Immunolocalization of Keratan Sulfate in Rat Spinal Tissues Using the Keratanase Generated BKS-1(+) Neoepitope: Correlation of Expression Patterns with the Class II SLRPs, Lumican and Keratocan


Abstract
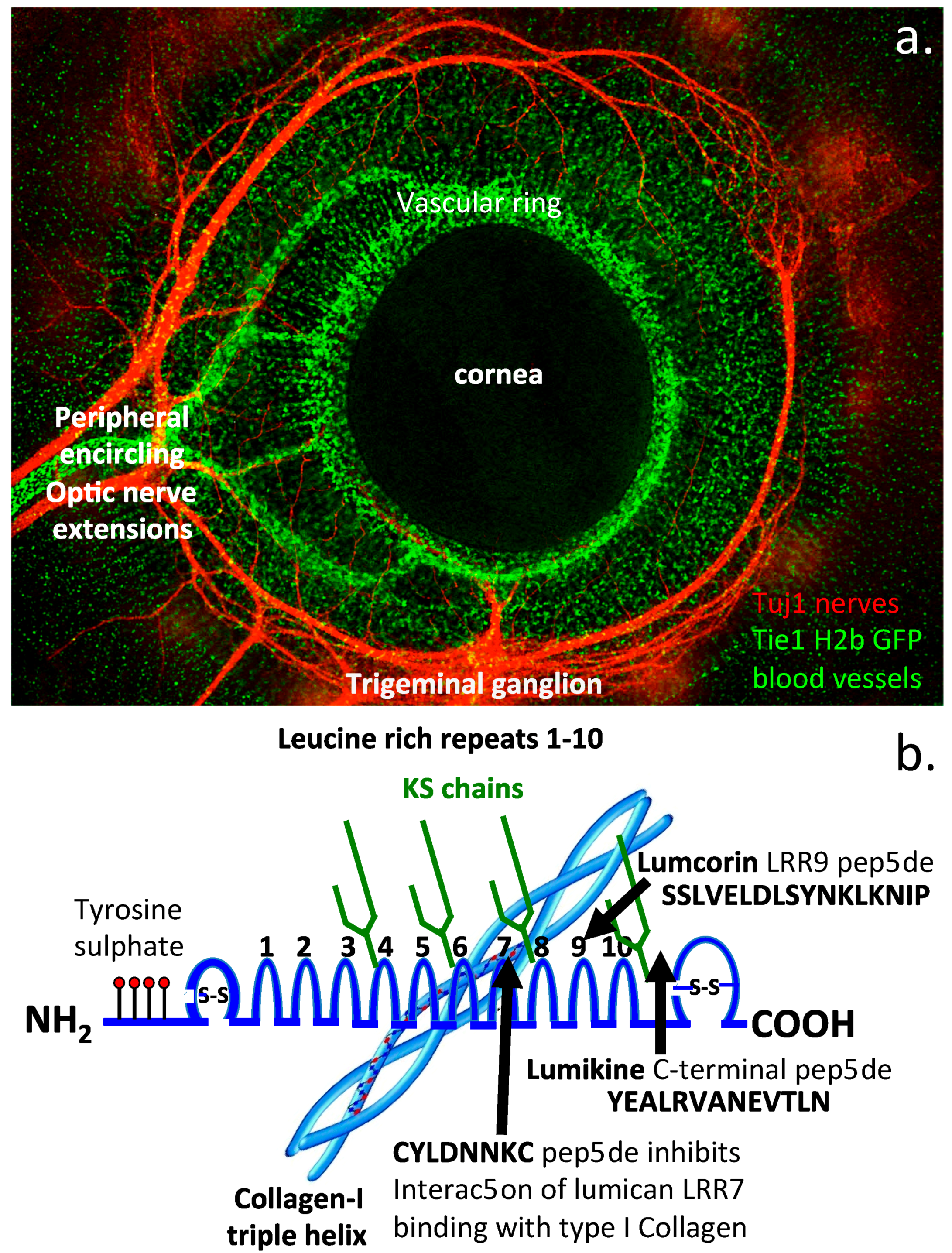
:1. Introduction
2. Materials and Methods
2.1. Tissues
2.2. Antibodies

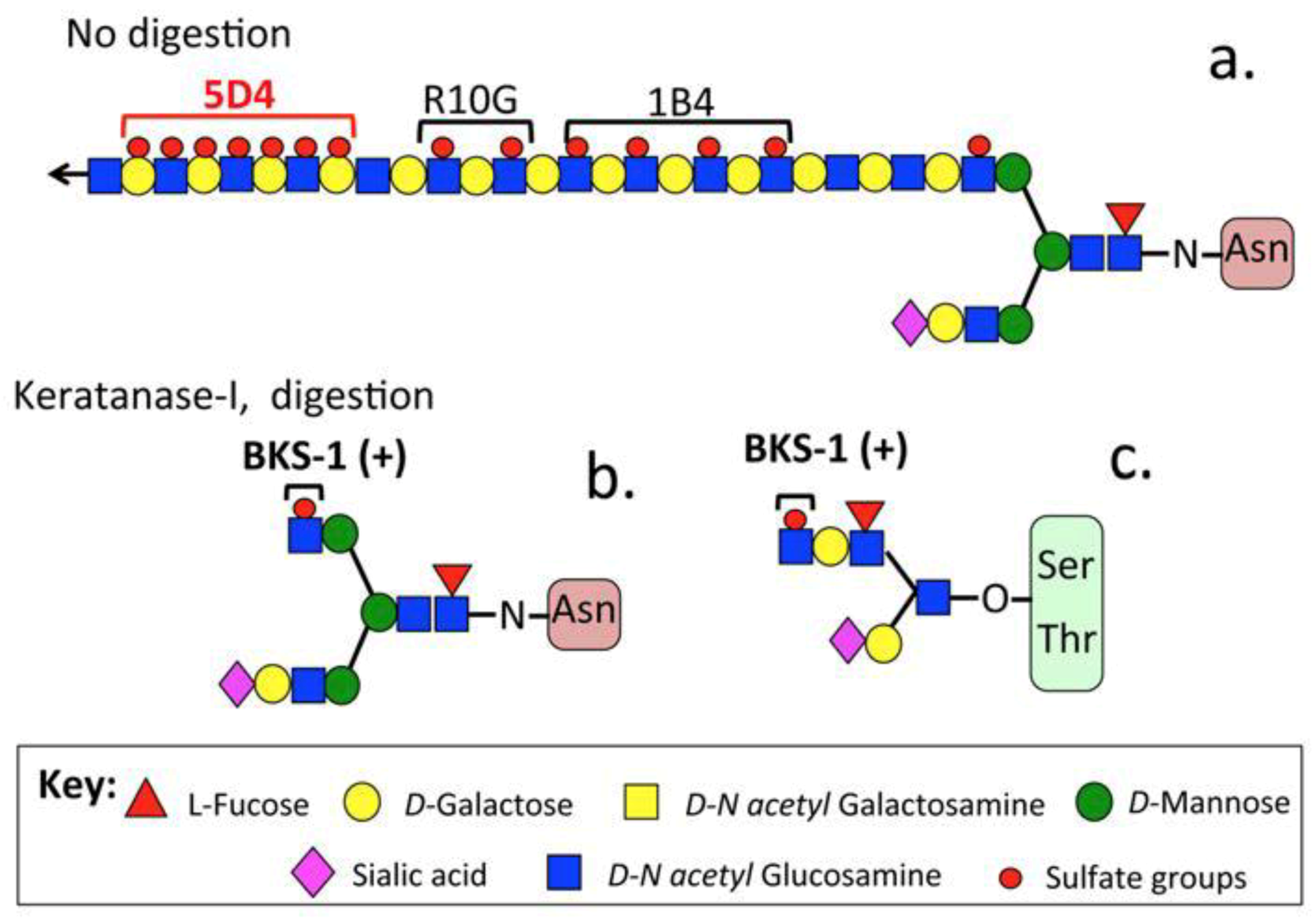{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody, Ab Class (Immunizing Antigen Used) | Source | Specificity | Reference |
|---|---|---|---|
| LUM-1, Mouse IgG monoclonal (lumican core protein) | Bridgeen Kerr, Cardiff University, UK | Mouse monoclonal IgG to 51 kDa core protein | [26] |
| KER-1, Mouse IgG monoclonal (keratocan core protein) | Bridgeen Kerr, Cardiff University, UK | Mouse monoclonal IgG to 38 kDa core protein | [26] |
| PR 353, Rabbit polyclonal (lumican C-terminal peptide LRVANEVTLN) | Peter Roughley, McGill University, Canada | Rabbit polyclonal Ab identifies LRVANEVTLN C-terminal peptide in lumican species | [17] |
| KTN, Rabbit polyclonal Ab, (C-terminal amino acids 227–257 of keratocan) | Abcam (cat # ab113115), UK | Rabbit polyclonal Ab to 38 kDa keratocan core protein | [27] |
| B9 Mouse monoclonal IgG (lumican core protein) | Santa Cruz (cat # sc-166871), USA | Rabbit monoclonal to 51 kDa clumican ore protein | [28,29] |
| BKS-1(+) Mouse monoclonal IgG (keratanase digested KS) | Bridgeen Kerr, Cardiff University, UK | Galactosamine-6-sulfate-galactose disaccharide in KS linkage region | [26] |
2.3. Histology
2.4. Immunohistochemistry
2.5. Extraction of PGs from Rat and Human Tissues
2.6. Chondroitinase ABC and Keratanase-I Digestion of Proteoglycan Samples
2.7. Lithium Dodecyl Sulfate PAGE and Western Blotting
3. Results
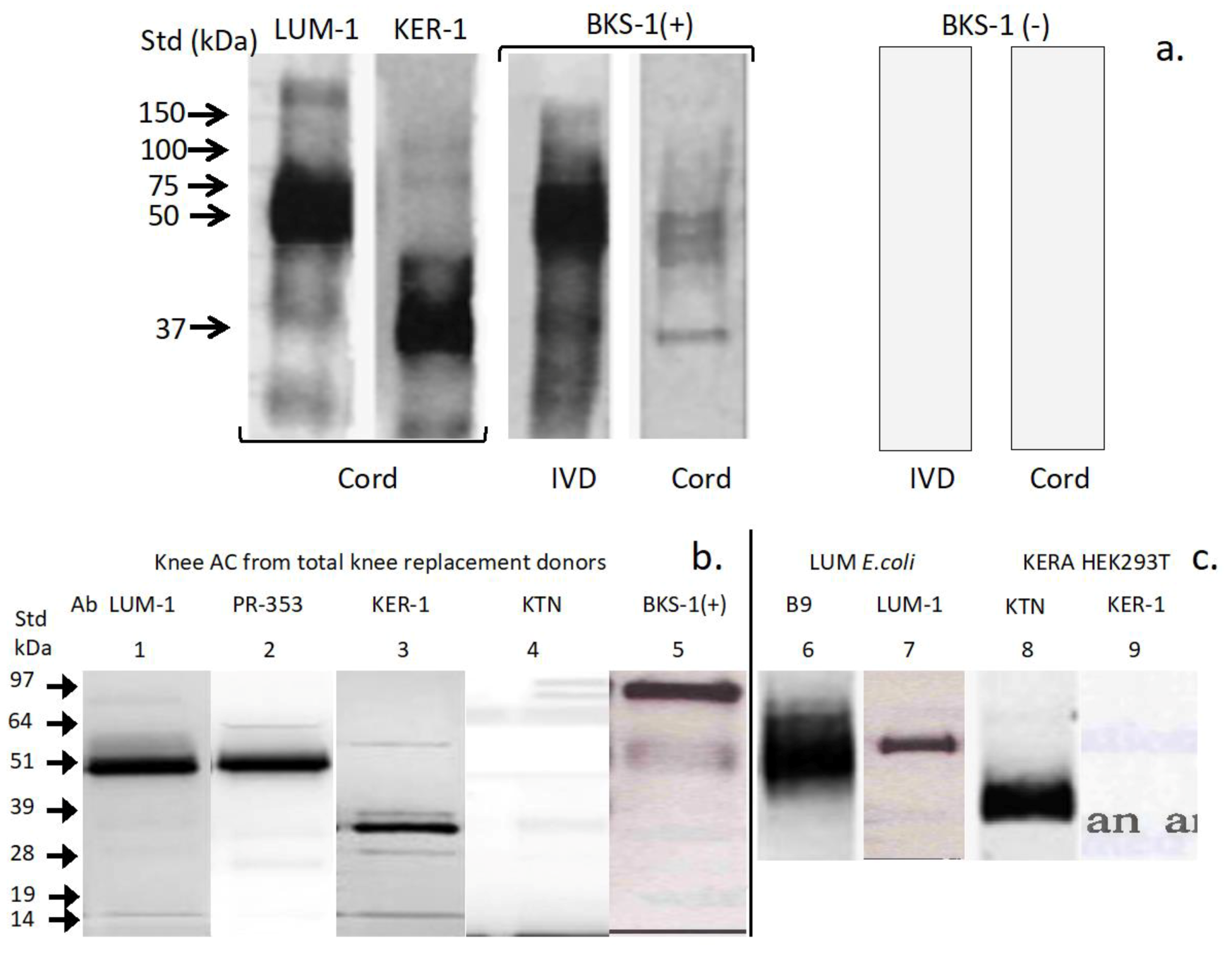
3.1. Western Blotting
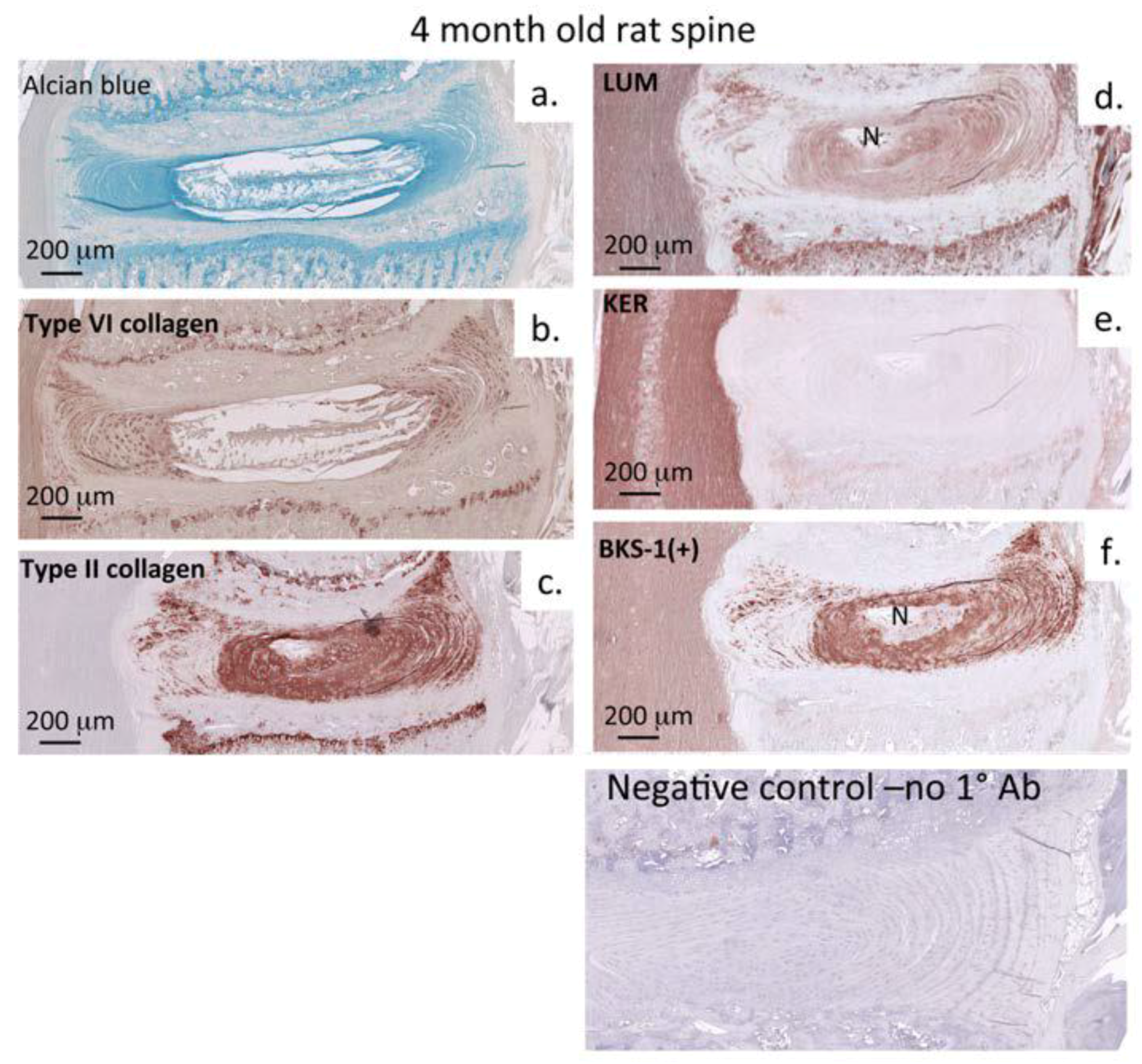
3.2. Immunohistochemistry: Adult Rat Spinal Tissue
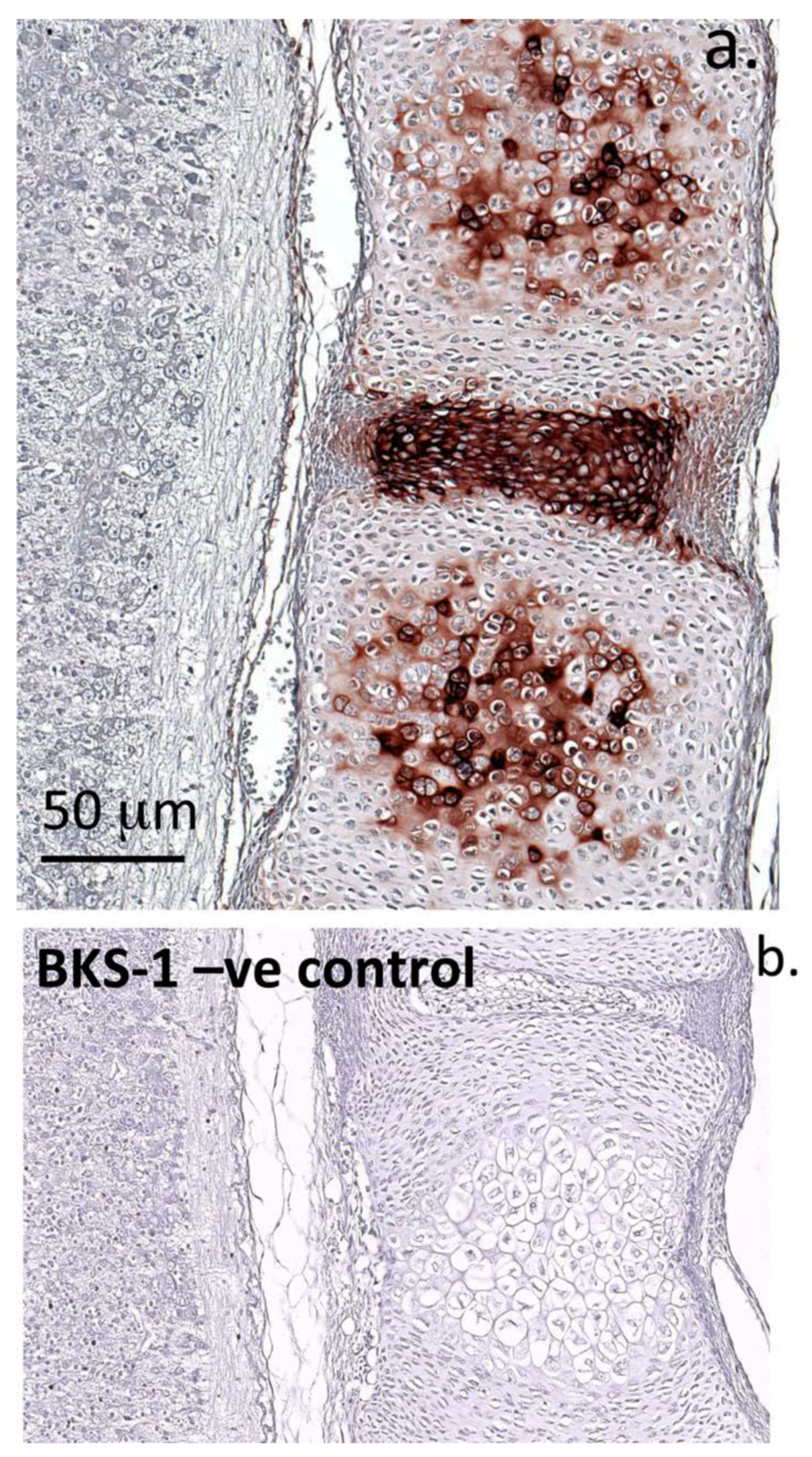
3.3. Immunohistochemistry: Fetal Rat Spinal Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Caterson, B.; Melrose, J. Keratan sulfate, a complex glycosaminoglycan with unique functional capability. Glycobiology 2018, 28, 182–206. [Google Scholar] [CrossRef] [PubMed]
- Funderburgh, J.L. MINI REVIEW Keratan sulfate: Structure, biosynthesis, and function. Glycobiology 2000, 10, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J. Keratan sulfate (KS)-proteoglycans and neuronal regulation in health and disease: The importance of KS -glycodynamics and interactive capability with neuroregulatory ligands. J. Neurochem. 2019, 149, 170–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caterson, B.; E Christner, J.; Baker, J.R. Identification of a monoclonal antibody that specifically recognizes corneal and skeletal keratan sulfate. Monoclonal antibodies to cartilage proteoglycan. J. Boil. Chem. 1983, 258, 8848–8854. [Google Scholar]
