
Potential Therapeutic Effects of New Ruthenium (III) Complex with Quercetin: Characterization, Structure, Gene Regulation, and Antitumor and Anti-Inflammatory Studies (RuIII/Q Novel Complex Is a Potent Immunoprotective Agent)

 , ,
, ,  ,
,
Abstract
:1. Intoduction
2. Materials and Methods
2.1. Chemicals and Analyses
| Type of Analysis | Models |
| Elemental analyses | Perkin Elmer CHN 2400 |
| Conductance | Jenway 4010 conductivity meter |
| FTIR spectra | Bruker FTIR Spectrophotometer |
| Electronic spectra | UV2 Unicam UV/Vis Spectrophotometer |
| Thermo gravimetric | TG/DTG–50H, Shimadzu thermo-gravimetric analyzer |
| TEM | JEOL 100 s microscopy |
In Vitro Release Profile of Q and Ru from Ruthenium(III) Complex with Q
2.2. Synthesisof New RuIII/Q III Complex
2.3. Animal Model
2.4. Experimental Design
2.5. Blood Samples
2.6. Preparation of Tissue Homogenates for Determination of the Redox State
2.7. Determination of Oxidative Stress Biomarker Activities in Either Brain and Testicular Tissues Homogenates
2.8. TNF-A and IL-6 Activities in the Testis Homogenates
2.9. Mitochondrial ROS Assay
2.10. Mitochondria Membrane Potential (MMP, Δψm) Assay
2.11. Fluorescent Detection of Mitochondrial Membrane Potential
2.12. Cytochrome C Oxidase Release Assay
2.13. Determination Of Plasma Homocysteine (Hcy) in Brain Homogenates
2.14. Determination Of Amyloid Beta (Aβ1-42) in Brain Homogenates
2.15. Determination of Gamma-Aminobutyric Acid (GABA) in Serum
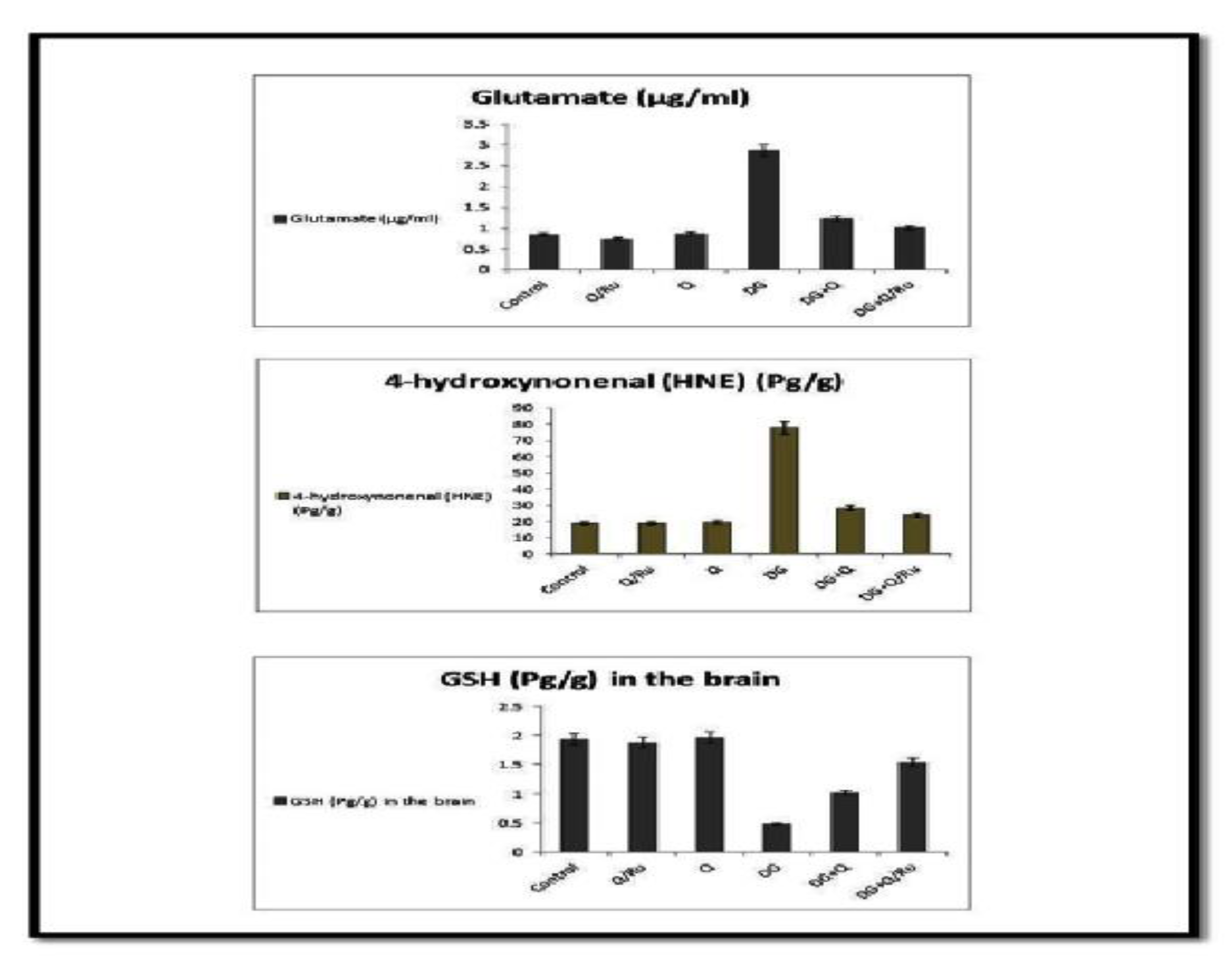
2.16. Determination of Glutamate in the Brain (GSH)
2.17. Determination of 4-Hydroxynonenal (HNE) in the Brain
2.18. Histological Changes and Transmission Electron Microscopy (TEM) Estimation
2.19. Single Cell Gel Electrophoresis (SCGE) (The Comet Assay)
2.20. Detection of Apoptosis Using Annexin V/PI Staining
2.21. Determination of Cytochrome-c
2.22. Determination of P53
2.23. Determination of Bax and BCl2
2.24. Determination of Caspase 3 and 9
2.25. RNA Isolation and Quantitative Reverse Transcription Polymerase Chain Reaction
2.26. Real Time-PCR
2.27. Cells Line
2.28. Cell Culture
2.29. Cell Culture Medium
2.30. Treatment of Cancer Cell Lines
2.31. Cytotoxic Activity (MTT Assay and IC50 Determination)
2.32. Counting of Cells
2.33. Flow Cytometric Analysis
2.34. Staining Procedure (Direct Staining Method)
2.35. Annexin V-FITC/PI Staining Assay
2.36. Analysis of Flow Cytometry of p53 and Bax Expression
2.37. Statistical Analysis
2.38. Sample Size Calculation:
3. Results
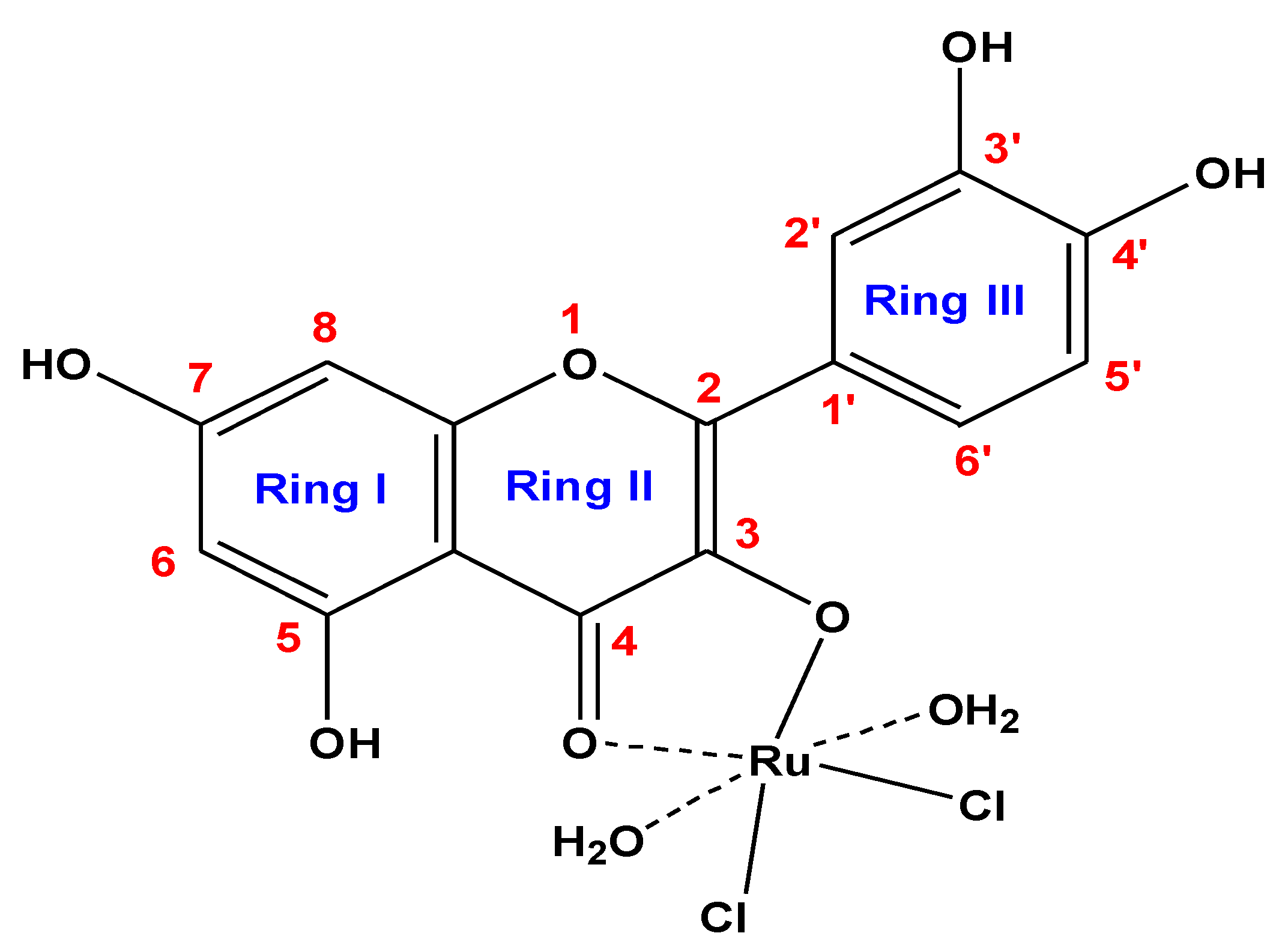
3.1. Chemical Composition Interpretations
