
A Simple, Affordable, Rapid, Stabilized, Colorimetric, Versatile RT-LAMP Assay to Detect SARS-CoV-2

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Specimens
2.2. RNA Isolation and RT-qPCR Amplification
2.3. Viral RNA-Positive Control and Patients’ RNA Samples Selected
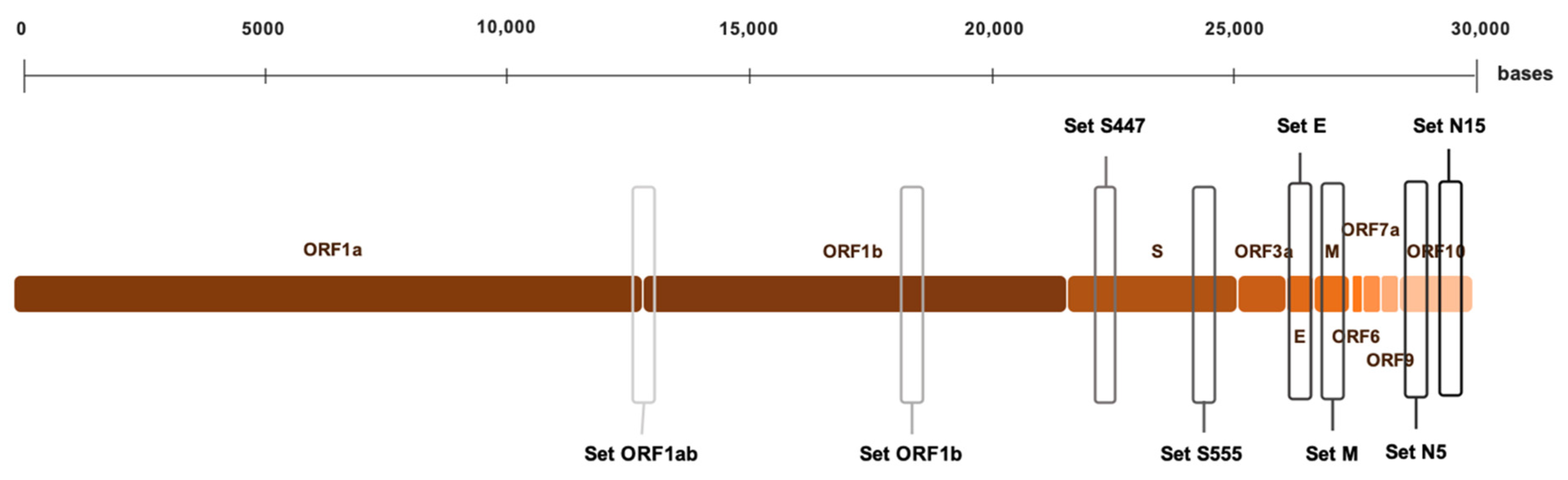
2.4. RT-LAMP Primer Design
2.5. RT-LAMP Reaction
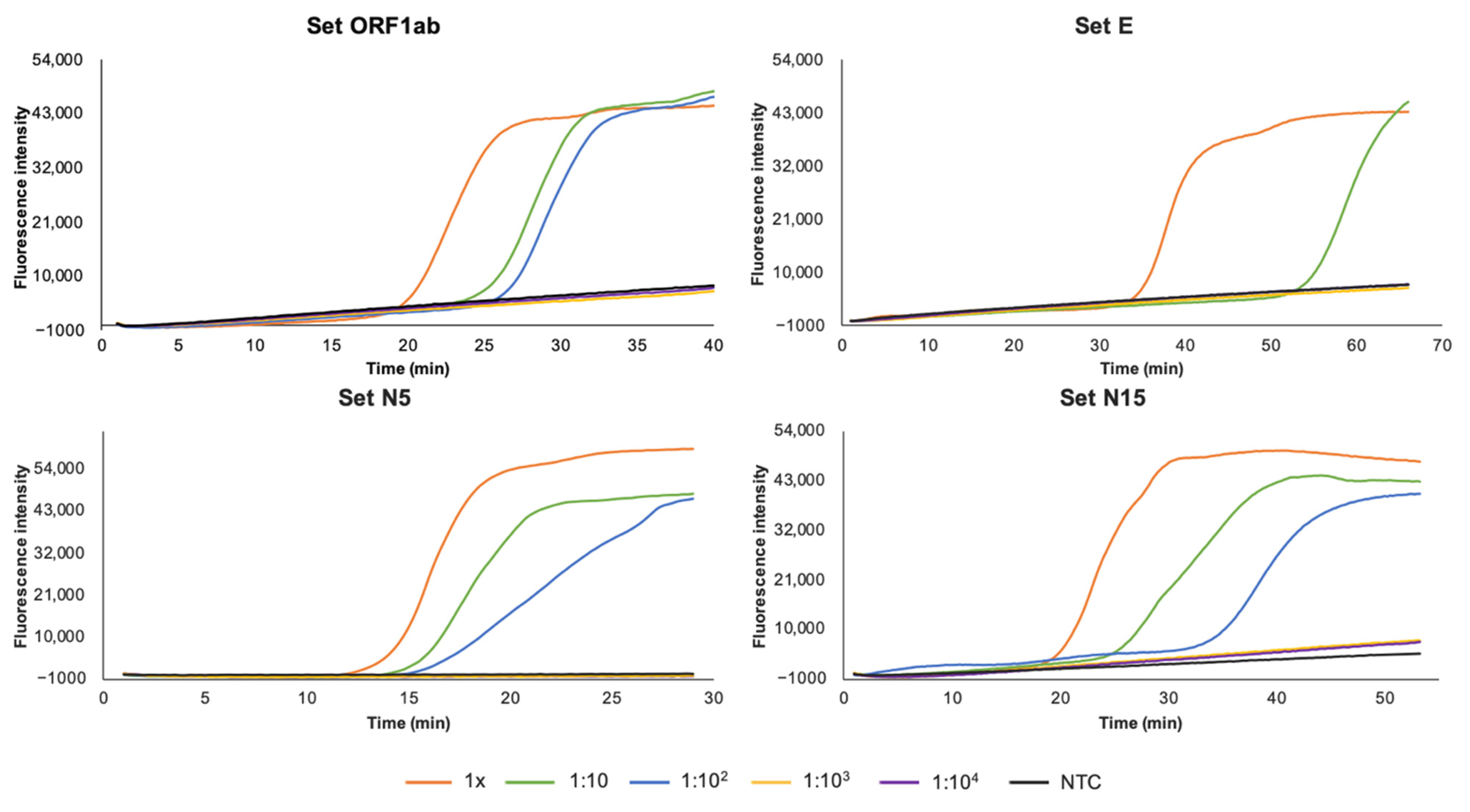
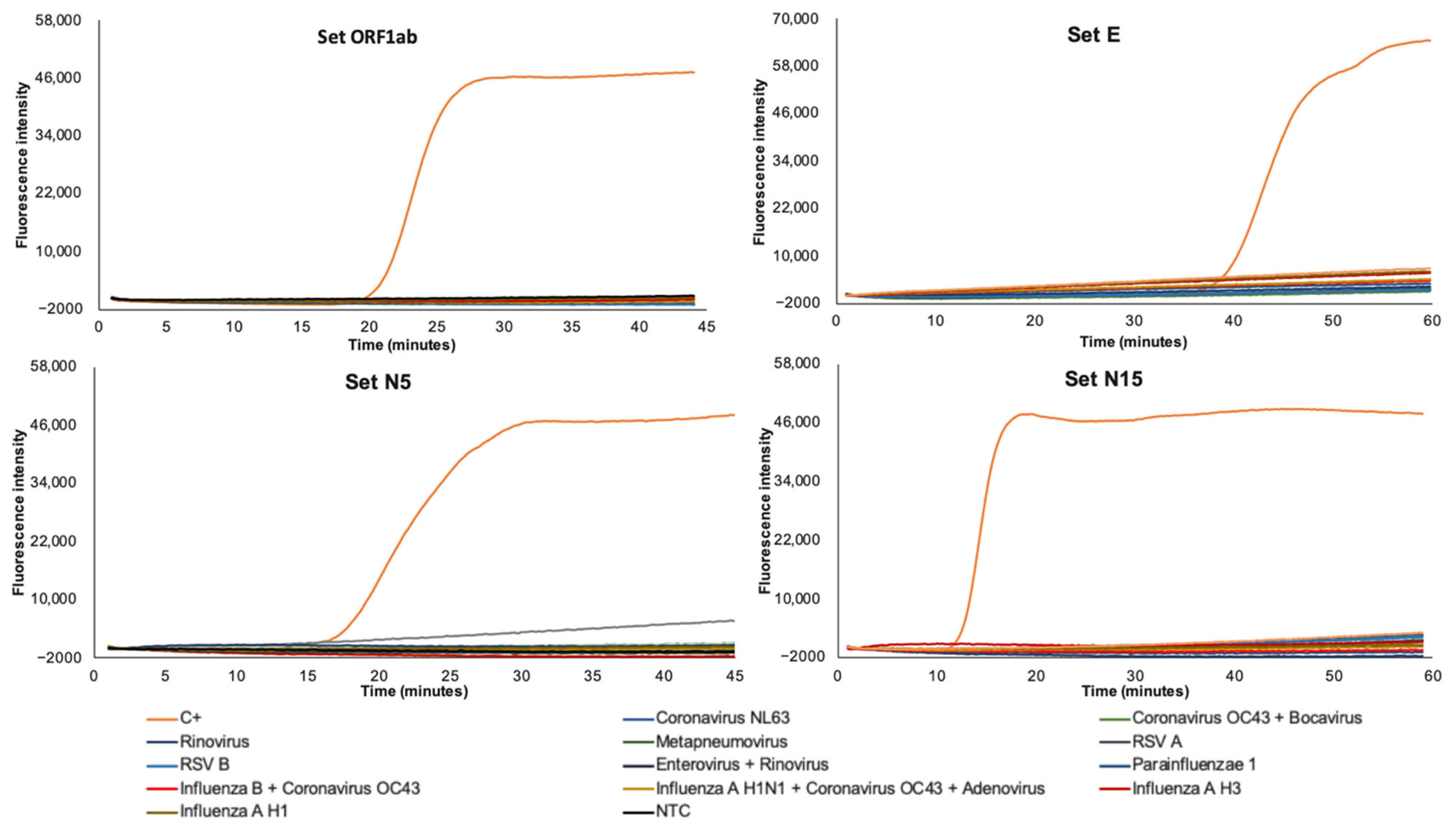
2.6. Sensitivity and Specificity
2.7. Stabilization for Long-Term Room-Temperature Storage: Dry-RT-LAMP
2.8. Estimation of the Shelf-Life of the Dry-RT-LAMP Mixes
3. Results
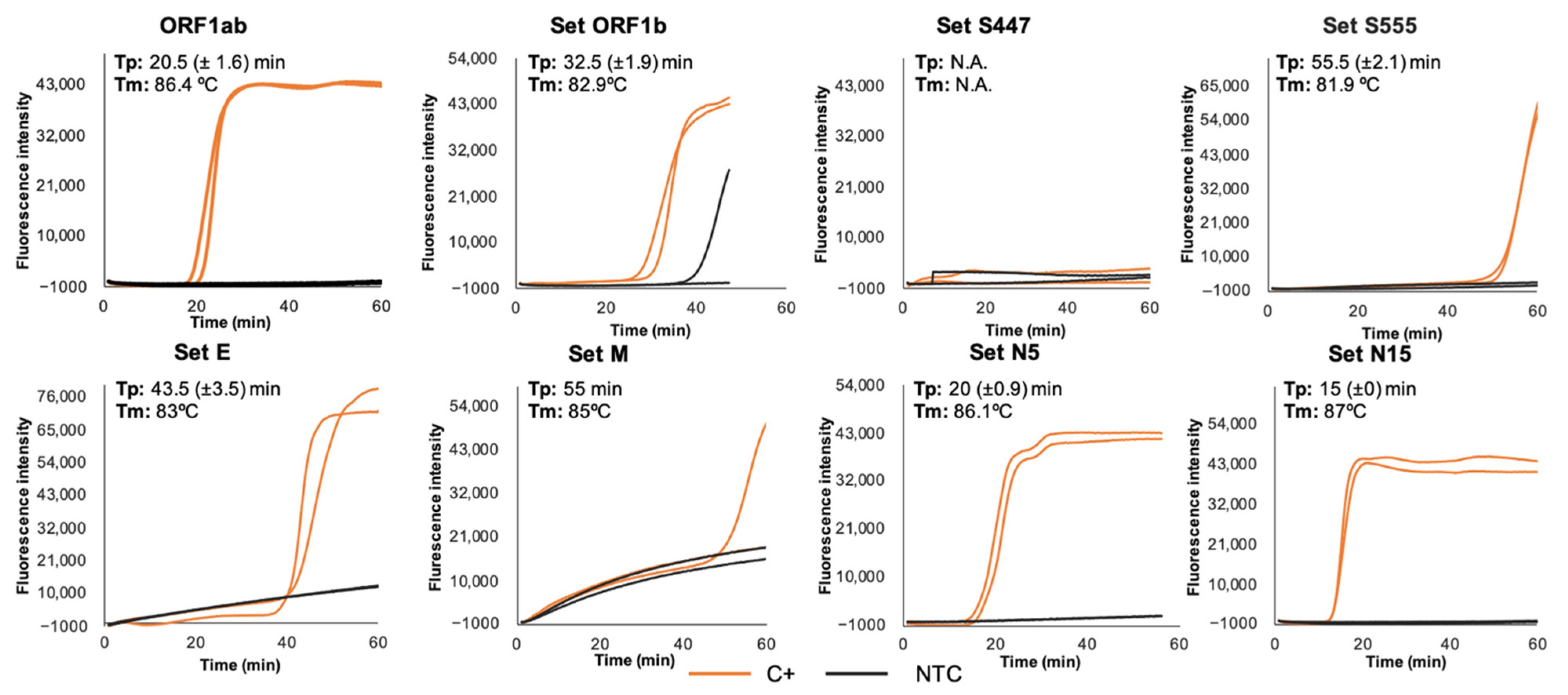
3.1. RT-LAMP Primer Sets Screening and Selection
3.2. Sensitivity and Specificity of RT-LAMP
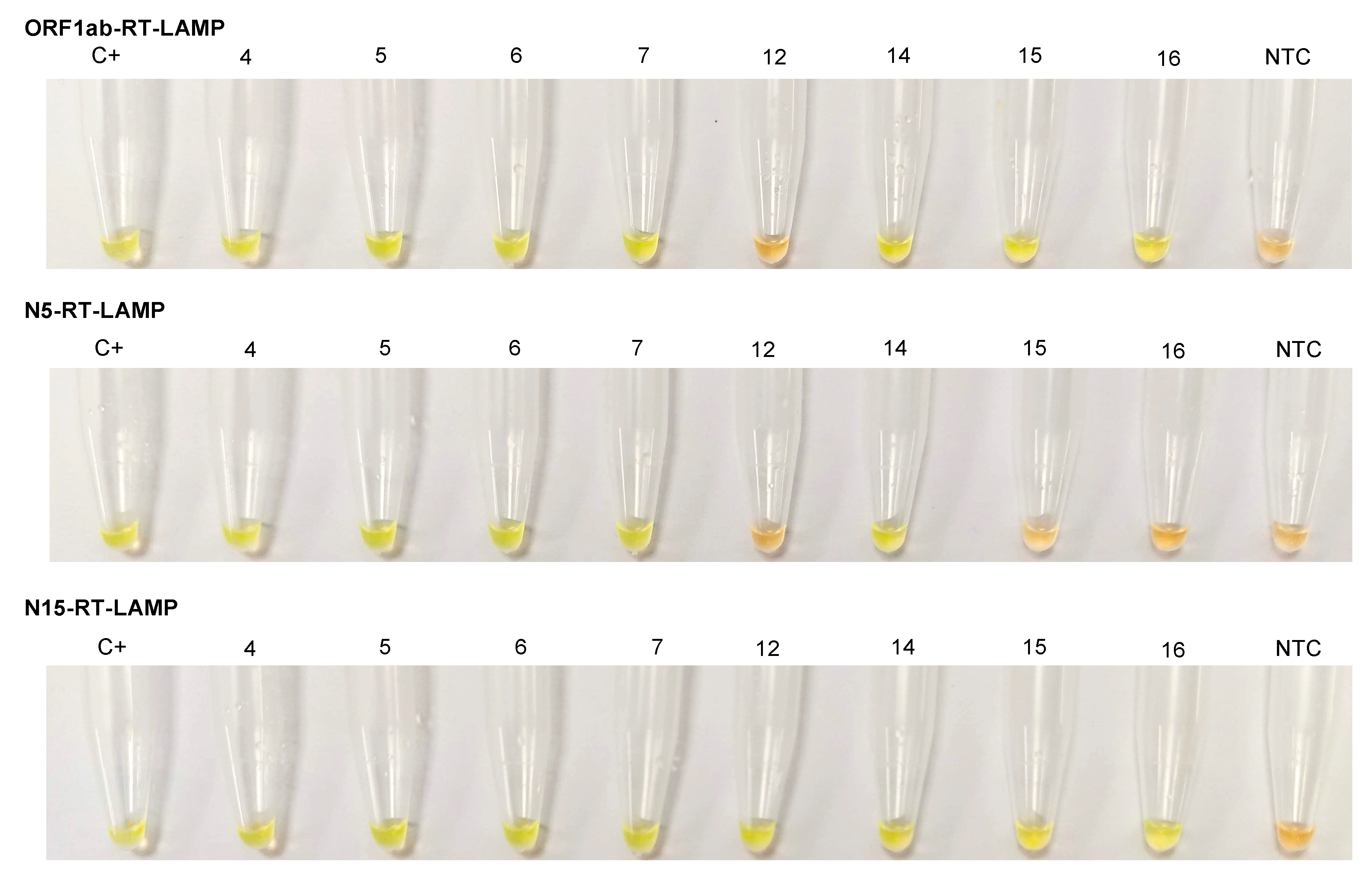
3.3. Clinical Samples Testing
3.4. Stability and Functionality Over Time of Dry-RT-LAMP Mixes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Dhama, K.; Khan, S.; Tiwari, R.; Sircar, S.; Bhat, S.; Malik, Y.S.; Singh, K.P.; Chaicumpa, W.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. Coronavirus Disease 2019–COVID-19. Clin. Microbiol. Rev. 2020, 33, e00028-20. [Google Scholar] [CrossRef] [PubMed]
- Backer, J.A.; Klinkenberg, D.; Wallinga, J. Incubation period of 2019 novel coronavirus (2019-nCoV) infections among travellers from Wuhan, China, 20–28 January 2020. Eurosurveillance 2020, 25, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oran, D.P.; Topol, E.J. Prevalence of asymptomatic SARS-CoV-2 infection: A narrative review. Ann. Intern. Med. 2020, 173, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Baj, J.; Karakuła-Juchnowicz, H.; Teresiński, G.; Buszewicz, G.; Ciesielka, M.; Sitarz, E.; Forma, A.; Karakuła, K.; Flieger, W.; Portincasa, P.; et al. COVID-19: Specific and non-specific clinical manifestations and symptoms: The current state of knowledge. J. Clin. Med. 2020, 9, 1753. [Google Scholar] [CrossRef]
