
Heat Treatment of Reishi Medicinal Mushroom (Ganoderma lingzhi) Basidiocarp Enhanced Its β-glucan Solubility, Antioxidant Capacity and Lactogenic Properties

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Instruments
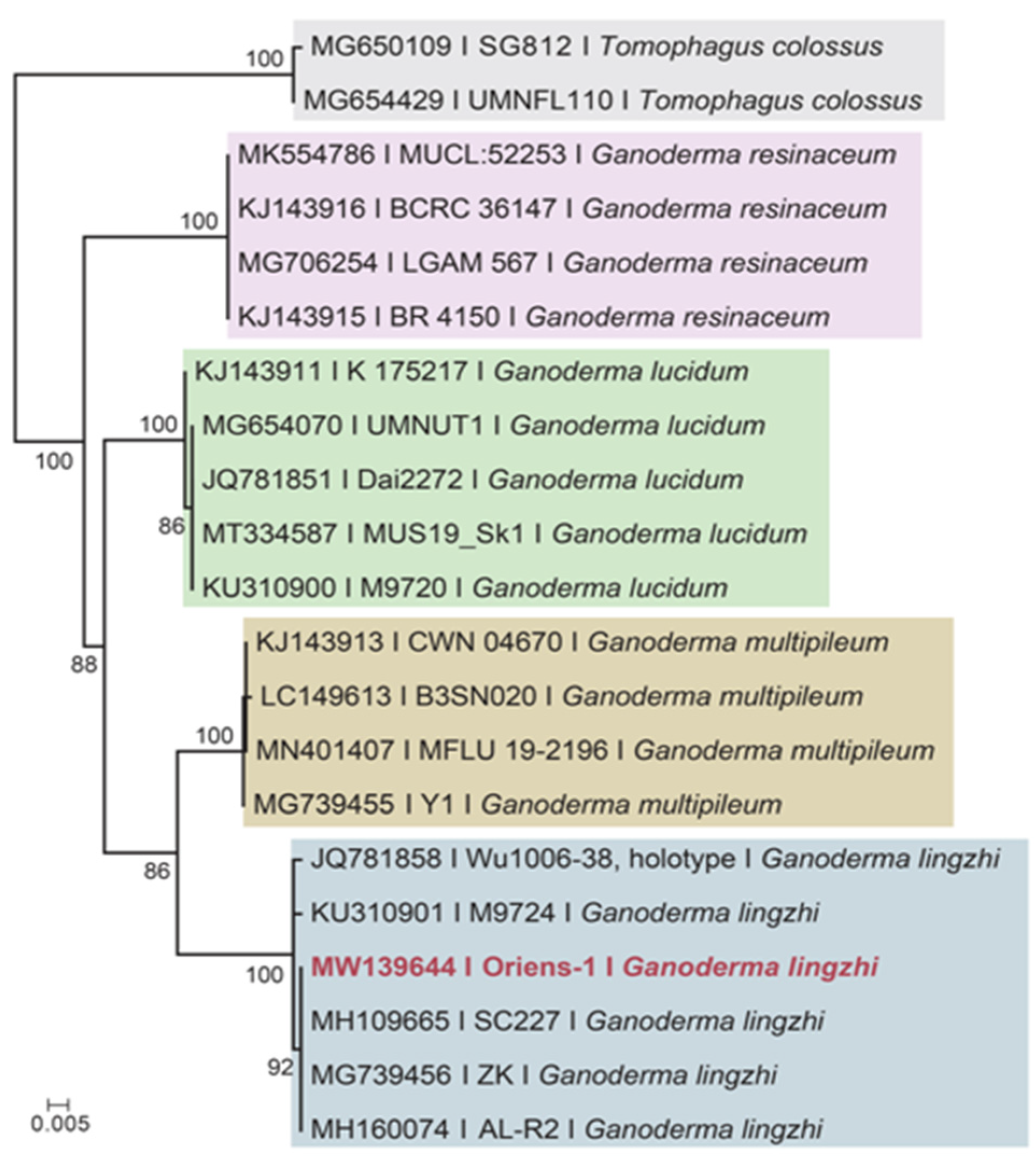
2.2. DNA Molecular Marker and Identification of Reishi Mushroom
2.3. Heat Treatment and Extraction
2.4. Determination of Total Phenolics Content (TPC)
2.5. Free Radical-Scavenging Activity (DPPH and ABTS Assay)
2.6. Measurement of α-glucan and Reducing Sugars
2.7. Measurement of Total and β-glucan Content
2.8. Bacterial Cell Culture and Growth Rate Determination
2.9. Statistics
3. Results and Discussion
3.1. Taxonomical Identification of Reishi Mushroom
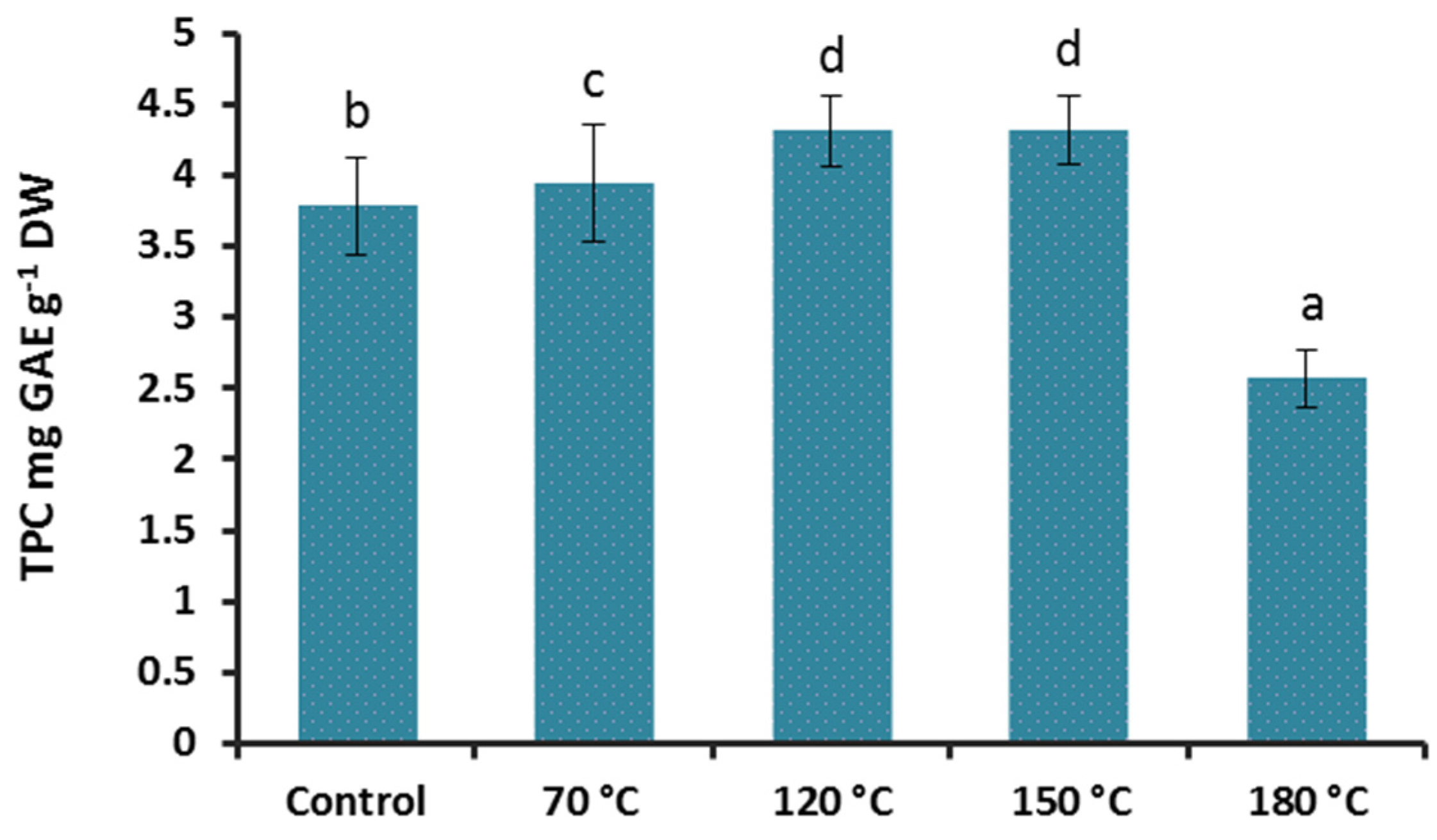
3.2. Effect of Heat Treatment on Reducing Sugars and Total Phenolics Content (TPC) of G. lingzhi
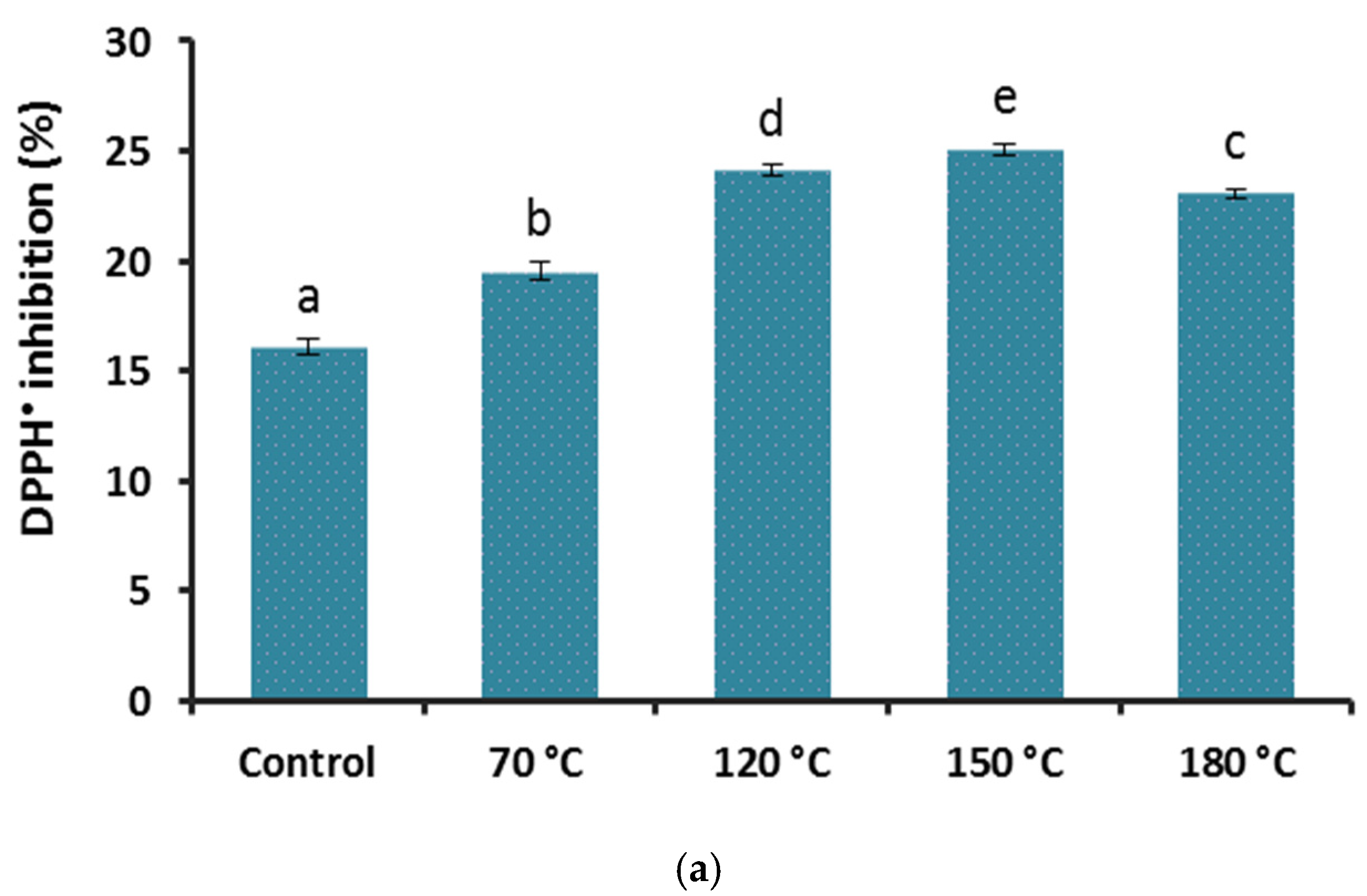
3.3. Effect of Heat Treatment on Antioxidant Properties of G. lingzhi
3.4. Effect of Heat Treatment on Glucans Content of G. lingzhi
3.5. Lactogenic and Antibacterial Properties of Heat-Treated G. lingzhi
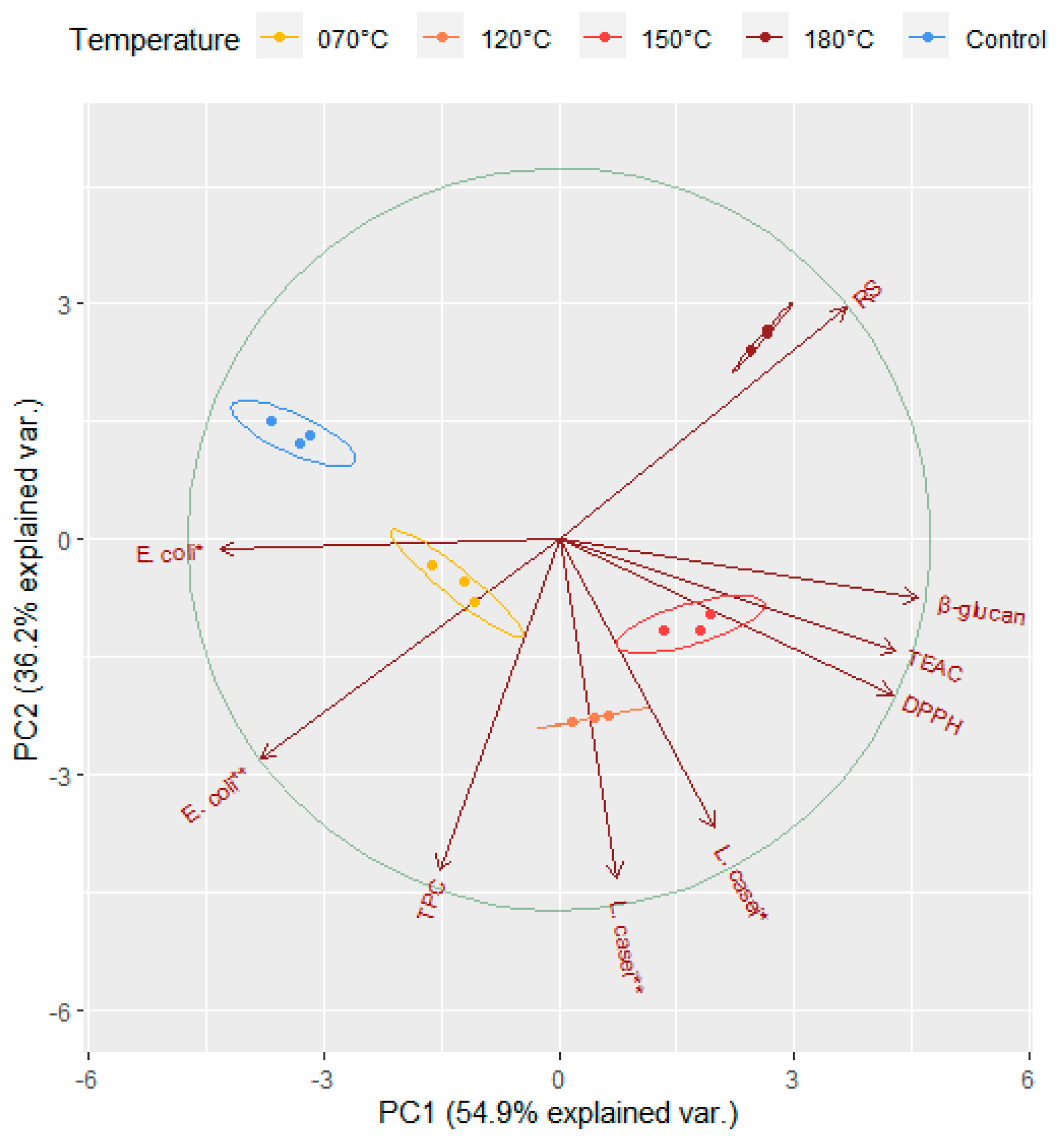
3.6. Principal Component Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Nicoli, M.C.; Anese, M.; Parpinel, M. Influence of processing on the antioxidant properties of fruit and vegetables. Trends Food Sci. Technol. 1999, 10, 94–100. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Liu, R.H. Processed sweet corn has higher antioxidant activity. J. Agric. Food Chem. 2002, 50, 4959–4964. [Google Scholar] [CrossRef]