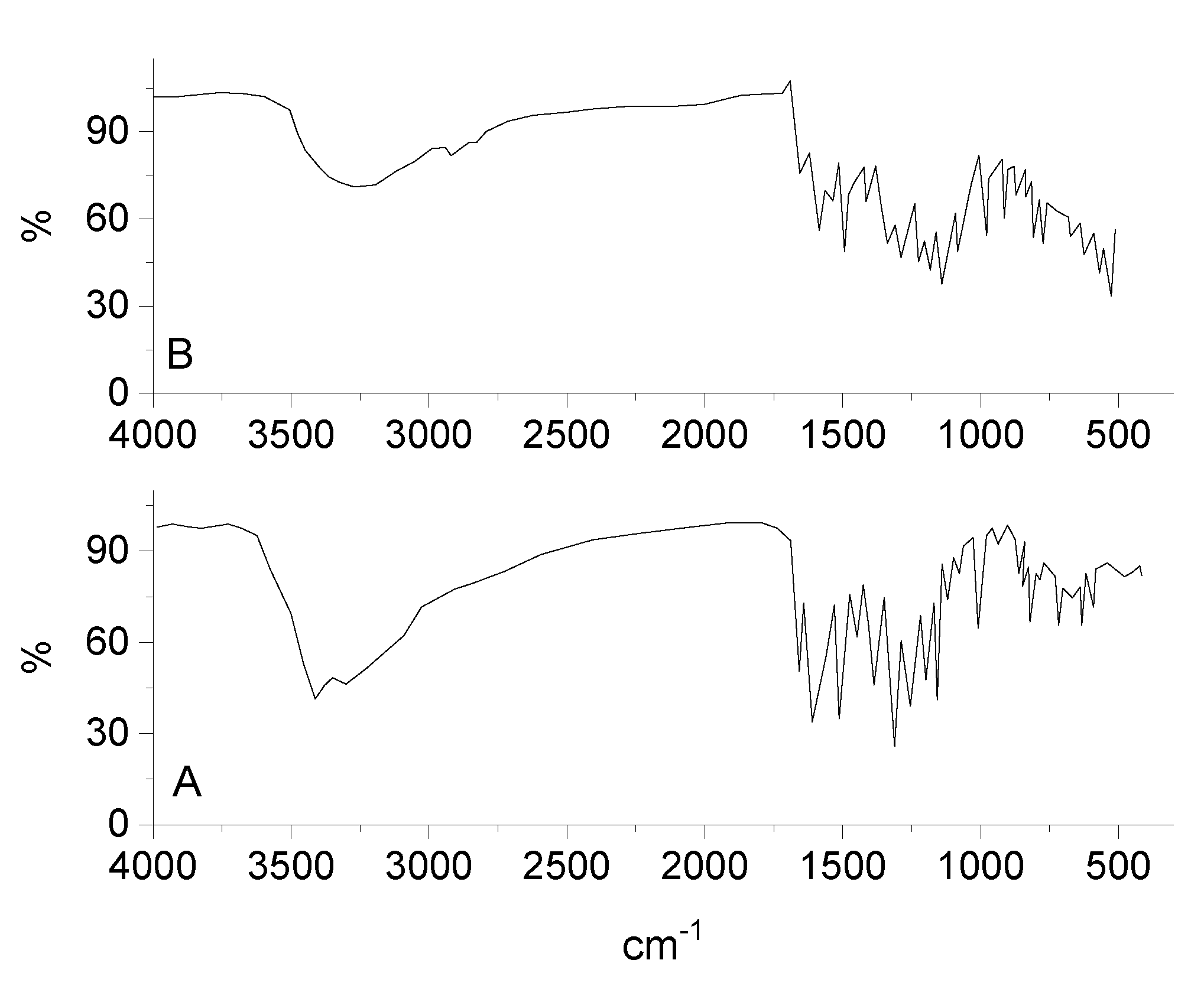
3.2. Infrared Spectra
3.3. Electronic Spectra
3.4. Thermal Analysis
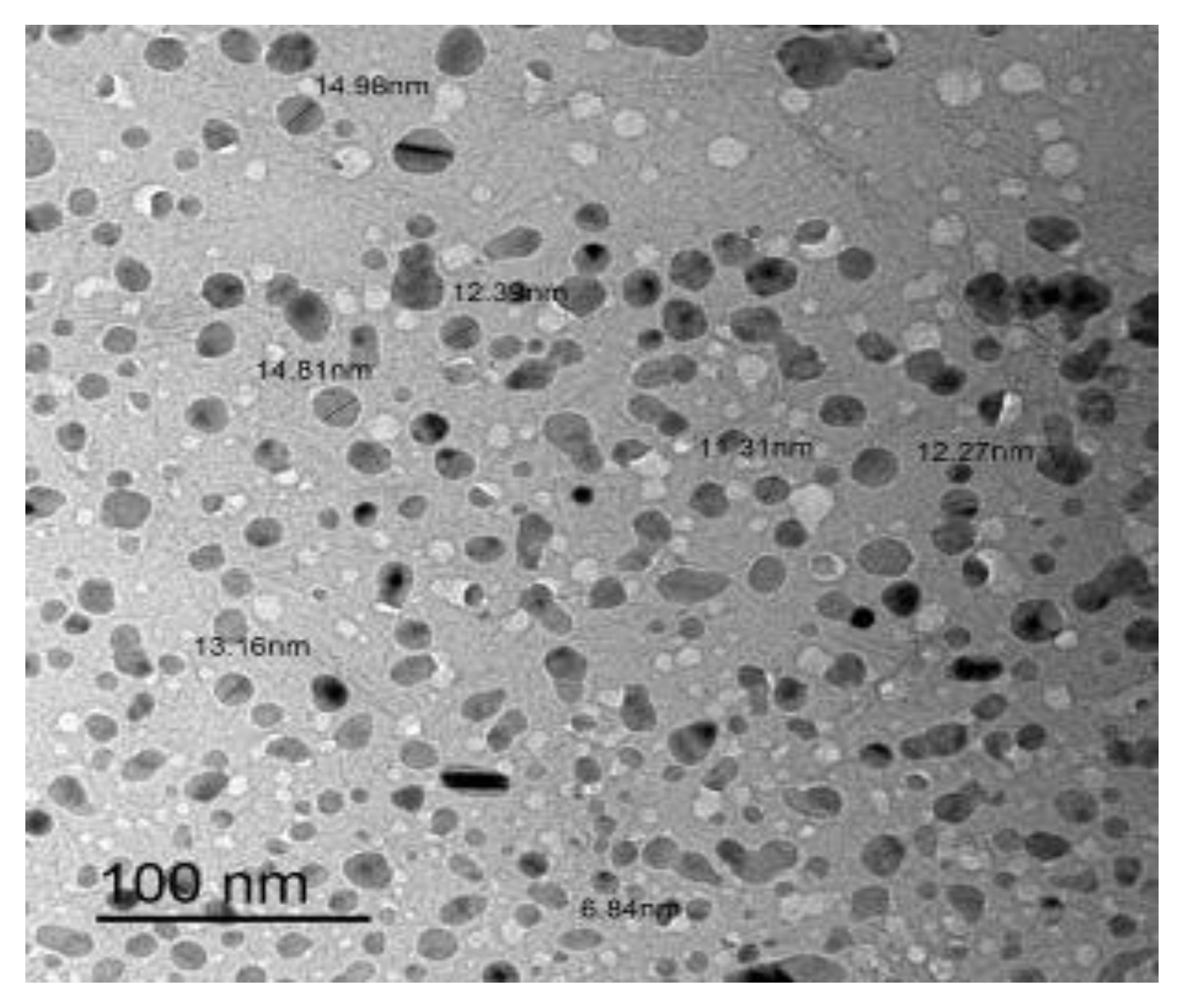
3.5. Transmission Electron Microscopy (TEM) Investigation
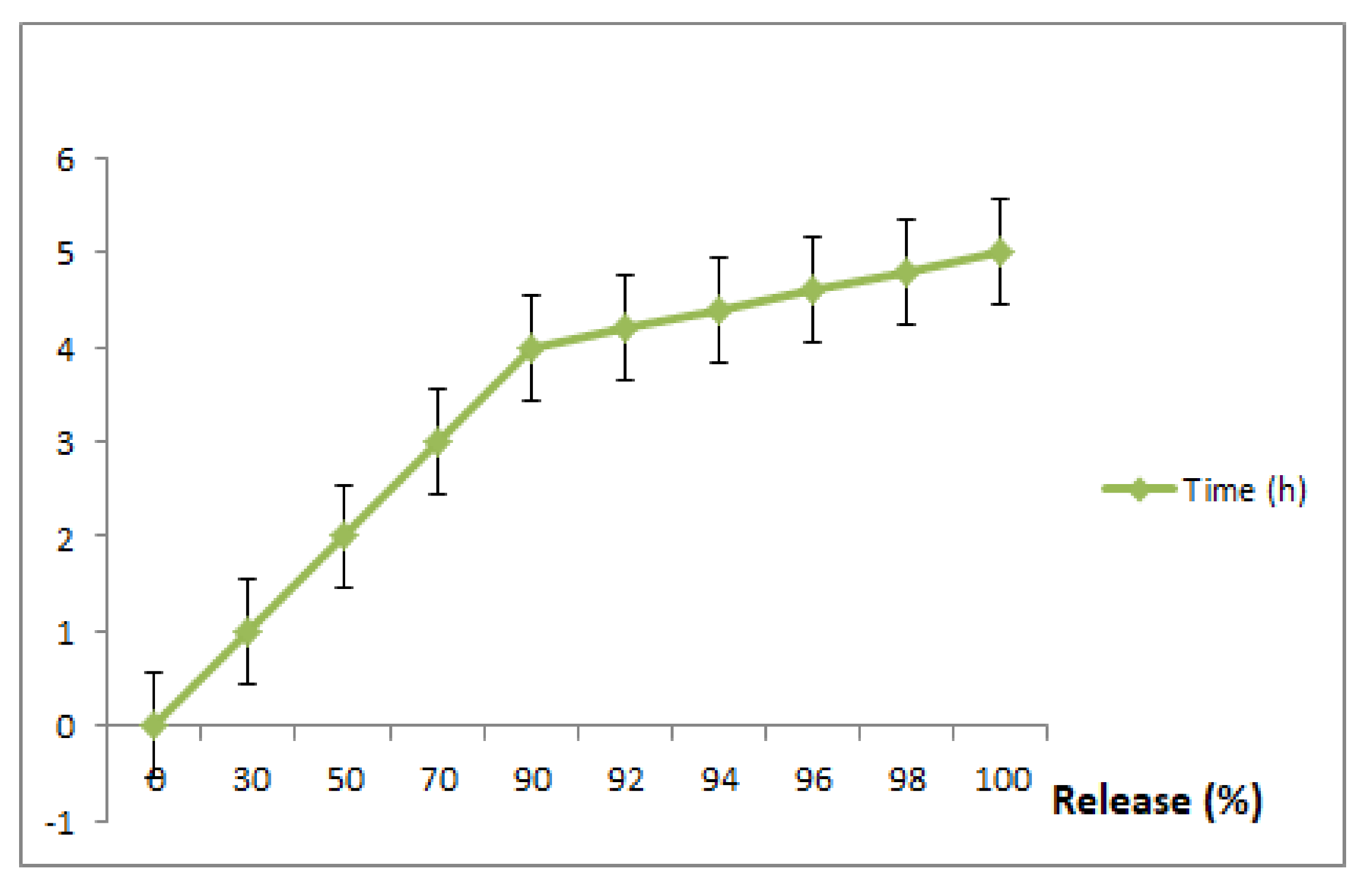
3.6. In Vitro Drug Release
3.7. Biological Interpretations
3.7.1. Determination of Cell Viability Percentage (%) (MTT Assay)
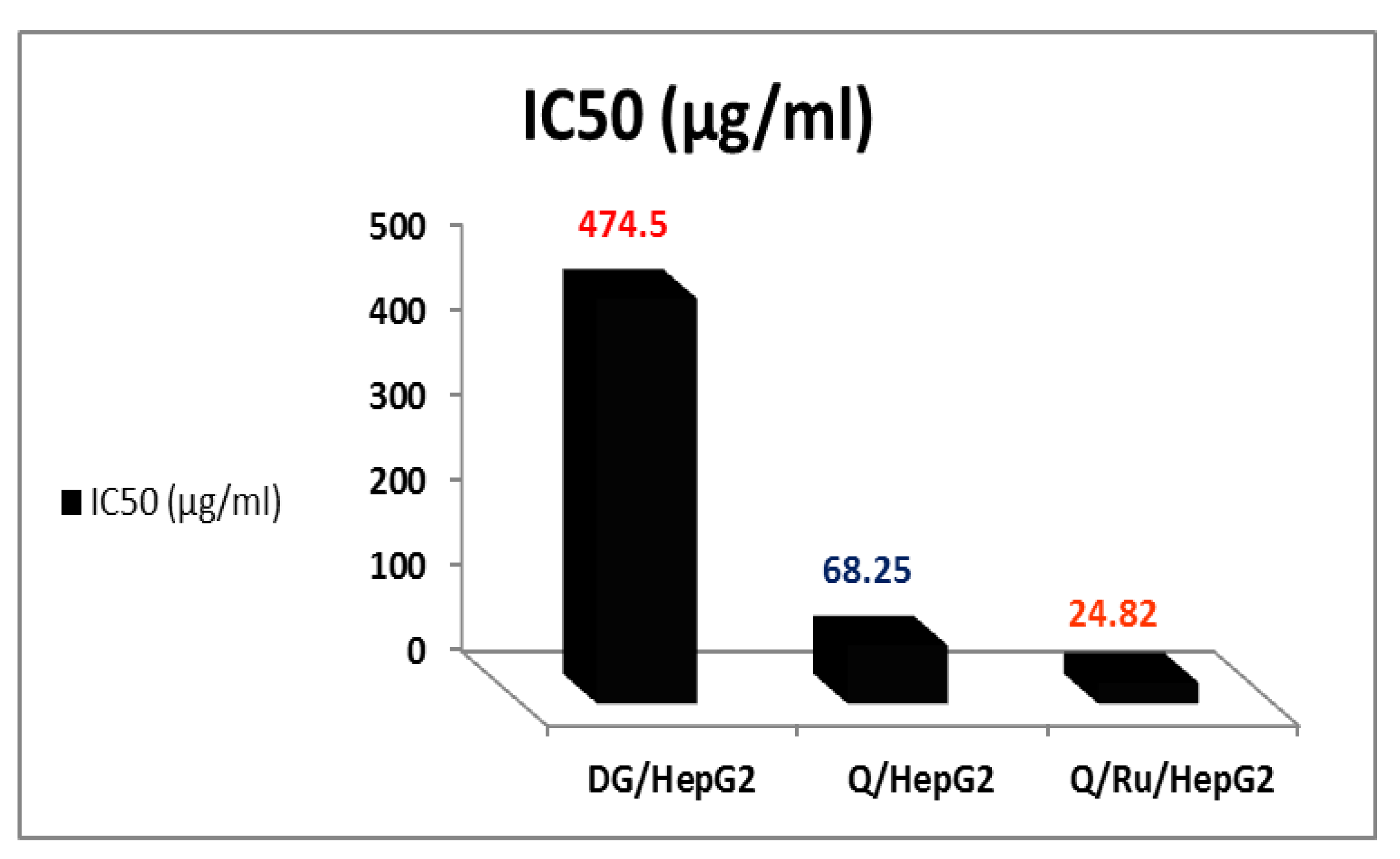
3.7.2. Determination of IC50 Doses
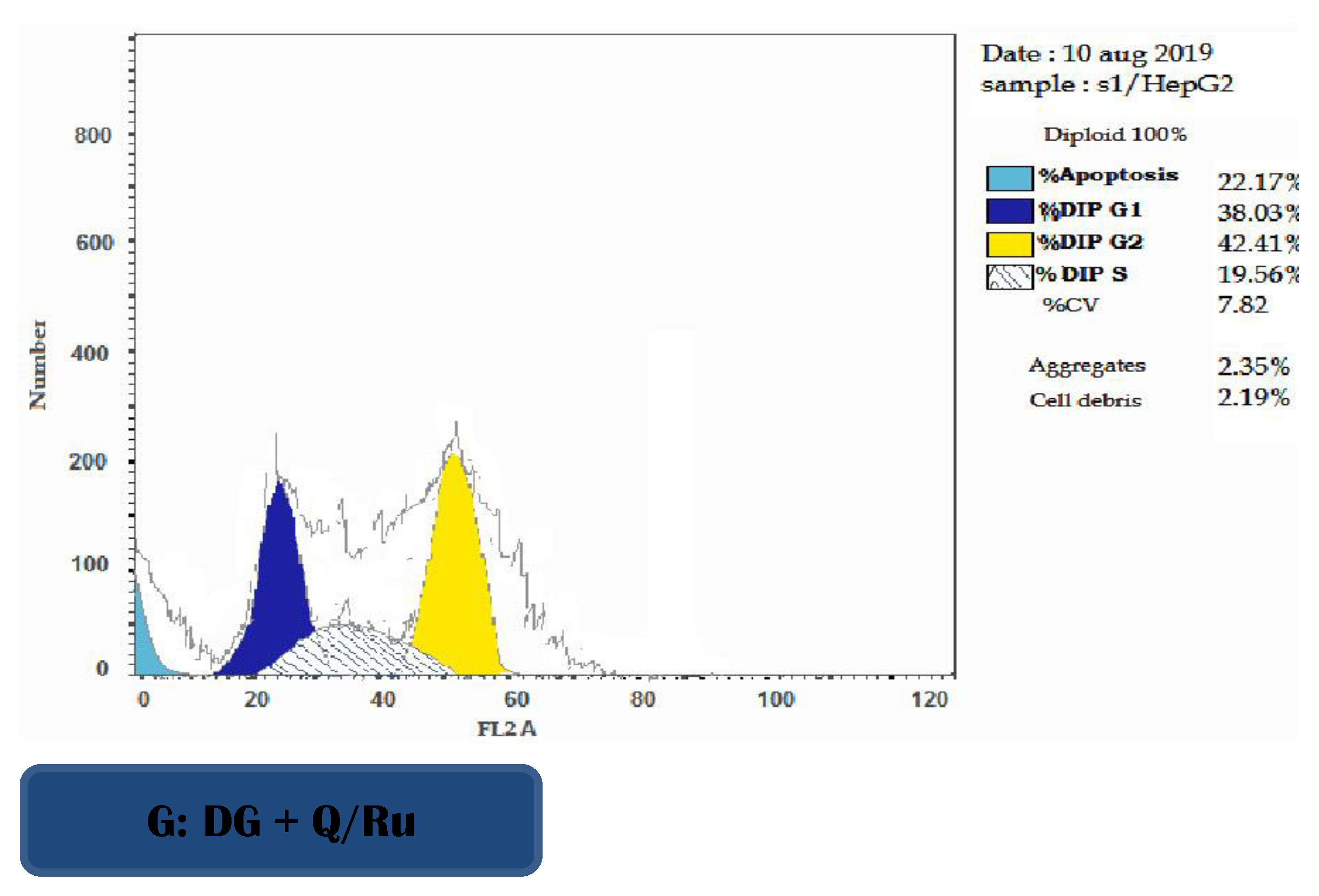
3.7.3. Determination of Apoptosis Using Annexin V/PI in HepG2 Cells
3.7.4. Cell Cycle Analysis
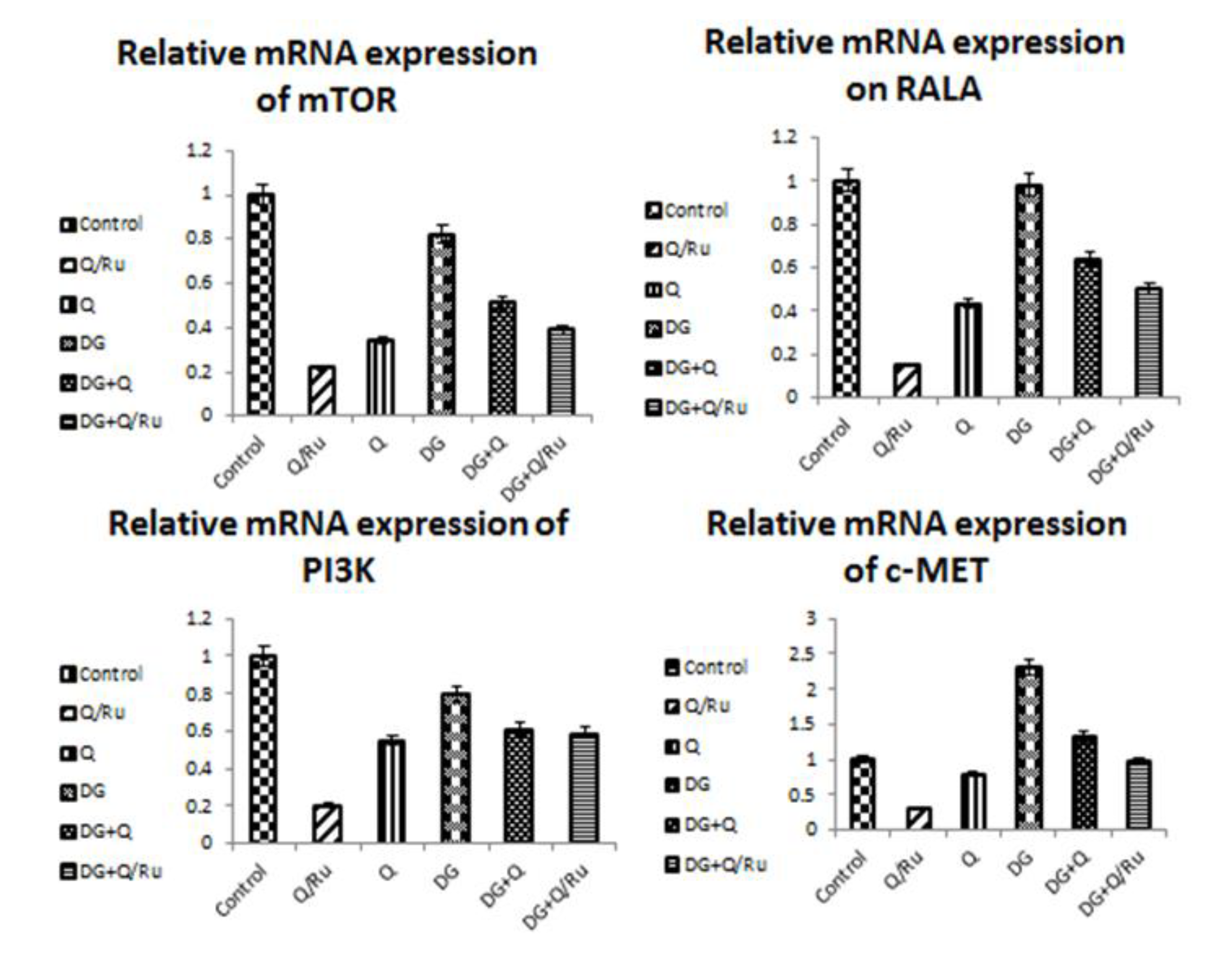
3.7.5. Gene Expression for mTOR, RALA, PI3K and c-Met Expression in HepG2 Cells
3.7.6. Gene Expression for Apoptotic GenesP53 Expression in HepG2 Cells
3.7.7. Bax Expression in HepG2 Cells
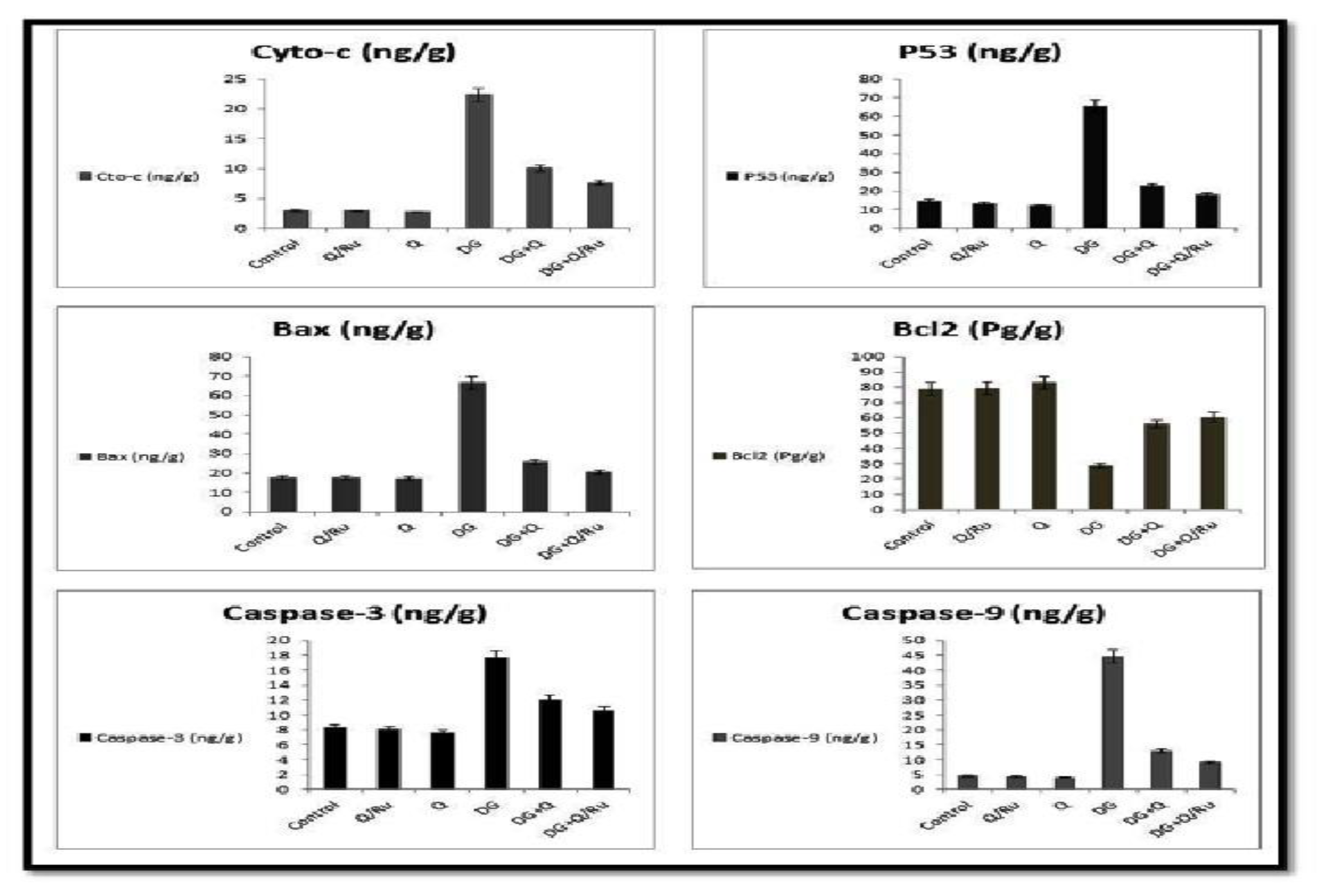
3.7.8. Gene Expression for cyto-c, Caspase-3, P53, Bax, BCl2, and Caspase-9 Expression in Testicular Tissues
3.7.9. Effect on Glutamate, 4-hydroxynonenal (HNE), GSH, HCY, Amyloid Beta and GABA Levels
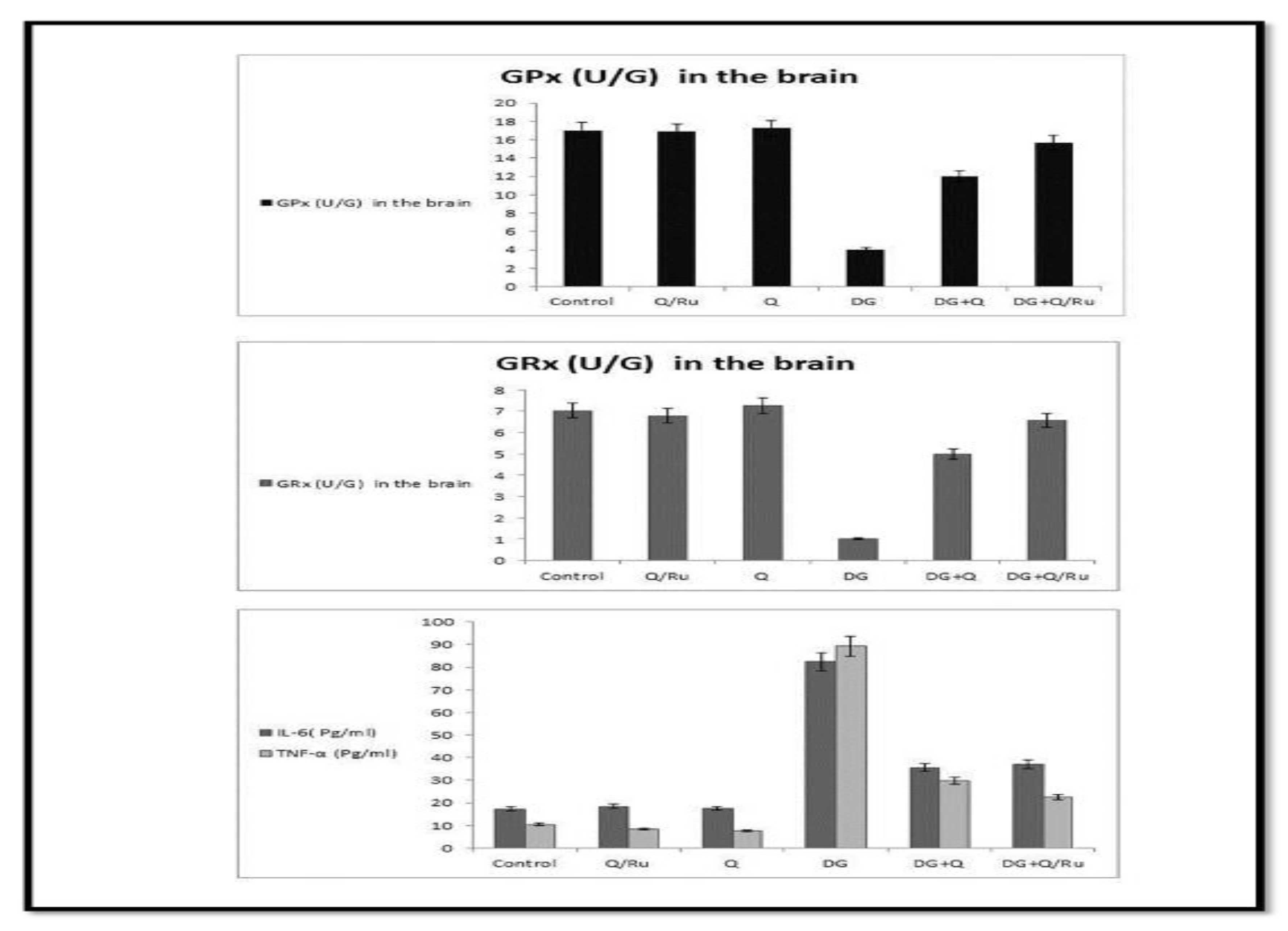
3.7.10. Effect on IL-6 and TNF-α
3.7.11. Oxidative Stress Enzyme Markers
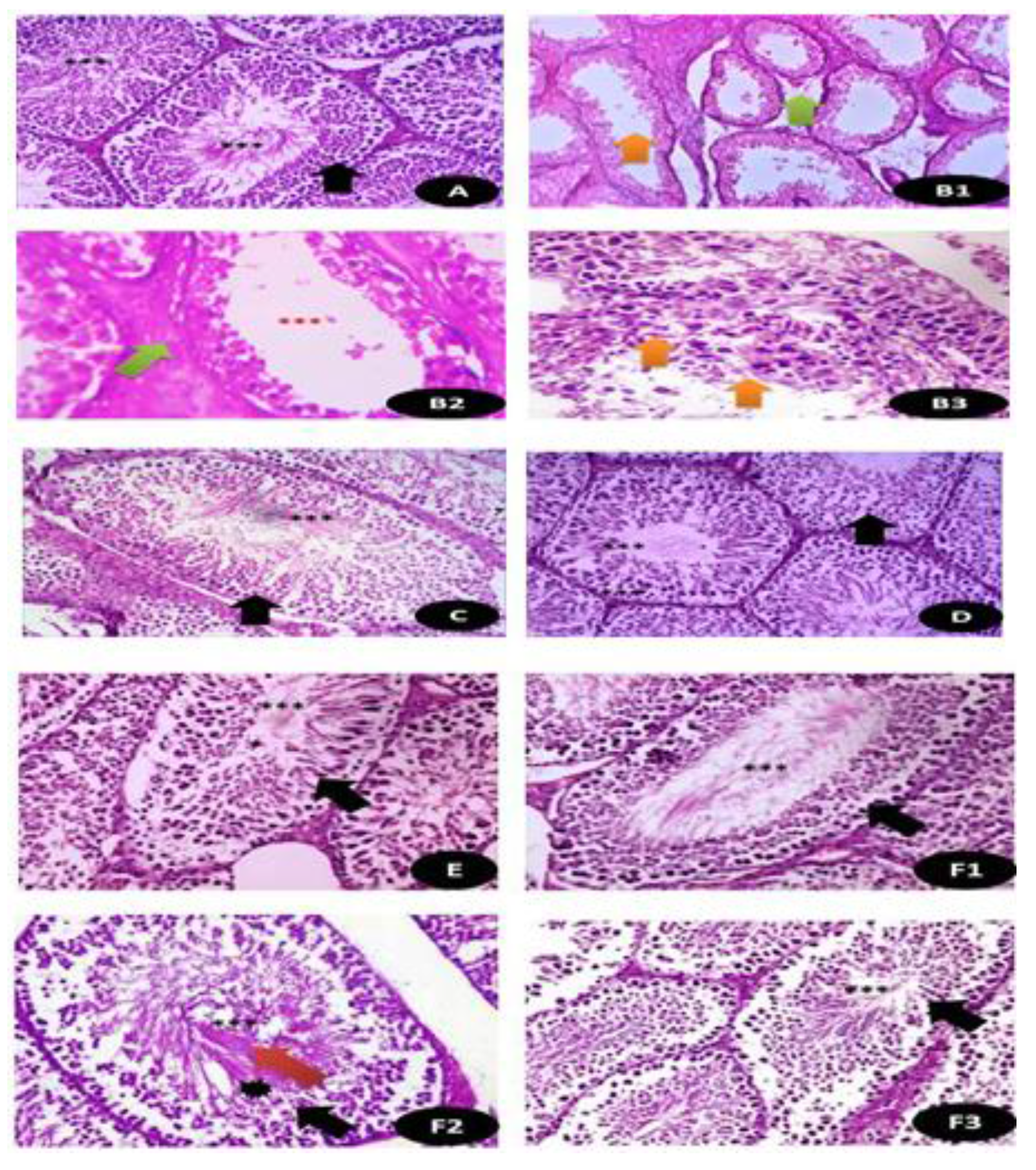
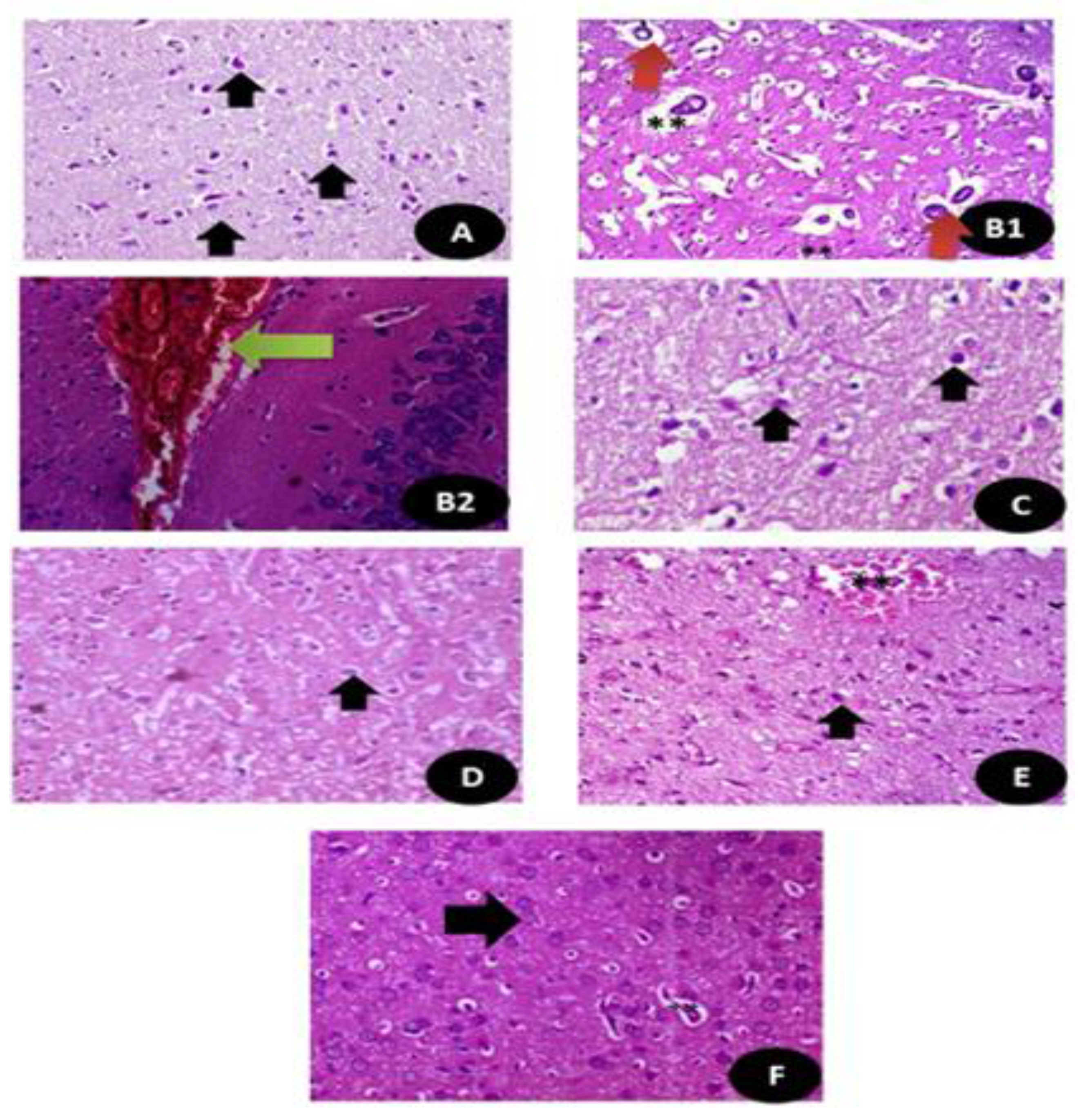
3.7.12. Histological Examination Testicular and Brain Tissues (Cerebral Cortex)
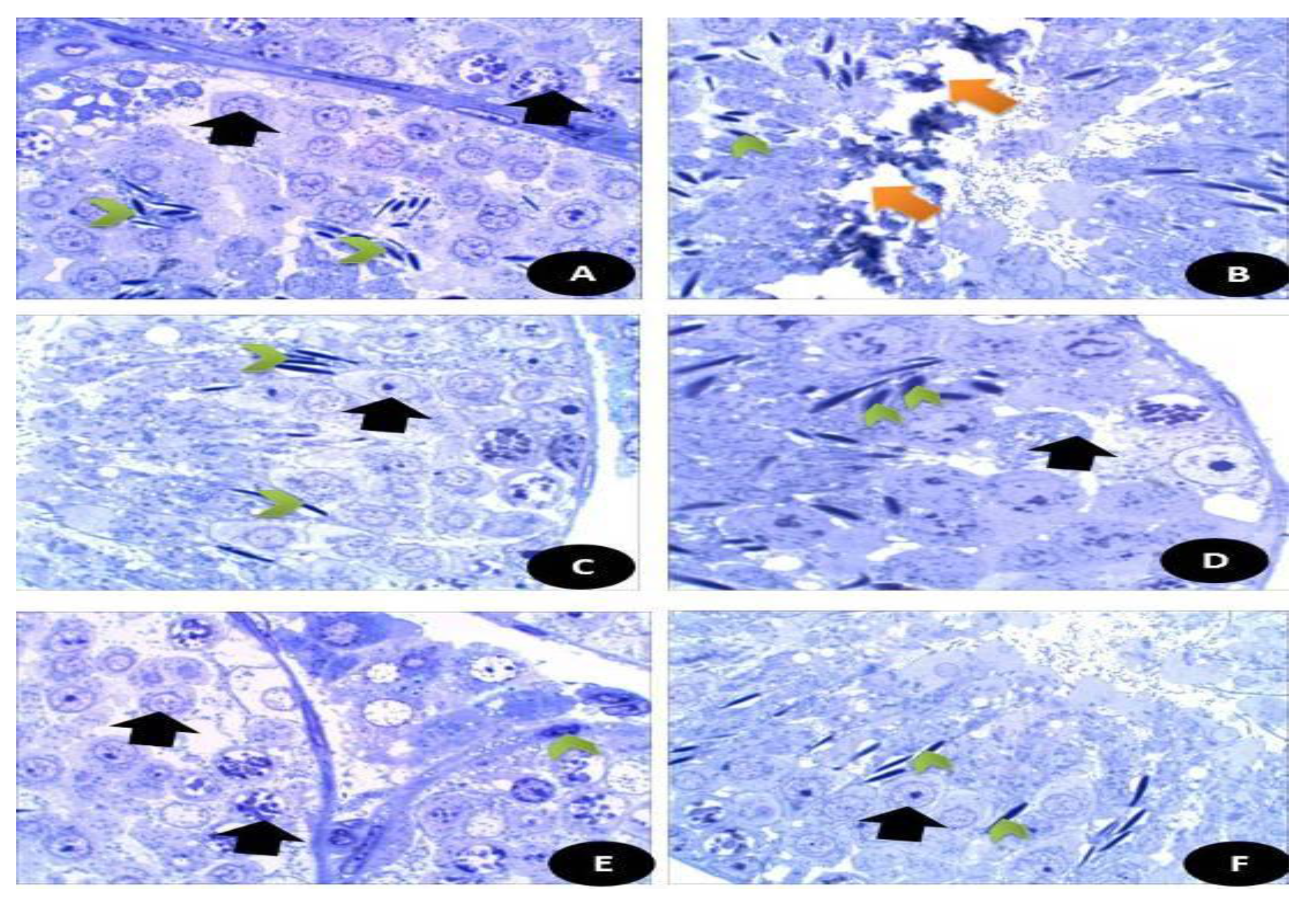
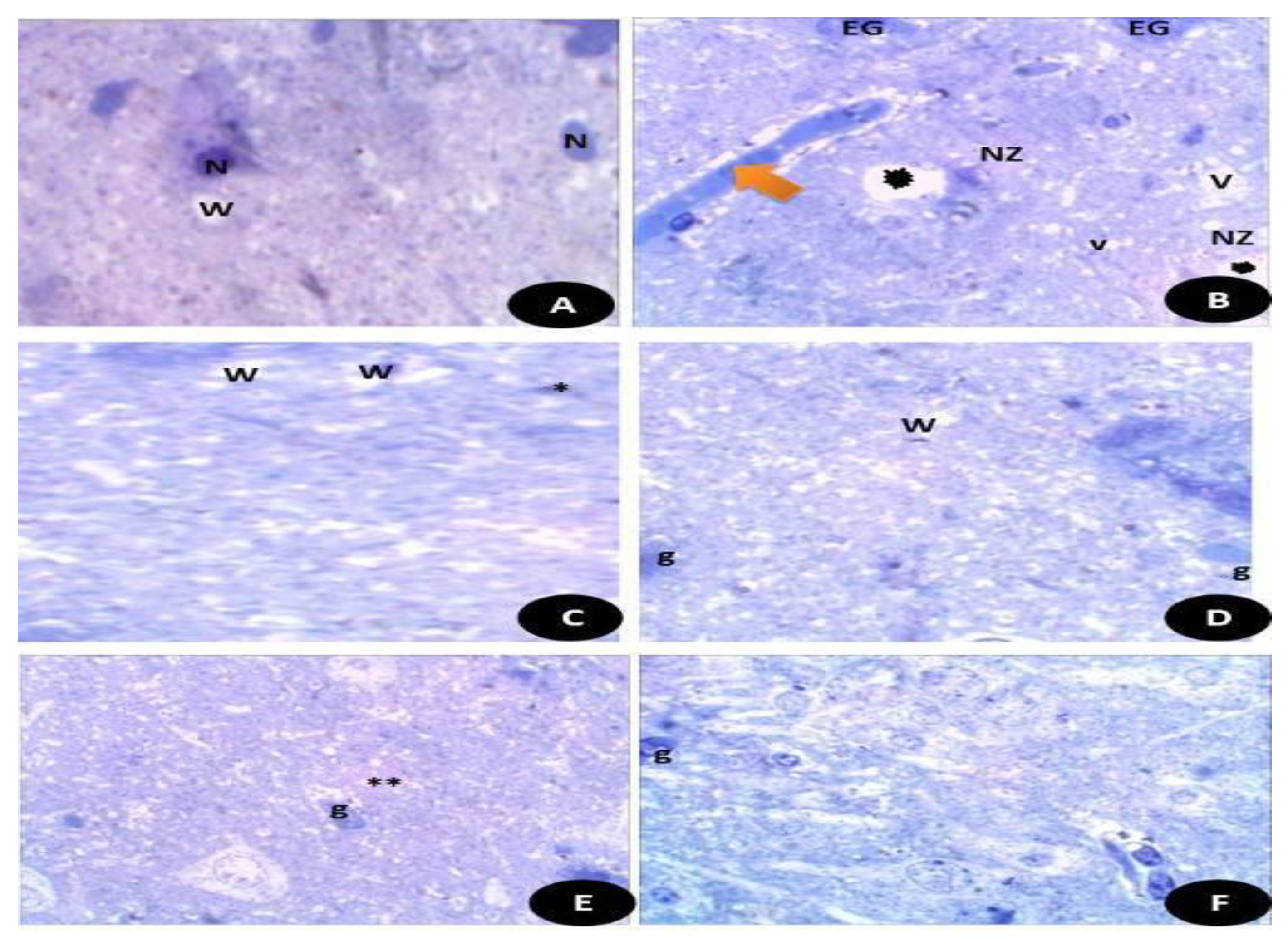
3.7.13. Semi Thin Sections of Testicular and Brain Tissues (Cerebral Cortex)
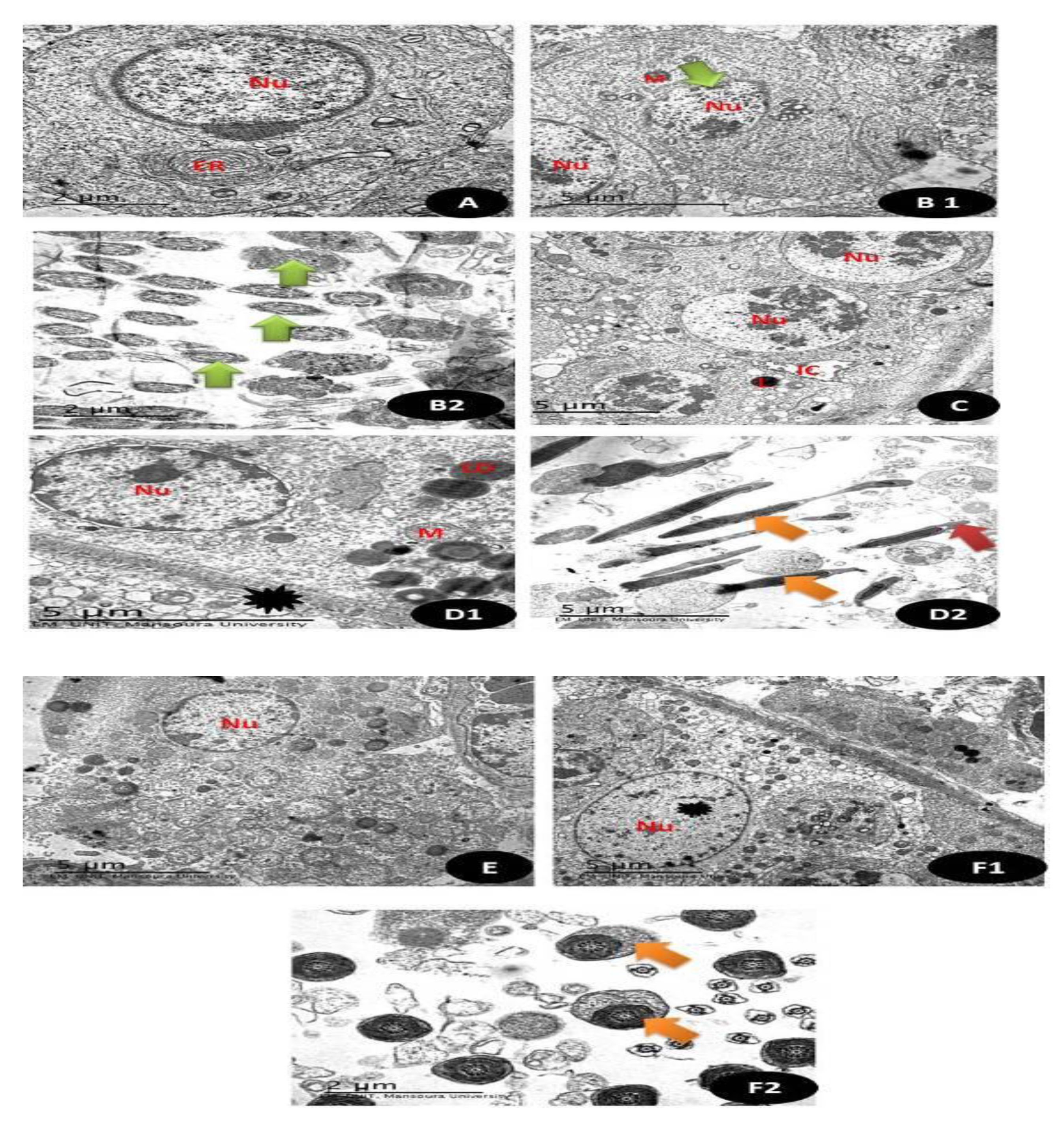
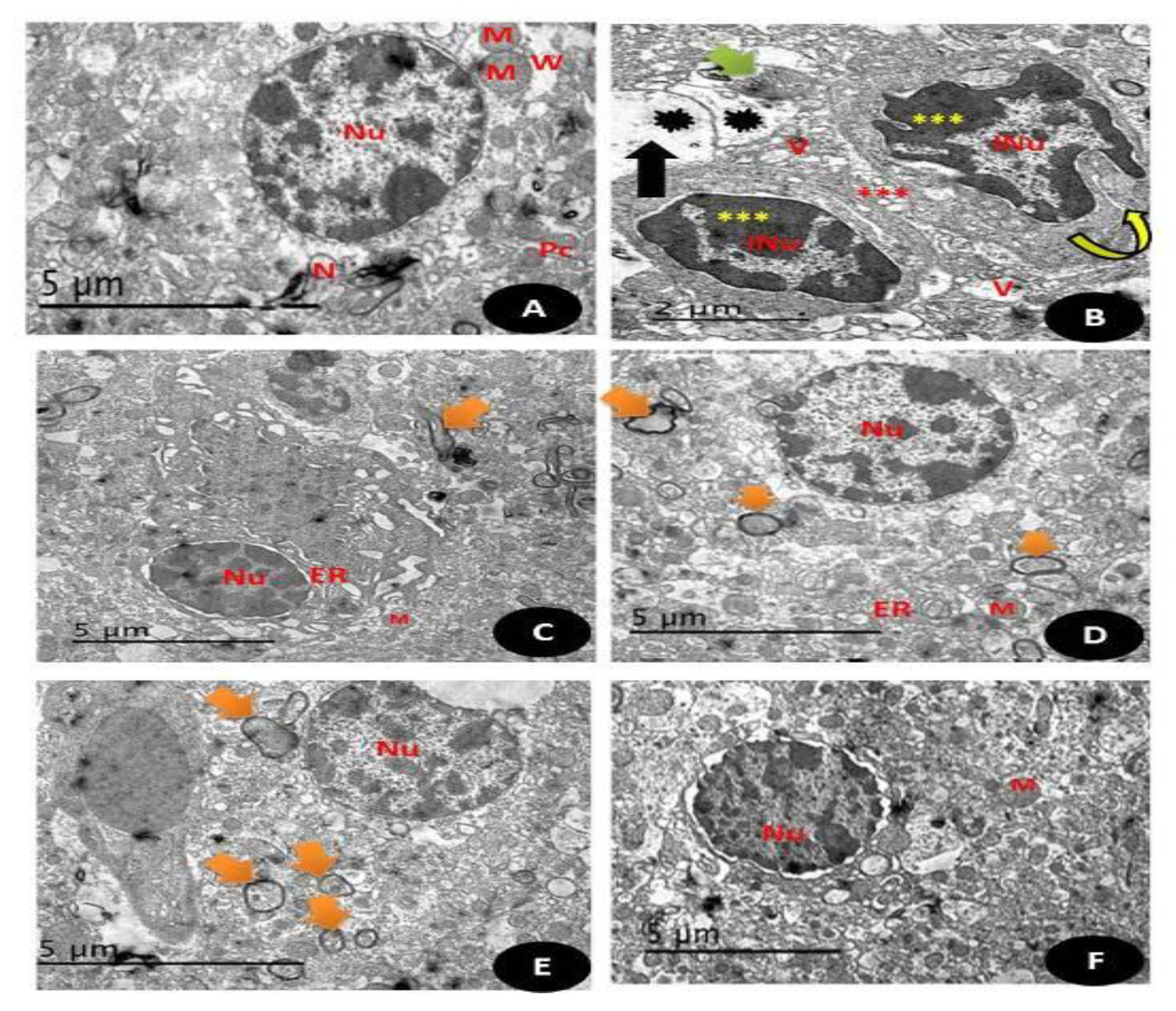
3.7.14. TEM Examination (Testicular and Brain Tissues)
3.7.15. Comet Assay
3.7.16. Mitochondrial Membrane Potential
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Betteridge, D.J. What is oxidative stress? Metabolism 2000, 49, 3–8. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P. Redefining oxidative stress. Antioxid. Redox Signal. 2006, 8, 1865–1879. [Google Scholar] [CrossRef]
- Bin, G.; Qing, G.; Zhan, W.; Shao, J.-B.; Liu, K.; Du, Z.-D.; Gong, S.-S. D-Galactose-induced oxidative stress and mitochondrial dysfunction in the cochlear basilar membrane: An in vitro aging model. Biogerontology 2020, 21, 311–323. [Google Scholar]
- Al-Baqami, N.M.; Hamza, R.Z. Synergistic antioxidant capacities of vanillin and chitosan nanoparticles against reactive oxygen species, hepatotoxicity, and genotoxicity induced by aging in male Wistar rats. Hum. Exp. Toxicol. 2021, 40, 183–202. [Google Scholar] [CrossRef] [PubMed]
- Barnham, K.J.; Masters, C.L.; Bush, A.I. Neurodegenerative diseases and oxidative stress. Nat. Rev. Drug Discov. 2004, 3, 205–214. [Google Scholar] [CrossRef]