- Liu, K.; Fang, Y.Y.; Deng, Y.; Liu, W.; Wang, M.F.; Ma, J.P.; Xiao, W.; Wang, Y.N.; Zhong, M.H.; Li, C.H.; et al. Clinical characteristics of novel coronavirus cases in tertiary hospitals in Hubei Province. Chin. Med. J. 2020, 133, 1025–1031. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Zhou, Y.; Ye, J.; Abdullah AL-maskri, A.A.; Kang, Y.; Zeng, S.; Cai, S. Recent advances and perspectives of nucleic acid detection for coronavirus. J. Pharm. Anal. 2020, 10, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Carter, L.J.; Garner, L.V.; Smoot, J.W.; Li, Y.; Zhou, Q.; Saveson, C.J.; Sasso, J.M.; Gregg, A.C.; Soares, D.J.; Beskid, T.R.; et al. Assay techniques and test development for COVID-19 diagnosis. ACS Cent. Sci. 2020, 6, 591–605. [Google Scholar] [CrossRef]
- Esbin, M.N.; Whitney, O.N.; Chong, S.; Maurer, A.; Darzacq, X.; Tjian, R. Overcoming the bottleneck to widespread testing: A rapid review of nucleic acid testing approaches for COVID-19 detection. RNA 2020, 26, 771–783. [Google Scholar] [CrossRef]
- Loeffelholz, M.J.; Tang, Y.W. Laboratory diagnosis of emerging human coronavirus infections–the state of the art. Emerg. Microbes Infect. 2020, 9, 747–756. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [Green Version]
- Notomi, T.; Mori, Y.; Tomita, N.; Kanda, H. Loop-mediated isothermal amplification (LAMP): Principle, features, and future prospects. J. Microbiol. 2015, 53, 1–5. [Google Scholar] [CrossRef] [PubMed]
- García-Bernalt Diego, J.; Fernández-Soto, P.; Crego-Vicente, B.; Alonso-Castrillejo, S.; Febrer-Sendra, B.; Gómez-Sánchez, A.; Vicente, B.; López-Abán, J.; Muro, A. Progress in loop-mediated isothermal amplification assay for detection of Schistosoma mansoni DNA: Towards a ready-to-use test. Sci. Rep. 2019, 9, 14744. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.P.; Othman, S.; Lau, Y.L.; Radu, S.; Chee, H.Y. Loop-mediated isothermal amplification (LAMP): A versatile technique for detection of micro-organisms. J. Appl. Microbiol. 2018, 124, 626–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustine, R.; Hasan, A.; Das, S.; Ahmed, R.; Mori, Y.; Notomi, T.; Kevadiya, B.D.; Thakor, A.S. Loop-mediated isothermal amplification (Lamp): A rapid, sensitive, specific, and cost-effective point-of-care test for coronaviruses in the context of covid-19 pandemic. Biology 2020, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Cui, J.; Huang, L.; Du, B.; Chen, L.; Xue, G.; Li, S.; Zhang, W.; Zhao, L.; Sun, Y.; et al. Rapid and visual detection of 2019 novel coronavirus (SARS-CoV-2) by a reverse transcription loop-mediated isothermal amplification assay. Clin. Microbiol. Infect. 2020, 26, 773–779. [Google Scholar] [CrossRef]
