbonylated in S. cerevisiae in response to a high-dose (5 mM) H2O2 treatment is comprised of ribosomal and ribosome-associated proteins [90,91].
Among the reactive derivatives produced by oxidation of cellular components, aldehydes such as malonaldidehyde (MDA) and 4-hydroxy-2-nonenal (HNE) operate as agents of secondary protein carbonylation. In cells, both of these aldehydes originate from the ROS-induced peroxidation of fatty acids and are commonly used as oxidative stress markers [80]. In human colorectal carcinoma RKO cells, 18 translation-related HNE-modified proteins, including six r-proteins, were significantly increased after treatment with different doses of HNE [92]. How modification with this product of lipid peroxidation may affect ribosome function remains unknown.
In addition to carbonylation, proteomic studies provided evidence of widespread crosslinking between r-proteins and the ribosome′s rRNA backbone in response to acute oxidative stress. In yeast cells treated with high doses of H2O2, 37 r-proteins were reported to form cross-links with RNA [91]. The functional impact of the rRNA-protein crosslinking is yet to be determined.
5. Ubiquitination of r-Proteins Following Oxidative Stress
In addition to attacking ribosome components, ROS may alter activities of ribosome-modifying enzymes and thus affect the ribosome indirectly. The modification of eukaryotic ribosomes with the 76-amino acid polypeptide ubiquitin (Ub) is one such example. Ubiquitination of r-proteins on ribosomes made nonfunctional by a mutation in rRNA was first observed a decade ago [93], and later shown to occur on ribosomes isolated from cells treated with H2O2 [94].
Ub moieties can be covalently attached to Lys residues present either on the target protein or on another Ub polypeptide. Given that Ub contains seven Lys residues, different types of poly-Ub chains can be assembled, including those linked through Lys48 (K48-Ub chains) or through Lys63 (K63-Ub chains). It is generally accepted that the K48-Ub chains primarily label proteins for degradation by proteasomes, while the K63-Ub chains play regulatory roles [95]. By applying a new mass spectrometry-based technique called Ub-DiGGer to detect specific Ub linkages, Silva and colleagues have recently shown that ribosomes are prominent targets of polyubiquitination with K63-Ub chains [96,97]. Modification with K63-Ub chains was detected in r-proteins in yeast and mammalian cells grown under normal conditions and found to increase during oxidative stress. The K63-Ub chains decorated r-proteins located on the solvent-exposed surfaces of the ribosome and were particularly abundant around the head of the 40S subunit. The yeasts Rad6 and Bre1 were identified as the E2/E3 enzymes responsible for the ubiquitination of r-proteins with K63-Ub chains, while deubiquitinase Ubp2 was implicated in removing the K63-Ub chains from the ribosome. On the mechanistic side, the control of the Rad6/Bre1-Ubp2 circuit is accomplished through the oxidation of a catalytically active Cys residue in Ubp2; this oxidation inactivates the enzyme, shifting the balance towards the ubiquitinated ribosomes. The same group later identified r-proteins as a major group of oxidized cellular targets that undergo K48-Ub chain polyubiquitination and subsequent degradation [98]. The attachment of K48-Ub chains was found to occur in two major stages: as an immediate response to oxidative stress and during the recovery period. Together, these findings illustrate the complexity of r-protein ubiquitination and hint at a potentially important role of this type of ribosome modification as a regulatory mechanism in the changing redox environment.
6. Role of Metals in the Effects of Oxidants on the Ribosome
Although readily diffusible, H2O2 reacts poorly with most biological macromolecules [99]. In DNA studies, it has been well established that mutagenic properties of H2O2 depend largely on its reactions with redox-active transition metals [1]. The best documented example involves ferrous iron (Fe2+), which can damage biomolecules through the generation of the highly reactive hydroxyl radical (OH•) in the Fenton reaction:
L-Fe2+ + H2O2 → L-Fe3+ + OH• + OH¯.
When the ligand (L) represents a nucleic acid or a protein, metal binding often occurs in a sequence and structure-dependent manner. This creates active centers for localized generation of OH•, resulting in site-specific damage [100,101]. Complexes of higher oxidation states of iron can damage macromolecules through additional mechanisms that are dependent on the nature of the ligand and are not always easy to precisely define [99].
In our recent studies, we have found that intracellular iron is responsible for site-specific cleavages in yeast rRNA that occur in ribosomes when cells experience oxidative stress [102,103]. One prominent cleavage in the expansion segment 7 (ES7L) of 25S rRNA was detectable by northern analysis within one minute of treating yeast cells with ROS inducers and could also be reproduced by redox cycling of iron bound to purified ribosomes in vitro [103]. Iron binding to ribosomes in cells appears to play roles that, depending on context, could be either regulatory or destructive. Iron-mediated cleavage in ES7L occurs in wild-type cells during mild redox perturbations that make cells more resistant to subsequent oxidative challenges [102], suggesting a role in the adaptive response to stress. Mutations that lead to perturbations in iron homeostasis, by contrast, provoke excessive rRNA fragmentation and loss of cell viability after mild oxidant challenges, from which wild-type cells fully recover [103].
When tested in anoxic conditions, Fe2+ ions can functionally substitute Mg2+ in rRNA folding and cell-free translation, owing to the close geometry of the RNA-Mg2+ and RNA-Fe2+ complexes and possibly reflecting the ribosome′s origins in an iron-rich and oxygen-poor prebiotic environment [104,105]. Given iron′s potential to promote free-radical formation, it is not surprising that its metabolism in aerobic organisms is tightly regulated [106]. Notably, in vitro reactions using high concentrations of Fe2+ and H2O2, similar to those used for hydroxyl radical footprinting [107], produce massive degradation of rRNA [108]. It is generally accepted that most intracellular iron is chelated but can be exchanged between its biological ligands [109]. Thus, under normal circumstances when intracellular iron is properly controlled, iron is more likely to promote limited, site-specific modifications in ribosomes upon their encounter with ROS. It is noteworthy that aside from RNA strand cleavages, iron can potentiate many other types of covalent RNA and protein modifications induced by ROS [101]. In a way, iron binding can make the ribosome more “redox-aware”.
Binding of other metals can also induce chemical changes in biological macromolecules, either through metal ions directly reacting with the bound molecules or by facilitating localized generation of ROS. For example, amino acid residues next to various metal-binding sites in proteins have been shown to be highly prone to metal-catalyzed oxidation (MCO) [77], which could represent another mechanism for metal-dependent ROS effects on the ribosome. Binding of Pb2+ to purified E. coli ribosomes was shown to generate site-specific cleavages in rRNA, which were counteracted by divalent metal ions competing for binding to the same sites [110]. As an illustration of the contribution of environmental metals to ribosome damage, studies in mussels exposed to Cu2+, Cd2+ and Hg2+ found increased levels of 8-oxo-G and strand breaks in rRNA [111]. It will be important to determine to what extent metals may be responsible for potentiating ribosome damage in other situations, including human disease. As discussed above, accumulation of oxidative damage in rRNA is observed in brain tissues in a number of pathological states associated with disruptions in proteostasis [53,55,61,62]. Likewise, abnormal iron homeostasis and its altered cellular distribution is a well-documented feature in brain pathologies, including neurodegenerative conditions such as AD and Parkinson′s diseases, as well as traumatic brain injury [112,113,114]. Copper is another transition metal capable of promoting redox damage to nucleic acids and proteins [100], with known links to neurodegeneration [115].
7. How Oxidative Modifications of the Ribosome May Affect Cellular Functions
Although a variety of modifications in ribosomal components caused by oxidative stress have been identified to date (Table 1), few studies have attempted to address the biological role of these modifications. Subjecting ribosomes to an intense oxidant treatment was shown to reduce incorporation of labeled Phe into products of cell-free translation reactions driven by poly(U) [108]. Oxidative damage to specific nucleobases or specific amino acid residues in r-proteins is probably more physiologically relevant, but also more difficult to address experimentally. A recent study by the Polacek group [51] stands out for its elegant use of an atomic mutagenesis approach to reconstitute T. aquaticus 50S subunits with a single oxidized base at several functionally important positions around the ribosome′s peptidyl transferase center. Remarkably, while some of these single-base substitutions were found to inhibit translation of model mRNA substrates in vitro, others had no measurable effect or even increased the yield of the translated product. These varied outcomes highlight the complexity of oxidant effects on ribosome functions in translation. The situation is likely even more complex in vivo, where ribosomes have to synthesize much more challenging polypeptides and correctly process numerous regulatory signals.