- Karasek, M. Melatonin, human aging, and age-related diseases. Exp. Gerontol. 2004, 3, 1723–1729. [Google Scholar] [CrossRef]
- Sampath, K.; Sathiyaraj, S.; Jayabalakrishnan, C. Evaluation of DNA-binding, DNA cleavage, antioxidant and cytotoxic activity of mononuclear ruthenium(II) carbonyl complex of benzaldehyde 4-phenyl-3-thiosemicarbazones. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2013, 115, 287–296. [Google Scholar] [CrossRef]
- Carlotti, M.E.; Sapino, S.; Ugazio, E.; Caron, G. On the complexation of quercetin with methyl-b-cyclodextrin: Photostability and antioxidant studies. J. Incl. Phenom. Macrocycl. Chem. 2011, 70, 81–90. [Google Scholar] [CrossRef]
- Pinelo, M.; Manzocco, L.; Nunez, M.J.; Nicoli, C. Solvent effect on quercetin antioxidant capacity. Food Chem. 2004, 88, 201–207. [Google Scholar] [CrossRef]
- Calabrò, M.L.; Tommasini, S.; Donato, P.; Raneri, D.; Stancanelli, R.; Ficarra, P.; Ficarra, R.; Costa, C.; Catania, S.; Rustichelli, C.; et al. Effects of a- and b-cyclodextrin complexation on the physico-chemical properties and antioxidant activity of some 3-hydroxyflavones. J. Pharm. Biomed. Anal. 2004, 35, 365–377. [Google Scholar] [CrossRef]
- Rezende, B.A.; Cortes, S.F.; de Sousa, F.B.; Lula, I.S.; Schmitt, M.; Sinisterra, R.D.; Lemos, V.S. Complexation with b-cyclodextrin confers oral activity on the flavonoid dioclein. Int. J. Pharm. 2009, 367, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Mielcarek, J. Photochemical stability of the inclusion complexes formed by modified 1,4-dihydropyridine derivatives with b-cyclodextrin. J. Pharm. Biomed. Anal. 1997, 15, 681–686. [Google Scholar] [CrossRef]
- Albini, A.; Fasani, E. Photochemistry of drugs: An overview and practical problems. In Drugs: Photochemistry and Photostability; Albini, A., Fasani, E., Eds.; The Royal Society of Chemistry: Cambridge, UK, 1998; pp. 1–74. [Google Scholar]
- Xie, X.; Shen, Q.; Cao, L.; Chen, Y.; Ma, L.; Xiao, Q.; Yu, C.; Fu, Z. Depression caused by long-term stress regulates premature aging and is possibly associated with disruption of circadian rhythms in mice. Physiol. Behav. 2019, 199, 100–110. [Google Scholar] [CrossRef]
- Azam, M.; Warad, I.; Al-Resayes, S.; Shakir, M.; Ullah, M.F.; Ahmad, A.; Sarkar, F.H. A novel Ru (II) complex derived from hydroxydiamine as a potential antitumor agent: Synthesis and Structural Characterization. Inorg. Chem. Commun. 2012, 20, 252–258. [Google Scholar] [CrossRef]