- Gealy, E.C.; Kerr, B.C.; Young, R.D.; Tudor, D.; Hayes, A.; Hughes, C.; Caterson, B.; Quantock, A.J.; Ralphs, J.R. Differential expression of the keratan sulphate proteoglycan, keratocan, during chick corneal embryogenesis. Histochem. Cell Boil. 2007, 128, 551–555. [Google Scholar] [CrossRef]
- Mehmet, H.; Scudder, P.; Tang, P.W.; Hounsell, E.F.; Feizi, T.; Caterson, B. The antigenic determinants recognized by three monoclonal antibodies to keratan sulphate involve sulphated hepta- or larger oligosaccharides of the poly (N-acetyllactosamine) series. JBIC J. Boil. Inorg. Chem. 1986, 157, 385–391. [Google Scholar] [CrossRef]
- Nakao, H.; Nagai, Y.; Kojima, A.; Toyoda, H.; Kawasaki, N.; Kawasaki, T. Binding specificity of R-10G and TRA-1-60/81, and substrate specificity of keratanase II studied with chemically synthesized oligosaccharides. Glycoconj. J. 2017, 34, 789–795. [Google Scholar] [CrossRef]
- Feizi, T.; A Childs, R.; Watanabe, K.; I Hakomori, S. Three types of blood group I specificity among monoclonal anti-I autoantibodies revealed by analogues of a branched erythrocyte glycolipid. J. Exp. Med. 1979, 149, 975–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, R.D.; Akama, T.O.; Liskova, P.; Ebenezer, N.D.; Allan, B.; Kerr, B.; Caterson, B.; Fukuda, M.N.; Quantock, A.J. Differential immunogold localisation of sulphated and unsulphated keratan sulphate proteoglycans in normal and macular dystrophy cornea using sulphation motif-specific antibodies. Histochem. Cell Boil. 2006, 127, 115–120. [Google Scholar] [CrossRef]
- Quantock, A.J.; Young, R.D.; Akama, T.O. Structural and biochemical aspects of keratan sulphate in the cornea. Cell. Mol. Life Sci. 2009, 67, 891–906. [Google Scholar] [CrossRef]
- Akhtar, S.; Kerr, B.C.; Hayes, A.; Hughes, C.; Meek, K.M.; Caterson, B. Immunochemical Localization of Keratan Sulfate Proteoglycans in Cornea, Sclera, and Limbus Using a Keratanase-Generated Neoepitope Monoclonal Antibody. Investig. Opthalmology Vis. Sci. 2008, 49, 2424–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Xia, K.; Lin, L.; Zhang, F.; Yu, Y.; Ange, K.S.; Han, X.; Edsinger, E.; Sohn, J.J.; Linhardt, R.J. Structural and Functional Components of the Skate Sensory Organ Ampullae of Lorenzini. ACS Chem. Boil. 2018, 13, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Josberger, E.E.; Hassanzadeh, P.; Deng, Y.; Sohn, J.; Rego, M.J.; Amemiya, C.T.; Rolandi, M. Proton conductivity in ampullae of Lorenzini jelly. Sci. Adv. 2016, 2, e1600112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivier, E.; Sorrell, J.M.; Ackerly, M.; Robertson, M.J.; A Rasmussen, R.; Levine, H.; Anderson, P. Developmental regulation of a mucinlike glycoprotein selectively expressed on natural killer cells. J. Exp. Med. 1993, 178, 2023–2033. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, J.-I.; Okada, K.; Shinohara, Y. Glycomics of human embryonic stem cells and human induced pluripotent stem cells. Glycoconj. J. 2016, 33, 707–715. [Google Scholar] [CrossRef]