As already mentioned, aside from promoting direct oxidation of ribosomal components, oxidative stress may also influence activities of ribosome-modifying enzymes. Given the dearth of mechanistic studies, for the most part we can currently only speculate about the functional significance of individual modifications. One plausible scenario is that an altered behavior of the modified ribosomes might contribute to the rapid translational shifts that occur after cell exposure to oxidants [12]. Other modifications might mark individual ribosomal subunits for a mechanical action by cellular enzymes, sequestration, or degradation. Indeed, recent studies identified yeast Rps3 (uS3) as a ubiquitination substrate that controls the release of the 40S subunit from translational stalls, followed by the subunit′s decay [116,117]. Ub modifications of the ribosome were also observed in yeast during starvation as part of ribophagy, a specialized form of autophagy that removes excessive ribosomes to the vacuole [118], and during the unfolded protein response to proteotoxic stress [119], known to perturb redox balance in cells [120,121].
While changes in r-protein ubiquitination manifest prominently after oxidant treatments, this likely represents only the tip of the iceberg. A recently performed affinity purification of mammalian ribosomes identified several hundred ribosome-associated proteins including RNA- and protein-modifying enzymes, as well as proteins involved in redox regulation [122]. Another group of potentially interesting redox effectors are Fe(II)/2-oxoglutarate (2OG)-dependent oxygenases [123], shown to catalyze hydroxylation of Arg, Pro, and His residues in r-proteins in both prokaryotes and eukaryotes [124,125,126,127]. One 2OG-dependent oxygenase, RlhA, is responsible for hydroxylation of the cytosine residue C2501 in the peptidyl transferase center of the E. coli LSU [128] and hypothesized to play a role in environmental stress responses [129]. 2OG-dependent oxygenases depend on the presence of reduced iron in their active center [130] and might potentially modulate ribosome functions in a redox- and metal-dependent manner.
The specificity of ribosome modifications observed under mild oxidative stress, such as site-specific rRNA cleavages [102] or Ub modifications of specific r-proteins [96], is consistent with the idea that some of the oxidative stress-driven ribosome modifications may play regulatory roles. More studies are clearly needed to determine how redox-induced modifications in ribosomes might help cells to cope with various stress-inducing conditions.
Obviously, not all changes in ribosomes caused by ROS occur in the context of adaptive cellular responses. Fragmentation of rRNA has been observed in mammalian and yeast studies as part of programmed cell death [131,132]. Whether the destruction of the cell’s ribosomes helps to execute the apoptotic program or simply represents an end result of the nuclease release and/or increased ROS production during this process [133] remains unknown. The cause and effect relationships also remain insufficiently well understood in various cases of increased rRNA oxidation associated with organ and tissue pathology discussed above. Another series of intriguing observations links accumulation of oxidative lesions in RNA to age-related disease [134]. Among unicellular organisms, an increased level of protein carbonylation in ribosomal particles has been observed in the nonculturable population of cells from a stationary-phase E. coli culture [135]. The idea that the competence of the protein-synthetic apparatus may be one of the factors in the aging process, as proposed in the early days of molecular biology by Orgel [136], has been difficult to prove, but as more information about ribosome damage becomes available, this point warrants a thorough reinvestigation.
8. Mechanisms for Dealing with Ribosome Damage: Reduce, Repair, Recycle
Cells have evolved enzymes to eliminate 8-oxo-GTP from the nucleotide pool as a way to reduce its incorporation into nucleic acids [137,138,139,140]. Thus, newly synthesized ribosomes should be relatively free of oxidative lesions in their rRNA, but they may accumulate oxidative damage during their lifetime in the cell. A substantial amount of oxidation was detected in rRNA isolated from E. coli cells, even if the cells were not subjected to any special oxidant treatment [51]. Interestingly, E. coli cells under normal growth conditions were found to contain lower levels of 8-oxo-G in rRNA than the rest of cellular RNA [40]. Theoretically, this could be due to either efficient elimination of damaged ribosomes from cells or the presence of some kind of repair mechanisms specific for ribosomes. Repair systems are essential for neutralizing the oxidative damage in DNA molecules incurred in the aerobic environment [1,141]. It is generally assumed that damaged RNA is degraded rather than repaired, a notion well supported by studies of mRNA [34,36,142]. Unlike mRNAs, which can be relatively easily remade, ribosomes are large and complex molecular machines that are expensive for the cell to build; destroying the entire ribosome seems a wasteful strategy if damage is repairable. In fact, replacement of some damaged r-proteins in fully assembled ribosomes has been already demonstrated [143]. Repair of oxidized Cys and Met residues in r-proteins should, in principle, also be possible through the activities of oxidoreductases such as thioredoxins, glutaredoxins, and methionine sulfoxide reductases [79]. To date, the most compelling case for repair of environmental damage in RNA is provided by oxidative demethylases, enzymes shown to reverse alkylation damage in both DNA and RNA [144]. It remains to be seen whether this and other types of damage to rRNA molecules, including oxidative damage, can be repaired in cells.
A number of nucleases have been implicated in the decay of rRNA in cytoplasmic ribosomes [145,146,147,148]. To effectively participate in oxidative damage control, these enzymes must either recognize oxidative lesions themselves or be guided to the damaged ribosomes by cofactors. A few examples of selectivity towards oxidative lesions in RNA have been described for human proteins [149,150], although it is unknown whether these proteins might contribute to decay of damaged rRNA. Lack of bacterial polynucleotide phosphorylase (PNPase), which binds to oxidized RNA, was shown to cause higher accumulation of 8-oxo-G and hypersensitivity of cells to H2O2 [151,152]. A similar behavior was reported for the human mitochondrial PNPase, although the precise mechanism by which PNPase promotes the selective reduction of oxidized RNA in the cell requires further investigation [153]. Studies in algae suggested a role for RBCL, a subunit of the Rubisco protein, in the management of oxidized RNA within chloroplasts [154]. Mechanisms that counteract oxidative damage may also operate during ribosome assembly steps. The apurinic/apyrimidinic endonuclease APE1, shown to cleave abasic sites within RNA in vitro [155], was found to accumulate in the nucleolus in interphase HeLa cells, making this protein well positioned to serve as a quality control factor in ribosome biosynthesis [156].
The complex folding of rRNA and its association with ribosomal proteins makes ribosomes rather difficult substrates for nucleases. How the large ribosomal particles are dismantled for the effective digestion of rRNA is not well understood. Studies demonstrating the role of the ubiquitin-proteasome system in eukaryotic ribosome degradation [93,94,117] point to the possible contribution of components of this system to the disassembly of ribosomal particles. Interestingly, inhibition of the proteasome in primary neuron and astrocyte cultures was found to strongly increase 8-oxo-G content in RNA, accompanied by diminished 18S and 28S rRNA levels and the appearance of cross-linked RNA species [157]. Autophagy is another general mechanism that eukaryotic cells utilize to eliminate excessive ribosomes [158]. Starvation conditions have been shown to greatly increase autophagic delivery of ribosomes to the lysosome, a process termed ribophagy [118,159,160]. The idea that lysosome-targeting ribophagy mechanisms [118,161] may also operate on damaged ribosomes is attractive, but still awaits direct experimental confirmation. Using ribosomal protein fusions with the pH-sensitive reporter Keima, An and Harper recently observed a modestly increased ribophagic flux after treatment of HEK293 cells with H2O2 [159]. Future studies will be needed to elucidate details of ribosome degradation mechanisms and determine the roles of the cytosolic and lysosome machineries in removing oxidatively damaged ribosomes from the translating pool.
9. Conclusions and Perspectives
In contrast to the progress achieved in deciphering normal ribosome functions in translation, our understanding of the oxidative damage to ribosomes and its impact on cell physiology is still very limited. It is important to keep in mind that oxidative stress comes in many “shades of grey”, with variations in the intensity, duration, types and location of the generated ROS and their primary cellular targets [2]. Thus, it is likely that a spectrum of oxidative modifications of the ribosome exists in cells. One intriguing possibility, which still needs experimental validation, is that localized redox modifications of rRNA and r-proteins, occurring when ROS levels in a cell increase, might confer distinctive properties to some ribosomes, thereby facilitating adaptive processes such as translation of stress-response genes (Figure 1). In effect, the total cellular ribosomal pool could separate into functionally nonidentical subpopulations under stress, each one performing their duties differently from the others. Considering experimental challenges in the way of rigorously proving ribosome specialization [162,163], we expect that establishing the existence of such ROS-induced ribosome heterogeneity would similarly require a lot of supporting biochemical work. Severe oxidative stress is likely to result in widespread ribosome damage (Figure 1). As a result of the ribosome dysfunction, newly translated proteins may be inactive, interfere with normal cellular processes through dominant-negative mechanisms or promote pathological aggregate formation, all of which could have broad implications for human disease.