- Lamb, L.E.; Bartolone, S.N.; Ward, E.; Chancellor, M.B. Rapid detection of novel coronavirus/Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) by reverse transcription-loop-mediated isothermal amplification. PLoS ONE 2020, 15, e0234682. [Google Scholar] [CrossRef]
- Park, G.S.; Ku, K.; Baek, S.H.; Kim, S.J.; Kim, S.I.; Kim, B.T.; Maeng, J.S. Development of reverse transcription loop-mediated isothermal amplification assays targeting severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). J. Mol. Diagn. 2020, 22, 729–735. [Google Scholar] [CrossRef]
- Lu, R.; Wu, X.; Wan, Z.; Li, Y.; Zuo, L.; Qin, J.; Jin, X.; Zhang, C. Development of a novel reverse transcription loop-mediated isothermal amplification method for rapid detection of SARS-CoV-2. Virol. Sin. 2020, 12250. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.E.; Lim, B.; Hsu, C.C.; Xiong, D.; Wu, W.; Yu, Y.; Jia, H.; Wang, Y.; Zeng, Y.; Ji, M.; et al. RT-LAMP for rapid diagnosis of coronavirus SARS-CoV-2. Microb. Biotechnol. 2020, 13, 950–961. [Google Scholar] [CrossRef] [Green Version]
- Lalli, M.A.; Chen, X.; Langmade, S.J.; Fronick, C.C.; Sawyer, S.; Burcea, L.C.; Fulton, R.S.; Heinz, M.; Buchser, W.J. Rapid and extraction-free detection of SARS-CoV-2 from saliva with colorimetric LAMP. medRxiv 2020. [Google Scholar] [CrossRef]
- Yu, L.; Wu, S.; Hao, X.; Dong, X.; Mao, L.; Pelechano, V.; Chen, W.-H.; Yin, X. Rapid detection of COVID-19 Coronavirus using a reverse transcriptional loop-mediated isothermal amplification (RT-LAMP) diagnostic platform. Clin. Chem. 2020, 66, 975–977. [Google Scholar] [CrossRef]
- El-Tholoth, M.; Bau, H.H.; Song, J. A Single and Two-Stage, Closed-Tube, Molecular Test for the 2019 Novel Coronavirus (COVID-19) at home, clinic, and points of entry. ChemRxiv 2020. [Google Scholar] [CrossRef]