- Ahn, G.-O.; Botting, K.J.; Patterson, A.V.; Ware, D.C.; Tercel, M.; Wilson, W.R. Radiolytic and cellular reduction of a novel hypoxia-activated cobalt (III) prodrug of a chloromethylbenzindoline DNA minor groove alkylator. Biochem. Pharmacol. 2006, 71, 1683–1694. [Google Scholar] [CrossRef]
- Levina, A.; Mitra, A.; Lay, P.A. Recent developments in ruthenium anticancer drugs. Metallomics 2009, 1, 458–470. [Google Scholar] [CrossRef]
- Anthony, E.J.; Bolitho, E.M.; Bridgewater, H.E.; Carter, O.W.L.; Donnelly, J.M.; Imberti, C.; Lant, E.C.; Lermyte, F.; Needham, R.J.; Palau, M.; et al. Metallodrugs are unique: Opportunities and challenges of discovery and development. Chem. Sci. 2020, 11, 12888–12917. [Google Scholar] [CrossRef]
- Hamza, R.Z.; Diab, A.A. Testicular protective and antioxidant effects of selenium nanoparticles on Monosodium glutamate-induced testicular composition alterations in male mice. Toxicol. Rep. 2020, 7, 254–260. [Google Scholar] [CrossRef]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Gilgun-Sherki, Y.; Melamed, E.; Offen, D. Oxidative stress induced-neurodegenerative diseases: The need for antioxidants that penetrate the blood brain barrier. Neuropharmacology 2001, 40, 959–975. [Google Scholar] [CrossRef]
- Guo, Y.; Bruno, R.S. Endogenous and exogenous mediators of quercetin bioavailability. J. Nutr. Biochem. 2015, 26, 201–210. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kang, J.S.; Park, J.H.Y.; Lee, Y.J.; Choi, J.S.; Kang, Y.H. Polyphenolic flavonoidsdiffer in their antiapoptotic efficacy in hydrogen peroxide-treated humanvascular endothelial cells. J. Nutr. 2003, 133, 985–991. [Google Scholar] [CrossRef] [Green Version]
- Hamza, R.Z.; AL-Thubiti, E.H.; Abd Al-Kader, S. The Antioxidant Activity of Quercetin and its Effect on Acrylamide Hepatotoxicity in Liver of Rats. Lat. Am. J. Pharm. 2019, 38, 2057–2062. [Google Scholar]
- Zaplatic, E.; Bule, M.; Shah, S.Z.A.; Uddin, M.S.; Niaz, K. Molecular mechanisms underlying protective role of quercetin in attenuating Alzheimer’s disease. Life Sci. 2019, 224, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Zribi, N.; Chakroun, N.F.; Abdallah, F.B.; Elleuch, H.; Sellami, A.; Gargouri, J.; Rebai, T.; Fakhfakh, F.; Keskes, L.A. Effect of freezing–thawing process and quercetin on human sperm survival and DNA integrity. Cryobiology 2012, 65, 326–331. [Google Scholar] [CrossRef]
- Silva, E.C.; Cajueiro, J.F.; Silva, S.V. Effect of antioxidants resveratrol and quercetin on in vitro evaluation of frozen ram sperm. Theriogenology 2012, 77, 1722–1726. [Google Scholar] [CrossRef]