- Conrad, A.H.; Zhang, Y.; Tasheva, E.S.; Conrad, G.W. Proteomic analysis of potential keratan sulfate, chondroitin sulfate A, and hyaluronic acid molecular interactions. Investig. Opthalmology Vis. Sci. 2010, 51, 4500–4515. [Google Scholar] [CrossRef] [Green Version]
- Melrose, J.; Fuller, E.S.; Roughley, P.J.; Smith, M.M.; Kerr, B.; Hughes, C.; Caterson, B.; Little, C.B. Fragmentation of decorin, biglycan, lumican and keratocan is elevated in degenerate human meniscus, knee and hip articular cartilages compared with age-matched macroscopically normal and control tissues. Arthritis Res. Ther. 2008, 10, R79. [Google Scholar] [CrossRef] [Green Version]
- Hayes, A.; Isaacs, M.D.; Hughes, C.; Caterson, B.; Ralphs, J.R. Collagen fibrillogenesis in the development of the annulus fibrosus of the intervertebral disc. Eur. Cell Mater. 2011, 22, 226–241. [Google Scholar] [CrossRef]
- Kotwal, R.; Harris, A.M.; Wright, A.E.; Hodgson, P.; Hughes, C.E.; Roberts, S.; Richardson, J.; Caterson, B.; Dent, C. Monoclonal antibodies recognising keratan sulphate (KS) proteoglycans, “KS-stubs” and KS sulphation motifs used in ELISA to quantify changes in proteoglycan metabolism in musculoskeletal diseases. J. Bone Jt. Surg. 2007, 91B, 292. [Google Scholar]
- Okayasu, K.; Ohtani, Y.; Takemura, T.; Uchibori, K.; Tamaoka, M.; Furuiye, M.; Miyazaki, Y.; Inase, N.; Yoshizawa, Y. Nonspecific interstitial pneumonia (NSIP) associated with anti-KS antibody: Differentiation from idiopathic NSIP. Intern. Med. 2009, 48, 1301–1306. [Google Scholar] [CrossRef] [Green Version]
- Yasogawa, Y.; Takano, Y.; Okayasu, I.; Kakita, A. The 5D4 antibody (anti-cyclin D1/D2) related antigen: Cytoplasmic staining is correlated to the progression of gastric cancer. Pathol. Int. 1998, 48, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Melrose, J.; Caterson, B.; Roughley, P.; Eisenstein, S.M.; Roberts, S. A comparative evaluation of the small leucine-rich proteoglycans of pathological human intervertebral discs. Eur. Spine J. 2012, 21, 154–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melrose, J.; Smith, S.M.; Fuller, E.S.; Young, A.A.; Roughley, P.J.; Dart, A.; Little, C.B. Biglycan and fibromodulin fragmentation correlates with temporal and spatial annular remodelling in experimentally injured ovine intervertebral discs. Eur. Spine J. 2007, 16, 2193–2205. [Google Scholar] [CrossRef] [Green Version]
- A Young, A.; Smith, M.M.; Smith, S.M.; A Cake, M.; Ghosh, P.; A Read, R.; Melrose, J.; Sonnabend, D.H.; Roughley, P.J.; Little, C.B. Regional assessment of articular cartilage gene expression and small proteoglycan metabolism in an animal model of osteoarthritis. Arthritis Res. Ther. 2005, 7, R852–R861. [Google Scholar] [CrossRef] [Green Version]
