Formation of Aberrant Myotubes by Myoblasts Lacking Myosin VI Is Associated with Alterations in the Cytoskeleton Organization, Myoblast Adhesion and Fusion

 , and
, and 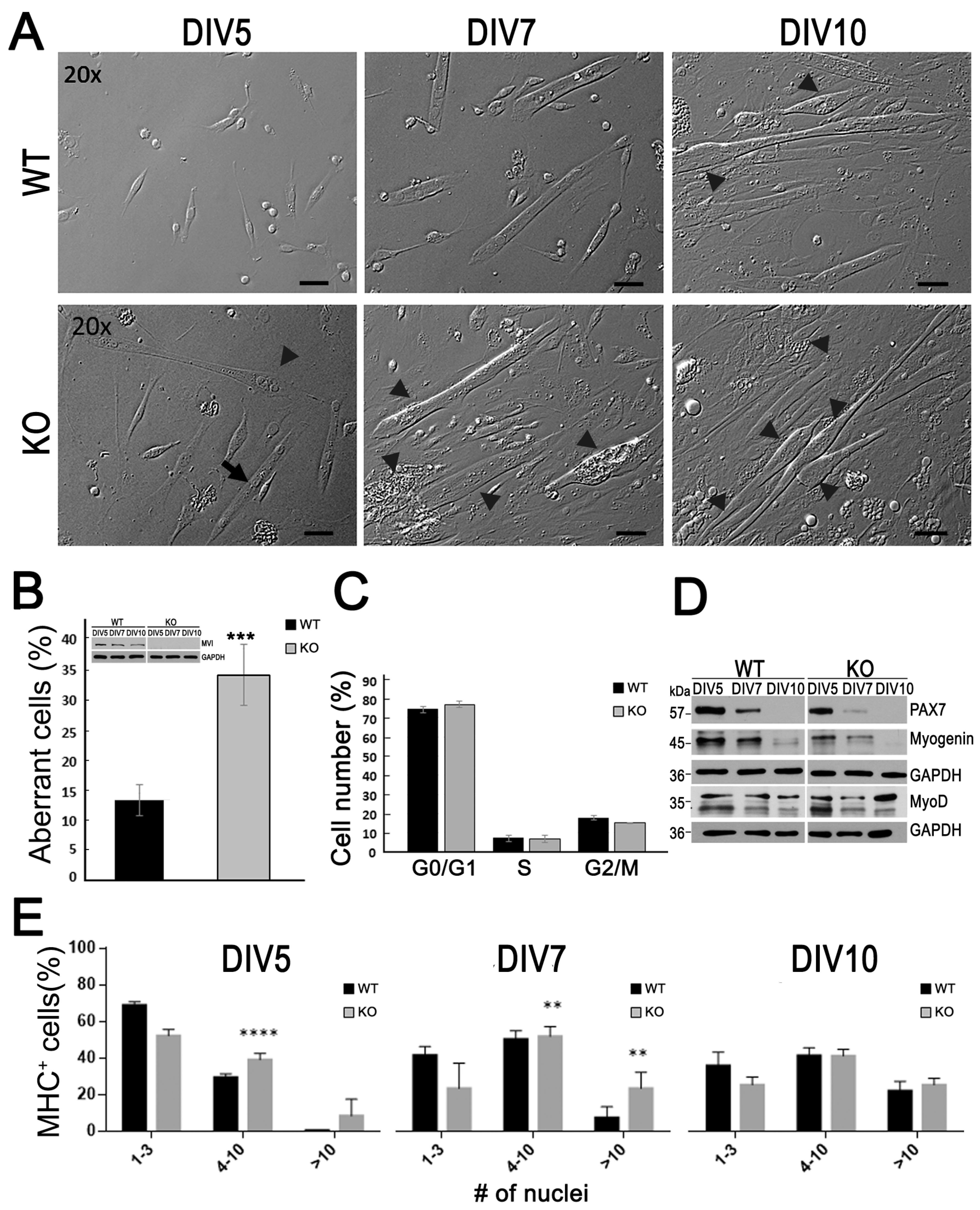{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Myoblast Isolation
2.3. Microscopy and Imaging
2.4. Immunoblotting
2.5. Antibodies and Fluorescent Markers
2.6. Cell Cycle Analysis
2.7. Statistical Analysis
3. Results
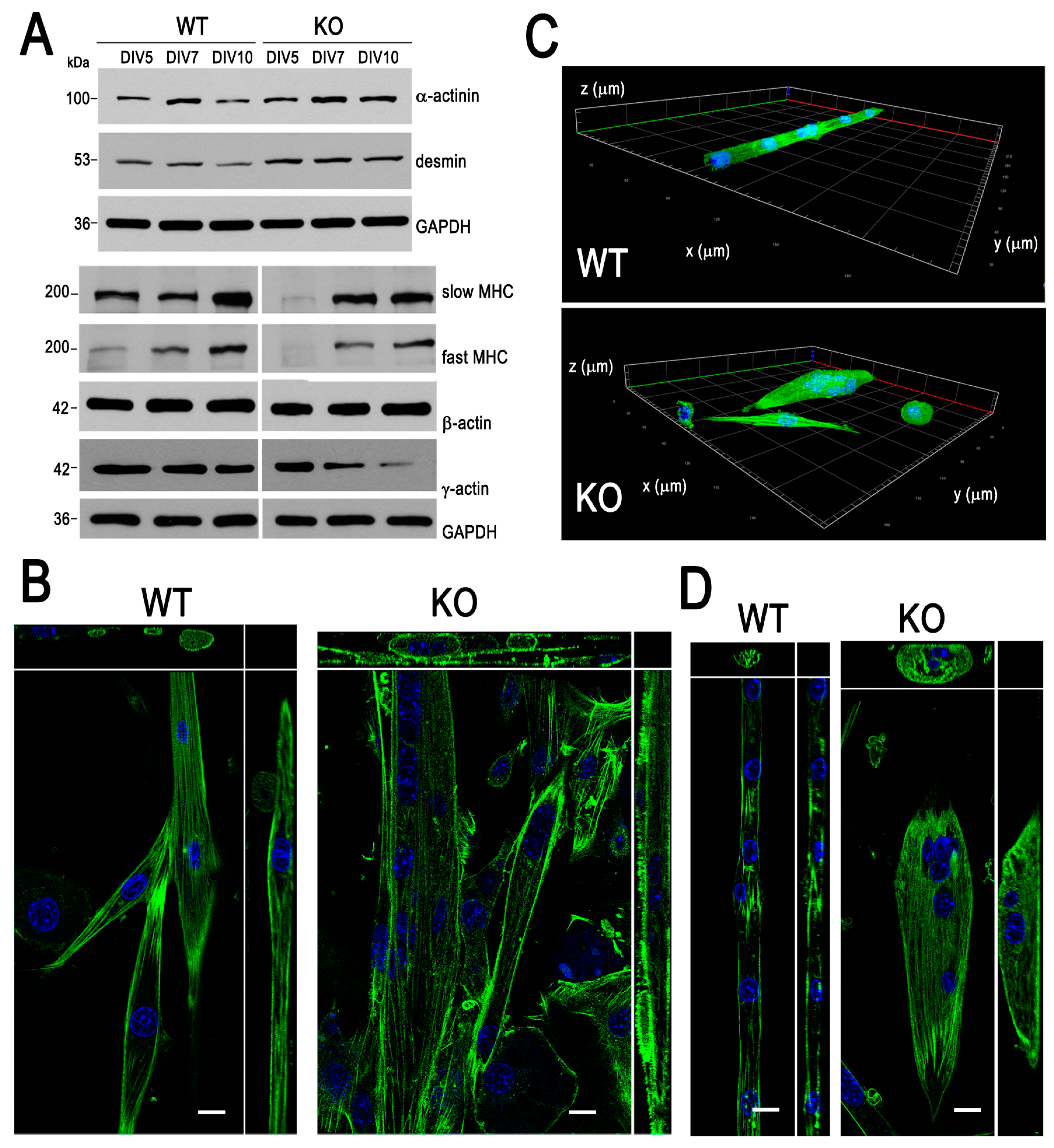
3.1. Aberrant Myotube Formation by Myoblasts Lacking MVI
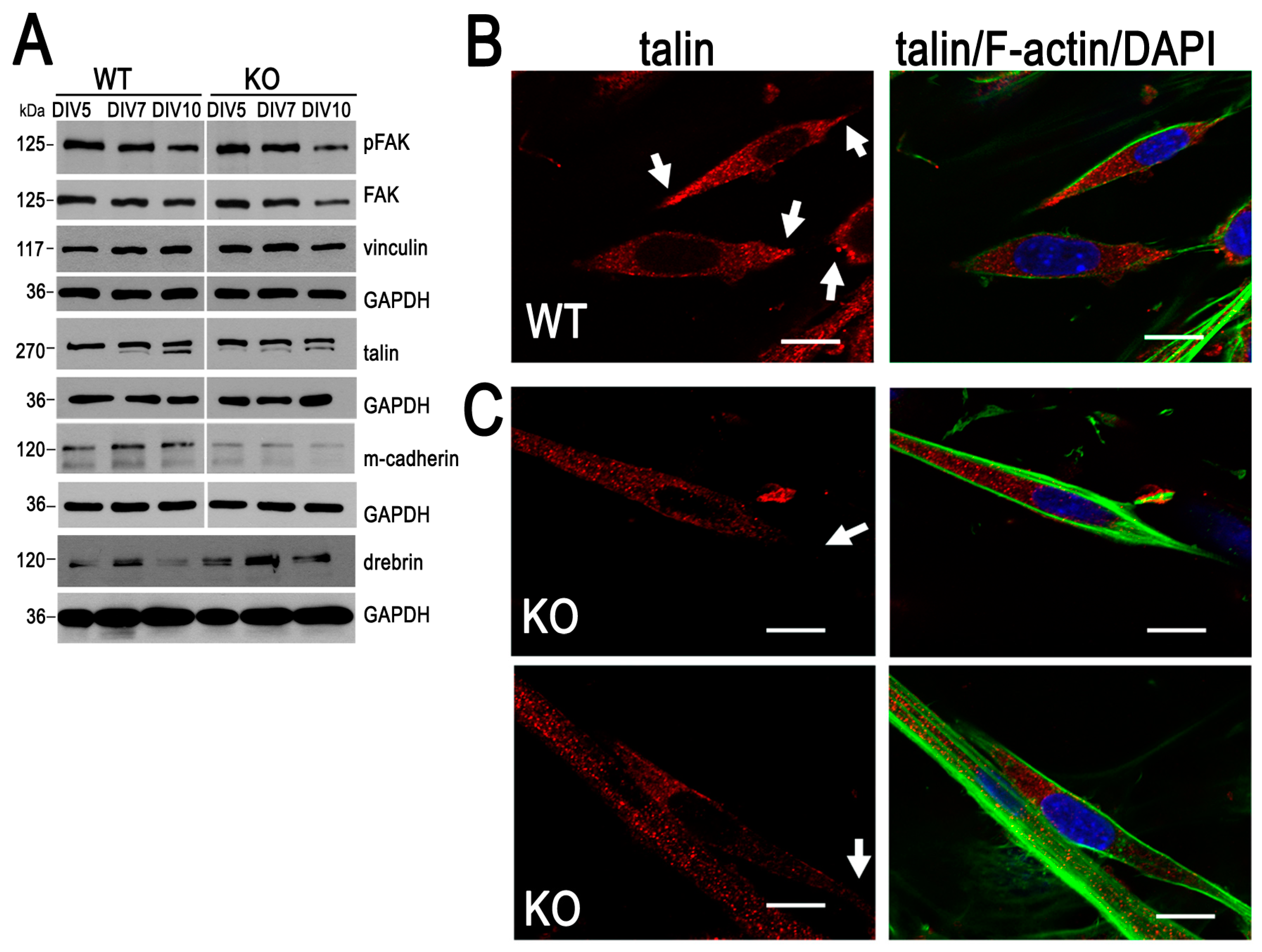
3.2. Cell Adhesion Is Impaired in Myoblasts Lacking MVI
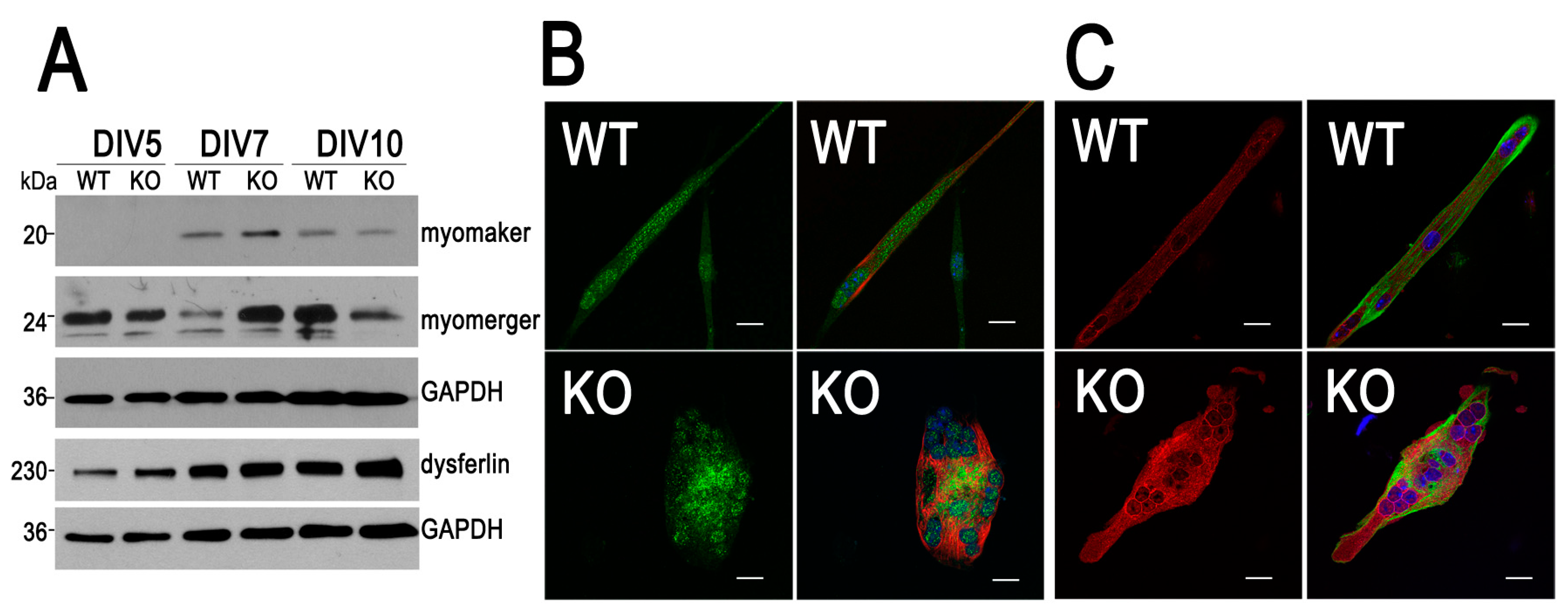
3.3. Expression Pattern of Myomaker and Myomerger Is Altered During Differentiation of Myoblasts Lacking MVI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Odronitz, F.; Kollmar, M. Drawing the tree of eukaryotic life based on the analysis of 2,269 manually annotated myosins from 328 species. Genome Boil. 2007, 8, R196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, C.; Coles, D.; Entwistle, A.; Peckham, M. Myogenic cells express multiple myosin isoforms. J. Muscle Res. Cell Motil. 1997, 18, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Salamon, M.; Millino, C.; Raffaello, A.; Mongillo, M.; Sandri, C.; Bean, C.; Negrisolo, E.; Pallavicini, A.; Valle, G.; Zaccolo, M.; et al. Human MYO18B, a novel unconventional myosin heavy chain expressed in striated muscles moves into the myonuclei upon differentiation. J. Mol. Boil. 2003, 326, 137–149. [Google Scholar] [CrossRef]
- Redowicz, M.J. Unconventional myosins in muscle. Eur. J. Cell Boil. 2007, 86, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Röder, I.V.; Strack, S.; Reischl, M.; Dahley, O.; Khan, M.M.; Kassel, O.; Zaccolo, M.; Rudolf, R. Participation of Myosin Va and Pka Type I in the Regeneration of Neuromuscular Junctions. PLoS ONE 2012, 7, e40860. [Google Scholar] [CrossRef]