- Baek, Y.H.; Um, J.; Antigua, K.J.C.; Park, J.H.; Kim, Y.; Oh, S.; Kim, Y.i.; Choi, W.S.; Kim, S.G.; Jeong, J.H.; et al. Development of a reverse transcription-loop-mediated isothermal amplification as a rapid early-detection method for novel SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 998–1007. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Wu, X.; Wan, Z.; Li, Y.; Jin, X.; Zhang, C. A novel reverse transcription loop-mediated isothermal amplification method for rapid detection of sars-cov-2. Int. J. Mol. Sci. 2020, 21, 2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, Y.L.; Ismail, I.; Mustapa, N.I.; Lai, M.Y.; Tuan Soh, T.S.; Hassan, A.; Peariasamy, K.M.; Lee, Y.L.; Chong, Y.M.; Sam, I.-C.; et al. Real-time reverse transcription loop-mediated isothermal amplification for rapid detection of SARS-CoV-2. PeerJ 2020, 8, e9278. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, L. A high-throughput strategy for COVID-19 testing based on next-generation sequencing. medRxiv 2020. [Google Scholar] [CrossRef]
- Butler, D.J.; Mozsary, C.; Meydan, C.; Danko, D.; Fox, J.; Rosiene, J.; Shaiber, A.; Afshinnekoo, E.; MacKay, M.; Sedlazeck, F.J. Host, viral, and environmental transcriptome profiles of the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Servellita, V.; Singh, J.; Miao, X.; Streithorst, J.A.; Granados, A.; Sotomayor-Gonzalez, A.; et al. CRISPR–Cas12-based detection of SARS-CoV-2. Nat. Biotechnol. 2020, 38, 870–874. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Real-Time RT-PCR Diagnostic Panel For Emergency Use Only; Centre for Disease and Control Prevention: Atlanta, GA, USA, 2020; Volume 4. [Google Scholar]
- Rödel, J.; Egerer, R.; Suleyman, A.; Sommer-Schmid, B.; Baier, M.; Henke, A.; Edel, B.; Löffler, B. Use of the variplexTM SARS-CoV-2 RT-LAMP as a rapid molecular assay to complement RT-PCR for COVID-19 diagnosis. J. Clin. Virol. 2020, 132, 104616. [Google Scholar] [CrossRef]
- Thompson, D.; Lei, Y. Mini review: Recent progress in RT-LAMP enabled COVID-19 detection. Sens. Actuators Rep. 2020, 2, 100017. [Google Scholar] [CrossRef]
- Rabe, B.A.; Cepko, C. SARS-CoV-2 detection using isothermal amplification and a rapid, inexpensive protocol for sample inactivation and purification. Proc. Natl. Acad. Sci. USA 2020, 117, 24450–24458. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, G.S. Shelf life of medical devices. In Fact Sheet: FDA Good Guidance Practices; Center for Devices and Radiological Health: Silver Spring, MD, USA, 1991; Volume 27. [Google Scholar]