- Rees, S.; Waggett, A.; Kerr, B.; Probert, J.; Gealy, E.; Dent, C.; Caterson, B.; Hughes, C. Immunolocalisation and expression of keratocan in tendon. Osteoarthr. Cartil. 2009, 17, 276–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, B. Keratan sulphate metabolism in connective tissue proteoglycans. Ph.D. Thesis, Cardiff University, Cardiff, UK, 2005. [Google Scholar]
- Türker, E.; Garreis, F.; Khajavi, N.; Reinach, P.S.; Joshi, P.; Brockmann, T.; Lucius, A.; Ljubojevic, N.; Turan, E.; Cooper, D.; et al. Vascular Endothelial Growth Factor (VEGF) Induced Downstream Responses to Transient Receptor Potential Vanilloid 1 (TRPV1) and 3-Iodothyronamine (3-T1AM) in Human Corneal Keratocytes. Front. Endocrinol. 2018, 9, 670. [Google Scholar] [CrossRef] [Green Version]
- Brosseau, J.-P.; Liao, C.-P.; Wang, Y.; Ramani, V.; Vandergriff, T.; Lee, M.; Patel, A.; Ariizumi, K.; Le, L. NF1 heterozygosity fosters de novo tumorigenesis but impairs malignant transformation. Nat. Commun. 2018, 9, 5014. [Google Scholar] [CrossRef]
- Jana, S.; Zhang, H.; Lopaschuk, G.D.; Freed, D.H.; Sergi, C.; Kantor, P.F.; Oudit, G.Y.; Kassiri, Z. Disparate Remodeling of the Extracellular Matrix and Proteoglycans in Failing Pediatric Versus Adult Hearts. J. Am. Hear. Assoc. 2018, 7, e010427. [Google Scholar] [CrossRef] [Green Version]
- Melrose, J.; Smith, S.; Cake, M.; Read, R.; Whitelock, J. Comparative spatial and temporal localisation of perlecan, aggrecan and type I, II and IV collagen in the ovine meniscus: An ageing study. Histochem. Cell Boil. 2005, 124, 225–235. [Google Scholar] [CrossRef]
- Nikdin, H.; Olsson, M.-L.; Hultenby, K.; Sugars, R. Osteoadherin Accumulates in the Predentin towards the Mineralization Front in the Developing Tooth. PLoS ONE 2012, 7, e31525. [Google Scholar] [CrossRef] [Green Version]
- Ramstad, V.; Franzen, A.; Heinegård, D.; Wendel, M.; Reinholt, F. Ultrastructural Distribution of Osteoadherin in Rat Bone Shows a Pattern Similar to That of Bone Sialoprotein. Calcif. Tissue Int. 2003, 72, 57–64. [Google Scholar] [CrossRef]
- Rehn, A.P.; Cerny, R.; Sugars, R.; Kaukua, N.; Wendel, M. Osteoadherin is Upregulated by Mature Osteoblasts and Enhances Their In Vitro Differentiation and Mineralization. Calcif. Tissue Int. 2008, 82, 454–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommarin, Y.; Wendel, M.; Shen, Z.; Hellman, U.; Heinegård, D. Osteoadherin, a Cell-binding Keratan Sulfate Proteoglycan in Bone, Belongs to the Family of Leucine-rich Repeat Proteins of the Extracellular Matrix. J. Boil. Chem. 1998, 273, 16723–16729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugars, R.; Olsson, M.-L.; Marchner, S.; Hultenby, K.; Wendel, M. The glycosylation profile of osteoadherin alters during endochondral bone formation. Bone 2013, 53, 459–467. [Google Scholar] [CrossRef]