- Sun, Y.; Chiu, T.T.; Foley, K.P.; Bilan, P.J.; Klip, A. Myosin Va mediates Rab8A-regulated GLUT4 vesicle exocytosis in insulin-stimulated muscle cells. Mol. Boil. Cell 2014, 25, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Luna, E.J. Unconventional myosins muscle into myofibrils. J. Boil. Chem. 2019, 294, 7219–7220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karolczak, J.; Sobczak, M.; Łukasz, M.; Yeghiazaryan, M.; Jakubiec-Puka, A.; Ehler, E.; Sławińska, U.; Wilczyński, G.M.; Rędowicz, M.J. Myosin VI in skeletal muscle: Its localization in the sarcoplasmic reticulum, neuromuscular junction and muscle nuclei. Histochem. Cell Boil. 2012, 139, 873–885. [Google Scholar] [CrossRef] [Green Version]
- Karolczak, J.; Weis, S.; Ehler, E.; Kierdaszuk, B.; Berdyński, M.; Zekanowski, C.; Kamińska, A.M.; Redowicz, M.J. Myosin VI Localization and Expression in Striated Muscle Pathology. Anat. Rec. Adv. Integr. Anat. Evol. Boil. 2014, 297, 1706–1713. [Google Scholar] [CrossRef]
- A Mohiddin, S.; Ahmed, Z.M.; Griffith, A.J.; Tripodi, D.; Friedman, T.B.; Fananapazir, L.; Morell, R. Novel association of hypertrophic cardiomyopathy, sensorineural deafness, and a mutation in unconventional myosin VI (MYO6). J. Med. Genet. 2004, 41, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Hegan, P.S.; Lanahan, A.A.; Simons, M.; Mooseker, M.S. Myosin VI and cardiomyopathy: Left ventricular hypertrophy, fibrosis, and both cardiac and pulmonary vascular endothelial cell defects in the Snell’s waltzer mouse. Cytoskelet 2015, 72, 373–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waxse, B.J.; Sengupta, P.; Hesketh, G.G.; Lippincott-Schwartz, J.; Buss, F. Myosin VI facilitates connexin 43 gap junction accretion. J. Cell Sci. 2017, 130, 827–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karolczak, J.; Pavlyk, I.; Łukasz, M.; Sobczak, M.; Niewiadomski, P.; Rzhepetskyy, Y.; Sikorska, A.; Nowak, N.; Pomorski, P.; Proszynski, T.; et al. Involvement of unconventional myosin VI in myoblast function and myotube formation. Histochem. Cell Boil. 2015, 144, 21–38. [Google Scholar] [CrossRef] [Green Version]
