
Identification and Transferability of Tetracycline Resistance in Streptococcus thermophilus during Milk Fermentation, Storage, and Gastrointestinal Transit



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Minimum Inhibitory Concentration (MIC) Determination
2.3. Identification of Antibiotic Resistance Genes
2.4. Identification of Genomic Islands and Mobile Elements
2.5. Transferability of Resistance Genes during Strain Fermentation and Storage
2.6. Transferability of Resistance Genes during Gastrointestinal Transit
3. Results
3.1. Minimal Inhibitory Concentrations for S. thermophilus Strains
3.2. Antibiotic Resistance Genes Investigation
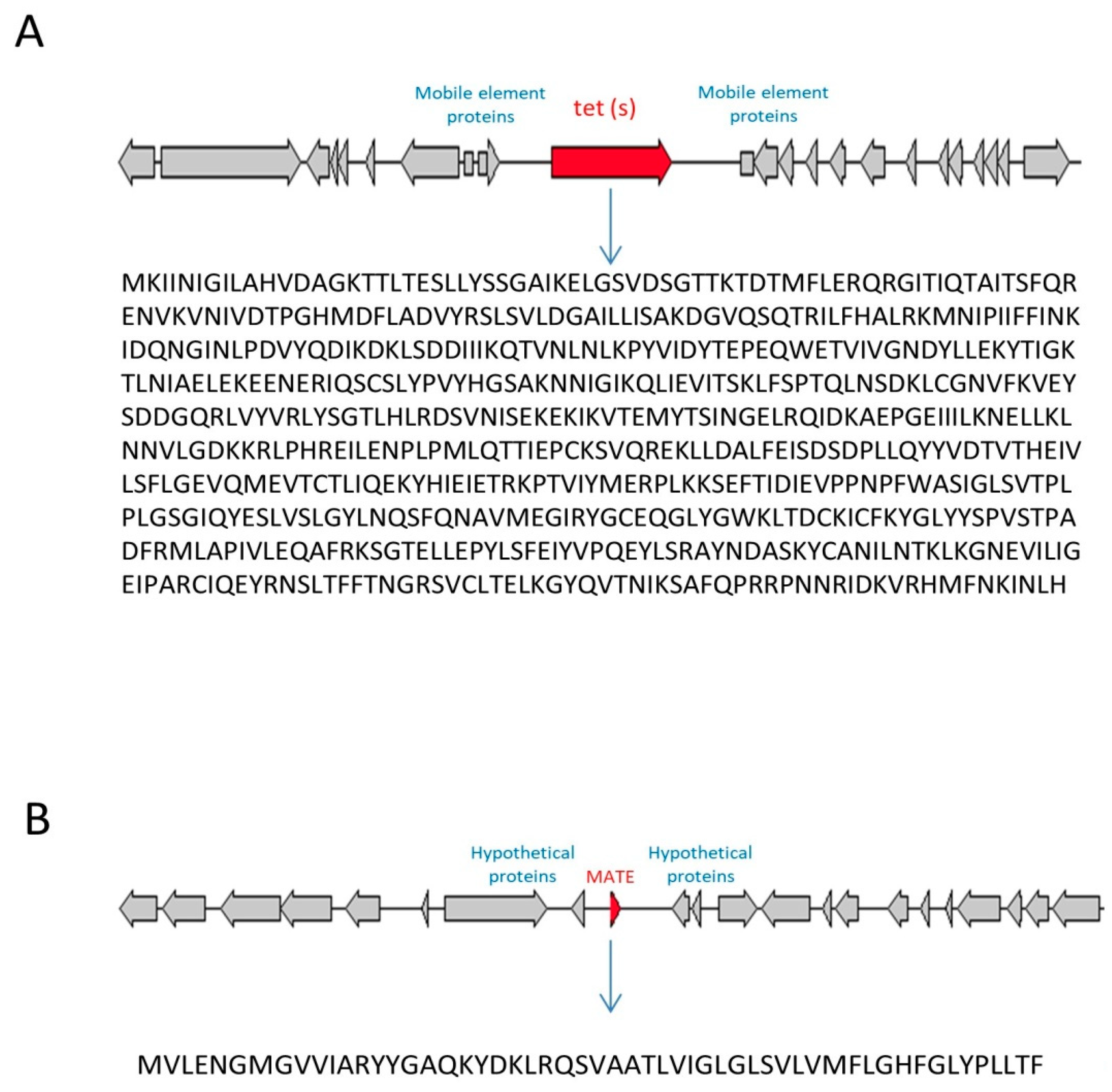
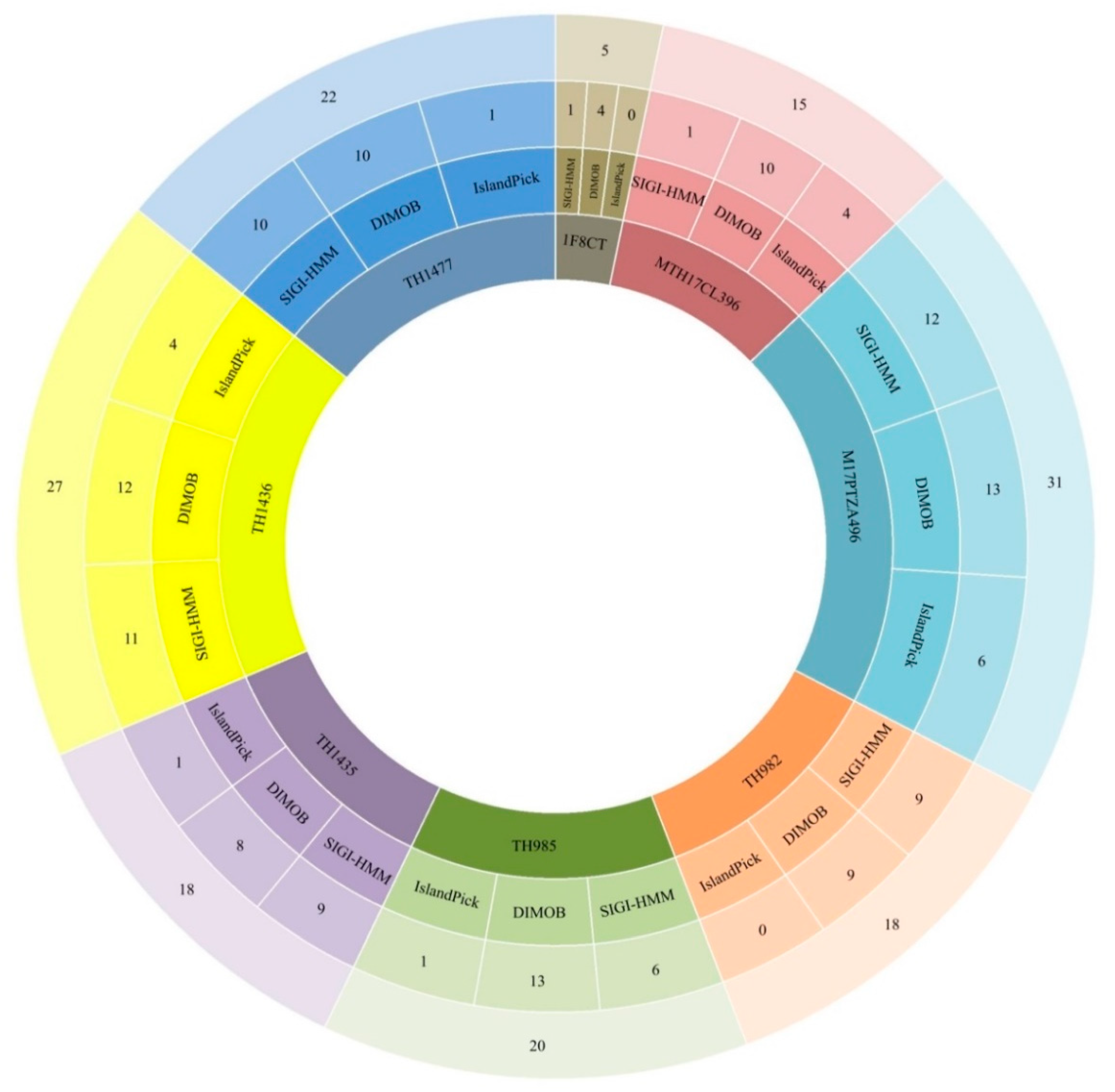
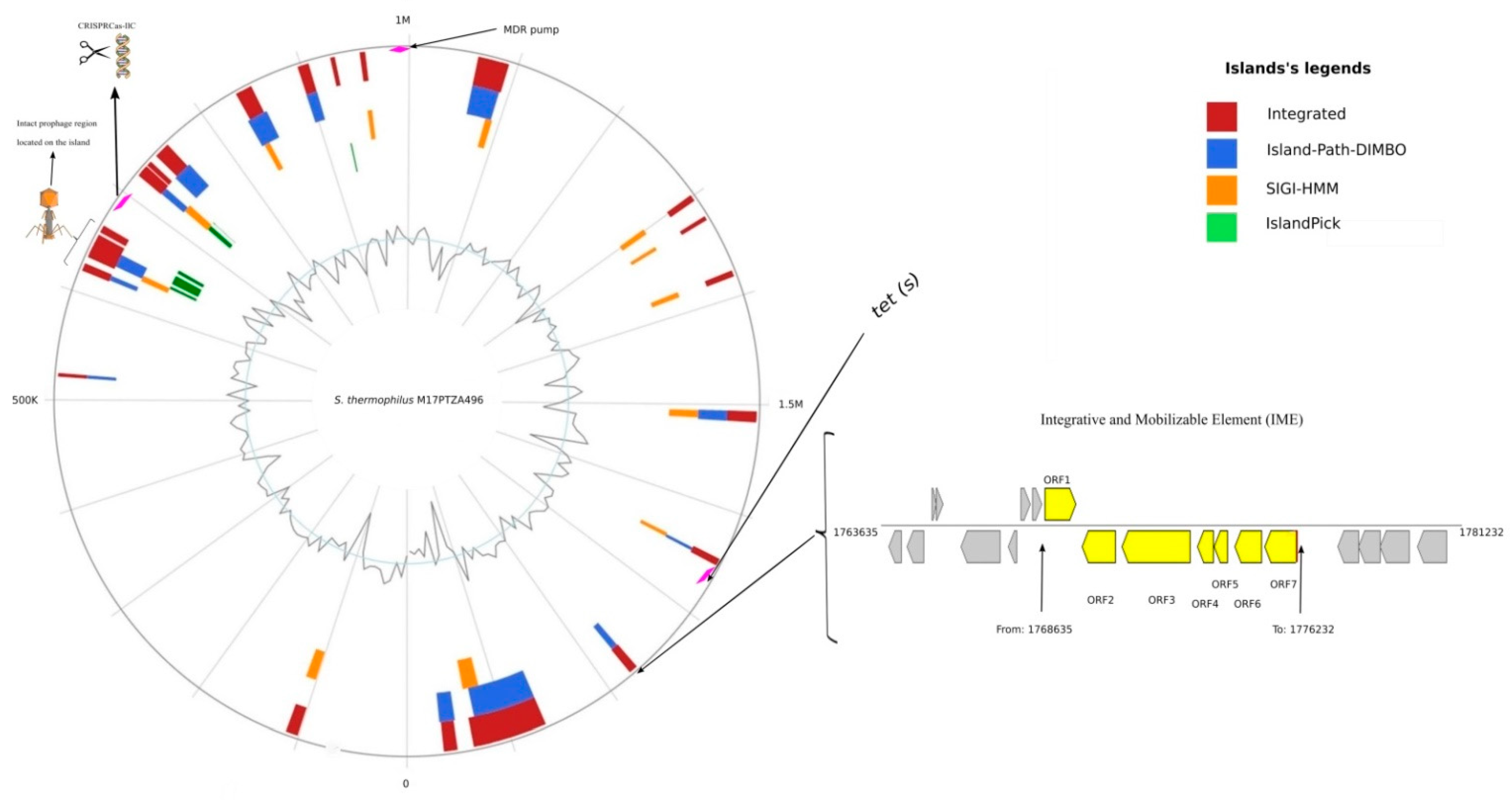
3.3. Identification of Genomic Islands (GI) and Mobile Elements
3.4. Horizontal Transfer of the Tet(S) Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Watve, M.G.; Tickoo, R.; Jog, M.M.; Bhole, B.D. How many antibiotics are produced by the genus Streptomyces? Arch. Microbiol. 2001, 176, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Aminov, R. History of antimicrobial drug discovery: Major classes and health impact. Biochem. Pharmacol. 2017, 133, 4–19. [Google Scholar] [CrossRef]
- Campedelli, I.; Mathur, H.; Salvetti, E.; Clarke, S.; Rea, M.C.; Torriani, S.; Ross, R.P.; Hill, C.; O’Toole, P.W. Genus-Wide assessment of antibiotic resistance in Lactobacillus spp. Appl. Environ. Microbiol. 2019, 85, e01738-18. [Google Scholar]
- Flórez, A.B.; Mayo, B. Antibiotic Resistance-Susceptibility profiles of Streptococcus thermophilus isolated from raw milk and genome analysis of the genetic basis of acquired resistances. Front. Microbiol. 2017, 8, 2608. [Google Scholar] [CrossRef]
- Devirgiliis, C.; Caravelli, A.; Coppola, D.; Barile, S.; Perozzi, G. Antibiotic resistance and microbial composition along the manufacturing process of Mozzarella di Bufala Campana. Int. J. Food Microbiol. 2008, 128, 378–384. [Google Scholar] [CrossRef]
- Soares-Santos, V.; Barreto, A.S.; Semedo-Lemsaddek, T. Characterization of Enterococci from Food and Food-Related Settings. J. Food Prot. 2015, 78, 1320–1326. [Google Scholar] [CrossRef]