- Beta, T.; Hwang, T. Influence of heat and moisture treatment on carotenoids, phenolic content, and antioxidant capacity of orange maize flour. Food Chem. 2018, 246, 58–64. [Google Scholar] [CrossRef]
- Kim, M.Y.; Lee, B.W.; Lee, H.U.; Lee, Y.Y.; Kim, M.H.; Lee, J.Y.; Lee, B.K.; Woo, K.S.; Kim, H.J. Phenolic compounds and antioxidant activity in sweet potato after heat treatment. J. Sci. Food Agric. 2019, 99, 6833–6840. [Google Scholar] [CrossRef]
- Yilmaz, Y.; Toledo, R. Antioxidant activity of water-soluble Maillard reaction products. Food Chem. 2005, 93, 273–278. [Google Scholar] [CrossRef]
- Papp, V. Global diversity of the genus Ganoderma: Taxonomic uncertainties and challenges. In Macrofungi: Diversity, Ecology and Biotechnology, 1st ed.; Sridhar, K.R., Deshmukh, S.K., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 10–33. [Google Scholar] [CrossRef]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- De Silva, D.D.; Rapior, S.; Sudarman, E.; Stadler, M.; Xu, J.; Aisyah Alias, S.; Hyde, K.D. Bioactive metabolites from macrofungi: Ethnopharmacology, biological activities and chemistry. Fungal Divers. 2013, 62, 1–40. [Google Scholar] [CrossRef]
- Hapuarachchi, K.K.; Cheng, C.R.; Wen, T.C.; Jeewon, R.; Kakumyan, P. Mycosphere Essays 20: Therapeutic potential of Ganoderma species: Insights into its use as traditional medicine. Mycosphere 2017, 8, 1653–1694. [Google Scholar] [CrossRef]
