Effects of Two Trichoderma Strains on Plant Growth, Rhizosphere Soil Nutrients, and Fungal Community of Pinus sylvestris var. mongolica Annual Seedlings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Organisms and Growth Conditions
2.2. Experimental Design and Seedling Inoculation
2.3. Sampling and Analysis of Seedlings
2.4. Soil Properties Analysis
2.5. Fungal Diversity Analysis
2.6. Data Analyses
3. Results and Analysis
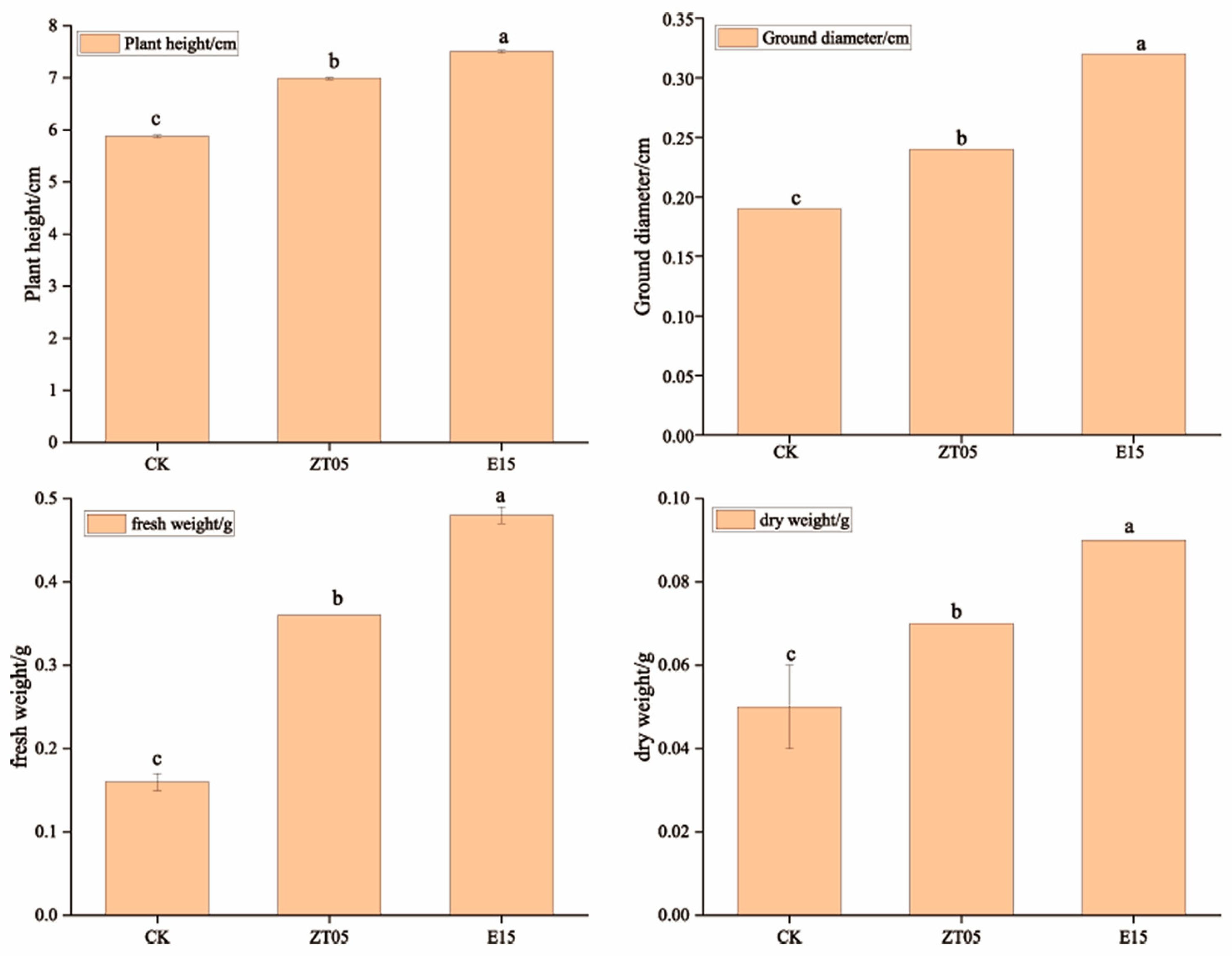
3.1. Effects of Trichoderma Inoculation on Seedling Growth
3.1.1. Seedling Height
3.1.2. Seedling Diameter
3.1.3. Seedling Biomass
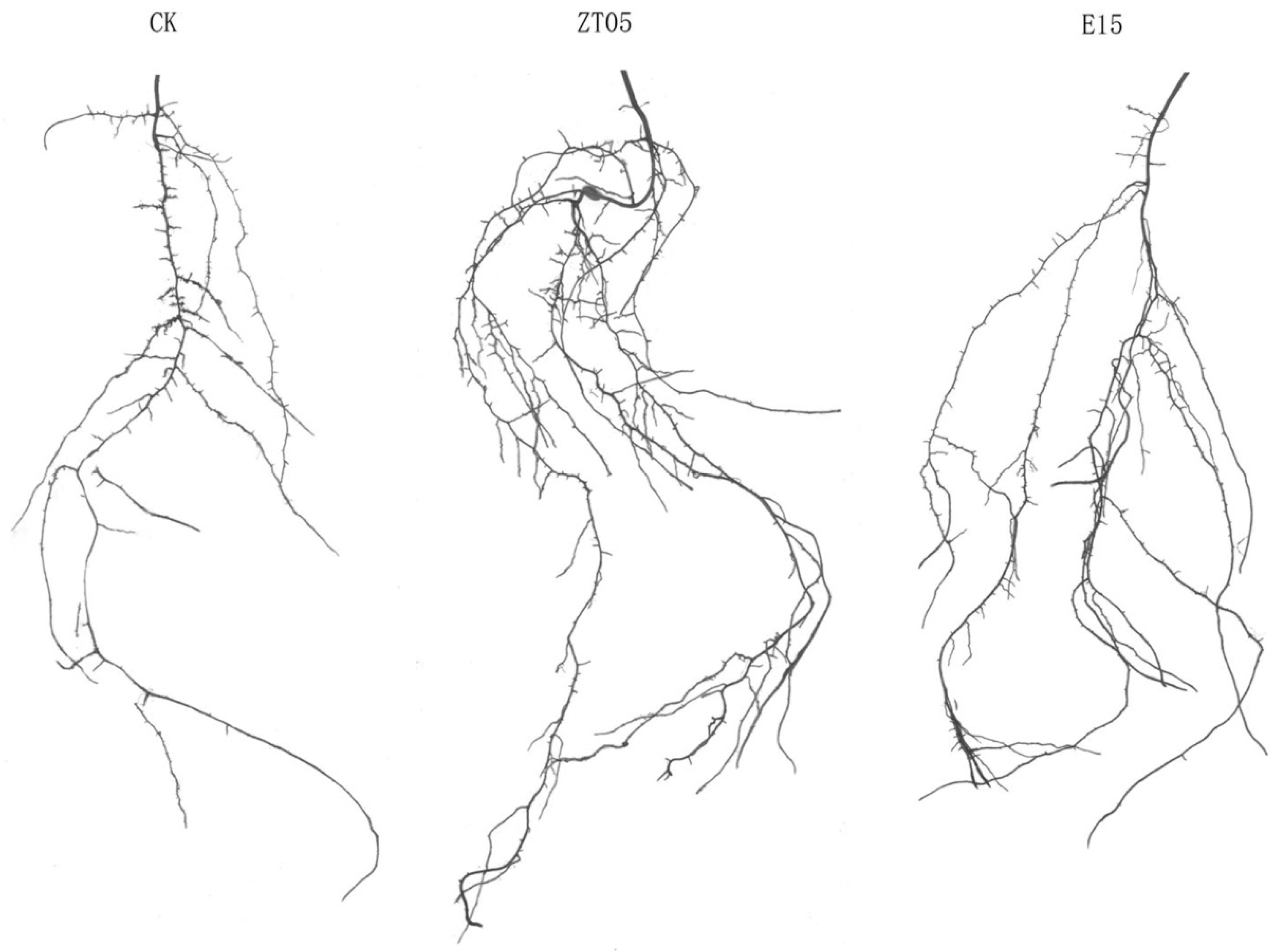
3.2. Effect of Trichoderma Inoculation on the Root Structure of Seedlings
3.3. Effects of Trichoderma Inoculation on Physicochemical Properties of Seedling Rhizosphere Soil
3.4. Effects of Trichoderma Inoculation on Rhizosphere Soil Enzyme Activity
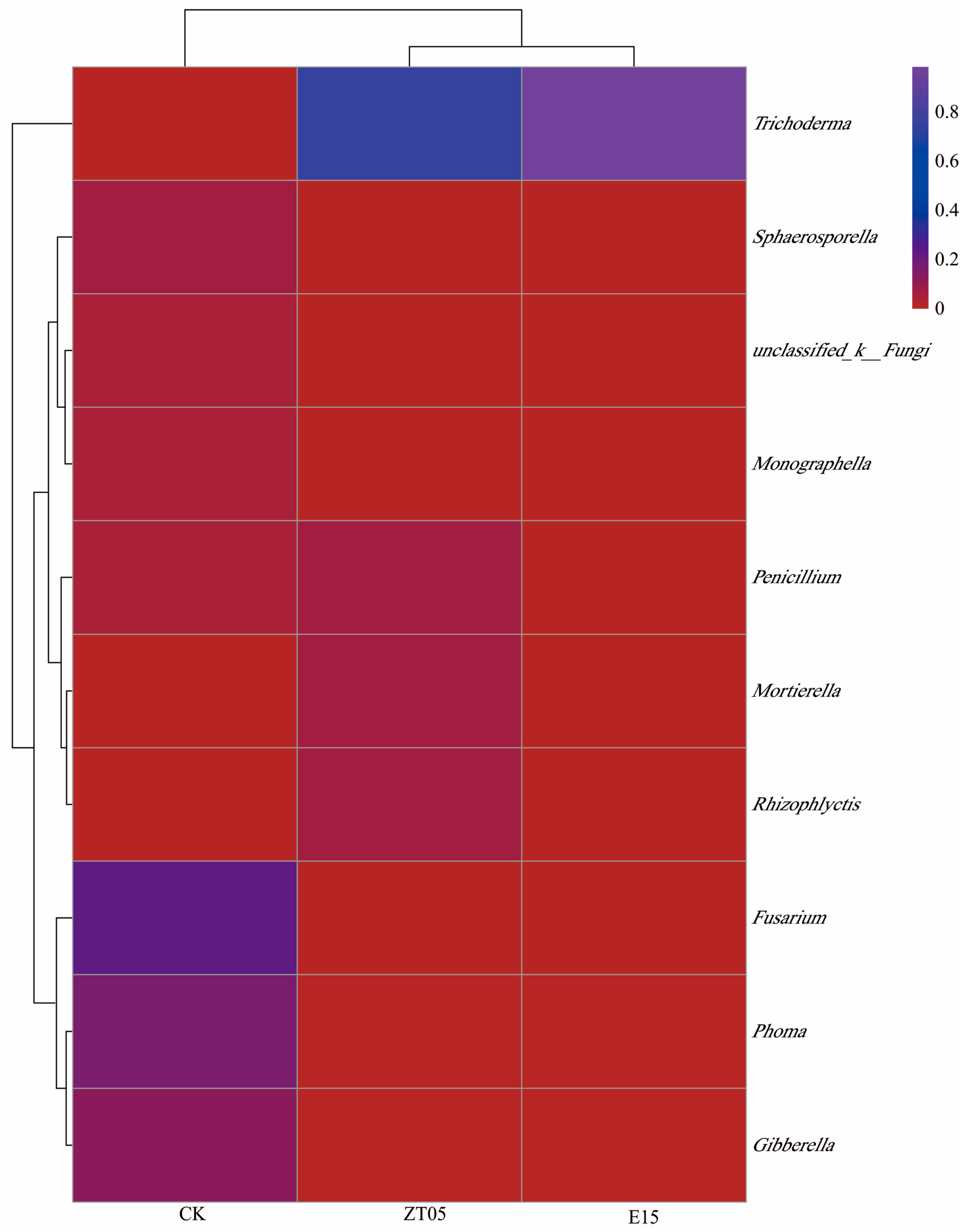
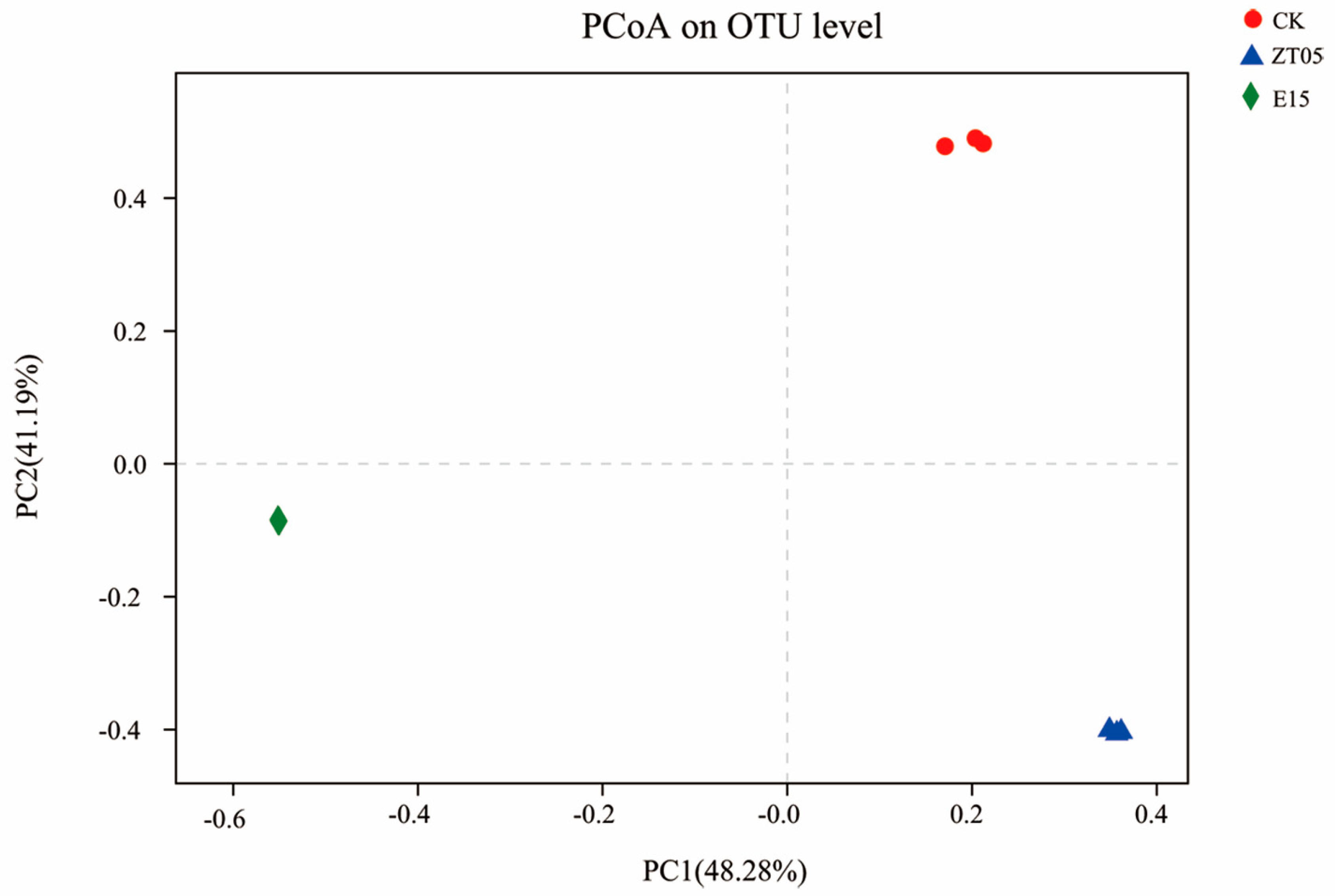
3.5. Effect on the Diversity of Rhizosphere Fungi of Seedlings
3.5.1. Soil Sample Sequencing Results and Sampling Depth Verification
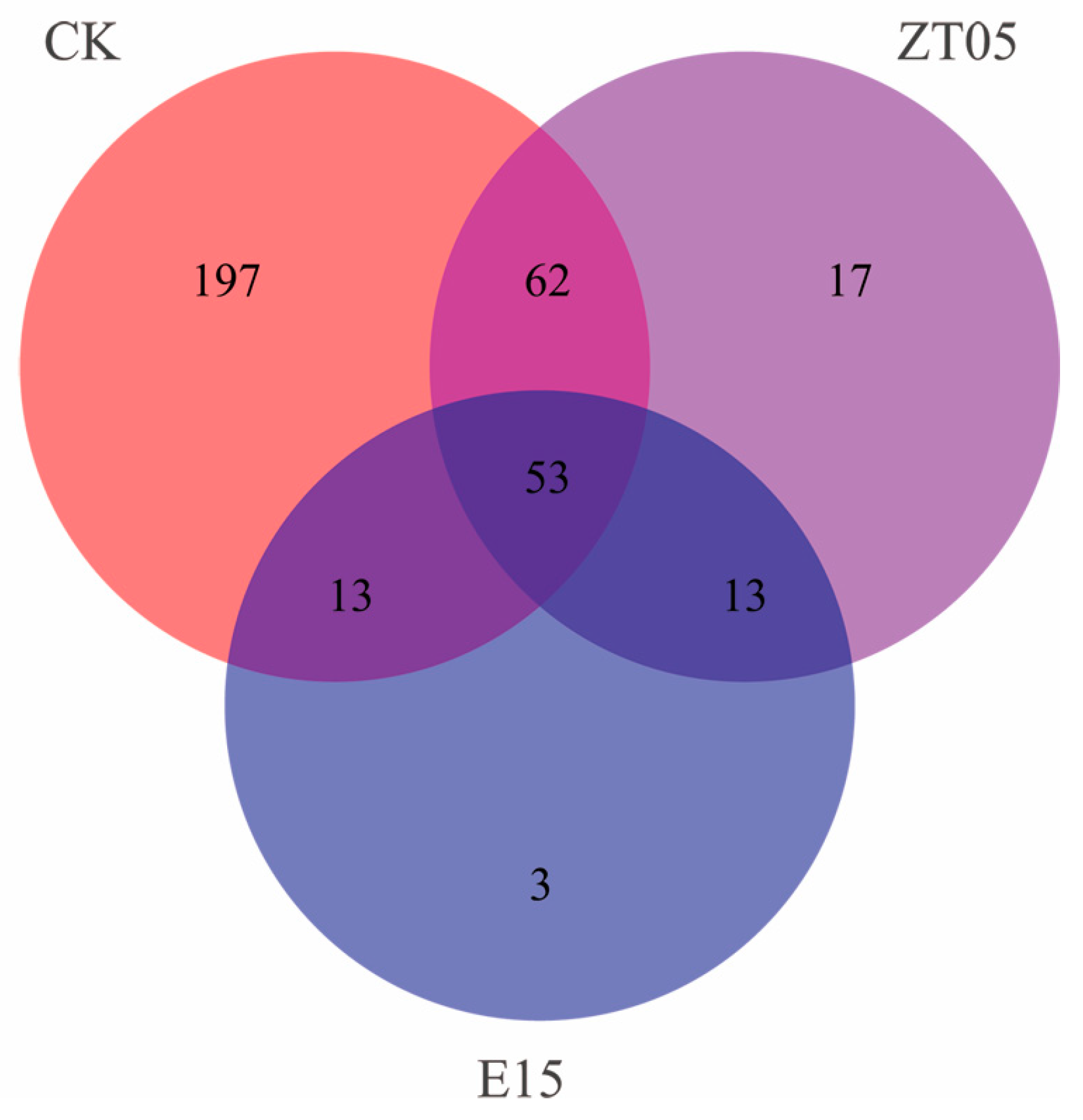
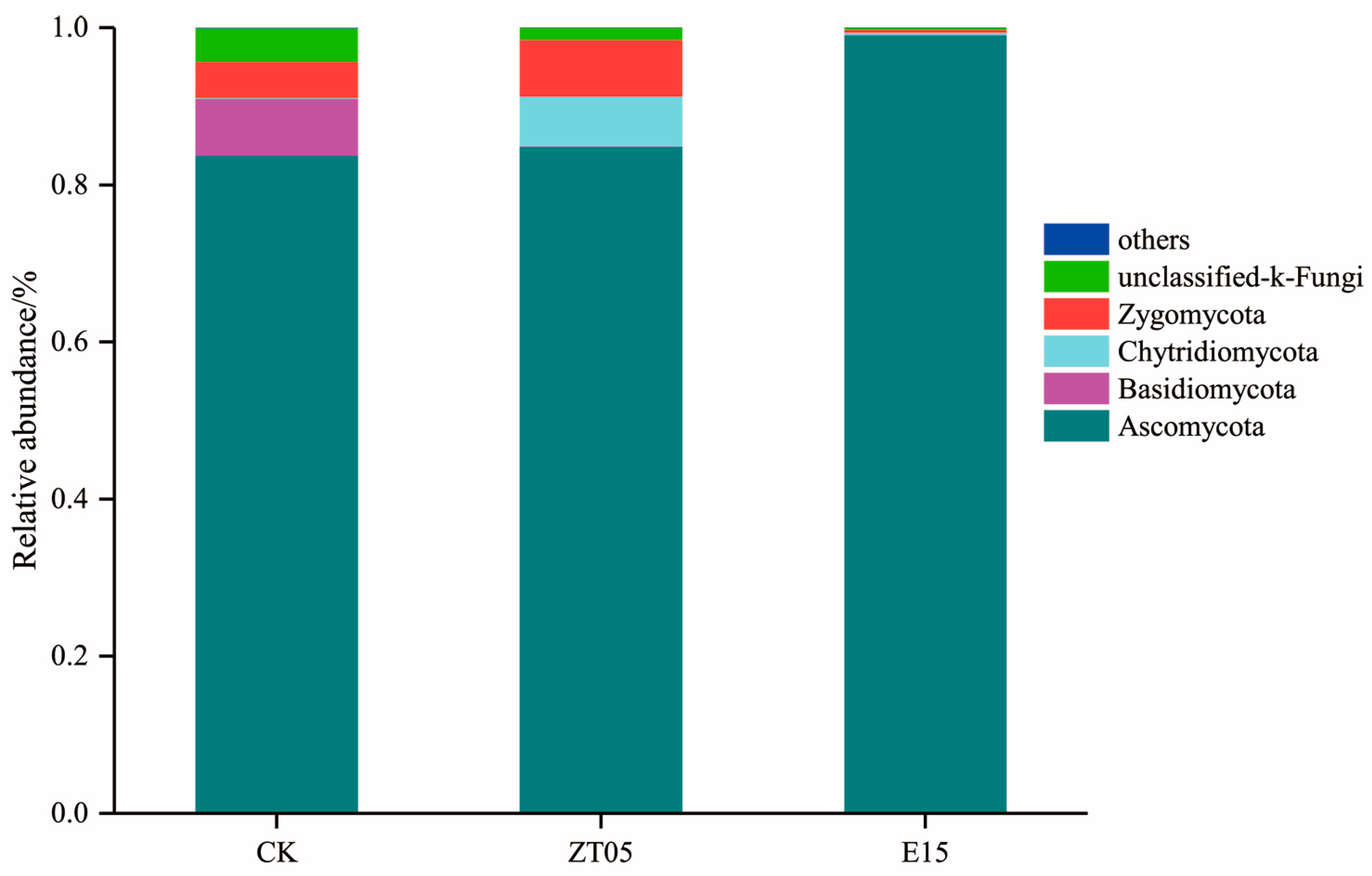
3.5.2. Distribution of Soil Fungal Community
3.5.3. Analysis of α Diversity Index
3.5.4. Analysis of β Diversity Index
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lorito, M.; Harman, G.E.; Mastouri, F. Translational research on Trichoderma: From omics to the field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Brotman, Y.; Kapuganti, J.G.; Viterbo, A. Trichoderma. Curr. Biol. 2010, 20, R390–R391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yedidia, I.I.; Benhamou, N.; Chet, I.I. Induction of Defense Responses in Cucumber Plants (Cucumis sativus L.) by the Biocontrol Agent Trichoderma harzianum. Appl. Environ. Microbiol. 1999, 65, 1061–1070. [Google Scholar] [PubMed]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced Systemic Resistance and Plant Responses to Fungal Biocontrol Agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras-Cornejo, H.A.; Macias-Rodriguez, L.; Cortes-Penagos, C.; Lopez-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yuan, J.; Yang, X.; Cui, Y.; Chen, L.; Ran, W.; Shen, Q. Putative Trichoderma harzianum mutant promotes cucumber growth by enhanced production of indole acetic acid and plant colonization. Plant Soil 2013, 368, 433–444. [Google Scholar] [CrossRef]
- Anil, K.; Lakshmi, T. Phosphate solubilization potential and phosphatase activity of rhizospheric Trichoderma spp. Braz. J. Microbiol. 2010, 41, 787–795. [Google Scholar]