- Tarrah, A.; Pakroo, S.; Junior, W.J.F.L.; Guerra, A.F.; Corich, V.; Giacomini, A. Complete Genome Sequence and Carbohydrates-Active EnZymes (CAZymes) Analysis of Lactobacillus paracasei DTA72, a Potential Probiotic Strain with Strong Capability to Use Inulin. Curr. Microbiol. 2020, 77, 2867–2875. [Google Scholar] [CrossRef]
- Tarrah, A.; Pakroo, S.; Corich, V.; Giacomini, A. Whole-Genome sequence and comparative genome analysis of Lactobacillus paracasei DTA93, a promising probiotic lactic acid bacterium. Arch. Microbiol. 2020, 202, 1997–2003. [Google Scholar] [CrossRef]
- Gómez-Gómez, C.; Blanco-Picazo, P.; Brown-Jaque, M.; Quirós, P.; Rodríguez-Rubio, L.; Cerdà-Cuellar, M.; Muniesa, M. Infectious phage particles packaging antibiotic resistance genes found in meat products and chicken feces. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Liu, M.; Li, X.; Xie, Y.; Bi, D.; Sun, J.; Li, J.; Tai, C.; Deng, Z.; Ou, H.-Y. ICEberg 2.0: An updated database of bacterial integrative and conjugative elements. Nucleic Acids Res. 2019, 47, D660–D665. [Google Scholar] [CrossRef]
- Guédon, G.; Libante, V.; Coluzzi, C.; Payot-Lacroix, S.; Leblond-Bourget, N. The Obscure World of Integrative and Mobilizable Elements, Highly Widespread Elements that Pirate Bacterial Conjugative Systems. Genes 2017, 8, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellanger, X.; Payot, S.; Leblond-Bourget, N.; Guédon, G. Conjugative and mobilizable genomic islands in bacteria: Evolution and diversity. FEMS Microbiol. Rev. 2014, 38, 720–760. [Google Scholar] [CrossRef] [Green Version]
- Irlinger, F.; Mounier, J. Microbial interactions in cheese: Implications for cheese quality and safety. Curr. Opin. Biotechnol. 2009, 20, 142–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.; Novak, J.; Knocke, W.; Pruden, A. Elevation of antibiotic resistance genes at cold temperatures: Implications for winter storage of sludge and biosolids. Lett. Appl. Microbiol. 2014, 59, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Schjørring, S.; Krogfelt, K.A. Assessment of Bacterial Antibiotic Resistance Transfer in the Gut. Int. J. Microbiol. 2011, 2011, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tarrah, A.; Noal, V.; Giaretta, S.; Treu, L.; Duarte, V.D.S.; Corich, V.; Giacomini, A. Effect of different initial pH on the growth of Streptococcus macedonicus and Streptococcus thermophilus strains. Int. Dairy J. 2018, 86, 65–68. [Google Scholar] [CrossRef]