- Thuwanut, P.; Chatdarong, K.; Johannisson, A. Cryopreservation of epididymal cat spermatozoa: Effects of in vitro antioxidative enzymes supplementation and lipid peroxidation induction. Theriogenology 2010, 73, 1076–1087. [Google Scholar] [CrossRef]
- Dhanya, G.; Uma Maheswari, P.; MeeraSheriffa Begum, K.M. Synergic formulation of onion peel quercetin loaded chitosan-cellulose hydrogel with green zinc oxide nanoparticles towards controlled release, biocompatibility, antimicrobial and anticancer activity. Int. J. Biol. Macromol. 2019, 132, 784–794. [Google Scholar]
- Refat, M.S.; Hamza, R.Z.; Adam, A.M.A.; Saad, H.A.; Gobouri, A.A.; Al-Salmi, F.A.; Altalhi, T.; El-Megharbel, S.M. Synthesis of N,N′-bis(1,5-dimethyl-2-phenyl-1,2-dihydro-3-oxopyrazol-4-yl) sebacamide that ameliorate osteoarthritis symptoms and improve bone marrow matrix structure and cartilage alterations induced by monoiodoacetate in the rat model: “Suggested potent anti-inflammatory agent against COVID-19”. Hum. Exp. Toxicol. 2021, 40, 325–341. [Google Scholar] [PubMed]
- Zhang, Z.F.; Fan, S.H.; Zheng, Y.L. Purple sweet potato color attenuates oxidative stress and inflammatory response induced by D-galactose in mouse liver. Food Chem. Toxicol. 2009, 47, 496–501. [Google Scholar] [CrossRef]
- Kalantari, H.; Foruozandeh, H.; Khodayar, M.J.; Siahpoosh, A.; Saki, N.; Kheradmand, P. Antioxidant and hepatoprotective effects of Capparis spinosa L. fractions and Quercetin on tert-butyl hydroperoxide- induced acute liver damage in mice. J. Tradit. Complementary Med. 2018, 8, 120–127. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Europ. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Meth. Enzymol. 1984, 105, 121–126. [Google Scholar]
- Couri, D.; Abdel-Rahman, M.S. Effect of chlorine dioxide and metabolites on glutathione-dependent system in rat, mouse and chicken blood. J. Environ. Pathol. Toxicol. 1980, 3, 451–460. [Google Scholar]
- Hafeman, D.G.; Sunde, R.A.; Hoekstra, W.G. Effect of dietary selenium on erythrocyte and liver glutathione peroxidase in the rat. J. Nutrit. 1974, 104, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Wright, H.L.; Moots, R.J.; Bucknall, R.C.; Edwards, S.W. Review Neutrophil function in inflammation and inflammatory diseases. Rheumatology 2010, 49, 1618–1631. [Google Scholar] [CrossRef] [Green Version]
- Gabe, M. Techniques Histologiques [Histological Techniques]; Masson Publisher: Paris, France, 1968. [Google Scholar]
- Weakley, B.; Beginner, S. Handbook in Biological Transmission Electron Microscopy, 2nd ed.; Churchill Livingstone: London, UK, 1981. [Google Scholar]