- Saravanakumar, K.; Shanmuga, A.V.; Kathiresan, K. Effect of Trichoderma on soil phosphate solubilization and growth improvement of Avicennia marina. Aquat. Bot. 2013, 104, 101–105. [Google Scholar] [CrossRef]
- Elena, B.; Paolo, A.; Fabio, P.; Moreno, T. Use of Trichoderma Spp. and Arbuscular Mycorrhizal Fungi to Increase Soil Beneficial Population of Bacteria in a Nectarine Commercial Orchard: Effect on Root Growth, Nutrient Acquisition and Replanting Disease. J. Plant Nutr. 2015, 39, 1147–1155. [Google Scholar]
- Zhang, F.; Zhu, Z.; Yang, X.; Ran, W.; Shen, Q. Trichoderma harzianum T-E5 significantly affects cucumber root exudates and fungal community in the cucumber rhizosphere. Appl. Soil Ecol. 2013, 72, 41–48. [Google Scholar] [CrossRef]
- Savazzini, F.; Longa, C.M.O.; Pertot, I. Impact of the biocontrol agent Trichoderma atroviride SC1 on soil microbial communities of a vineyard in northern Italy. Soil Biol. Biochem. 2009, 41, 1457–1465. [Google Scholar] [CrossRef]
- Zhou, X.; Jia, H.; Ge, X.; Wu, F. Effects of vanillin on the community structures and abundances of Fusarium and Trichoderma spp. in cucumber seedling rhizosphere. J. Plant Interact. 2018, 13, 45–50. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chang, Y.C.; Baker, R.; Kleifeld, O.; Chet, I. Increased growth of plants in the presence of the biological control agent Trichoderma harzianum. Plant Dis. 1986, 70, 145–148. [Google Scholar] [CrossRef]
- Eugenia, M.D.; Belén; Sara, D.; Hermosa, R.; Monte, E.; Carlos, N. Transcriptomic response of Arabidopsis thaliana after 24h incubation with the biocontrol fungus Trichoderma harzianum. J. Plant Physiol. 2012, 169, 614–620. [Google Scholar]
- Contreras-Cornejo, H.; Macías-Rodríguez, L.I.; Alfaro, C.R.; López-Bucio, J. Trichoderma spp. Improve growth of Arabidopsis seedlings under salt stress through enhanced root development, osmolite production, and Na⁺ elimination through root exudates. Mol. Plant Microbe Interact. 2014, 27, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Spor, A.; Edel-Hermann, V.; Heraud, C.; Breuil, M.C.; Bizouard, F.; Toyoda, S.; Yoshida, N.; Steinberg, C.; Philippot, L. N2O production, a widespread trait in fungi. Sci. Rep. 2015, 5, 9697. [Google Scholar] [CrossRef]
- Khan, M.Y.; Haque, M.M.; Molla, A.H.; Rahman, M.; Alam, M.Z. Antioxidant compounds and minerals in tomatoes by, Trichoderma-enriched biofertilizer and their relationship with the soil environments. J. Integr. Agric. 2017, 16, 691–703. [Google Scholar] [CrossRef]