- Avraham, K.B.; Hasson, T.; Steel, K.P.; Kingsley, D.M.; Russell, L.B.; Mooseker, M.S.; Copeland, N.G.; Jenkins, N.A. The mouse Snell’s waltzer deafness gene encodes an unconventional myosin required for structural integrity of inner ear hair cells. Nat. Genet. 1995, 11, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Li, A.; Chen, L.-Q.; Houdusse, A.; Selvin, P.R.; Sweeney, H.L. The unique insert at the end of the myosin VI motor is the sole determinant of directionality. Proc. Natl. Acad. Sci. USA 2007, 104, 778–783. [Google Scholar] [CrossRef] [Green Version]
- Aschenbrenner, L.; Lee, T.; Hasson, T. Myo6 Facilitates the Translocation of Endocytic Vesicles from Cell Peripheries. Mol. Boil. Cell 2003, 14, 2728–2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Au, J.S.-Y.; Puri, C.; Ihrke, G.; Kendrick-Jones, J.; Buss, F. Myosin VI is required for sorting of AP-1B–dependent cargo to the basolateral domain in polarized MDCK cells. J. Cell Boil. 2007, 177, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Redowicz, M.J. Myosins and pathology: Genetics and biology. Acta Biochim. Pol. 2002, 49, 789–804. [Google Scholar] [CrossRef]
- Gotoh, N.; Yan, Q.; Du, Z.; Biemesderfer, D.; Kashgarian, M.; Mooseker, M.S.; Wang, T. Altered renal proximal tubular endocytosis and histology in mice lacking myosin-VI. Cytoskelet. 2010, 67, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Yano, H.; Ninan, I.; Zhang, H.; A Milner, T.; Arancio, O.; Chao, M.V. BDNF-mediated neurotransmission relies upon a myosin VI motor complex. Nat. Neurosci. 2006, 9, 1009–1018. [Google Scholar] [CrossRef]
- Zakrzewski, P.; Suwińska, A.; Lenartowski, R.; Rędowicz, M.J.; Buss, F.; Lenartowska, M. Myosin VI maintains the actin-dependent organization of the tubulobulbar complexes required for endocytosis during mouse spermiogenesis. Boil. Reprod. 