Based on the information available to us today, some of the challenging questions to be addressed in the future studies include the following:
Which ribosomal parts can undergo oxidation without losing functionality and which ones are easily damaged by oxidants?
How do ROS-induced ribosome modifications affect translation and to what extent do these modifications help cells to fight oxidative stress?
What are the consequences of ribosome damage for cell growth, survival, and tissue functions? How much ribosome damage can different cell types tolerate?
Is there a special degradation pathway(s) for ribosomes made dysfunctional by oxidative damage?
Another challenge will be to understand to what extent ROS-induced alterations in ribosome functions act as a driving force in disease etiology and progression, as well as the full diagnostic value of these alterations as a biomarker. Only a handful of studies to date have made an attempt to address these issues, and the presented evidence remains mostly circumstantial. It is clear that many interesting discoveries still lie ahead.
Funding
This work was funded by the NIH grant R01GM114308 to N.S.
Conflicts of Interest
The authors declare no conflict of interest
References
- Imlay, J.A. Cellular defenses against superoxide and hydrogen peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. Reconciling the chemistry and biology of reactive oxygen species. Nat. Chem. Biol. 2008, 4, 278–286. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Cobley, J.N.; Fiorello, M.L.; Bailey, D.M. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol. 2018, 15, 490–503. [Google Scholar] [CrossRef] [PubMed]
- Hornsveld, M.; Dansen, T.B. The Hallmarks of Cancer from a Redox Perspective. Antioxid. Redox Signal. 2016, 25, 300–325. [Google Scholar] [CrossRef] [PubMed]
- Madamanchi, N.R.; Vendrov, A.; Runge, M.S. Oxidative stress and vascular disease. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 29–38. [Google Scholar] [CrossRef]
- Shenton, D.; Smirnova, J.B.; Selley, J.N.; Carroll, K.; Hubbard, S.J.; Pavitt, G.D.; Ashe, M.P.; Grant, C.M. Global translational responses to oxidative stress impact upon multiple levels of protein synthesis. J. Biol. Chem. 2006, 281, 29011–29021. [Google Scholar] [CrossRef]
- Topf, U.; Suppanz, I.; Samluk, L.; Wrobel, L.; Böser, A.; Sakowska, P.; Knapp, B.; Pietrzyk, M.K.; Chacinska, A.; Warscheid, B. Quantitative proteomics identifies redox switches for global translation modulation by mitochondrially produced reactive oxygen species. Nat. Commun. 2018, 9, 324. [Google Scholar] [CrossRef]
- Zhu, M.; Dai, X. Maintenance of translational elongation rate underlies the survival of Escherichia coli during oxidative stress. Nucleic Acids Res. 2019, 47, 7592–7604. [Google Scholar] [CrossRef]
- Costello, J.L.; Kershaw, C.J.; Castelli, L.M.; Talavera, D.; Rowe, W.; Sims, P.F.G.; Ashe, M.P.; Grant, C.M.; Hubbard, S.J.; Pavitt, G.D. Dynamic changes in eIF4F-mRNA interactions revealed by global analyses of environmental stress responses. Genome Biol. 2017, 18, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerashchenko, M.V.; Lobanov, A.V.; Gladyshev, V.N. Genome-wide ribosome profiling reveals complex translational regulation in response to oxidative stress. Proc. Natl. Acad. Sci. USA 2012, 109, 17394–17399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blevins, W.R.; Tavella, T.; Moro, S.G.; Blasco-Moreno, B.; Closa-Mosquera, A.; Díez, J.; Carey, L.B.; Albà, M.M. Extensive post-transcriptional buffering of gene expression in the response to severe oxidative stress in baker’s yeast. Sci. Rep. 2019, 9, 11005. [Google Scholar] [CrossRef] [PubMed]
- Doronina, V.A.; Staniforth, G.L.; Speldewinde, S.H.; Tuite, M.F.; Grant, C.M. Oxidative stress conditions increase the frequency of de novo formation of the yeast [PSI+] prion. Mol. Microbiol. 2015, 96, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Zhang, Y.; Zeng, H.; Novoa, I.; Lu, P.D.; Calfon, M.; Sadri, N.; Yun, C.; Popko, B.; Paules, R.; et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol. Cell 2003, 11, 619–633. [Google Scholar] [CrossRef]
- Ling, J.; Söll, D. Severe oxidative stress induces protein mistranslation through impairment of an aminoacyl-tRNA synthetase editing site. Proc. Natl. Acad. Sci. USA 2010, 107, 4028–4033. [Google Scholar] [CrossRef] [Green Version]
- Nagano, T.; Yutthanasirikul, R.; Hihara, Y.; Hisabori, T.; Kanamori, T.; Takeuchi, N.; Ueda, T.; Nishiyama, Y. Oxidation of translation factor EF-G transiently retards the translational elongation cycle in Escherichia coli. J. Biochem. 2015, 158, 165–172. [Google Scholar] [CrossRef]
- Netzer, N.; Goodenbour, J.M.; David, A.; Dittmar, K.A.; Jones, R.B.; Schneider, J.R.; Boone, D.; Eves, E.M.; Rosner, M.R.; Gibbs, J.S.; et al. Innate immune and chemically triggered oxidative stress modifies translational fidelity. Nature 2009, 462, 522–526. [Google Scholar] [CrossRef]
- Sideri, T.C.; Koloteva-Levine, N.; Tuite, M.F.; Grant, C.M. Methionine oxidation of Sup35 protein induces formation of the [PSI+] prion in a yeast peroxiredoxin mutant. J. Biol. Chem. 2011, 286, 38924–38931. [Google Scholar] [CrossRef]
- Chan, C.T.Y.; Dyavaiah, M.; DeMott, M.S.; Taghizadeh, K.; Dedon, P.C.; Begley, T.J. A quantitative systems approach reveals dynamic control of tRNA modifications during cellular stress. PLoS Genet. 2010, 6, e1001247. [Google Scholar] [CrossRef]
- Endres, L.; Dedon, P.C.; Begley, T.J. Codon-biased translation can be regulated by wobble-base tRNA modification systems during cellular stress responses. RNA Biol. 2015, 12, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, C.; Begley, T.J.; Dedon, P.C. tRNA modifications regulate translation during cellular stress. FEBS Lett. 2014, 588, 4287–4296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czech, A.; Wende, S.; Mörl, M.; Pan, T.; Ignatova, Z. Reversible and rapid transfer-RNA deactivation as a mechanism of translational repression in stress. PLoS Genet. 2013, 9, e1003767. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Feng, J.; Liu, Q.; Sun, F.; Tie, Y.; Zhu, J.; Xing, R.; Sun, Z.; Zheng, X. Stress induces tRNA cleavage by angiogenin in mammalian cells. FEBS Lett. 2009, 583, 437–442. [Google Scholar] [CrossRef]
- Raina, M.; Ibba, M. tRNAs as regulators of biological processes. Front. Genet. 2014, 5, 171. [Google Scholar] [CrossRef] [Green Version]
- Gebetsberger, J.; Zywicki, M.; Künzi, A.; Polacek, N. tRNA-derived fragments target the ribosome and function as regulatory non-coding RNA in Haloferax volcanii. Archaea 2012, 2012, 260909. [Google Scholar] [CrossRef]
- Ivanov, P.; Emara, M.M.; Villen, J.; Gygi, S.P.; Anderson, P. Angiogenin-induced tRNA fragments inhibit translation initiation. Mol. Cell 2011, 43, 613–623. [Google Scholar] [CrossRef]