- Tarrah, A.; Noal, V.; Treu, L.; Giaretta, S.; Duarte, V.D.S.; Corich, V.; Giacomini, A. Short communication: Comparison of growth kinetics at different temperatures of Streptococcus macedonicus and Streptococcus thermophilus strains of dairy origin. J. Dairy Sci. 2018, 101, 7812–7816. [Google Scholar] [CrossRef]
- Huddleston, J.R. Horizontal gene transfer in the human gastrointestinal tract: Potential spread of antibiotic resistance genes. Infect. Drug Resist. 2014, 7, 167. [Google Scholar] [CrossRef] [Green Version]
- Mater, D.D.G.; Bretigny, L.; Firmesse, O.; Flores, M.-J.; Mogenet, A.; Bresson, J.-L.; Corthier, G. Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus survive gastrointestinal transit of healthy volunteers consuming yogurt. FEMS Microbiol. Lett. 2005, 250, 185–187. [Google Scholar] [CrossRef] [Green Version]
- Tarrah, A.; Treu, L.; Giaretta, S.; Duarte, V.; Corich, V.; Giacomini, A. Differences in Carbohydrates Utilization and Antibiotic Resistance Between Streptococcus macedonicus and Streptococcus thermophilus Strains Isolated from Dairy Products in Italy. Curr. Microbiol. 2018, 75, 1334–1344. [Google Scholar] [CrossRef] [PubMed]
- Blaiotta, G.; Sorrentino, A.; Ottombrino, A.; Aponte, M. Short communication: Technological and genotypic comparison between Streptococcus macedonicus and Streptococcus thermophilus strains coming from the same dairy environment. J. Dairy Sci. 2011, 94, 5871–5877. [Google Scholar] [CrossRef] [Green Version]
- Zhalnina, K.; Dias, R.; Quadros, P.D.D.; Davis-Richardson, A.; Camargo, F.A.O.; Clark, I.M.; McGrath, S.P.; Hirsch, P.R.; Triplett, E.W. Soil pH Determines Microbial Diversity and Composition in the Park Grass Experiment. Microb. Ecol. 2015, 69, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Iyer, R.; Tomar, S.; Maheswari, T.U.; Singh, R. Streptococcus thermophilus strains: Multifunctional lactic acid bacteria. Int. Dairy J. 2010, 20, 133–141. [Google Scholar] [CrossRef]
- Cho, H.; Park, K.-E.; Kim, K.-S. Genome analysis of Streptococcus salivarius subsp. thermophilus type strain ATCC 19258 and its comparison to equivalent strain NCTC 12958. Arch. Microbiol. 2021, 203, 1843–1849. [Google Scholar] [CrossRef]