2020, 102, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, P.; Rędowicz, M.J.; Buss, F.; Lenartowska, M. Loss of myosin VI expression affects acrosome/acroplaxome complex morphology during mouse spermiogenesis. Boil. Reprod. 2020. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.J.; Liu, G.; Zhou, W.; Chen, X. Myosin VI Is a Mediator of the p53-Dependent Cell Survival Pathway. Mol. Cell. Boil. 2006, 26, 2175–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vreugde, S.; Ferrai, C.; Miluzio, A.; Hauben, E.; Marchisio, P.C.; Crippa, M.; Bussi, M.; Biffo, S. Nuclear Myosin VI Enhances RNA Polymerase II-Dependent Transcription. Mol. Cell 2006, 23, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, H.L.; Houdusse, A. Myosin VI Rewrites the Rules for Myosin Motors. Cell 2010, 141, 573–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, H.L.; Houdusse, A. What can myosin VI do in cells? Curr. Opin. Cell Boil. 2007, 19, 57–66. [Google Scholar] [CrossRef]
- Chibalina, M.V.; Puri, C.; Kendrick-Jones, J.; Buss, F. Potential roles of myosin VI in cell motility. Biochem. Soc. Trans. 2009, 37, 966–970. [Google Scholar] [CrossRef] [Green Version]
- Majewski, L.; Nowak, J.; Sobczak, M.; Karatsai, O.; Havrylov, S.; Lenartowski, R.; Suszek, M.; Lenartowska, M.; Rędowicz, M.J. Myosin VI in the nucleus of neurosecretory PC12 cells: Stimulation-dependent nuclear translocation and interaction with nuclear proteins. Nuclear 2018, 9, 125–141. [Google Scholar] [CrossRef] [Green Version]
- A Tumbarello, D.; Kendrick-Jones, J.; Buss, F. Myosin VI and its cargo adaptors – linking endocytosis and autophagy. J. Cell Sci. 2013, 126, 2561–2570. [Google Scholar] [CrossRef] [Green Version]
- Spudich, G.; Chibalina, M.V.; Au, J.S.-Y.; Arden, S.D.; Buss, F.; Kendrick-Jones, J. Myosin VI targeting to clathrin-coated structures and dimerization is mediated by binding to Disabled-2 and PtdIns(4,5)P2. Nature 2006, 9, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Mukherjea, M.; Ali, M.Y.; Kikuti, C.; Safer, D.; Yang, Z.; Sirkia, H.; Ropars, V.; Houdusse, A.; Warshaw, D.M.; Sweeney, H.L. Myosin