- Ivanov, P.; O’Day, E.; Emara, M.M.; Wagner, G.; Lieberman, J.; Anderson, P. G-quadruplex structures contribute to the neuroprotective effects of angiogenin-induced tRNA fragments. Proc. Natl. Acad. Sci. USA 2014, 111, 18201–18206. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Sun, L.; Kragler, F. The phloem-delivered RNA pool contains small noncoding RNAs and interferes with translation. Plant. Physiol. 2009, 150, 378–387. [Google Scholar] [CrossRef]
- Dai, D.-P.; Gan, W.; Hayakawa, H.; Zhu, J.-L.; Zhang, X.-Q.; Hu, G.-X.; Xu, T.; Jiang, Z.-L.; Zhang, L.-Q.; Hu, X.-D.; et al. Transcriptional mutagenesis mediated by 8-oxoG induces translational errors in mammalian cells. Proc. Natl. Acad. Sci. USA 2018, 115, 4218–4222. [Google Scholar] [CrossRef] [Green Version]
- Simms, C.L.; Hudson, B.H.; Mosior, J.W.; Rangwala, A.S.; Zaher, H.S. An active role for the ribosome in determining the fate of oxidized mRNA. Cell Rep. 2014, 9, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Chock, P.B.; Stadtman, E.R. Oxidized messenger RNA induces translation errors. Proc. Natl. Acad. Sci. USA 2007, 104, 66–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnikov, S.; Ben-Shem, A.; Garreau de Loubresse, N.; Jenner, L.; Yusupova, G.; Yusupov, M. One core, two shells: Bacterial and eukaryotic ribosomes. Nat. Struct. Mol. Biol. 2012, 19, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Malla, S.; Shin, B.; Li, J.M. Battle against RNA oxidation: Molecular mechanisms for reducing oxidized RNA to protect cells. Wiley Interdiscip Rev. RNA 2014, 5, 335–346. [Google Scholar] [CrossRef]
- Nunomura, A.; Lee, H.-G.; Zhu, X.; Perry, G. Consequences of RNA oxidation on protein synthesis rate and fidelity: Implications for the pathophysiology of neuropsychiatric disorders. Biochem. Soc. Trans. 2017, 45, 1053–1066. [Google Scholar] [CrossRef]
- Simms, C.L.; Zaher, H.S. Quality control of chemically damaged RNA. Cell. Mol. Life Sci. 2016, 73, 3639–3653. [Google Scholar] [CrossRef] [Green Version]
- Wurtmann, E.J.; Wolin, S.L. RNA under attack: Cellular handling of RNA damage. Crit. Rev. Biochem. Mol. Biol. 2009, 44, 34–49. [Google Scholar] [CrossRef] [Green Version]
- Küpfer, P.A.; Leumann, C.J. Oxidative damage on RNA nucleobases. In Chemical biology of nucleic acids: Fundamentals and clinical applications; RNA Technologies; Springer: Berlin/Heidelberg, Germany, 2014; pp. 75–94. ISBN 978-3-642-54452-1. [Google Scholar]
- Tanaka, M.; Han, S.; Song, H.; Küpfer, P.A.; Leumann, C.J.; Sonntag, W.E. An assay for RNA oxidation induced abasic sites using the Aldehyde Reactive Probe. Free Radic. Res. 2011, 45, 237–247. [Google Scholar] [CrossRef]
- Liu, M.; Gong, X.; Alluri, R.K.; Wu, J.; Sablo, T.; Li, Z. Characterization of RNA damage under oxidative stress in Escherichia coli. Biol. Chem. 2012, 393, 123–132. [Google Scholar] [CrossRef]
- Steenken, S. Purine bases, nucleosides, and nucleotides: Aqueous solution redox chemistry and transformation reactions of their radical cations and e- and OH adducts. Chem. Rev. 1989, 89, 503–520. [Google Scholar] [CrossRef]
- Radak, Z.; Boldogh, I. 8-Oxo-7,8-dihydroguanine: Links to gene expression, aging, and defense against oxidative stress. Free Radic. Biol. Med. 2010, 49, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, E.N.; Simms, C.L.; Keedy, H.E.; Zaher, H.S. Insights into the base-pairing preferences of 8-oxoguanosine on the ribosome. Nucleic Acids Res. 2019, 47, 9857–9870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.J.; Gibala, K.S.; Ayele, T.; Deventer, K.V.; Resendiz, M.J.E. Biophysical properties, thermal stability and functional impact of 8-oxo-7,8-dihydroguanine on oligonucleotides of RNA-a study of duplex, hairpins and the aptamer for preQ1 as models. Nucleic Acids Res. 2017, 45, 2099–2111. [Google Scholar] [CrossRef] [PubMed]
- Noeske, J.; Cate, J.H. Structural basis for protein synthesis: Snapshots of the ribosome in motion. Curr. Opin. Struct. Biol. 2012, 22, 743–749. [Google Scholar] [CrossRef]
- Hosseini, M.; Roy, P.; Sissler, M.; Zirbel, C.L.; Westhof, E.; Leontis, N. How to fold and protect mitochondrial ribosomal RNA with fewer guanines. Nucleic Acids Res. 2018, 46, 10946–10968. [Google Scholar] [CrossRef]
- Ramrath, D.J.F.; Niemann, M.; Leibundgut, M.; Bieri, P.; Prange, C.; Horn, E.K.; Leitner, A.; Boehringer, D.; Schneider, A.; Ban, N. Evolutionary shift toward protein-based architecture in trypanosomal mitochondrial ribosomes. Science 2018, 362, eaau7735. [Google Scholar] [CrossRef]
- Hofer, T.; Badouard, C.; Bajak, E.; Ravanat, J.-L.; Mattsson, A.; Cotgreave, I.A. Hydrogen peroxide causes greater oxidation in cellular RNA than in DNA. Biol. Chem. 2005, 386, 333–337. [Google Scholar] [CrossRef]
- Wamer, W.G.; Wei, R.R. In vitro photooxidation of nucleic acids by ultraviolet A radiation. Photochem. Photobiol. 1997, 65, 560–563. [Google Scholar]
- Görg, B.; Qvartskhava, N.; Keitel, V.; Bidmon, H.J.; Selbach, O.; Schliess, F.; Häussinger, D. Ammonia induces RNA oxidation in cultured astrocytes and brain in vivo. Hepatology 2008, 48, 567–579. [Google Scholar] [CrossRef]
- Willi, J.; Küpfer, P.; Evéquoz, D.; Fernandez, G.; Katz, A.; Leumann, C.; Polacek, N. Oxidative stress damages rRNA inside the ribosome and differentially affects the catalytic center. Nucleic Acids Res. 2018, 46, 1945–1957. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.; Lin, C.-L.G. Oxidative damage to RNA: Mechanisms, consequences, and diseases. Cell. Mol. Life Sci. 2010, 67, 1817–1829. [Google Scholar] [CrossRef] [PubMed]
- Nunomura, A.; Moreira, P.I.; Castellani, R.J.; Lee, H.-G.; Zhu, X.; Smith, M.A.; Perry, G. Oxidative damage to RNA in aging and neurodegenerative disorders. Neurotox Res. 2012, 22, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, H.E.; Specht, E.; Broedbaek, K.; Henriksen, T.; Ellervik, C.; Mandrup-Poulsen, T.; Tonnesen, M.; Nielsen, P.E.; Andersen, H.U.; Weimann, A. RNA modifications by oxidation: A novel disease mechanism? Free Radic. Biol. Med. 2012, 52, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Nunomura, A.; Perry, G.; Pappolla, M.A.; Wade, R.; Hirai, K.; Chiba, S.; Smith, M.A. RNA oxidation is a prominent feature of vulnerable neurons in Alzheimer’s disease. J. Neurosci. 1999, 19, 1959–1964. [Google Scholar] [CrossRef]
- Zhang, J.; Perry, G.; Smith, M.A.; Robertson, D.; Olson, S.J.; Graham, D.G.; Montine, T.J. Parkinson’s disease is associated with oxidative damage to cytoplasmic DNA and RNA in substantia nigra neurons. Am. J. Pathol. 1999, 154, 1423–1429. [Google Scholar] [CrossRef]
- Nunomura, A.; Chiba, S.; Kosaka, K.; Takeda, A.; Castellani, R.J.; Smith, M.A.; Perry, G. Neuronal RNA oxidation is a prominent feature of dementia with Lewy bodies. Neuroreport 2002, 13, 2035–2039. [Google Scholar] [CrossRef]