- Treu, L.; Vendramin, V.; Bovo, B.; Campanaro, S.; Corich, V.; Giacomini, A. Genome Sequences of Four Italian Streptococcus thermophilus Strains of Dairy Origin. Genome Announc. 2014, 2, e00126-14. [Google Scholar] [CrossRef] [Green Version]
- Treu, L.; Vendramin, V.; Bovo, B.; Campanaro, S.; Corich, V.; Giacomini, A. Genome Sequences of Streptococcus thermophilus Strains MTH17CL396 and M17PTZA496 from Fontina, an Italian PDO Cheese. Genome Announc. 2014, 2, e00067-14. [Google Scholar] [CrossRef] [Green Version]
- Treu, L.; Vendramin, V.; Bovo, B.; Campanaro, S.; Corich, V.; Giacomini, A. Whole-Genome Sequences of Streptococcus thermophilus Strains TH1435 and TH1436, Isolated from Raw Goat Milk. Genome Announc. 2014, 2, e01129-13. [Google Scholar] [CrossRef] [Green Version]
- Tarrah, A.; Duarte, V.D.S.; Pakroo, S.; Corich, V.; Giacomini, A. Genomic and phenotypic assessments of safety and probiotic properties of Streptococcus macedonicus strains of dairy origin. Food Res. Int. 2020, 130, 108931. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. Simon Fraser University Research Computing Group IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Néron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In SilicoDetection and Typing of Plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xie, Y.; Liu, M.; Tai, C.; Sun, J.; Deng, Z.; Ou, H.-Y. oriTfinder: A web-based tool for the identification of origin of transfers in DNA sequences of bacterial mobile genetic elements. Nucleic Acids Res. 2018, 46, W229–W234. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel On Additives And Products Or Substances Used in Animal Feed FEEDAP. Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 2704. [Google Scholar] [CrossRef]
- Vendramin, V.; Treu, L.; Campanaro, S.; Lombardi, A.; Corich, V.; Giacomini, A. Genome comparison and physiological characterization of eight Streptococcus thermophilus strains isolated from Italian dairy products. Food Microbiol. 2017, 63, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Palmer, K.L.; Gilmore, M.S. Multidrug-Resistant Enterococci Lack CRISPR-cas. MBio 2010, 1, e00227-10. [Google Scholar] [CrossRef] [Green Version]