- Harman, G.E. Multifunctional fungal plant symbionts: New tools to enhance plant growth and productivity. New Phytol. 2011, 189, 647–649. [Google Scholar] [CrossRef]
- Mbarki, S.; Cerdà, A.; Brestic, M.; Mahendra, R.; Abdelly, C.; Pascual, J.A. Vineyard compost supplemented with\r, trichodermaharzianum\r, t78 improve saline soil quality. Land Degrad. Dev. 2016, 28, 1028–1037. [Google Scholar] [CrossRef]
- Zhang, F.G. The Affects and Mechanisms of Puta Five Trichoredma harzianum Mutant and ITS Bio-Organic Fertilizer on Growth of Cucumber; Nanjing Agricultural University: Nanjing, China, 2015; pp. 15–18. [Google Scholar]
- Assigbetsé, K.; Ciss, I.; Bakhoum, N.; Dieng, L. Effect of inoculation of acacia senegal mature trees with mycorrhiza and rhizobia on soil properties and microbial community structure. In Proceedings of the EGU General Assembly Conference Abstracts, Vienna, Austria, 22–27 April 2012; pp. 2012–8004. [Google Scholar]
- Wagner, K.; Apostolakis, A.; Daliakopoulos, I.; Tsanis, I. Can Tomato Inoculation with Trichoderma Compensate Yield and Soil Health Deficiency due to Soil Salinity? In Proceedings of the Egu General Assembly Conference Abstracts, Vienna, Austria, 17–22 April 2016. [Google Scholar] [CrossRef]
- Yadav, R.L.; Shukla, S.K.; Suman, A.; Singh, P.N. Trichoderma inoculation and trash management effects on soil microbial biomass, soil respiration, nutrient uptake and yield of ratoon sugarcane under subtropical conditions. Biol. Fertil. Soils 2009, 45, 461–468. [Google Scholar] [CrossRef]
- Zhu, J.-J.; Li, F.-Q.; Xu, M.-L.; Kang, H.-Z.; Wu, X.-Y. Le rôle des champignons ectomycorrhiziens dans la réduction du dépérissement du pin sur sols sableux semi-arides du nord de la Chine: Une approche expérimentale. Ann. For. 2008, 65, 304. [Google Scholar]
- Yin, D.C.; Deng, X.; Ilan, C.; Song, R.Q. Physiological Responses of Pinus sylvestris var. Mongolica Seedlings to the Interaction Between Suillus luteus and Trichoderma virens. Curr. Microbiol. 2014, 69, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.C.; Song, R.Q.; Qi, J.Y.; Deng, X. Ectomycorrhizal fungus enhances drought tolerance of Pinus sylvestris var. mongolica seedlings and improves soil condition. J. For. Res. 2018, 29, 1775–1788. [Google Scholar] [CrossRef]