- Wendel, M.; Sommarin, Y.; Heinegård, D. Bone Matrix Proteins: Isolation and Characterization of a Novel Cell-binding Keratan Sulfate Proteoglycan (Osteoadherin) from Bovine Bone. J. Cell Boil. 1998, 141, 839–847. [Google Scholar] [CrossRef]
- Petersson, U.; Hultenby, K.; Wendel, M. Identification, distribution and expression of osteoadherin during tooth formation. Eur. J. Oral Sci. 2003, 111, 128–136. [Google Scholar] [CrossRef]
- Hildebrand, A.; Romaris, M.; Rasmussen, L.M.; Heinegård, D.; Twardzik, D.R.; A Border, W.; Ruoslahti, E. Interaction of the small interstitial proteoglycans biglycan, decorin and fibromodulin with transforming growth factor β. Biochem. J. 1994, 302, 527–534. [Google Scholar] [CrossRef]
- Chen, S.; Birk, D.E. The regulatory roles of small leucine-rich proteoglycans in extracellular matrix assembly. FEBS J. 2013, 280, 2120–2137. [Google Scholar] [CrossRef] [Green Version]
- I Inkinen, R.; Lammi, M.J.; Lehmonen, S.; Puustjärvi, K.; Kääpä, E.; I Tammi, M. Relative increase of biglycan and decorin and altered chondroitin sulfate epitopes in the degenerating human intervertebral disc. J. Rheumatol. 1998, 25, 506–514. [Google Scholar]
- Johnstone, B.; Markopoulos, M.; Neame, P.; Caterson, B. Identification and characterization of glycanated and non-glycanated forms of biglycan and decorin in the human intervertebral disc. Biochem. J. 1993, 292, 661–666. [Google Scholar] [CrossRef] [Green Version]
- Roughley, P.J.; White, R.J.; Magny, M.C.; Liu, J.; Pearce, R.H.; Mort, J.S. Non-proteoglycan forms of biglycan increase with age in human articular cartilage. Biochem. J. 1993, 295, 421–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theocharis, A.D.; Karamanos, N.K.; Papageorgakopoulou, N.; Tsiganos, C.P.; A Theocharis, D. Isolation and characterization of matrix proteoglycans from human nasal cartilage. Compositional and structural comparison between normal and scoliotic tissues. Biochim. et Biophys. Acta. 2002, 1569, 117–126. [Google Scholar] [CrossRef]
- Burton-Wurster, N.; Liu, W.; Matthews, G.L.; Lust, G.; Roughley, P.J.; Glant, T.T.; Cs-Szabó, G. TGF beta 1 and biglycan, decorin, and fibromodulin metabolism in canine cartilage. Osteoarthr. Cartil. 2003, 11, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Septier, D.; Hall, R.; Embery, G.; Goldberg, M. Immunoelectron Microscopic Visualization of Pro- and Secreted Forms of Decorin and Biglycan in the Predentin and During Dentin Formation in the Rat Incisor. Calcif. Tissue Int. 2001, 69, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Dunlevy, J.R.; Neame, P.J.; Vergnes, J.-P.; Hassell, J.R. Identification of theN-Linked Oligosaccharide Sites in Chick Corneal Lumican and Keratocan That Receive Keratan Sulfate. J. Boil. Chem. 1998, 273, 9615–9621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashida, Y.; Akama, T.O.; Beecher, N.; Lewis, P.; Young, R.D.; Meek, K.M.; Kerr, B.; Hughes, C.; Caterson, B.; Tanigami, A.; et al. Matrix morphogenesis in cornea is mediated by the modification of keratan sulfate by GlcNAc 6-O-sulfotransferase. Proc. Natl. Acad. Sci. USA 2006, 103, 13333–13338. [Google Scholar] [CrossRef] [Green Version]