- Pendergast, J.S.; Yamazaki, S. Mammalian peripheral circadian oscillators are temperature compensated. J. Biol. Rhythms 2008, 23, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Rolando, J.C.; Jue, E.; Schoepp, N.G.; Ismagilov, R.F. Real-time, digital LAMP with commercial microfluidic chips reveals the interplay of efficiency, speed, and background amplification as a function of reaction temperature and time. Anal. Chem. 2019, 91, 1034–1042. [Google Scholar] [CrossRef] [Green Version]
- Rolando, J.C.; Jue, E.; Barlow, J.T. Real-time kinetics and high-resolution melt curves in single-molecule digital LAMP to differentiate and study specific and non-specific amplification. Nucleic Acids Res. 2020, e42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D. One-pot detection of COVID-19 with real-time reverse-transcription loop-mediated isothermal amplification (RT-LAMP) assay and Visual RT-LAMP assay. bioRxiv 2020. [Google Scholar] [CrossRef]
- Schmid-Burgk, J.L.; Li, D.; Feldman, D.; Slabicki, M.; Borrajo, J.; Strecker, J.; Cleary, B.; Regev, A.; Zhang, F. LAMP-Seq: Population-scale COVID-19 diagnostics using a compressed barcode space. bioRxiv 2020. [Google Scholar] [CrossRef]
- Khailany, R.A.; Safdar, M.; Ozaslan, M. Genomic characterization of a novel SARS-CoV-2 Rozhgar. Gene Rep. 2020, 19, 1–6. [Google Scholar] [CrossRef]
- Callaway, B.E. Making sense of coronavirus mutations. Nature 2020, 585, 174–177. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Pan, W.; Arasthfer, A.; Fang, W.; Ling, L.; Fang, H.; Daneshnia, F.; Yu, J.; Liao, W.; Pei, H.; et al. Development and validation of a rapid, single-step reverse transcriptase loop-mediated isothermal amplification (RT-LAMP) System potentially to be used for reliable and high-throughput screening of COVID-19. Front. Cell. Infect. Microbiol. 2020, 10, 331. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, D.; Yang, P.; Poon, L.L.M.; Wang, Q. Viral load of SARS-CoV-2 in clinical samples. Lancet Infect. Dis. 2020, 20, 411–412. [Google Scholar] [CrossRef]