- John, C.J.; Jishma, P.; Karthika, N.R.; Nidheesh, K.S.; Radhakrishnan, E.K. Pseudomonas fluorescens R68 assisted enhancement in growth and fertilizer utilization of Amaranthus tricolor (L.). 3 Biotech 2017, 7, 256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.G.; Huo, Y.Q.; Cobb, A.B.; Luo, G.W.; Zhou, J.Q.; Yang, G.W.; Gail, W.T.W.; Zhang, Y.J. Trichoderma biofertilizer links to altered soil chemistry, altered microbial communities, and improved grassland biomass. Front. Microbiol. 2018, 9, 848. [Google Scholar] [CrossRef] [PubMed]
- Nunzio, F.; Valeria, V.; Woo, S.L.; Olimpia, P.; Armando, D.R.; Laura, G.; Romano, I.; Lombardi, N.; Napolitano, M.; Colla, G.; et al. Trichoderma-Based Biostimulants Modulate Rhizosphere Microbial Populations and Improve N Uptake Efficiency, Yield, and Nutritional Quality of Leafy Vegetables. Front. Plant Sci. 2018, 9, 743. [Google Scholar] [Green Version]
- Yin, D.C.; Deng, X.; Chet, I. Inhibiting effect and mechanism of Trichoderma virens T43 on four major species of forest pathogen. Chin. J. Ecol. 2014, 33, 1911–1919. [Google Scholar]
- Yin, D.C.; Yang, L.B.; Deng, X.; Chet, I. Song RQ How Trichoderma virens affects growth indicators, physiological and biochemical parameters of Pinus sylvestris var. mongolica seedlings. J. Beijing For. Univ. 2015, 37, 78–83. [Google Scholar]
- Qi, J.Y.; Yin, D.C.; Song, R.Q. Effects of Suillusluteus on Salt Resistance of Pinussylvestris var. mongolica Seedlings. J. Jilin Agric. Univ. 2019, 50, 108–113. [Google Scholar]
- Deng, X.; Song, X.S.; Song, R.Q. Effect of inoculating Phialocephala fortinii D575 and Suillus luteus N94 on the growth of Pinus sylvestris var. mongolica and its resistant to damping-off. For. Pest Dis. Jan. 2017, 36, 21–25. [Google Scholar]
- Bao, S.D. Soil and Plant Analysis, 3rd ed.; Agricultural Press of China: Beijing, China, 2000; pp. 25–109. [Google Scholar]
- Collignon, C.; Uroz, S.; Turpault, M.P.; Frey-Klett, P. Seasons differently impact the structure of mineral weathering bacterial communities in beech and spruce stands. Soil Biol. Biochem. 2011, 43, 2012–2022. [Google Scholar] [CrossRef]
- Shanmugam, V.; Verma, R.; Rajkumar, S.; Naruka, D.S. Bacterial diversity and soil enzyme activity in diseased and disease free apple rhizosphere soils. Ann. Microbiol. 2011, 61, 765–772. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Uroz, S.; Oger, P.; Tisserand, E.; Cébron, A.; Turpault, M.P.; Buée, M.; Boer, W.D.; Leveau, J.H.J.; Frey-Klett, P. Specific impacts of beech and Norway spruce on the structure and diversity of the rhizosphere and soil microbial communities. Sci. Rep. 2016, 6, 27756. [Google Scholar] [CrossRef]