- Ammor, M.S.; Flórez, A.B.; Mayo, B. Antibiotic resistance in non-enterococcal lactic acid bacteria and bifidobacteria. Food Microbiol. 2007, 24, 559–570. [Google Scholar] [CrossRef]
- Panel, E.F. Technical Guidance for assessing the safety of feed additives for the environment. EFSA J. 2008, 6, 842. [Google Scholar]
- Arioli, S.; Guglielmetti, S.; Amalfitano, S.; Viti, C.; Marchi, E.; Decorosi, F.; Giovannetti, L.; Mora, D. Characterization of tetA-like gene encoding for a major facilitator superfamily efflux pump in Streptococcus thermophilus. FEMS Microbiol. Lett. 2014, 355, 61–70. [Google Scholar] [CrossRef]
- Ge, B.; Jiang, P.; Han, F.; Saleh, N.K.; Dhiman, N.; Fedorko, D.P.; Nelson, N.A.; Meng, J. Identification and Antimicrobial Susceptibility of Lactic Acid Bacteria from Retail Fermented Foods. J. Food Prot. 2007, 70, 2606–2612. [Google Scholar] [CrossRef]
- Domig, K.J.; Zycka-Krzesinska, J.; Bardowski, J.; Morelli, L. Molecular assessment of erythromycin and tetracycline resistance genes in lactic acid bacteria and bifidobacteria and their relation to the phenotypic resistance. Int. J. Probiotics Prebiotics 2008, 3, 271–280. [Google Scholar]
- Kim, S.-R.; Nonaka, L.; Suzuki, S. Occurrence of tetracycline resistance genes tet(M) and tet(S) in bacteria from marine aquaculture sites. FEMS Microbiol. Lett. 2004, 237, 147–156. [Google Scholar] [CrossRef]
- Hedayatianfard, K.; Akhlaghi, M.; Sharifiyazdi, H. Detection of tetracycline resistance genes in bacteria isolated from fish farms using polymerase chain reaction. In Veterinary Research Forum; Faculty of Veterinary Medicine, Urmia University: Urmia, Iran, 2014; p. 269. [Google Scholar]
- Mishra, M.N.; Daniels, L. Characterization of the MSMEG_2631 Gene (mmp) Encoding a Multidrug and Toxic Compound Extrusion (MATE) Family Protein in Mycobacterium smegmatis and Exploration of Its Polyspecific Nature Using Biolog Phenotype MicroArray. J. Bacteriol. 2013, 195, 1610–1621. [Google Scholar] [CrossRef] [Green Version]
- Dridi, L.; Tankovic, J.; Petit, J.-C. CdeA of Clostridium difficile, a new multidrug efflux transporter of the MATE family. Microb. Drug Resist. 2004, 10, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Jasir, A.; Tanna, A.; Noorani, A.; Mirsalehian, A.; Efstratiou, A.; Schalen, C. High rate of tetracycline resistance inStreptococcus pyogenes in Iran: An epidemiological study. J. Clin. Microbiol. 2000, 38, 2103–2107. [Google Scholar] [CrossRef] [PubMed]