- Rao, W.W.-Y.; Liu, C.-Y. Roles of lumican and keratocan on corneal transparency. Glycoconj. J. 2002, 19, 275–285. [Google Scholar]
- Dunlevy, J.R.; Beales, M.P.; Berryhill, B.L.; Cornuet, P.K.; Hassell, J.R. Expression of the Keratan Sulfate Proteoglycans Lumican, Keratocan and Osteoglycin/Mimecan During Chick Corneal Development. Exp. Eye Res. 2000, 70, 349–362. [Google Scholar] [CrossRef]
- Wiese, S.; Faissner, A. The role of extracellular matrix in spinal cord development. Exp. Neurol. 2015, 274, 90–99. [Google Scholar] [CrossRef]
- Gaudet, A.; Popovich, P.G. Extracellular matrix regulation of inflammation in the healthy and injured spinal cord. Exp. Neurol. 2014, 258, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Pietraszek-Gremplewicz, K.; Brézillon, S.; Perreau, C.; Malicka-Błaszkiewicz, M.; Maquart, F.-X.; Wegrowski, Y. Lumican – Derived Peptides Inhibit Melanoma Cell Growth and Migration. PLoS ONE 2013, 8, e76232. [Google Scholar]
- Pietraszek-Gremplewicz, K.; Chatron-Colliet, A.; Brézillon, S.; Perreau, C.; Jakubiak-Augustyn, A.; Krotkiewski, H.; Maquart, F.-X.; Wegrowski, Y. Lumican: A new inhibitor of matrix metalloproteinase-14 activity. FEBS Lett. 2014, 588, 4319–4324. [Google Scholar] [CrossRef] [Green Version]
- Brézillon, S.; Pietraszek-Gremplewicz, K.; Maquart, F.-X.; Wegrowski, Y. Lumican effects in the control of tumour progression and their links with metalloproteinases and integrins. FEBS J. 2013, 280, 2369–2381. [Google Scholar] [CrossRef] [PubMed]
- Stasiak, M.; Boncela, J.; Perreau, C.; Karamanou, K.; Chatron-Colliet, A.; Proult, I.; Przygodzka, P.; Chakravarti, S.; Maquart, F.-X.; Kowalska, M.A.; et al. Lumican Inhibits SNAIL-Induced Melanoma Cell Migration Specifically by Blocking MMP-14 Activity. PLoS ONE 2016, 11, e0150226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeltz, C.; Brézillon, S.; Perreau, C.; Ramont, L.; Maquart, F.-X.; Wegrowski, Y. Lumcorin: A leucine-rich repeat 9-derived peptide from human lumican inhibiting melanoma cell migration. FEBS Lett. 2009, 583, 3027–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricard-Blum, S.; Salza, R. Matricryptins and matrikines: Biologically active fragments of the extracellular matrix. Exp. Dermatol. 2014, 23, 457–463. [Google Scholar] [CrossRef]
- Sivaraman, K.; Shanthi, C. Matrikines for therapeutic and biomedical applications. Life Sci. 2018, 214, 22–33. [Google Scholar] [CrossRef]
- Maquart, F.-X.; Bellon, G.; Brassart-Pasco, S.; Monboisse, J. Matrikines in the regulation of extracellular matrix degradation. Biochim. 2005, 87, 353–360. [Google Scholar] [CrossRef]