VI must dimerize and deploy its unusual lever arm in order to perform its cellular roles. Cell Rep. 2014, 8, 1522–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łukasz, M.; Sobczak, M.; Havrylov, S.; Jóźwiak, J.; Redowicz, M.J. Dock7: A GEF for Rho-family GTPases and a novel myosin VI-binding partner in neuronal PC12 cells. Biochem. Cell Boil. 2012, 90, 565–574. [Google Scholar] [CrossRef]
- Karolczak, J.; Sobczak, M.; Skowronek, K.; Redowicz, M.J. A Kinase Anchoring Protein 9 Is a Novel Myosin VI Binding Partner That Links Myosin VI with the PKA Pathway in Myogenic Cells. BioMed Res. Int. 2015, 2015, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehka, L.; Redowicz, M.J. Mechanisms regulating myoblast fusion: A multilevel interplay. Semin. Cell Dev. Biol. 2020, in press. [Google Scholar] [CrossRef]
- Krauss, R.S.; Joseph, G.A.; Goel, A.J. Keep Your Friends Close: Cell–Cell Contact and Skeletal Myogenesis. Cold Spring Harb. Perspect. Boil. 2017, 9, a029298. [Google Scholar] [CrossRef] [Green Version]
- Ludolph, D.C.; Konieczny, S.F. Transcription factor families: Muscling in on the myogenic program. FASEB J. 1995, 9, 1595–1604. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; Dezonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Doetschman, T.C.; Eppenberger, H.M. Comparison of M-line and other myofibril components dur-ing reversible phorbol ester treatment. Eur. J. Cell Biol. 1984, 33, 265–274. [Google Scholar]
- Lin, J.J. Assembly of different isoforms of actin and tropomyosin into the skeletal tropomyosin-enriched microfilaments during differentiation of muscle cells in vitro. J. Cell Boil. 1986, 103, 2173–2183. [Google Scholar] [CrossRef] [Green Version]