- Collins, A.R.; Dunsinka, M. Oxidation of cellular DNA measured with the comet assay. In Methods in Molecular Biology: Oxidative Stress Biomarkers and Antioxidant Protocols; Armstrong, D., Ed.; Humana Press: Totowa, NJ, USA, 2002; pp. 147–159. [Google Scholar]
- Donato, M.T.; Tolosa, L.; Gómez-Lechón, M.J. Culture and functional characterization of human hepatoma HepG2 cells. In Protocols in In Vitro Hepatocyte Research. Methods in Molecular Biology (Methods and Protocols); Humana Press: New York, NY, USA, 2015; pp. 77–93. [Google Scholar]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Lisa, M. Cell viability assays. In Assay Guidance Manual; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2016. [Google Scholar]
- Malek, S.N.A.; Shin, S.K.; Wahab, N.A.; Yaacob, H. Cytotoxic components of Pereskia bleo (Kunth) DC.(Cactaceae) leaves. Molecules 2009, 14, 1713–1724. [Google Scholar] [CrossRef]
- Lugli, E.; Roederer, M.; Cossarizza, A. Data analysis in flow cytometry: The future just started. Cytom. Part A 2010, 77, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Cottet-Rousselle, C.; Ronot, X.; Leverve, X.; Mayol, J.F. Cytometric assessment of mitochondria using fluorescent probes. Cytom. Part A 2011, 79, 405–425. [Google Scholar] [CrossRef]
- Han, Y.; Lo, Y.H. Imaging Cells in Flow Cytometer Using Spatial-Temporal Transformation. Sci. Rep. 2015, 5, 13267. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, N.; Ami, Y.; Terao, K.; Tashiro, F.; Honda, M. Upgrading of flow cytometric analysis for absolute counts, cytokines and other antigenic molecules of cynomolgus monkeys (Macaca fascicularis) by using anti-human cross-reactive antibodies. Exp. Anim. 2000, 49, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Van Engeland, M.; Nieland, L.J.W.; Ramaekers, F.C.S.; Schutte, B.; Reutelingsperger, C.P.M. Annexin V-affinity assay: A review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry 1998, 31, 1–9. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Rieger, A.M.; Nelson, K.L.; Konowalchuk, J.D.; Barreda, D.R. Modified Annexin V/Propidium Iodide Apoptosis Assay for Accurate Assessment of Cell Death. J. Vis. Exp. 2011, JoVE, 2597. [Google Scholar] [CrossRef] [PubMed]
- IBM. IBM SPSS Statistics for Windows, Version 27; IBM Corp.: Armonk, NY, USA, 2020; Available online: http://www-01.ibm.com/support/docview.wss?uid=swg27049428 (accessed on 12 August 2020).
- Dean, A.; Sullivan, K.; Soe, M. OpenEpi: Open-Source Epidemiologic Statistics for Public Health. 2013. Available online: https://www.OpenEpi.com (accessed on 6 April 2013).