- Ricard-Blum, S.; Vallet, S.D. Fragments generated upon extracellular matrix remodeling: Biological regulators and potential drugs. Matrix Boil. 2019, 75–76, 170–189. [Google Scholar] [CrossRef]
- Yamanaka, O.; Yuan, Y.; Coulson-Thomas, V.J.; Gesteira, T.F.; Call, M.K.; Zhang, Y.; Zhang, J.; Chang, S.-H.; Xie, C.; Liu, C.-Y.; et al. Lumican Binds ALK5 to Promote Epithelium Wound Healing. PLoS ONE 2013, 8, e82730. [Google Scholar] [CrossRef]
- Gesteira, T.F.; Coulson-Thomas, V.J.; Yuan, Y.; Zhang, J.; Nader, H.B.; Kao, W.W. Lumican Peptides: Rational Design Targeting ALK5/TGFBRI. Sci. Rep. 2017, 7, 42057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyers, A.; Yang, B.; Solakyildirim, K.; Yee, V.; Li, L.; Zhang, F.; Linhardt, R.J. Isolation of bovine corneal keratan sulfate and its growth factor and morphogen binding. FEBS J. 2013, 280, 2285–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrad, A.H.; Conrad, G.W. The keratocan gene is expressed in both ocular and non-ocular tissues during early chick development. Matrix Boil. 2003, 22, 323–337. [Google Scholar] [CrossRef]
- Schwend, T.; Deaton, R.J.; Zhang, Y.; Caterson, B.; Conrad, G.W. Corneal Sulfated Glycosaminoglycans and Their Effects on Trigeminal Nerve Growth Cone Behavior In Vitro: Roles for ECM in Cornea Innervation. Investig. Opthalmology Vis. Sci. 2012, 53, 8118–8137. [Google Scholar] [CrossRef]
- Schwend, T.; Lwigale, P.Y.; Conrad, G.W. Nerve repulsion by the lens and cornea during cornea innervation is dependent on Robo–Slit signaling and diminishes with neuron age. Dev. Boil. 2012, 363, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.L.; Tuszynski, M.H. Spinal Cord Injury Elicits Expression of Keratan Sulfate Proteoglycans by Macrophages, Reactive Microglia, and Oligodendrocyte Progenitors. J. Neurosci. 2002, 22, 4611–4624. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-Y.; Kim, H.; Kim, K.; Lee, H.; Lee, S.; Lee, D. Arhgap17, a RhoGTPase activating protein, regulates mucosal and epithelial barrier function in the mouse colon. Sci. Rep. 2016, 6, 26923. [Google Scholar] [CrossRef] [Green Version]
- Bacaj, T.; Wu, D.; Yang, X.; Morishita, W.; Zhou, P.; Xu, W.; Malenka, R.C.; Südhof, T.C. Synaptotagmin-1 and synaptotagmin-7 trigger synchronous and asynchronous phases of neurotransmitter release. Neuron 2013, 80, 947–959. [Google Scholar] [CrossRef] [Green Version]
- Südhof, T.C. A molecular machine for neurotransmitter release: Synaptotagmin and beyond. Nat. Med. 2013, 19, 1227–1231. [Google Scholar] [CrossRef]
- Richnau, N.; Aspenström, P. RICH, a Rho GTPase-activating Protein Domain-containing Protein Involved in Signaling by Cdc42 and Rac1. J. Boil. Chem. 2001, 276, 35060–35070. [Google Scholar] [CrossRef] [Green Version]