- Yang, L.B.; Sui, X.; Zhu, D.G.; Cui, F.X.; Li, J.B.; Song, R.Q.; Ni, H.W. Study on fungal communities characteristics of different Larixgmelini forest types in cold temperate zone. J. Cent. South Univ. For. Technol. 2017, 37, 12. [Google Scholar]
- Sui, X.; Zhang, R.T.; Yang, L.B.; Xu, N.; Chai, C.R.; Wang, J.F.; Fu, X.L.; Zhong, H.X.; Xing, J.H.; Zhang, Y.; et al. Effect of simulation nitrogen deposition on bacterial diversity of Deyeuxia angustifolia in wetland of Sanjiang plain. Pratacult. Sci. 2016, 33, 589–598. [Google Scholar]
- Yong, C.; Jiang, Y.M.; Huang, H.; Muo, L.C.; Ru, J.L.; Zhao, J.H.; Xiao, S. Long-term and high-concentration heavy-metal contamination strongly influences the microbiome and functional genes in Yellow River sediments. Sci. Total Environ. 2018, 637, 1400–1412. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Deng, Y.; Shen, L.N.; Wen, C.Q.; Wen, C.Q.; Yan, Q.Y.; Ning, D.L.; Qin, Y.J.; Xue, K.; Wu, L.Y.; et al. Temperature mediates continental-scale diversity of microbes in forest soils. Nat. Commun. 2016, 7, 12083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef] [PubMed]
- De, S.R.; Adriana, A.; Passaglia, L.M.P. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar]
- Berg, G. Plant–microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Mastouri, F.; Björkman, T.; Harman, G.E. Seed treatment with Trichoderma harzianum alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef]
- Zaidi, N.W.; Dar, M.H.; Singh, S.; Singh, U.S. Chapter 38-Trichoderma Species as Abiotic Stress Relievers in Plants. In Biotechnology & Biology of Trichoderma; Elsevier: Amsterdam, The Netherlands, 2014; pp. 515–525. [Google Scholar]
- Harman, G.E. Trichoderma—Not just for biocontrol anymore. Phytoparasitica 2011, 39, 103–108. [Google Scholar] [CrossRef]
- Howell, C.R.; Hanson, L.E.; Stipanovic, R.D. Induction of terpenoid synthesis in cotton roots and control of rhizoctonia solani by seed treatments with trichodermavirens. Phytopathology 2000, 90, 248–252. [Google Scholar] [CrossRef]
- Bal, U.; Altintas, S. Effects of Trichoderma harzianum on the yield and fruit quality of tomato plants (Lycopersicon esculentum) grown in an unheated greenhouse. Aust. J. Exp. Agric. 2006, 46, 131–136. [Google Scholar] [CrossRef]
- Shen, Z.; Ruan, Y.; Chao, X.; Zhang, J.; Li, R.; Shen, Q. Rhizosphere microbial community manipulated by 2 years of consecutive biofertilizer application associated with banana Fusarium wilt disease suppression. Biol. Fertil. Soils 2015, 51, 553–562. [Google Scholar] [CrossRef]