- Corman, V.M.; Jones, T.C.; Mühlemann, B.; Veith, T.; Biele, G.; Zuchowski, M.; Hofmann, J.; Stein, A.; Edelmann, A.; Drosten, C. An analysis of SARS-CoV-2 viral load by patient age. medRxiv 2020. [Google Scholar] [CrossRef]
- Pujadas, E.; Ibeh, N.; Hernandez, M.M.; Waluszko, A.; Sidorenko, T.; Flores, V.; Shiffrin, B.; Chiu, N.; Young-Francois, A.; Nowak, M.D.; et al. Comparison of SARS-CoV-2 detection from nasopharyngeal swab samples by the Roche cobas 6800 SARS-CoV-2 test and a laboratory-developed real-time RT-PCR test. J. Med. Virol. 2020, 92, 1695–1698. [Google Scholar] [CrossRef]
- Yu, F.; Yan, L.; Wang, N.; Yang, S.; Wang, L.; Tang, Y.; Gao, G.; Wang, S.; Ma, C.; Xie, R.; et al. Quantitative detection and viral load analysis of SARS-CoV-2 in infected patients. Clin. Infect. Dis. 2020, 71, 793–798. [Google Scholar] [CrossRef] [Green Version]
- Khorosheva, E.M.; Karymov, M.A.; Selck, D.A.; Ismagilov, R.F. Lack of correlation between reaction speed and analytical sensitivity in isothermal amplification reveals the value of digital methods for optimization: Validation using digital real-time RT-LAMP. Nucleic Acids Res. 2016, 44, e10. [Google Scholar] [CrossRef]
- Hayashida, K.; Kajino, K.; Hachaambwa, L.; Namangala, B.; Sugimoto, C. Direct blood dry LAMP: A rapid, stable, and easy diagnostic tool for human African trypanosomiasis. PLoS Negl. Trop. Dis. 2015, 9, e0003578. [Google Scholar] [CrossRef]
- Chander, Y.; Koelbl, J.; Puckett, J.; Moser, M.J.; Klingele, A.J.; Liles, M.R.; Carrias, A.; Mead, D.A.; Schoenfeld, T.W. A novel thermostable polymerase for RNA and DNA loop-mediated isothermal amplification (LAMP). Front. Microbiol. 2014, 5, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.; Müller, T.G.; Khalid, D.; Sonntag-Buck, V.; Heuser, A.M.; Glass, B.; Meurer, M.; Morales, I.; Schillak, A.; Freistaedter, A.; et al. SARS-CoV-2 RNA extraction using magnetic beads for rapid large-scale testing by RT-qPCR and RT-LAMP. Viruses 2020, 12, 863. [Google Scholar] [CrossRef]
- Margolin, E.; Burgers, W.A.; Sturrock, E.D.; Mendelson, M.; Chapman, R.; Douglass, N.; Williamson, A.L.; Rybicki, E.P. Prospects for SARS-CoV-2 diagnostics, therapeutics and vaccines in Africa. Nat. Rev. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