- Che, Y.; Wang, J.-F.; Shao, L.; Young, T. Oxidative damage to RNA but not DNA in the hippocampus of patients with major mental illness. J. Psychiatry Neurosci 2010, 35, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Martinet, W.; De Meyer, G.R.Y.; Herman, A.G.; Kockx, M.M. RNA damage in human atherosclerosis: Pathophysiological significance and implications for gene expression studies. RNA Biol. 2005, 2, 4–7. [Google Scholar] [CrossRef]
- Warner, J.R. The economics of ribosome biosynthesis in yeast. Trends Biochem. Sci. 1999, 24, 437–440. [Google Scholar] [CrossRef]
- Ding, Q.; Markesbery, W.R.; Chen, Q.; Li, F.; Keller, J.N. Ribosome dysfunction is an early event in Alzheimer’s disease. J. Neurosci. 2005, 25, 9171–9175. [Google Scholar] [CrossRef]
- Ding, Q.; Markesbery, W.R.; Cecarini, V.; Keller, J.N. Decreased RNA, and increased RNA oxidation, in ribosomes from early Alzheimer’s disease. Neurochem. Res. 2006, 31, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Poole, L.B. The basics of thiols and cysteines in redox biology and chemistry. Free Radic. Biol. Med. 2015, 80, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Le Moan, N.; Clement, G.; Le Maout, S.; Tacnet, F.; Toledano, M.B. The Saccharomyces cerevisiae proteome of oxidized protein thiols: Contrasted functions for the thioredoxin and glutathione pathways. J. Biol. Chem. 2006, 281, 10420–10430. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.-M.; Duong, D.M.; Peng, J.; Jones, D.P. Protein Cysteines Map to Functional Networks According to Steady-state Level of Oxidation. J. Proteom. Bioinform 2011, 4, 196–209. [Google Scholar]
- Leichert, L.I.; Gehrke, F.; Gudiseva, H.V.; Blackwell, T.; Ilbert, M.; Walker, A.K.; Strahler, J.R.; Andrews, P.C.; Jakob, U. Quantifying changes in the thiol redox proteome upon oxidative stress in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 8197–8202. [Google Scholar] [CrossRef]
- Brandes, N.; Reichmann, D.; Tienson, H.; Leichert, L.I.; Jakob, U. Using quantitative redox proteomics to dissect the yeast redoxome. J. Biol. Chem. 2011, 286, 41893–41903. [Google Scholar] [CrossRef]
- Knoefler, D.; Thamsen, M.; Koniczek, M.; Niemuth, N.J.; Diederich, A.-K.; Jakob, U. Quantitative in vivo redox sensors uncover oxidative stress as an early event in life. Mol. Cell 2012, 47, 767–776. [Google Scholar] [CrossRef]
- Menger, K.E.; James, A.M.; Cochemé, H.M.; Harbour, M.E.; Chouchani, E.T.; Ding, S.; Fearnley, I.M.; Partridge, L.; Murphy, M.P. Fasting, but Not Aging, Dramatically Alters the Redox Status of Cysteine Residues on Proteins in Drosophila melanogaster. Cell Rep. 2015, 11, 1856–1865. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.; Weiss, S.J.; Levine, R.L. Methionine oxidation and reduction in proteins. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 901–905. [Google Scholar] [CrossRef]
- Caldwell, P.; Luk, D.C.; Weissbach, H.; Brot, N. Oxidation of the methionine residues of Escherichia coli ribosomal protein L12 decreases the protein’s biological activity. Proc. Natl. Acad. Sci. USA 1978, 75, 5349–5352. [Google Scholar] [CrossRef]
- Koteliansky, V.E.; Domogatsky, S.P.; Gudkov, A.T. Dimer state of protein L7/L12 and EF-G-dependent reactions of ribosomes. Eur. J. Biochem. 1978, 90, 319–323. [Google Scholar] [CrossRef]
- Berk, V.; Cate, J.H.D. Insights into protein biosynthesis from structures of bacterial ribosomes. Curr. Opin. Struct. Biol. 2007, 17, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.C.; Möller, W. Structure and function of the acidic ribosomal stalk proteins. Curr. Protein Pept. Sci. 2002, 3, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groitl, B.; Jakob, U. Thiol-based redox switches. Biochim. Biophys. Acta 2014, 1844, 1335–1343. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R.; Levine, R.L. Free radical-mediated oxidation of free amino acids and amino acid residues in proteins. Amino Acids 2003, 25, 207–218. [Google Scholar] [CrossRef]
- Nyström, T. Role of oxidative carbonylation in protein quality control and senescence. EMBO J. 2005, 24, 1311–1317. [Google Scholar] [CrossRef]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.-F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G.; Pizzimenti, S.; Daga, M.; Dianzani, C.; Arcaro, A.; Cetrangolo, G.P.; Giordano, G.; Cucci, M.A.; Graf, M.; Gentile, F. Lipid Peroxidation-Derived Aldehydes, 4-Hydroxynonenal and Malondialdehyde in Aging-Related Disorders. Antioxidants 2018, 7, 102. [Google Scholar] [CrossRef]
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein carbonylation as a major hallmark of oxidative damage: Update of analytical strategies. Mass Spectrom Rev. 2014, 33, 79–97. [Google Scholar] [CrossRef]
- Suzuki, Y.J.; Carini, M.; Butterfield, D.A. Protein carbonylation. Antioxid. Redox Signal. 2010, 12, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Madian, A.G.; Regnier, F.E. Proteomic identification of carbonylated proteins and their oxidation sites. J. Proteome Res. 2010, 9, 3766–3780. [Google Scholar] [CrossRef] [PubMed]
- Lemma-Gray, P.; Weintraub, S.T.; Carroll, C.A.; Musatov, A.; Robinson, N.C. Tryptophan 334 oxidation in bovine cytochrome c oxidase subunit I involves free radical migration. FEBS Lett. 2007, 581, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.W.; Fahy, E.; Murray, J.; Capaldi, R.A.; Ghosh, S.S. Oxidative post-translational modification of tryptophan residues in cardiac mitochondrial proteins. J. Biol. Chem. 2003, 278, 19587–19590. [Google Scholar] [CrossRef]
- Todorovski, T.; Fedorova, M.; Hoffmann, R. Mass spectrometric characterization of peptides containing different oxidized tryptophan residues. J. Mass Spectrom 2011, 46, 1030–1038. [Google Scholar] [CrossRef]
- Baraibar, M.A.; Ladouce, R.; Friguet, B. Proteomic quantification and identification of carbonylated proteins upon oxidative stress and during cellular aging. J. Proteom. 2013, 92, 63–70. [Google Scholar] [CrossRef]
- Lennicke, C.; Rahn, J.; Heimer, N.; Lichtenfels, R.; Wessjohann, L.A.; Seliger, B. Redox proteomics: Methods for the identification and enrichment of redox-modified proteins and their applications. Proteomics 2016, 16, 197–213. [Google Scholar] [CrossRef]
- Bollineni, R.C.; Hoffmann, R.; Fedorova, M. Proteome-wide profiling of carbonylated proteins and carbonylation sites in HeLa cells under mild oxidative stress conditions. Free Radic. Biol. Med. 2014, 68, 186–195. [Google Scholar] [CrossRef]
- Mirzaei, H.; Regnier, F. Creation of allotypic active sites during oxidative stress. J. Proteome Res. 2006, 5, 2159–2168. [Google Scholar] [CrossRef]
- Mirzaei, H.; Regnier, F. Protein-RNA cross-linking in the ribosomes of yeast under oxidative stress. J. Proteome Res. 2006, 5, 3249–3259. [Google Scholar] [CrossRef]