- Fu, L.; Penton, C.R.; Ruan, Y.; Shen, Z.; Xue, C.; Li, R.; Shen, Q. Inducing the rhizosphere microbiome by biofertilizer application to suppress banana Fusarium wilt disease. Soil Biol. Biochem. 2017, 104, 39–48. [Google Scholar] [CrossRef]
- Xiong, W.; Guo, S.; Jousset, A.; Zhao, Q.; Shen, Q. Bio-fertilizer application induces soil suppressiveness against Fusarium wilt disease by reshaping the soil microbiome. Soil Biol. Biochem. 2017, 114, 238–247. [Google Scholar] [CrossRef]
- Huang, X.; Chen, L.; Ran, W.; Shen, Q.; Yang, X. Trichoderma harzianum strain SQR-T37 and its bio-organic fertilizer could control Rhizoctonia solani damping-off disease in cucumber seedlings mainly by the mycoparasitism. Appl. Microbiol. Biotechnol. 2011, 91, 741–755. [Google Scholar] [CrossRef] [PubMed]
- Casimiro, I.; Marchant, A.; Bhalerao, R.P.; Beeckman, T.; Dhooge, S.; Swarup, R.; Neil, G.; Inze, D.; Sandberg, G.; Pedro, P.J.; et al. Auxin Transport Promotes Arabidopsis Lateral Root Initiation. Plant Cell 2001, 13, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotasthane, A.; Agrawal, T.; Kushwah, R.; Rahatkar, O.V. In-vitro antagonism of Trichoderma spp. Against Sclerotium rolfsii and Rhizoctonia solani and their response towards growth of cucumber, bottle gourd and bitter gourd. Eur. J. Plant Pathol. 2015, 141, 523–543. [Google Scholar] [CrossRef]
- José, L.R.; Guerrero, R.F.; Rosario, H.G.; Collado, I.G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 2008, 7, 89–123. [Google Scholar]
- Hoagland, R.E. Toxic Metabolites of Fungal Biocontrol Agents. Fungi as Biocontrol Agents Progress Problems & Potential; CABI: Wallingford, UK, 2001; Volume 149, pp. 46–331. [Google Scholar]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Barbetti, M.J.; Li, H.; Woo, S.L.; Lorito, M. A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol. Mol. Plant Pathol. 2008, 72, 80–86. [Google Scholar] [CrossRef]
- Li, R.X.; Cai, F.; Pang, G.; Shen, Q.R.; Li, R.; Chen, W. Solubilisation of Phosphate and Micronutrients by Trichoderma harzianum and Its Relationship with the Promotion of Tomato Plant Growth. PLoS ONE 2015, 10, e0130081. [Google Scholar] [CrossRef]
- Elkatatny, M.H. Enzyme production and nitrogen fixation by free, immobilized and coimmobillzed inoculants of Trichoderma harzianum and Azospirillum brasilense and their possible role in growth promotion of tomato. Food Technol. Biotechnol. 2010, 48, 161–174. [Google Scholar]