- Petrie, A.; Sabin, C. Medical Statistics at a Glance, 3rd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2009; Volume 23. [Google Scholar] [CrossRef]
- Geary, W.J. The Use of Conductivity Measurements in Organic Solvents for the Characterisation of Coordination Compounds. Coord. Chem. Rev. 1971, 7, 81–122. [Google Scholar] [CrossRef]
- Nakamoto, K. Infrared and Raman Spectra of Inorganic and Coordination Compounds, 4th ed.; Wiley: New York, NY, USA, 1986. [Google Scholar]
- Deacon, G.B.; Philips, R.J. Relationships between the carbon-oxygen stretching frequencies of carboxylato complexes and the type of carboxylate coordination. Coord. Chem. Rev. 1980, 33, 227–250. [Google Scholar] [CrossRef]
- Sertçelik, M.; Özbek, F.E.; Taslimi, P.; Durman, M.; Özdemir, M.; Yalçın, B.; Necefoğlu, H.; Hökelek, T. Synthesis, spectroscopic characterization, crystal structure, density functional theory studies and biological properties of coordination complex Ni(II) 2-fluorobenzoate with 3-hydroxypyridine. Appl. Organomet. Chem. 2020, 34, 9. [Google Scholar] [CrossRef]
- Figgis, B.N. Introduction to Ligand Fields; Interscience Publishers: Geneva, Switzerland, 1966. [Google Scholar]
- Ullah, R.; Jo, M.H.; Riaz, M.; Alam, S.I.; Saeed, K.; Ali, W.; Rehman, I.U.; Ikram, M.; Kim, M.O. Glycine, the smallest amino acid, confers neuroprotection against D-galactose-induced neurodegeneration and memory impairment by regulating c-Jun N-terminal kinase in the mouse brain. J. Neuroinflamm. 2020, 17, 303. [Google Scholar] [CrossRef] [PubMed]
- Ali, T.; Badshah, H.; Kim, T.H.; Kim, M.O. Melatonin attenuates D-galactose induced memory impairment, neuroinflammation and neurodegeneration via RAGE/NF-K B/JNK signaling pathway in aging mouse model. J. Pineal Res. 2015, 58, 71–85. [Google Scholar] [CrossRef]
- Thazin, S.; Wasana, P.; Nipon, C.; Siriporn, C.C. Role of D-galactose-induced brain aging and its potential used for therapeutic interventions. Exp. Gerontol. 2018, 101, 13–36. [Google Scholar]
- Acosta, P.B.; Gross, K.C. Hidden sources of galactose in the environment. Eur. J. Pediatr. 1995, 154, S87–S92. [Google Scholar] [CrossRef]
- Boespflug, E.L.; Eliassen, J.C.; Dudley, J.A.; Shidler, M.D.; Kalt, W.; Summer, S.S.; Stein, A.L.; Stover, A.N.; Krikorian, R. Enhanced neural activation with blueberry supplementation in mild cognitive impairment. Nutr. Neurosci. 2017, 21, 297–305. [Google Scholar] [CrossRef]
- Coelho, A.I.; Berry, G.T.; Rubio-Gozalbo, M.E. Galactose metabolism and health. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Cura, A.J.; Carruthers, A. Role of monosaccharide transport proteins in carbohydrate assimilation, distribution, metabolism, and homeostasis. Compr. Physiol. 2012, 2, 863–914. [Google Scholar] [PubMed] [Green Version]
- Berry, G.T. Classic galactosemia and clinical variant galactosemia. In GeneReviews®; Pagon, R.A., Adam, M.P., Ardinger, H.H., Wallace, S.E., Amemiya, A., Bean, L.J.H., Bird, T.D., Ledbetter, N., Mefford, H.C., Smith, R.J.H., et al., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Morava, E. Galactose supplementation in phosphoglucomutase-1 deficiency; review and outlook for a novel treatable CDG. Mol. Genet. Metab. 2014, 112, 275–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydın, A.F.; Çoban, J.; Doğan-Ekici, I.; Betül-Kalaz, E.; Doğru-Abbasoğlu, S.; Uysal, M. Carnosine and taurine treatments diminished brain oxidative stress and apoptosis in D-galactose aging model. Metab. Brain Dis. 2016, 31, 337–345. [Google Scholar] [CrossRef]
- Rehman, S.U.; Shah, S.A.; Ali, T.; Chung, J.I.; Kim, M.O. Anthocyanins Reversed D-Galactose-Induced Oxidative Stress and Neuroinflammation Mediated Cognitive Impairment in Adult Rats. Mol. Neurobiol. 2017, 54, 255–271. [Google Scholar] [CrossRef]
- Jiang, Q.; Isquith, J.; Zipeto, M.A.; Diep, R.H.; Pham, J.; Santos, N.D.; Reynoso, E.; Chau, J.; Leu, H.; Lazzari, E.; et al. Hyper-Editing of Cell-Cycle Regulatory and Tumor Suppressor RNA Promotes Malignant Progenitor Propagation. Cancer Cell 2019, 35, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.-W.; Sun, Q.-X.; Liu, Y.-H.; Gao, W.; Li, Y.-H.; Lu, K.; Wang, Z. Chronic administration of Liu Wei Dihuang protects rat’s brain against D-galactose-induced impairment of cholinergic system. Acta Physiol. Sin. 2011, 63, 245–255. [Google Scholar]
- Wu, W.; Wang, X.; Xiang, Q.; Meng, X.; Peng, Y.; Du, N.; Liu, Z.; Sun, Q.; Wang, C.; Liu, X. Astaxanthin alleviates brain aging in rats by attenuating oxidative stress and increasing BDNF levels. Food Funct. 2014, 5, 158–166. [Google Scholar] [CrossRef]
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Rehman, S.U.; Ahmad, A.; Yoon, G.-H.; Khan, M.; Abid, M.N.; Kim, M.O. Inhibition of c-Jun N-Terminal Kinase Protects Against Brain Damage and Improves Learning and Memory After Traumatic Brain Injury in Adult Mice. Cereb. Cortex 2018, 28, 2854–2872. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, Q.; Zhuang, S.; Wen, Y.; Cheng, W.; Zeng, Z.; Jiang, T.; Tang, C. Effect of Anoectochilus roxburghii flavonoids extract on H2O2—Induced oxidative stress in LO2 cells and D-gal induced aging mice model. J. Ethnopharmacol. 2020, 254, 112670. [Google Scholar] [CrossRef]
- Perruchot, M.; Gondret, F.; Robert, F.; Dupuis, E.; Quesnel, H.; Dessauge, F. Effect of the flavonoid baicalin on the proliferative capacity of bovine mammary cells and their ability to regulate oxidative stress. PeerJ 2019, 7, e6565. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, H.; Liu, B.; Shan, Y.; Zhou, H.; Qi, X.; Wu, W.; Jia, L. Combination of chick embryo and nutrient mixture prevent D-galactose-induced cognitive deficits, immune impairment and oxidative stress in aging rat model. Sci. Rep. 2019, 9, 4092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mede, T.; Jäger, M.; Schubert, U.S. “Chemistry-on-the-complex”: Functional Ru IIpolypyridyl-type sensitizers as divergent building blocks. Chem. Soc. Rev. 2018, 47, 7577–7627. [Google Scholar] [CrossRef] [PubMed]
- Lucio, G.C.; Garrick, J.M.; Roquè, P.J.; Pellacani, C. Mechanisms of Neuroprotection by Quercetin: Counteracting Oxidative Stress and More. Oxidative Med. Cell. Longev. 2016, 2016, 2986796. [Google Scholar]
- Sharma, V.; Mishra, M.; Ghosh, S.; Tewari, R.; Basu, A.; Seth, P.; Sen, E. Modulation of interleukin-1 mediated inflammatory response in human astrocytes by flavonoids: Implications in neuroprotection. Brain Res. Bull. 2007, 73, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.A.; Ackerman, S.L. Oxidative stress, cell cycle, and neurodegeneration. J. Clin. Investig. 2003, 111, 785–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Sun, L.; Liu, Z.; Wang, H.; Xu, C. Protection afforded by quercetin against H2O2-induced apoptosis on PC12 cells via activating PI3K/Akt signal pathway. J. Recept. Signal Transduct. 2016, 36, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.J.; Jeong, J.H.; Bing, G.; Park, E.S.; Chae, J.S.; Yen, T.P.H.; Kim, W.K.; Wie, M.B.; Jung, B.D.; Kim, H.J.; et al. Kainate-induced mitochondrial oxidative stress contributes to hippocampal degeneration in senescence-accelerated mice. Cell Signal 2008, 20, 645–658. [Google Scholar] [CrossRef]
- Fu, Y.; Tang, M.; Bai, L.; Fan, Y. The role of apoptosis inducing factor in the apoptosis of retinal pigment epithelium cells induced by oxidative stress. Cell. Mol. Biol. 2016, 62, 36–41. [Google Scholar]
- Moreira, A.C.; Branco, A.F.; Sampaio, S.F.; Cunha-Oliveira, T.; Martins, T.R.; Holy, J.; Oliveira, P.J.; Sardão, V.A. Mitochondrial apoptosis-inducing factor is involved in doxorubicin induced toxicity on H9c2 cardiomyoblasts. Biochim. Biophys. Acta 2014, 1842, 2468–2478. [Google Scholar] [CrossRef] [Green Version]
- Hamza, R.Z.; EL-Megharbel, S.M.; Altalhi, T.; Gobouri, A.A.; Alrogi, A.A. Hypolipidemic and hepatoprotective synergistic effects of selenium nanoparticles and vitamin. E against acrylamide-induced hepatic alterations in male albino mice. Appl Organometal Chem. 2020, 34, e5458. [Google Scholar] [CrossRef]
- Refat, M.S.; Hamza, R.Z.; Adam, A.M.A.; Saad, H.A.; Gobouri, A.A.; Al-Harbi, F.S.; Al-Salmi, F.A.; Altalhi, T.; El-Meharbel, S.M. Quercetin/Zinc complex and stem cells: A new drug therapy to ameliorate glycometabolic control and pulmonary dysfunction in diabetes mellitus: Structural characterization and genetic studies. PLoS ONE 2021, 16, e0246265. [Google Scholar] [CrossRef]