- Maddugoda, M.P.; Crampton, M.S.; Shewan, A.; Yap, A.S. Myosin VI and vinculin cooperate during the morphogenesis of cadherin cell–cell contacts in mammalian epithelial cells. J. Cell Boil. 2007, 178, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Geisbrecht, E.R.; Montell, D.J. Myosin VI is required for E-cadherin-mediated border cell migration. Nature 2002, 4, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Krauss, R.S. Regulation of Skeletal Myoblast Differentiation by Drebrin; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2017; Volume 1006, pp. 361–373. [Google Scholar]
- Rehm, K.; Panzer, L.; Van Vliet, V.; Genot, E.; Linder, S. Drebrin preserves endothelial integrity by stabilizing nectin at adherens junctions. J. Cell Sci. 2013, 126, 3756–3769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malla, R.R.; Vempati, R.K. Talin: A Potential Drug Target for Cancer Therapy. Curr. Drug Metab. 2020, 21, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; You, W.; Wang, Y.; Shan, T. The regulatory role of Myomaker and Myomixer–Myomerger–Minion in muscle development and regeneration. Cell. Mol. Life Sci. 2019, 77, 1551–1569. [Google Scholar] [CrossRef] [PubMed]
- Millay, U.P.; O’Rourke, J.R.; Sutherland, L.B.; Bezprozvannaya, S.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Myomaker is a membrane activator of myoblast fusion and muscle formation. Nature 2013, 499, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Vashisht, A.A.; O’Rourke, J.; Corbel, S.Y.; Moran, R.; Romero, A.; Miraglia, L.; Zhang, J.; Durrant, E.; Schmedt, C.; et al. The microprotein Minion controls cell fusion and muscle formation. Nat. Commun. 2017, 8, 15664. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas, A.M.; González-Jamett, A.M.; Cea, L.A.; Bevilacqua, J.A.; Caviedes, P. Dysferlin function in skeletal muscle: Possible pathological mechanisms and therapeutical targets in dysferlinopathies. Exp. Neurol. 2016, 283, 246–254. [Google Scholar] [CrossRef]
- McGrath, M.J.; Cottle, D.; Nguyen, M.-A.; Dyson, J.M.; Coghill, I.D.; Robinson, P.A.; Holdsworth, M.; Cowling, B.S.; Hardeman, E.C.; Mitchell, C.A.; et al. Four and a Half LIM Protein 1 Binds Myosin-binding Protein C and Regulates Myosin Filament Formation and Sarcomere Assembly. J. Boil. Chem. 2006, 281, 7666–7683. [Google Scholar] [CrossRef] [Green Version]