- Pelagio-Flores, R.; Esparza-Reynoso, S.; Amira, G.V.; López-Bucio, J.; Alfredo, H.E. richoderma-Induced Acidification Is an Early Trigger for Changes in Arabidopsis Root Growth and Determines Fungal Phytostimulation. Front. Plant Sci. 2017, 8, 822. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Mondejar, R.; Bernal-Vicente, A.; Ros, M.; Tittarelli, F.; Canali, S.; Intrigiolo, F.; Pascual, J.A. Utilisation of citrus compost-based growing media amended with Trichoderma harzianum T-78 in Cucumis melo L. seedling production. Bioresour. Technol. 2010, 101, 3718–3723. [Google Scholar] [CrossRef] [PubMed]
- Yedidia, I.; Srivastva, A.K.; Kapulnik, Y.; Chet, I. Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants. Plant Soil 2001, 235, 235–242. [Google Scholar] [CrossRef]
- Li, Y.T.; Hwang, S.G.; Huang, Y.M.; Huang, C.H. Effects of Trichoderma asperellum on nutrient uptake and Fusarium wilt of tomato. Crop Prot. 2017, 110, 275–282. [Google Scholar] [CrossRef]
- Zhai, T.; Wang, Y.; Liu, Z.; Zhao, M.; Chang, Y.; Zhang, R. Trichoderma asperellum ACCC30536 inoculation improves soil nutrition and leaf artemisinin production in Artemisia annua. Acta Physiol. Plant. 2019, 41, 46. [Google Scholar] [CrossRef]
- Yu, L.; Nicolaisen, M.; Larsen, J.; Ravnskov, S. Organic fertilization alters the community composition of root associated fungi in Pisum sativum. Soil Biol. Biochem. 2013, 58, 36–41. [Google Scholar] [CrossRef]
- Galletti, S.; Fornasier, F.; Cianchetta, S.; Lazzeri, L. Soil incorporation of brassica materials and seed treatment with Trichoderma harzianum: Effects on melon growth and soil microbial activity. Ind. Crops Prod. 2015, 75, 73–78. [Google Scholar] [CrossRef]
- McLean, K.L.; Dodd, S.L.; Minchin, R.F.; Ohkura, M.; Bienkowski, D.; Stewart, A. Non-target impacts of the biocontrol agent Trichoderma atroviride on plant health and soil microbial communities in two native ecosystems in New Zealand. Aust. Plant Pathol. 2013, 43, 33–45. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | CK | ZT05 | E15 |
|---|---|---|---|
| Root length/cm | 68.14 ± 0.81 B | 85.25 ± 0.57 A | 70.48 ± 1.88 B |
| Surface area/cm2 | 11.59 ± 0.10 C | 22.97 ± 0.72 A | 13.70 ± 0.31 B |
| Average diameter/mm | 0.53 ± 0.00 C | 0.56 ± 0.01 A | 0.55 ± 0.00 B |
| Apical number | 100.90 ± 2.13 C | 147.20 ± 0.99 A | 123.20 ± 2.09 B |
| Bifurcation number | 17.20 ± 0.36 C | 30.00 ± 0.63 A | 22.60 ± 0.83 B |
| Index | CK | ZT05 | E15 |
|---|---|---|---|
| OM g/kg | 84.27 ± 0.35 A | 69.77 ± 0.46 C | 77.91 ± 0.67 B |
| TN g/kg | 2.20 ± 0.01 C | 2.61 ± 0.00 A | 2.40 ± 0.01 B |
| AN mg/kg | 206.26 ± 0.03 B | 219.59 ± 0.59 A | 191.41 ± 0.36 C |