- Wen, Z.; Xie, J.; Xin, H.; Fang, L.; Qi, W.; Yang, F.; Cai, D.; Niu, Y. Identifcation and quantifcation of triterpenoids in lingzhi or reishi medicinal mushroom, Ganoderma lucidum (Agaricomycetes) with HPLC-MSMS methods. Int. J. Med. Mushrooms. 2018, 20, 919–934. [Google Scholar] [CrossRef]
- Sharma, C.; Bhardwaj, N.; Sharma, A.; Tuli, H.S.; Batra, P.; Beniwal, V.; Gupta, G.K.; Sharma, A.K. Bioactive metabolites of Ganoderma lucidum: Factors, mechanism and broad spectrum therapeutic potential. J. Herb. Med. 2019, 17, 100268. [Google Scholar] [CrossRef]
- Lu, J.; He, R.; Sun, P.; Zhang, F.; Linhardt, R.J.; Zhang, A. Molecular mechanisms of bioactive polysaccharides from Ganoderma lucidum (Lingzhi), a review. Int. J. Biol. Macromol. 2020, 150, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Meneses, M.E.; Martínez-Carrera, D.; Torres, N.; Sánchez-Tapia, M.; Aguilar-López, M.; Morales, P.; Sobal, M.; Bernabé, T.; Escudero, H.; Granados-Portillo, O.; et al. Hypocholesterolemic properties and prebiotic effects of Mexican Ganoderma lucidum in C57BL/6 mice. PLoS ONE. 2016, 11, e0159631. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Huang, G.; Li, X.; Leong, W.; Xia, W.; Hsiao, W.W. Mushroom polysaccharides from Ganoderma lucidum and Poria cocos reveal prebiotic functions. J. Funct. Foods 2018, 41, 191–201. [Google Scholar] [CrossRef]