- Roman, W.; Gomes, E.R. Nuclear positioning in skeletal muscle. Semin. Cell Dev. Boil. 2018, 82, 51–56. [Google Scholar] [CrossRef]
- Masters, T.A.; A Tumbarello, D.; Chibalina, M.V.; Buss, F. MYO6 Regulates Spatial Organization of Signaling Endosomes Driving AKT Activation and Actin Dynamics. Cell Rep. 2017, 19, 2088–2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobczak, M.; Chumak, V.; Pomorski, P.; Wojtera, E.; Łukasz, M.; Nowak, J.; Yamauchi, J.; Redowicz, M.J. Interaction of myosin VI and its binding partner DOCK7 plays an important role in NGF-stimulated protrusion formation in PC12 cells. Biochim. Biophys. Acta (BBA) Bioenerg. 2016, 1863, 1589–1600. [Google Scholar] [CrossRef] [PubMed]
- Lv, K.; Chen, L.; Li, Y.; Li, Z.; Zheng, P.; Liu, Y.; Chen, J.; Teng, J. Trip6 Promotes Dendritic Morphogenesis through Dephosphorylated GRIP1-Dependent Myosin VI and F-Actin Organization. J. Neurosci. 2015, 35, 2559–2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckingham, M.; Rigby, P.W. Gene Regulatory Networks and Transcriptional Mechanisms that Control Myogenesis. Dev. Cell 2014, 28, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Cheng, W.; Hung, J.; Montell, D.; Geisbrecht, E.; Rosen, D.; Liu, J.; Naora, H. Lessons from border cell migration in the Drosophila ovary: A role for myosin VI in dissemination of human ovarian cancer. Proc. Natl. Acad. Sci. USA 2004, 101, 8144–8149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, B.R.; Nestor-Bergmann, A.; Liang, X.; Gupta, S.; Duszyc, K.; Gauquelin, E.; Gomez, G.A.; Budnar, S.; Marcq, P.; Jensen, O.E.; et al. A Mechanosensitive RhoA Pathway that Protects Epithelia against Acute Tensile Stress. Dev. Cell 2018, 47, 439–452.e6. [Google Scholar] [CrossRef] [Green Version]
- Tuxworth, R.I.; Stephens, S.; Ryan, Z.C.; Titus, M.A. Identification of a Myosin VII-Talin Complex. J. Boil. Chem. 2005, 280, 26557–26564. [Google Scholar] [CrossRef] [Green Version]