| TP g/kg | 1.87 ± 0.00 B | 1.77 ± 0.00 C | 1.95 ± 0.00 A |
| AP mg/kg | 772.14 ± 0.54 B | 796.76 ± 0.54 A | 459.50 ± 2.0 C |
| TK g/kg | 7.08 ± 0.07 A | 7.13 ± 0.09 A | 7.06 ± 0.12 A |
| AK mg/kg | 235.86 ± 0.26 A | 161.41 ± 0.48 B | 135.38 ± 0.07 C |
| pH value | 5.68 ± 0.00 B | 5.75 ± 0.01 A | 5.74 ± 0.01 A |
| Sucrase activity U/g | 4.27 ± 0.01 C | 20.82 ± 0.01 A | 14.45 ± 0.04 B |
| Catalase activity U/g | 5.15 ± 0.03 C | 5.97 ± 0.00 A | 5.57 ± 0.05 B |
| Acid phosphatase activity U/g | 3.81 ± 0.09 C | 8.78 ± 0.02 A | 5.06 ± 0.02 B |
| Urease activity U/g | 938.26 ± 0.08 C | 1295.74 ± 0.06 A | 1094.43 ± 2.42 B |
| Samples | Shannon Index | Simpson Index | Chao1 Index | ACE Index | Coverage % |
|---|---|---|---|---|---|
| CK | 2.85 ± 0.44 A | 0.151 ± 0.07 A | 260.04 ± 16.39 A | 257.51 ± 17.23 A | 0.99 ± 0.00 A |
| ZT05 | 1.21 ± 0.15 B | 0.54 ± 0.08 B | 116.24 ± 3.43 B | 119.25 ± 3.71 B | 0.99 ± 0.00 A |
| E15 | 0.58 ± 0.02 B | 0.71 ± 0.01 B | 82.72 ± 5.71 B | 100.71 ± 12.45 B | 0.99 ± 0.00 A |
| p | 0.002 | 0.001 | 0.00 | 0.00 | 0.03 |
| pH Value | Organic Matter | Available Nitrogen | Total Nitrogen | Available Phosphorus | Total Phosphorus | Available Potassium | Total Potassium | Sucrase Activity | Catalase Activity | Urease Activity | Acid Phosphatase Activity | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ace | −0.97 | 0.76 | 0.29 | −0.80 | 0.55 | −0.04 | 1.00 | −0.13 | −0.88 | −0.82 | −0.76 | −0.61 |
| Chao1 | −0.95 | 0.72 | 0.36 | −0.75 | 0.60 | −0.12 | 1.00** | −0.64 | −0.84 | −0.77 | −0.71 | −0.56 |
| Shannon | −0.92 | 0.65 | 0.49 | −0.69 | 0.68 | −0.21 | 1.00 | 0.03 | −0.79 | −0.71 | −0.64 | −0.47 |
| Simpson | 0.90 | −0.62 | −0.48 | 0.66 | −0.71 | 0.25 | −0.99 | −0.07 | 0.76 | 0.68 | 0.61 | 0.44 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of Two Trichoderma Strains on Plant Growth, Rhizosphere Soil Nutrients, and Fungal Community of Pinus sylvestris var. mongolica Annual Seedlings. Forests 2019, 10, 758. https://doi.org/10.3390/f10090758
Halifu S, Deng X, Song X, Song R. Effects of Two Trichoderma Strains on Plant Growth, Rhizosphere Soil Nutrients, and Fungal Community of Pinus sylvestris var. mongolica Annual Seedlings. Forests. 2019; 10(9):758. https://doi.org/10.3390/f10090758
Chicago/Turabian StyleHalifu, Saiyaremu, Xun Deng, Xiaoshuang Song, and Ruiqing Song. 2019. "Effects of Two Trichoderma Strains on Plant Growth, Rhizosphere Soil Nutrients, and Fungal Community of Pinus sylvestris var. mongolica Annual Seedlings" Forests 10, no. 9: 758. https://doi.org/10.3390/f10090758