- Khursheed, R.; Singh, S.K.; Wadhwa, S.; Gulati, M.; Awasthi, A. Therapeutic potential of mushrooms in diabetes mellitus: Role of polysaccharides. Int. J. Biol. Macromol. 2020, 164, 1194–1205. [Google Scholar] [CrossRef]
- Yin, C.; Noratto, G.D.; Fan, X.; Chen, Z.; Yao, F.; Shi, D.; Gao, H. The impact of mushroom polysaccharides on gut microbiota and its beneficial effects to host: A review. Carbohydr. Polym. 2020, 250, 116942. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, A.T.; Wei, Y.; Alneamah, S.J.A.; Al-Shawi, S.G.; Kadir, S.Y.A.; Zainol, J.; Liu, X. What is new in the preventive and therapeutic role of dairy products as nutraceuticals and functional foods? Biomed. Res. Int. 2021, 2021, 8823222. [Google Scholar] [CrossRef]
- Jeong, S.M.; Kim, S.Y.; Kim, D.R.; Jo, S.C.; Nam, K.C.; Ahn, D.U.; Lee, S.C. Effect of heat treatment on the antioxidant activity of extracts from citrus peels. J. Agric. Food Chem. 2004, 52, 3389–3393. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Tenorio, M.L.; Verkerk, R.; van Boekel, M.A.; Dekker, M. Thermal stability of phytochemicals, HMF and antioxidant activity in cape gooseberry (Physalis peruviana L.). J. Funct. Foods 2017, 32, 46–57. [Google Scholar] [CrossRef]