- Codreanu, S.G.; Zhang, B.; Sobecki, S.M.; Billheimer, D.D.; Liebler, D.C. Global analysis of protein damage by the lipid electrophile 4-hydroxy-2-nonenal. Mol. Cell Proteom. 2009, 8, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Kitabatake, M.; Sakata, T.; Miyata, A.; Ohno, M. A role for ubiquitin in the clearance of nonfunctional rRNAs. Genes Dev. 2009, 23, 963–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, K.; Kitabatake, M.; Sakata, T.; Ohno, M. 40S subunit dissociation and proteasome-dependent RNA degradation in nonfunctional 25S rRNA decay. EMBO J. 2012, 31, 2579–2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komander, D.; Rape, M. The ubiquitin code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef]
- Back, S.; Gorman, A.W.; Vogel, C.; Silva, G.M. Site-specific K63 ubiquitinomics provides insights into translation regulation under stress. J. Proteome Res. 2019, 18, 309–318. [Google Scholar] [CrossRef]
- Silva, G.M.; Finley, D.; Vogel, C. K63 polyubiquitination is a new modulator of the oxidative stress response. Nat. Struct. Mol. Biol. 2015, 22, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Manohar, S.; Jacob, S.; Wang, J.C.; Wiechecki, K.A.; Koh, H.W.; Simões, V.; Choi, H.; Vogel, C.; Silva, G.M. Polyubiquitin chains linked by lysine residue 48 (K48) selectively target oxidized proteins in vivo. Antioxid. Redox Signal. 2019. [Google Scholar] [CrossRef]
- Winterbourn, C.C. The biological chemistry of hydrogen peroxide. Meth. Enzymol. 2013, 528, 3–25. [Google Scholar]
- Chevion, M. A site-specific mechanism for free radical induced biological damage: The essential role of redox-active transition metals. Free Radic. Biol. Med. 1988, 5, 27–37. [Google Scholar] [CrossRef]
- Stadtman, E.R. Metal ion-catalyzed oxidation of proteins: Biochemical mechanism and biological consequences. Free Radic. Biol. Med. 1990, 9, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Shedlovskiy, D.; Zinskie, J.A.; Gardner, E.; Pestov, D.G.; Shcherbik, N. Endonucleolytic cleavage in the expansion segment 7 of 25S rRNA is an early marker of low-level oxidative stress in yeast. J. Biol. Chem. 2017, 292, 18469–18485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinskie, J.A.; Ghosh, A.; Trainor, B.M.; Shedlovskiy, D.; Pestov, D.G.; Shcherbik, N. Iron-dependent cleavage of ribosomal RNA during oxidative stress in the yeast Saccharomyces cerevisiae. J. Biol. Chem. 2018, 293, 14237–14248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athavale, S.S.; Petrov, A.S.; Hsiao, C.; Watkins, D.; Prickett, C.D.; Gossett, J.J.; Lie, L.; Bowman, J.C.; O’Neill, E.; Bernier, C.R.; et al. RNA folding and catalysis mediated by iron (II). PLoS ONE 2012, 7, e38024. [Google Scholar] [CrossRef] [PubMed]
- Bray, M.S.; Lenz, T.K.; Haynes, J.W.; Bowman, J.C.; Petrov, A.S.; Reddi, A.R.; Hud, N.V.; Williams, L.D.; Glass, J.B. Multiple prebiotic metals mediate translation. Proc. Natl. Acad. Sci. USA 2018, 115, 12164–12169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, D.J.R.; Merlot, A.M.; Huang, M.L.-H.; Bae, D.-H.; Jansson, P.J.; Sahni, S.; Kalinowski, D.S.; Richardson, D.R. Cellular iron uptake, trafficking and metabolism: Key molecules and mechanisms and their roles in disease. Biochim. Et Biophys. Acta (BBA) Mol. Cell Res. 2015, 1853, 1130–1144. [Google Scholar] [CrossRef] [Green Version]
- Tullius, T.D.; Dombroski, B.A. Hydroxyl radical “footprinting”: High-resolution information about DNA-protein contacts and application to lambda repressor and Cro protein. Proc. Natl. Acad. Sci. USA 1986, 83, 5469–5473. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Smith, M.A.; Zhu, X.; Baus, D.; Merrick, W.C.; Tartakoff, A.M.; Hattier, T.; Harris, P.L.; Siedlak, S.L.; Fujioka, H.; et al. Ribosomal RNA in Alzheimer disease is oxidized by bound redox-active iron. J. Biol. Chem. 2005, 280, 20978–20986. [Google Scholar] [CrossRef]
- Emerit, J.; Beaumont, C.; Trivin, F. Iron metabolism, free radicals, and oxidative injury. Biomed. Pharmacother. 2001, 55, 333–339. [Google Scholar] [CrossRef]
- Winter, D.; Polacek, N.; Halama, I.; Streicher, B.; Barta, A. Lead-catalysed specific cleavage of ribosomal RNAs. Nucleic Acids Res. 1997, 25, 1817–1824. [Google Scholar] [CrossRef] [Green Version]
- Kournoutou, G.G.; Giannopoulou, P.C.; Sazakli, E.; Leotsinidis, M.; Kalpaxis, D.L. Oxidative damage of 18S and 5S ribosomal RNA in digestive gland of mussels exposed to trace metals. Aquat. Toxicol. 2017, 192, 136–147. [Google Scholar] [CrossRef]
- Daglas, M.; Adlard, P.A. The Involvement of Iron in Traumatic Brain Injury and Neurodegenerative Disease. Front. Neurosci. 2018, 12, 981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kell, D.B. Towards a unifying, systems biology understanding of large-scale cellular death and destruction caused by poorly liganded iron: Parkinson’s, Huntington’s, Alzheimer’s, prions, bactericides, chemical toxicology and others as examples. Arch. Toxicol. 2010, 84, 825–889. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Dusek, P.; Roos, P.M.; Litwin, T.; Schneider, S.A.; Flaten, T.P.; Aaseth, J. The neurotoxicity of iron, copper and manganese in Parkinson’s and Wilson’s diseases. J. Trace Elem. Med. Biol. 2015, 31, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Limoncelli, K.A.; Merrikh, C.N.; Moore, M.J. ASC1 and RPS3: New actors in 18S nonfunctional rRNA decay. RNA 2017, 23, 1946–1960. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Li, S.; Kato, M.; Ikeuchi, K.; Ichimura, A.; Matsuo, Y.; Inada, T. Sequential Ubiquitination of Ribosomal Protein uS3 Triggers the Degradation of Non-functional 18S rRNA. Cell Rep. 2019, 26, 3400–3415.e7. [Google Scholar] [CrossRef] [PubMed]
- Kraft, C.; Deplazes, A.; Sohrmann, M.; Peter, M. Mature ribosomes are selectively degraded upon starvation by an autophagy pathway requiring the Ubp3p/Bre5p ubiquitin protease. Nat. Cell Biol. 2008, 10, 602–610. [Google Scholar] [CrossRef]
- Higgins, R.; Gendron, J.M.; Rising, L.; Mak, R.; Webb, K.; Kaiser, S.E.; Zuzow, N.; Riviere, P.; Yang, B.; Fenech, E.; et al. The Unfolded Protein Response Triggers Site-Specific Regulatory Ubiquitylation of 40S Ribosomal Proteins. Mol. Cell 2015, 59, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Chong, W.C.; Shastri, M.D.; Eri, R. Endoplasmic Reticulum Stress and Oxidative Stress: A Vicious Nexus Implicated in Bowel Disease Pathophysiology. Int. J. Mol. Sci. 2017, 18, 771. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double-edged sword? Antioxid. Redox Signal. 2007, 9, 2277–2293. [Google Scholar] [CrossRef]