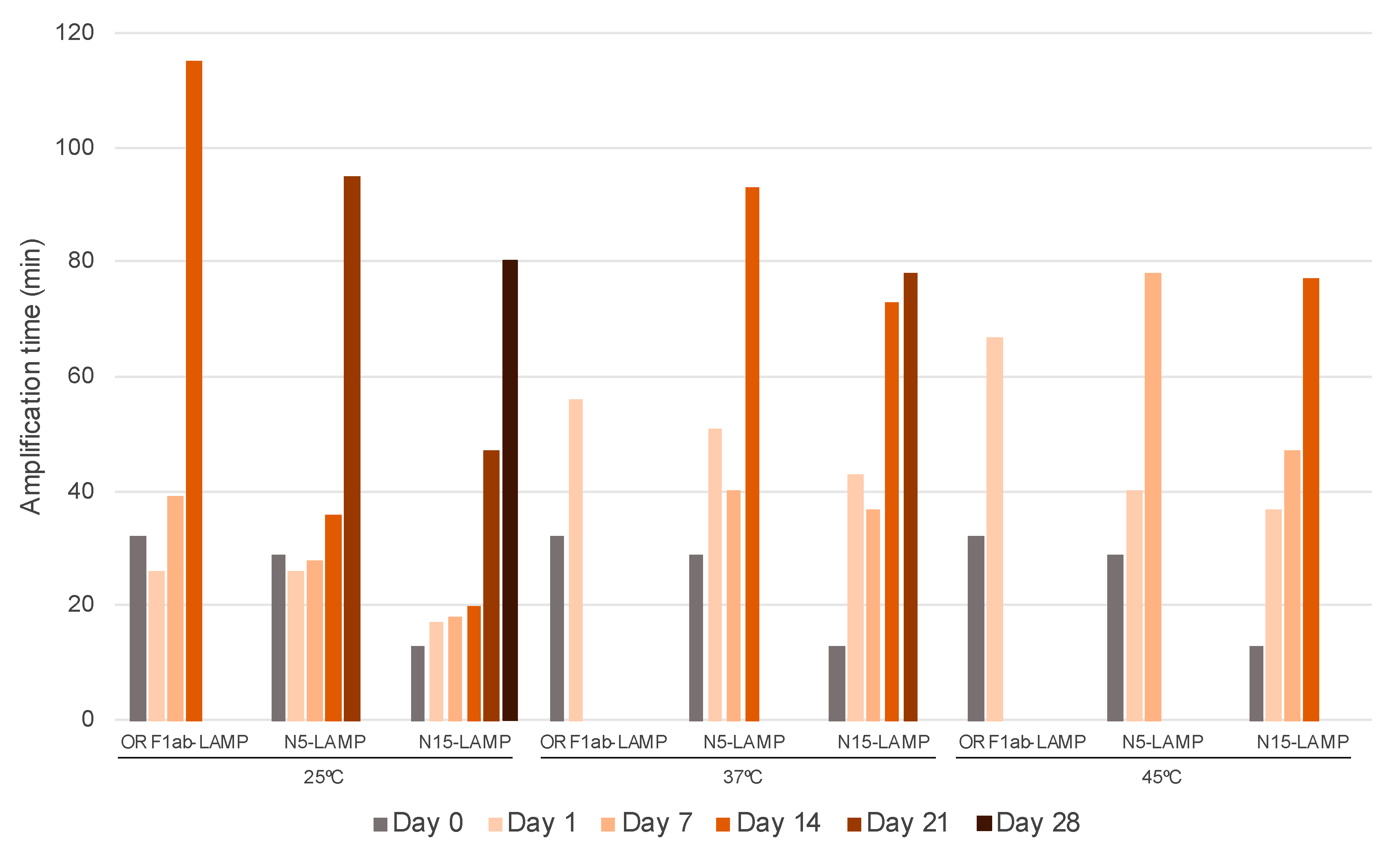{kind=link}
{kind=link}
| Set | Primer 1 | Sequence 5′→3′ | Length (nt) | Gene Position |
|---|---|---|---|---|
| ORF1ab 2 | F3 | TGCTTCAGTCAGCTGATG | 18 | 13,434–13,636 |
| B3 | TTAAATTGTCATCTTCGTCCTT | 22 | ||
| FIP | CAGTACTAGTGCCTGTGCCGCACAATCGTTTTTAAACGGGT | 41 | ||
| BIP | TCGTATACAGGGCTTTTGACATCTATCTTGGAAGCGACAACAA | 43 | ||
| LF | CTGCACTTACACCGCAA | 17 | ||
| LB | GTAGCTGGTTTTGCTAAATTCC | 22 | ||
| ORF1b 3 | F3 | CACAGACTTTGTGAATGAGTT | 21 | 15,654–15,896 |
| B3 | GTCAGTCTCAGTCCAACAT | 19 | ||
| FIP | CTATTGAAACACACAACAGCATCGCATATTTGCGTAAACATTTCTCA | 47 | ||
| BIP | TATGCATCTCAAGGTCTAGTGGCTATGCTTCAGACATAAAAACATTG | 47 | ||
| S447 3 | F3 | GTTTCTGCCTTTCCAACAA | 19 | 22,985–23,415 |
| B3 | AACAGGGACTTCTGTGCA | 18 | ||
| FIP | TCAAGAATCTCAAGTGTCTGTGGTGGCAGAGACATTGCTGA | 41 | ||
| BIP | ACCATGTTCTTTTGGTGGTGTCAACATCCTGATAAAGAACAGC | 43 | ||
| LF | TCACGGACAGCATCAGTAGTG | 21 | ||
| LB | CAGGAACAAATACTTCTAACCAGGT | 25 | ||
| S555 3 | F3 | CTATGCAAATGGCTTATAGGTT | 22 | 24,182–24,736 |
| B3 | AGTTGTTTAACAAGCGTGTT | 20 | ||
| FIP | GCACTATTAAATTGGTTGGCAATCATAATGGTATTGGAGTTACACAGA | 48 | ||
| BIP | ATTGGCAAAATTCAAGACTCACTTTTGTGCATTTTGGTTGACC | 43 | ||
| E 4 | F3 | TCATTCGTTTCGGAAGAGA | 19 | 26,245–26,472 |
| B3 | AGGAACTCTAGAAGAATTCAGAT | 23 | ||
| FIP | TGTAACTAGCAAGAATACCACGAAACAGGTACGTTAATAGTTAATAGCG | 49 | ||
| BIP | GCTTCGATTGTGTGCGTACTCGAGAGTAAACGTAAAAAGAAGG | 43 | ||
| M 4 | F3 | GTTTCCTATTCCTTACATGGATT | 23 | 26,597–26,801 |
| B3 | AGCCACATCAAGCCTACA | 18 | ||
| FIP | CCATAACAGCCAGAGGAAAATTAACCTTCTACAATTTGCCTATGCC | 46 | ||
| BIP | AACTTTAGCTTGTTTTGTGCTTGCACAAGCCATTGCGATAGC | 42 | ||
| N5 4 | F3 | CCAGAATGGAGAACGCAGTG | 20 | 28,355–28,570 |
| B3 | CCGTCACCACCACGAATT | 18 | ||
| FIP | AGCGGTGAACCAAGACGCAGGGCGCGATCAAAACAACG | 38 | ||
| BIP | AATTCCCTCGAGGACAAGGCGAGCTCTTCGGTAGTAGCCAA | 41 | ||
| LF | ATTATTGGGTAAACCTTGGGGC | 22 | ||
| LB | ATTAACACCAATAGCAGTCCAGATG | 25 | ||
| N15 4 | F3 | AGATCACATTGGCACCCG | 18 | 28,703–28,915 |
| B3 | CCATTGCCAGCCATTCTAGC | 20 | ||
| FIP | TGCTCCCTTCTGCGTAGAAGCCAATGCTGCAATCGTGCTAC | 41 | ||
| BIP | GGCGGCAGTCAAGCCTCTTCCCTACTGCTGCCTGGAGTT | 39 | ||