- Kwon, O.C.; Woo, K.S.; Kim, T.M.; Kim, D.J.; Hong, J.T.; Jeong, H.S. Physicochemical characteristics of garlic (Allium sativum L.) on the high temperature and pressure treatment. Korean J. Food Sci. Technol. 2006, 38, 331–336. [Google Scholar]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Choi, Y.; Lee, S.M.; Chun, J.; Lee, H.B.; Lee, J. Influence of heat treatment on the antioxidant activities and polyphenolic compounds of Shiitake (Lentinus edodes) mushroom. Food Chem. 2006, 99, 381–387. [Google Scholar] [CrossRef]
- Gujral, H.S.; Sharma, P.; Rachna, S. Effect of sand roasting on beta glucan extractability, physicochemical and antioxidant properties of oats. LWT 2011, 44, 2223–2230. [Google Scholar] [CrossRef]
- Kim, M.Y.; Yoon, N.; Lee, Y.J.; Woo, K.S.; Kim, H.Y.; Lee, J.; Jeong, H.S. Influence of thermal processing on free and bound forms of phenolics and antioxidant capacity of rice hull (Oryza sativa L.). Prev. Nutr. Food Sci. 2020, 25, 310–318. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Papp, V.; Dima, B. New systematic position of Aurantiporus alborubescens (Meruliaceae, Basidiomycota), a threatened old-growth forest polypore. Mycol. Prog. 2018, 17, 319–332. [Google Scholar] [CrossRef]
- Kiss, A.; Mirmazloum, I.; Naár, Z.; Némedi, E. Supplementation of lingzhi or reishi medicinal mushroom, Ganoderma lucidum (Agaricomycetes) extract enhanced the medicinal values and prebiotic index of Hungarian acacia honey. Int. J. Med. Mushrooms 2019, 21, 1167–1179. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Pavlik, M.; Zhou, S.; Zhang, J.; Tang, Q.; Feng, N.; Kurjak, D.; Pavlík, M., Jr.; Kunca, A. Comparative tnalysis of triterpene composition between Ganoderma lingzhi from China and G. lucidum from Slovakia under different growing conditions. Int. J. Med. Mushrooms 2020, 22, 793–802. [Google Scholar] [CrossRef]