- Simsek, D.; Tiu, G.C.; Flynn, R.A.; Byeon, G.W.; Leppek, K.; Xu, A.F.; Chang, H.Y.; Barna, M. The Mammalian Ribo-interactome Reveals Ribosome Functional Diversity and Heterogeneity. Cell 2017, 169, 1051–1065.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herr, C.Q.; Hausinger, R.P. Amazing Diversity in Biochemical Roles of Fe(II)/2-Oxoglutarate Oxygenases. Trends Biochem. Sci. 2018, 43, 517–532. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.; Wolf, A.; Feng, T.; Ho, C.-H.; Sekirnik, R.; Zayer, A.; Granatino, N.; Cockman, M.E.; Loenarz, C.; Loik, N.D.; et al. Oxygenase-catalyzed ribosome hydroxylation occurs in prokaryotes and humans. Nat. Chem. Biol. 2012, 8, 960–962. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.J.; Acevedo, J.M.; Loenarz, C.; Galagovsky, D.; Liu-Yi, P.; Pérez-Pepe, M.; Thalhammer, A.; Sekirnik, R.; Ge, W.; Melani, M.; et al. Sudestada1, a Drosophila ribosomal prolyl-hydroxylase required for mRNA translation, cell homeostasis, and organ growth. Proc. Natl. Acad. Sci. USA 2014, 111, 4025–4030. [Google Scholar] [CrossRef] [PubMed]
- Loenarz, C.; Sekirnik, R.; Thalhammer, A.; Ge, W.; Spivakovsky, E.; Mackeen, M.M.; McDonough, M.A.; Cockman, M.E.; Kessler, B.M.; Ratcliffe, P.J.; et al. Hydroxylation of the eukaryotic ribosomal decoding center affects translational accuracy. Proc. Natl. Acad. Sci. USA 2014, 111, 4019–4024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, R.S.; Liu-Yi, P.; Formenti, F.; Ge, W.; Sekirnik, R.; Fischer, R.; Adam, J.; Pollard, P.J.; Wolf, A.; Thalhammer, A.; et al. OGFOD1 catalyzes prolyl hydroxylation of RPS23 and is involved in translation control and stress granule formation. Proc. Natl. Acad. Sci. USA 2014, 111, 4031–4036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, S.; Sakai, Y.; Ishiguro, K.; Suzuki, T. Biogenesis and iron-dependency of ribosomal RNA hydroxylation. Nucleic Acids Res. 2017, 45, 12974–12986. [Google Scholar] [CrossRef] [Green Version]
- Havelund, J.F.; Giessing, A.M.B.; Hansen, T.; Rasmussen, A.; Scott, L.G.; Kirpekar, F. Identification of 5-hydroxycytidine at position 2501 concludes characterization of modified nucleotides in E. coli 23S rRNA. J. Mol. Biol. 2011, 411, 529–536. [Google Scholar] [CrossRef]
- Martinez, S.; Hausinger, R.P. Catalytic Mechanisms of Fe(II)- and 2-Oxoglutarate-dependent Oxygenases. J. Biol. Chem. 2015, 290, 20702–20711. [Google Scholar] [CrossRef] [Green Version]
- Houge, G.; Døskeland, S.O.; Bøe, R.; Lanotte, M. Selective cleavage of 28S rRNA variable regions V3 and V13 in myeloid leukemia cell apoptosis. FEBS Lett. 1993, 315, 16–20. [Google Scholar] [CrossRef] [Green Version]
- Mroczek, S.; Kufel, J. Apoptotic signals induce specific degradation of ribosomal RNA in yeast. Nucleic Acids Res. 2008, 36, 2874–2888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrone, G.G.; Tan, S.-X.; Dawes, I.W. Reactive oxygen species and yeast apoptosis. Biochim. Biophys. Acta 2008, 1783, 1354–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, K.D.; Noren Hooten, N.; Trzeciak, A.R.; Evans, M.K. Markers of oxidant stress that are clinically relevant in aging and age-related disease. Mech. Ageing Dev. 2013, 134, 139–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desnues, B.; Cuny, C.; Grégori, G.; Dukan, S.; Aguilaniu, H.; Nyström, T. Differential oxidative damage and expression of stress defence regulons in culturable and non-culturable Escherichia coli cells. EMBO Rep. 2003, 4, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Orgel, L.E. The maintenance of the accuracy of protein synthesis and its relevance to ageing. Proc. Natl. Acad. Sci. USA 1963, 49, 517–521. [Google Scholar] [CrossRef]
- Hayakawa, H.; Hofer, A.; Thelander, L.; Kitajima, S.; Cai, Y.; Oshiro, S.; Yakushiji, H.; Nakabeppu, Y.; Kuwano, M.; Sekiguchi, M. Metabolic fate of oxidized guanine ribonucleotides in mammalian cells. Biochemistry 1999, 38, 3610–3614. [Google Scholar] [CrossRef]
- Ishibashi, T.; Hayakawa, H.; Ito, R.; Miyazawa, M.; Yamagata, Y.; Sekiguchi, M. Mammalian enzymes for preventing transcriptional errors caused by oxidative damage. Nucleic Acids Res. 2005, 33, 3779–3784. [Google Scholar] [CrossRef] [Green Version]
- Taddei, F.; Hayakawa, H.; Bouton, M.; Cirinesi, A.; Matic, I.; Sekiguchi, M.; Radman, M. Counteraction by MutT protein of transcriptional errors caused by oxidative damage. Science 1997, 278, 128–130. [Google Scholar] [CrossRef]
- Takagi, Y.; Setoyama, D.; Ito, R.; Kamiya, H.; Yamagata, Y.; Sekiguchi, M. Human MTH3 (NUDT18) protein hydrolyzes oxidized forms of guanosine and deoxyguanosine diphosphates: Comparison with MTH1 and MTH2. J. Biol. Chem. 2012, 287, 21541–21549. [Google Scholar] [CrossRef]
- Shafirovich, V.; Geacintov, N.E. Removal of oxidatively generated DNA damage by overlapping repair pathways. Free Radic. Biol. Med. 2017, 107, 53–61. [Google Scholar] [CrossRef]
- Simms, C.L.; Yan, L.L.; Zaher, H.S. Ribosome Collision Is Critical for Quality Control during No-Go Decay. Mol. Cell 2017, 68, 361–373.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulk, A.; Liiv, A.; Peil, L.; Maiväli, U.; Nierhaus, K.; Remme, J. Ribosome reactivation by replacement of damaged proteins. Mol. Microbiol. 2010, 75, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Aas, P.A.; Otterlei, M.; Falnes, P.O.; Vågbø, C.B.; Skorpen, F.; Akbari, M.; Sundheim, O.; Bjørås, M.; Slupphaug, G.; Seeberg, E.; et al. Human and bacterial oxidative demethylases repair alkylation damage in both RNA and DNA. Nature 2003, 421, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.E.; LaRiviere, F.J.; Merrikh, C.N.; Moore, M.J. A convergence of rRNA and mRNA quality control pathways revealed by mechanistic analysis of nonfunctional rRNA decay. Mol. Cell 2009, 34, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Jacob, A.I.; Köhrer, C.; Davies, B.W.; RajBhandary, U.L.; Walker, G.C. Conserved bacterial RNase YbeY plays key roles in 70S ribosome quality control and 16S rRNA maturation. Mol. Cell 2013, 49, 427–438. [Google Scholar] [CrossRef]
- Pestov, D.G.; Shcherbik, N. Rapid cytoplasmic turnover of yeast ribosomes in response to rapamycin inhibition of TOR. Mol. Cell. Biol. 2012, 32, 2135–2144. [Google Scholar] [CrossRef]
- Sulthana, S.; Basturea, G.N.; Deutscher, M.P. Elucidation of pathways of ribosomal RNA degradation: An essential role for RNase, E. RNA 2016, 22, 1163–1171. [Google Scholar] [CrossRef]
- Hayakawa, H.; Fujikane, A.; Ito, R.; Matsumoto, M.; Nakayama, K.I.; Sekiguchi, M. Human proteins that specifically bind to 8-oxoguanine-containing RNA and their responses to oxidative stress. Biochem. Biophys. Res. Commun. 2010, 403, 220–224. [Google Scholar] [CrossRef]
- Ishii, T.; Hayakawa, H.; Igawa, T.; Sekiguchi, T.; Sekiguchi, M. Specific binding of PCBP1 to heavily oxidized RNA to induce cell death. Proc. Natl. Acad. Sci. USA 2018, 115, 6715–6720. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, H.; Kuwano, M.; Sekiguchi, M. Specific binding of 8-oxoguanine-containing RNA to polynucleotide phosphorylase protein. Biochemistry 2001, 40, 9977–9982. [Google Scholar] [CrossRef]