- Galdeen, S.A.; Stephens, S.; Thomas, D.D.; Titus, M.A. Talin Influences the Dynamics of the Myosin VII-Membrane Interaction. Mol. Boil. Cell 2007, 18, 4074–4084. [Google Scholar] [CrossRef] [Green Version]
- Swailes, N. Non-muscle myosins 2A and 2B drive changes in cell morphology that occur as myoblasts align and fuse. J. Cell Sci. 2006, 119, 3561–3570. [Google Scholar] [CrossRef] [Green Version]
- Schaller, M.D. Cellular functions of FAK kinases: Insight into molecular mechanisms and novel functions. J. Cell Sci. 2010, 123, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Willkomm, L.; Bloch, W. State of the Art in Cell–Cell Fusion. In Methods in Molecular Biology; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2015; Volume 1313, pp. 1–19. [Google Scholar]
- Mancini, A.; Sirabella, D.; Zhang, W.; Yamazaki, H.; Shirao, T.; Krauss, R.S. Regulation of myotube formation by the actin-binding factor drebrin. Skelet. Muscle 2011, 1, 36. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehka, L.; Topolewska, M.; Wojton, D.; Karatsai, O.; Alvarez-Suarez, P.; Pomorski, P.; Rędowicz, M.J. Formation of Aberrant Myotubes by Myoblasts Lacking Myosin VI Is Associated with Alterations in the Cytoskeleton Organization, Myoblast Adhesion and Fusion. Cells 2020, 9, 1673. https://doi.org/10.3390/cells9071673
Lehka L, Topolewska M, Wojton D, Karatsai O, Alvarez-Suarez P, Pomorski P, Rędowicz MJ. Formation of Aberrant Myotubes by Myoblasts Lacking Myosin VI Is Associated with Alterations in the Cytoskeleton Organization, Myoblast Adhesion and Fusion. Cells. 2020; 9(7):1673. https://doi.org/10.3390/cells9071673
Chicago/Turabian StyleLehka, Lilya, Małgorzata Topolewska, Dominika Wojton, Olena Karatsai, Paloma Alvarez-Suarez, Paweł Pomorski, and Maria Jolanta Rędowicz. 2020. "Formation of Aberrant Myotubes by Myoblasts Lacking Myosin VI Is Associated with Alterations in the Cytoskeleton Organization, Myoblast Adhesion and Fusion" Cells 9, no. 7: 1673. https://doi.org/10.3390/cells9071673