- Ryu, D.H.; Cho, J.Y.; Sadiq, N.B.; Kim, L.C.; Lee, B.; Hamayun, M.; Lee, T.S.; Kim, H.S.; Park, S.H.; Nho, C.W.; et al. Optimization of antioxidant, anti-diabetic, and anti-inflammatory activities and ganoderic acid content of differentially dried Ganoderma lucidum using response surface methodology. Food Chem. 2021, 335, 127645. [Google Scholar] [CrossRef]
- Stojković, D.S.; Barros, L.; Calhelha, R.C.; Glamočlija, J.; Ćirić, A.; Van Griensven, L.J.; Soković, M.; Ferreira, I.C. A detailed comparative study between chemical and bioactive properties of Ganoderma lucidum from different origins. Int. J. Food Sci. 2014, 65, 42–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heleno, S.A.; Barros, L.; Martins, A.; Queiroz, M.J.R.; Santos-Buelga, C.; Ferreira, I.C. Fruiting body, spores and in vitro produced mycelium of Ganoderma lucidum from Northeast Portugal: A comparative study of the antioxidant potential of phenolic and polysaccharidic extracts. Food Res. Int. 2012, 46, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Rufián-Henares, J.A.; Delgado-Andrade, C.; Morales, F.J. Assessing the Maillard reaction development during the toasting process of common flours employed by the cereal products industry. Food Chem. 