- Wu, J.; Jiang, Z.; Liu, M.; Gong, X.; Wu, S.; Burns, C.M.; Li, Z. Polynucleotide phosphorylase protects Escherichia coli against oxidative stress. Biochemistry 2009, 48, 2012–2020. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, Z. Human polynucleotide phosphorylase reduces oxidative RNA damage and protects HeLa cell against oxidative stress. Biochem. Biophys. Res. Commun. 2008, 372, 288–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, Y.; Dhaliwal, J.S.; Adjibade, P.; Uniacke, J.; Mazroui, R.; Zerges, W. Localized control of oxidized RNA. J. Cell Sci. 2015, 128, 4210–4219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berquist, B.R.; McNeill, D.R.; Wilson, D.M. Characterization of abasic endonuclease activity of human Ape1 on alternative substrates, as well as effects of ATP and sequence context on AP site incision. J. Mol. Biol. 2008, 379, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Vascotto, C.; Fantini, D.; Romanello, M.; Cesaratto, L.; Deganuto, M.; Leonardi, A.; Radicella, J.P.; Kelley, M.R.; D’Ambrosio, C.; Scaloni, A.; et al. APE1/Ref-1 interacts with NPM1 within nucleoli and plays a role in the rRNA quality control process. Mol. Cell. Biol. 2009, 29, 1834–1854. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Dimayuga, E.; Markesbery, W.R.; Keller, J.N. Proteasome inhibition increases DNA and RNA oxidation in astrocyte and neuron cultures. J. Neurochem. 2004, 91, 1211–1218. [Google Scholar] [CrossRef]
- An, H.; Harper, J.W. Ribosome Abundance Control Via the Ubiquitin-Proteasome System and Autophagy. J. Mol. Biol. 2019. [Google Scholar] [CrossRef]
- An, H.; Harper, J.W. Systematic analysis of ribophagy in human cells reveals bystander flux during selective autophagy. Nat. Cell Biol. 2018, 20, 135–143. [Google Scholar] [CrossRef]
- Huang, H.; Kawamata, T.; Horie, T.; Tsugawa, H.; Nakayama, Y.; Ohsumi, Y.; Fukusaki, E. Bulk RNA degradation by nitrogen starvation-induced autophagy in yeast. EMBO J. 2015, 34, 154–168. [Google Scholar] [CrossRef]
- Ossareh-Nazari, B.; Niño, C.A.; Bengtson, M.H.; Lee, J.-W.; Joazeiro, C.A.P.; Dargemont, C. Ubiquitylation by the Ltn1 E3 ligase protects 60S ribosomes from starvation-induced selective autophagy. J. Cell Biol. 2014, 204, 909–917. [Google Scholar] [CrossRef]
- Dinman, J.D. Pathways to specialized ribosomes: The Brussels lecture. J. Mol. Biol. 2016, 428, 2186–2194. [Google Scholar] [CrossRef] [PubMed]
- Genuth, N.R.; Barna, M. The discovery of ribosome heterogeneity and its implications for gene regulation and organismal life. Mol. Cell 2018, 71, 364–374. [Google Scholar] [CrossRef] [PubMed]
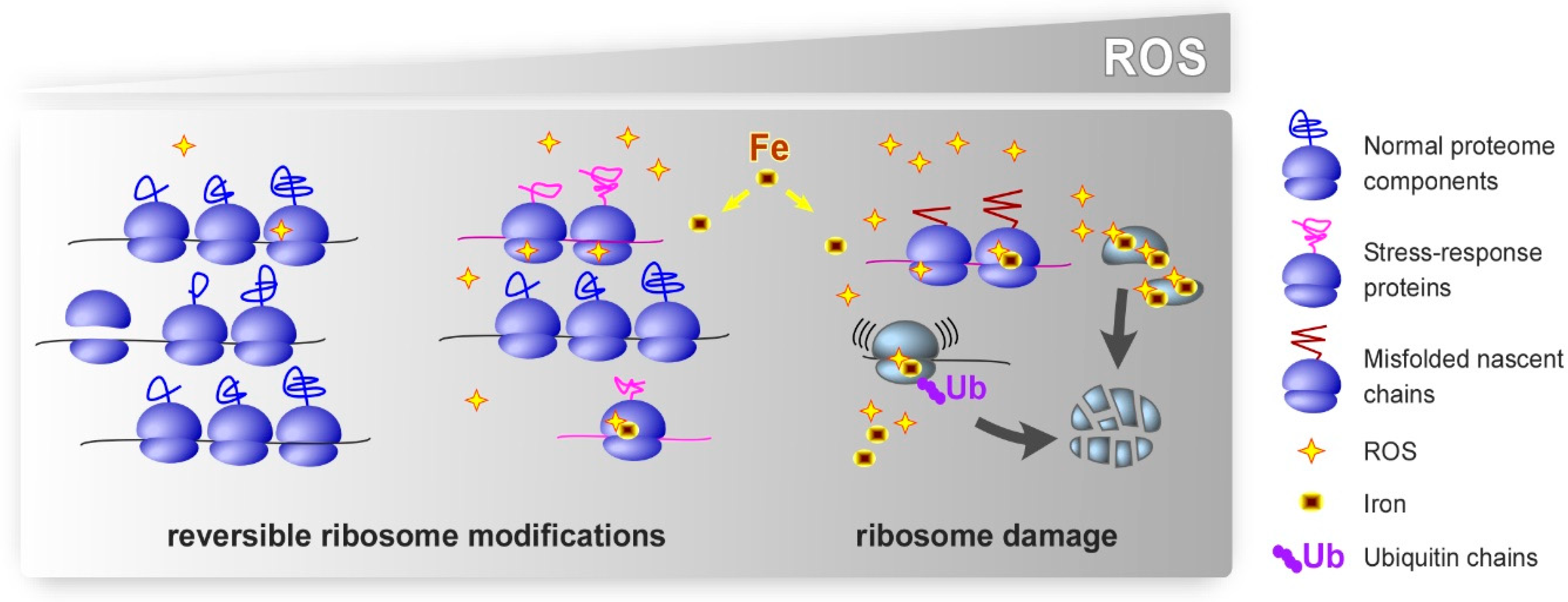
Figure 1.
Putative effects of oxidative stress on ribosomes. Cellular oxidative defenses neutralize excess ROS under basal conditions, which permits normal translation. We propose that low-level oxidative stress leads to largely reversible modifications in rRNA and r-proteins. These modifications could potentially promote selective translation of stress-response proteins and facilitate adaptive cellular responses. Strong or chronic oxidative stress inflicts excessive damage to ribosomes. Damaged ribosomes may be prone to making errors, become stalled during translation, or synthesize polypeptides that fail to fold correctly. In addition to ROS, intracellular iron contributes to the effects of oxidants by binding to specific sites on ribosomes, where it promotes localized redox reactions. Ubiquitination of r-proteins, one function of which appears to target dysfunctional ribosomes for degradation, has emerged as a prominent type of the secondary modifications of ribosomes under oxidative stress conditions.
Figure 1.
Putative effects of oxidative stress on ribosomes. Cellular oxidative defenses neutralize excess ROS under basal conditions, which permits normal translation. We propose that low-level oxidative stress leads to largely reversible modifications in rRNA and r-proteins. These modifications could potentially promote selective translation of stress-response proteins and facilitate adaptive cellular responses. Strong or chronic oxidative stress inflicts excessive damage to ribosomes. Damaged ribosomes may be prone to making errors, become stalled during translation, or synthesize polypeptides that fail to fold correctly. In addition to ROS, intracellular iron contributes to the effects of oxidants by binding to specific sites on ribosomes, where it promotes localized redox reactions. Ubiquitination of r-proteins, one function of which appears to target dysfunctional ribosomes for degradation, has emerged as a prominent type of the secondary modifications of ribosomes under oxidative stress conditions.


{kind=link}
Table 1.
Ribosomal RNA and protein modifications induced by oxidative stress.
| Type of Modification | Target | Organism | Reference |
|---|---|---|---|
| Guanine base | rRNA | Escherichia coli | [40,51] |
| oxidation | Mytilus galloprovincialis | [111] | |
| Rattus norvegicus | [50] | ||
| Homo sapiens | [61,62,108] | ||
| Strand scission | rRNA | Escherichia coli | [40] |
| Saccharomyces cerevisiae | [102,103,132] | ||
| Homo sapiens | [131] | ||
| rRNA-protein cross-links | rRNA, r-proteins | Saccharomyces cerevisiae | [91] |
| Met oxidation | r-proteins (LSU) | Escherichia coli | [71,72] |
| Cys oxidation | r-proteins (LSU + SSU) | Escherichia coli | [66] |
| Saccharomyces cerevisiae | [9,64,67] | ||
| Caenorhabditis elegans | [68] | ||
| Drosophila melanogaster | [69] | ||
| Homo sapiens (HT-29 cells) | [65] | ||
| Carbonylation | r-proteins (LSU + SSU) | Saccharomyces cerevisiae | [90,91] |
| r-proteins (LSU) | Homo sapiens (HeLa cells) | [89] | |
| Adduct formation | r-proteins (LSU + SSU) | Homo sapiens (RKO cells) | [92] |
| Ubiquitination | r-proteins (LSU + SSU) | Saccharomyces cerevisiae | [94,118] |
| r-proteins (SSU) | Saccharomyces cerevisiae | [116,117] | |
| Homo sapiens (HCT116 cells) | [119] | ||
| Ubiquitination | r-proteins (LSU + SSU) | Saccharomyces cerevisiae | [96,97] |
| (K63-Ub chains) | Mus musculus (HT22 cells) | [97] | |
| Ubiquitination (K48-Ub chains) | r-proteins (LSU + SSU) | Saccharomyces cerevisiae | [98] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shcherbik, N.; Pestov, D.G. The Impact of Oxidative Stress on Ribosomes: From Injury to Regulation. Cells 2019, 8, 1379. https://doi.org/10.3390/cells8111379
AMA Style
Shcherbik N, Pestov DG. The Impact of Oxidative Stress on Ribosomes: From Injury to Regulation. Cells. 2019; 8(11):1379. https://doi.org/10.3390/cells8111379
Chicago/Turabian StyleShcherbik, Natalia, and Dimitri G. Pestov. 2019. "The Impact of Oxidative Stress on Ribosomes: From Injury to Regulation" Cells 8, no. 11: 1379. https://doi.org/10.3390/cells8111379
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.