| LF | GCAATGTTGTTCCTTGAGGAAGTT | 24 | ||
| LB | CCTCATCACGTAGTCGCAACAG | 22 |
| RT-qPCR Ct Values | Real-Time RT-LAMP Tp Values | |||||||
|---|---|---|---|---|---|---|---|---|
| Groups | No. Sample | Commercial Kit 1 | ORF1ab | E/N | ORF1ab | N5 | N15 | E |
| Group 1 ORF1ab+ (Ct < 30) E/N+ | 1 | VIASURE | 22 | 27 | 15 | 14 | 9 | 27 |
| 2 | COBAS | 25 | 26 | 18 | 16 | 13 | 72 | |
| 3 | VIASURE | 25 | 29 | 15 | 15 | 19 | 43 | |
| 4 | VIASURE | 26 | 25 | 17 | 16 | 11 | 28 | |
| 5 | VIASURE | 27 | 31 | 17 | 18 | 13 | - | |
| 6 | VIASURE | 27 | 30 | 19 | 18 | 13 | 35 | |
| Group 2 ORF1ab+ (Ct > 30) E/N+ | 7 | VIASURE | 32 | 31 | 20 | 22 | 13 | 60 |
| 8 | COBAS | 32 | 35 | 35 | 23 | 49 | - | |
| 9 | VIASURE | 33 | 39 | 23 | 20 | 39 | - | |
| 10 | COBAS | 33 | 36 | 31 | - | 60 | - | |
| 11 | COBAS | 34 | 36 | - | - | 45 | - | |
| 12 | VIASURE | 39 | 36 | - | - | 36 | - | |
| Group 3 ORF1ab+ E− | 13 | COBAS | 36 | - | 39 | - | 29 | - |
| 14 | COBAS | 33 | - | 30 | 18 | 47 | - | |
| Group 4 ORF1ab- N+ | 15 | VIASURE | - | 25 | 34 | - | 46 | - |
| 16 | VIASURE | - | 38 | 28 | - | 31 | - | |
| 17 | VIASURE | - | 38 | - | - | - | - | |
| 18 | VIASURE | - | 40 | 43 | - | 60 | - | |
| 19 | VIASURE | - | 41 | - | - | 58 | - | |
| 20 | VIASURE | - | 41 | - | - | 30 | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Bernalt Diego, J.; Fernández-Soto, P.; Domínguez-Gil, M.; Belhassen-García, M.; Bellido, J.L.M.; Muro, A. A Simple, Affordable, Rapid, Stabilized, Colorimetric, Versatile RT-LAMP Assay to Detect SARS-CoV-2. Diagnostics 2021, 11, 438. https://doi.org/10.3390/diagnostics11030438
García-Bernalt Diego J, Fernández-Soto P, Domínguez-Gil M, Belhassen-García M, Bellido JLM, Muro A. A Simple, Affordable, Rapid, Stabilized, Colorimetric, Versatile RT-LAMP Assay to Detect SARS-CoV-2. Diagnostics. 2021; 11(3):438. https://doi.org/10.3390/diagnostics11030438
Chicago/Turabian StyleGarcía-Bernalt Diego, Juan, Pedro Fernández-Soto, Marta Domínguez-Gil, Moncef Belhassen-García, Juan Luis Muñoz Bellido, and Antonio Muro. 2021. "A Simple, Affordable, Rapid, Stabilized, Colorimetric, Versatile RT-LAMP Assay to Detect SARS-CoV-2" Diagnostics 11, no. 3: 438. https://doi.org/10.3390/diagnostics11030438