- Hamza, R.Z.; Al-Baqami, N.M. Testicular protective effects of ellagic acid on monosodium glutamate-induced testicular structural alterations in male rats. Ultrastruct. Pathol. 2019, 43, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Hamza, R.A.; Al-Eisa, R.A.; El-Shenawy, N.S. Efficacy of Mesenchymal Stem Cell and Vitamin D in the Treatment of Diabetes Mellitus Induced in a Rat Model: Pancreatic Tissues. Coatings 2021, 11, 317. [Google Scholar] [CrossRef]
- Pavanato, A.; Tuñón, M.J.; Sánchez-Campos, S.; Marroni, C.A.; Llesuy, S.; González-Gallego, J.; Marroni, N. Effects of quercetin onliver damage in rats with carbon tetrachloride-induced cirrhosis. Dig. Dis. Sci. 2003, 48, 824–829. [Google Scholar] [CrossRef]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Saw, C.L.; Guo, Y.; Yang, A.Y.; Paredes-Gonzalez, X.; Ramirez, C.; Pung, D.; Kong, A.-N.T. The berry constituents quercetin, kaempferol, and pterostilbene synergistically attenuate reactive oxygen species: Involvement of the Nrf2-ARE signaling pathway. Food Chem. Toxicol. 2014, 72, 303–311. [Google Scholar] [CrossRef]
- Ramyaa, P.; Krishnaswamy, R.; Padma, V.V. Quercetin modulates OTA-induced oxidative stress and redox signalling in HepG2 cells—up regulation of Nrf2 expression and down regulation of NF-kappaB and COX-2. Biochim. Biophys. Acta 2014, 1840, 681–692. [Google Scholar] [CrossRef]
- Uddin, M.S.; al Mamun, A.; Takeda, S.; Sarwar, M.S.; Begum, M.M. Analyzing the chance of dementia amid geriatric people: A cross-sectional pilot study in Bangladesh. Psychogeriatrics 2019, 19, 87–94. [Google Scholar] [CrossRef]
- Van Dyck, C.H. Anti-amyloid-β monoclonal antibodies for Alzheimer’s disease: Pitfalls and promise. Biol. Psychiatry 2018, 83, 311–319. [Google Scholar] [CrossRef] [Green Version]
- Ghanemi, A. Alzheimer’s disease therapies: Selected advances and future perspectives. Alex. J. Med. 2015, 51, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Banji, O.J.; Banji, D.; Ch, K. Curcumin and hesperidin improve cognition by suppressing mitochondrial dysfunction and apoptosis induced by D-galactose in rat brain. Food Chem. Toxicol. 2014, 74, 51–59. [Google Scholar] [CrossRef]
- Hsieh, H.M.; Wu, W.M.; Hu, M.L. Soy isoflavones attenuate oxidative stress and improve parameters related to aging and Alzheimer’s disease in C57BL/6J mice treated with D-galactose. Food Chem. Toxicol. 2009, 47, 625–632. [Google Scholar] [CrossRef]
- Golubev, A.; Hanson, A.D.; Gladyshev, V.N. Non-enzymatic molecular damage as a prototypic driver of aging. J. Biol. Chem. 2017, 292, 6029–6038. [Google Scholar] [CrossRef] [Green Version]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
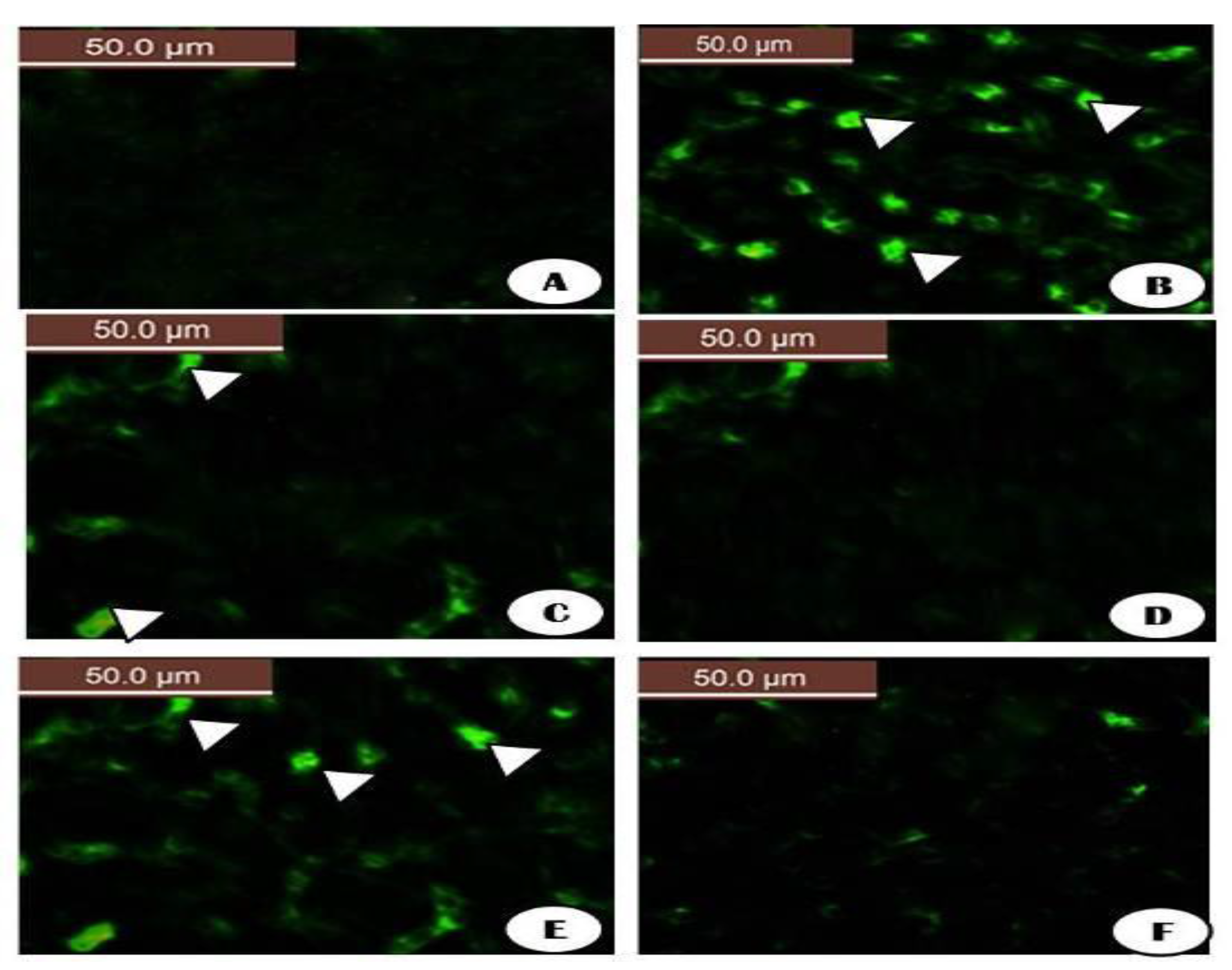{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| P53 | 5′-GTATTTGGATGACAGAAACACT-3′ | 5′-GTTACACATGTAGTTGTAGTGG-3′ |
| Bax | 5′-GAACTGGACAGTAACATGGAG-3′ | 5′-TTTGCTGGCAAAGTAGAAAAG-3′ |
| Bcl-2 | 5′-ATTGTGGCCTTCTTTGAGTT-3′ | 5′-GTTCAGGTACTCAGTCATCC-3′ |
| mTOR | 5′-TGAATTCAGCCAAGTTTAAGAG-3′ | 5′-TTCAATGCTGCATTTGTGTC-3′ |
| RALA | 5′-ATGAATTGGAACTGCAATGAAA-3′ | 5′-TCAGGACCATGATAAATGTAGG-3′ |
| PI3K | 5′-TCAGAGTTACTGTTTCAGAACA-3′ | 5′-GTAACATTCGAAGATCAAGACC-3′ |
| c-MET | 5′-CTGGTGTTGTCTCAATATCAAC-3′ | 5′-ATATCAGAGTCCCCACTAGTTA-3′ |
| Nrf2 | 5′-CAC ATT CCC AAA CAA GAT GC-3′ | 5′-TCT TTT TCC AGCGAG GAG AT-3′ |
| mTOR | RALA | PI3K | c-MET | |
|---|---|---|---|---|
| Group 2 | 0.82 ± 0.06 | 0.88 ± 0.04 | 0.80 ± 0.04 | 2.31 ± 0.10 |
| Group 3 | 0.54 ± 0.08 | 0.53 ± 0.02 | 0.55 ± 0.04 | 0.80 ± 0.04 |
| Group 4 | 0.22 ± 0.04 | 0.15 ± 0.04 | 0.20 ± 0.05 | 0.30 ± 0.10 |
| Group 5 | 0.52 ± 0.04 | 0.62 ± 0.12 | 0.60 ± 0.02 | 1.20 ± 0.02 |
| Group 6 | 0.38 ± 0.06 | 0.40 ± 0.16 | 0.55 ± 0.02 | 0.79 ± 0.16 |
| Post Hoc Power Analysis | ||||
| Groups 2 vs. 3 | 100% | 100% | 100% | 100% |
| Groups 2 vs. 4 | 100% | 100% | 100% | 100% |
| Groups 2 vs. 5 | 100% | 100% | 100% | 100% |
| Groups 2 vs. 6 | 100% | 100% | 100% | 100% |
| Groups 3 vs. 4 | 100% | 100% | 100% | 100% |
| Groups 3 vs. 5 | 13.7% | 81.7% | 99.1% | 100% |
| Groups 3 vs. 6 | 100% | 87.8% | 1.1% | 3.6% |
| Groups 4 vs. 5 | 100% | 100% | 100% | 100% |
| Groups 4 vs. 6 | 100% | 100% | 100% | 100% |
| Groups 5 vs. 6 | 100% | 98.9% | 100% | 100% |
| CAT (U/g) | SOD (U/g) | GRx (U/g) | MDA (U/g) | GPx (U/g) | |
|---|---|---|---|---|---|
| Group 1 | 5.0 ± 1.20 | 10.4 ± 1.69 | 7.7 ±1.01 | 4.4 ±0.52 | 14.5 ±1.69 |
| Group 2 | 1.2 ± 0.02 | 4.0 ± 0.57 | 2.0 ± 0.68 | 34.0 ± 4.36 | 3.6 ± 0.87 |
| Group 3 | 4.7 ± 0.85 | 10.0 ± 1.65 | 7.3 ± 1.02 | 4.0 ± 0.78 | 14.0 ± 3.02 |
| Group 4 | 5.3 ± 0.63 | 11.4 ± 2.01 | 8.3 ± 1.06 | 3.7 ± 0.65 | 15.4 ± 2.54 |
| Group 5 | 3.3 ± 0.45 | 7.3 ± 0.98 | 5.4 ± 0.69 | 18.4 ± 2.36 | 9.3 ± 1.02 |
| Group 6 | 4.8 ± 0.87 | 9.3 ± 1.54 | 7.0 ± 1.57 | 11.0 ± 1.98 | 12.4 ± 1.98 |
| Post Hoc Power Analysis | |||||
| Groups 1 vs. 2 | 100% | 100% | 100% | 100% | 100% |
| Groups 1 vs. 3 | 12.1% | 9.5% | 18.9% | 37.9% | 7.9% |
| Groups 1 vs. 4 | 13.5% | 31.5% | 35.5% | 90.3% | 20.7% |
| Groups 1 vs. 5 | 99.9% | 100% | 100% | 100% | 100% |
| Groups 1 vs. 6 | 7.3% | 46.2 | 30.6% | 100% | 87.8% |
| Groups 2 vs. 3 | 100% | 100% | 100% | 100% | 100% |
| Groups 2 vs. 4 | 100% | 100% | 100% | 100% | 100% |
| Groups 2 vs. 5 | 100% | 100% | 100% | 100% | 100% |
| Groups 2 vs. 6 | 100% | 100% | 100% | 100% | 100% |
| Groups 3 vs. 4 | 59.4% | 55.0% | 75.0% | 20.7% | 27.9% |
| Groups 3 vs. 5 | 99.9% | 99.9% | 100% | 100% | 100% |
| Groups 3 vs. 6 | 4.6% | 22.4% | 8.9% | 100% | 40.4% |
| Groups 4 vs. 5 | 100% | 100% | 100% | 100% | 100% |
| Groups 4 vs. 6 | 43.8% | 89.5% | 75.7% | 100% | 95.0% |
| Groups 5 vs. 6 | 100% | 98.9% | 95.1% | 100% | 100% |
| CAT (U/g) | SOD (U/g) | GRx (U/g) | MDA (U/g) | GPx (U/g) | |
|---|---|---|---|---|---|
| Group 1 | 4.7 ± 0.47 | 12.6 ± 1.58 | 6.4 ± 1.69 | 7.3 ± 1.25 | 17.0 ± 2.58 |
| Group 2 | 1.2 ± 0.02 | 4.4 ± 1.35 | 1.4 ± 0.53 | 50.3 ± 3.52 | 4.5 ± 0.69 |
| Group 3 | 4.5 ± 0.85 | 12.6 ± 2.36 | 6.0 ± 1.66 | 7.0 ± 1.52 | 16.1 ± 2.01 |
| Group 4 | 5.1 ± 0.63 | 13.3 ± 2.98 | 6.4 ± 1.36 | 7.0 ± 1.02 | 17.0 ± 2.65 |
| Group 5 | 3.5 ± 0.45 | 7.7 ± 1.25 | 4.4 ± 0.69 | 24.4 ± 2.65 | 9.3 ± 1.24 |
| Group 6 | 4.01 ± 0.87 | 10.7 ± 2.69 | 6.0 ± 0.69 | 14.0 ± 2.25 | 13.6 ± 2.58 |
| Post hoc power analysis | |||||
| Groups 1 vs. 2 | 100% | 100% | 100% | 100% | 100% |
| Groups 1 vs. 3 | 12.2% | 1.1% | 9.5% | 8.4% | 18.6% |
| Groups 1 vs. 4 | 50.4% | 12.3% | 1.1% | 10.7% | 1.1% |
| Groups 1 vs. 5 | 100% | 100% | 98.9% | 100% | 100% |
| Groups 1 vs. 6 | 77.1% | 65.5% | 13.3% | 100% | 95.0% |
| Groups 2 vs. 3 | 100% | 100% | 100% | 100% | 100% |
| Groups 2 vs. 4 | 100% | 100% | 100% | 100% | 100% |
| Groups 2 vs. 5 | 100% | 100% | 100% | 100% | 100% |
| Groups 2 vs. 6 | 100% | 100% | 100% | 100% | 100% |
| Groups 3 vs. 4 | 59.4% | 10.6% | 10.7% | 1.1% | 18.1% |
| Groups 3 vs. 5 | 98.1% | 100% | 93.2% | 100% | 100% |
| Groups 3 vs. 6 | 35.7% | 53.8% | 1.1% | 100% | 84.2% |
| Groups 4 vs. 5 | 100% | 100% | 99.9% | 100% | 100% |
| Groups 4 vs. 6 | 97.8% | 71.2% | 17.3% | 100% | 94.5% |
| Groups 5 vs. 6 | 50.7% | 97.5% | 100% | 100% | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Refat, M.S.; Hamza, R.Z.; Adam, A.M.A.; Saad, H.A.; Gobouri, A.A.; Al-Salmi, F.A.; Altalhi, T.A.; El-Megharbel, S.M. Potential Therapeutic Effects of New Ruthenium (III) Complex with Quercetin: Characterization, Structure, Gene Regulation, and Antitumor and Anti-Inflammatory Studies (RuIII/Q Novel Complex Is a Potent Immunoprotective Agent). Crystals 2021, 11, 367. https://doi.org/10.3390/cryst11040367
Refat MS, Hamza RZ, Adam AMA, Saad HA, Gobouri AA, Al-Salmi FA, Altalhi TA, El-Megharbel SM. Potential Therapeutic Effects of New Ruthenium (III) Complex with Quercetin: Characterization, Structure, Gene Regulation, and Antitumor and Anti-Inflammatory Studies (RuIII/Q Novel Complex Is a Potent Immunoprotective Agent). Crystals. 2021; 11(4):367. https://doi.org/10.3390/cryst11040367
Chicago/Turabian StyleRefat, Moamen S., Reham Z. Hamza, Abdel Majid A. Adam, Hosam A. Saad, Adil A. Gobouri, Fawziah A. Al-Salmi, Tariq A. Altalhi, and Samy M. El-Megharbel. 2021. "Potential Therapeutic Effects of New Ruthenium (III) Complex with Quercetin: Characterization, Structure, Gene Regulation, and Antitumor and Anti-Inflammatory Studies (RuIII/Q Novel Complex Is a Potent Immunoprotective Agent)" Crystals 11, no. 4: 367. https://doi.org/10.3390/cryst11040367