- Emaneini, M.; Mirsalehian, A.; Beigvierdi, R.; Fooladi, A.A.I.; Asadi, F.; Jabalameli, F.; Taherikalani, M. High Incidence of Macrolide and Tetracycline Resistance among Streptococcus Agalactiae Strains Isolated from Clinical Samples in Tehran, Iran. MAEDICA J. Clin. Med. 2014, 9, 157–161. [Google Scholar]
- Mathur, S.; Singh, R. Antibiotic resistance in food lactic acid bacteria—A review. Int. J. Food Microbiol. 2005, 105, 281–295. [Google Scholar] [CrossRef]
- Rizzotti, L.; Gioia, F.L.; Dellaglio, F.; Torriani, S. Characterization of Tetracycline-Resistant Streptococcus thermophilus Isolates from Italian Soft Cheeses. Appl. Environ. Microbiol. 2009, 75, 4224–4229. [Google Scholar] [CrossRef] [Green Version]
- Duarte, V.D.S.; Giaretta, S.; Campanaro, S.; Treu, L.; Armani, A.; Tarrah, A.; Paula, S.O.D.; Giacomini, A.; Corich, V. A Cryptic Non-Inducible Prophage Confers Phage-Immunity on the Streptococcus thermophilus M17PTZA496. Viruses 2018, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Tarrah, A.; De Castilhos, J.; Rossi, R.C.; Duarte, V.D.S.; Ziegler, D.R.; Corich, V.; Giacomini, A. In Vitro Probiotic Potential and Anti-Cancer Activity of Newly Isolated Folate-Producing Streptococcus thermophilus Strains. Front. Microbiol. 2018, 9, 2214. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Strains | Geographical Region | Isolation Matrix | Genome Size (Mbp) | Reference |
|---|---|---|---|---|
| S. thermophilus ATCC19258T | USA | Cow milk | - | [24] |
| S. thermophilus1F8CT | Veneto, Italy | Mozzarella curd (cow) | 1.74 | [25] |
| S. thermophilus MTH17CL396 | Valle d’Aosta, Italy | Fontina cheese (cow) | 1.82 | [26] |
| S. thermophilus M17PTZA496 | Valle d’Aosta, Italy | Fontina cheese (cow) | 2.13 | [26] |
| S. thermophilus TH982 | Campania, Italy | Mozzarella curd (buffalo) | 1.79 | [25] |
| S. thermophilus TH985 | Campania, Italy | Mozzarella whey (buffalo) | 1.83 | [25] |
| S. thermophilus TH1435 | Friuli Venezia Giulia, Italy | Goat milk | 1.79 | [27] |
| S. thermophilus TH1436 | Friuli Venezia Giulia, Italy | Goat milk | 1.79 | [27] |
| S. thermophilus TH1477 | Veneto, Italy | Cow milk | 1.90 | [25] |