- Wells, C.D.; Fawcett, J.P.; Traweger, A.; Yamanaka, Y.; Goudreault, M.; Elder, K.; Kulkarni, S.; Gish, G.; Virag, C.; Lim, C.; et al. A Rich1/Amot complex regulates the Cdc42 GTPase and apical-polarity proteins in epithelial cells. Cell 2006, 125, 535–548. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Zhou, P.; Wang, A.L.; Wu, D.; Zhao, M.; Südhof, T.C.; Brunger, A.T. The primed SNARE–complexin–synaptotagmin complex for neuronal exocytosis. Nat. 2017, 548, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Schivell, A.E.; Batchelor, R.H.; Bajjalieh, S.M. Isoform-specific, Calcium-regulated Interaction of the Synaptic Vesicle Proteins SV2 and Synaptotagmin. J. Boil. Chem. 1996, 271, 27770–27775. [Google Scholar] [CrossRef] [Green Version]
- Plachez, C.; Andrews, W.; Liapi, A.; Knoell, B.; Drescher, U.; Mankoo, B.; Zhe, L.; Mambetisaeva, E.; Annan, A.; Bannister, L. Robos are required for the correct targeting of retinal ganglion cell axons in the visual pathway of the brain. Mol. Cell. Neurosci. 2008, 37, 719–730. [Google Scholar] [CrossRef]
- Snow, D.M.; Lemmon, V.P.; Carrino, D.A.; Caplan, A.; Silver, J. Sulfated proteoglycans in astroglial barriers inhibit neurite outgrowth in vitro. Exp. Neurol. 1990, 109, 111–130. [Google Scholar] [CrossRef]
- Kriegstein, A.R.; Alvarez-Buylla, A. The glial nature of embryonic and adult neural stem cells. Annu. Rev. Neurosci. 2009, 32, 149–184. [Google Scholar] [CrossRef] [Green Version]
- Song, I.; Dityatev, A. Crosstalk between glia, extracellular matrix and neurons. Brain Res. Bull. 2018, 136, 101–108. [Google Scholar] [CrossRef]
- Theocharidis, U.; Long, K.R.; Ffrench-Constant, C.; Faissner, A. Regulation of the neural stem cell compartment by extracellular matrix constituents. Prog. Brain Res. 2014, 214, 3–28. [Google Scholar]
- Hashimoto, H.; Ishino, Y.; Jiang, W.; Yoshimura, T.; Takeda-Uchimura, Y.; Uchimura, K.; Kadomatsu, K.; Ikenaka, K. Keratan Sulfate Regulates the Switch from Motor Neuron to Oligodendrocyte Generation During Development of the Mouse Spinal Cord. Neurochem. Res. 2016, 41, 450–462. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayes, A.J.; Melrose, J. Immunolocalization of Keratan Sulfate in Rat Spinal Tissues Using the Keratanase Generated BKS-1(+) Neoepitope: Correlation of Expression Patterns with the Class II SLRPs, Lumican and Keratocan. Cells 2020, 9, 826. https://doi.org/10.3390/cells9040826
Hayes AJ, Melrose J. Immunolocalization of Keratan Sulfate in Rat Spinal Tissues Using the Keratanase Generated BKS-1(+) Neoepitope: Correlation of Expression Patterns with the Class II SLRPs, Lumican and Keratocan. Cells. 2020; 9(4):826. https://doi.org/10.3390/cells9040826
Chicago/Turabian StyleHayes, Anthony J., and James Melrose. 2020. "Immunolocalization of Keratan Sulfate in Rat Spinal Tissues Using the Keratanase Generated BKS-1(+) Neoepitope: Correlation of Expression Patterns with the Class II SLRPs, Lumican and Keratocan" Cells 9, no. 4: 826. https://doi.org/10.3390/cells9040826