| Antibiotics | EFSA Breakpoint | S. thermophilus Strains | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ATCC19258T | 1F8CT | MTH17CL396 | M17PTZA496 | TH982 | TH985 | TH1435 | TH1436 | TH1477 | ||
| Ampicillin | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Chloramphenicol | 4 | 4 | 4 | 4 | 4 | 2 | 4 | 4 | 4 | 4 |
| Ciprofloxacin | – | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Oxacillin | – | 4 | 1 | 0.5 | 0.5 | 1 | 1 | 8 | 1 | 1 |
| Erythromycin | 2 | <0.125 | <0.125 | <0.125 | <0.125 | <0.125 | <0.125 | <0.125 | <0.125 | <0.125 |
| Gentamycin | 32 | 16 | 16 | 16 | 16 | 8 | 16 | 16 | 16 | 16 |
| Kanamycin | 64 | 64 | 64 | 64 | 32 | 32 | 64 | 64 | 64 | 64 |
| Penicillin G | – | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 |
| Streptomycin | 64 | 32 | 64 | 64 | 32 | 16 | 64 | 32 | 32 | 32 |
| Tetracycline | 4 | 0.125 | 0.25 | 0.25 | 128 | 0.25 | 0.25 | 0.25 | 0.125 | 0.125 |
| Trimethoprim | – | >128 | >128 | >128 | >128 | >128 | >128 | >128 | >128 | >128 |
| Vancomycin | 4 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Neomycin | – | 64 | 32 | 64 | 64 | 32 | 32 | 64 | 64 | 64 |
| Rifampicin | – | 0.25 | 1 | 0.5 | 0.5 | 0.5 | 1 | 0.5 | 0.5 | 0.5 |
| Spectinomycin | – | 64 | 128 | 128 | 128 | 32 | 128 | 64 | 64 | 128 |
| Carbenicillin | – | 4 | 8 | 4 | 4 | 4 | 8 | 8 | 4 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarrah, A.; Pakroo, S.; Corich, V.; Giacomini, A. Identification and Transferability of Tetracycline Resistance in Streptococcus thermophilus during Milk Fermentation, Storage, and Gastrointestinal Transit. Fermentation 2021, 7, 65. https://doi.org/10.3390/fermentation7020065
Tarrah A, Pakroo S, Corich V, Giacomini A. Identification and Transferability of Tetracycline Resistance in Streptococcus thermophilus during Milk Fermentation, Storage, and Gastrointestinal Transit. Fermentation. 2021; 7(2):65. https://doi.org/10.3390/fermentation7020065
Chicago/Turabian StyleTarrah, Armin, Shadi Pakroo, Viviana Corich, and Alessio Giacomini. 2021. "Identification and Transferability of Tetracycline Resistance in Streptococcus thermophilus during Milk Fermentation, Storage, and Gastrointestinal Transit" Fermentation 7, no. 2: 65. https://doi.org/10.3390/fermentation7020065