2009, 114, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.S.; Chon, S.Y.; Bang, W.S.; Kim, M.K. Influence of roasting temperatures on the antioxidant properties, β-glucan content, and volatile flavor profiles of shiitake mushroom. Foods 2021, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Rani, P.; Lal, M.R.; Maheshwari, U.; Krishnan, S. Antioxidant potential of lingzhi or reishi medicinal mushroom, Ganoderma lucidum (Higher Basidiomycetes) cultivated on Artocarpus heterophyllus sawdust substrate in India. Int. J. Med. Mushrooms 2015, 17, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Xu, B.B. Antidiabetic and antioxidant activities of eight medicinal mushroom species from China. Int. J. Med. Mushrooms 2015, 17, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.K.; Dutta, S.D.; Ganguly, K.; Cho, S.J.; Lim, K.T. Mushroom-derived bioactive molecules as immunotherapeutic agents: A review. Molecules 2021, 26, 1359. [Google Scholar] [CrossRef]
- Vetvicka, V.; Teplyakova, T.V.; Shintyapina, A.B.; Korolenko, T.A. Effects of Medicinal Fungi-Derived β-Glucan on Tumor Progression. J. Fungi. 2021, 7, 250. [Google Scholar] [CrossRef]
- Vlassopoulou, M.; Yannakoulia, M.; Pletsa, V.; Zervakis, G.I.; Kyriacou, A. Effects of fungal beta-glucans on health–a systematic review of randomized controlled trials. Food Funct. 2021, 12, 3366–3380. [Google Scholar] [CrossRef]
- Hwang, I.W.; Kim, B.M.; Kim, Y.C.; Lee, S.H.; Chung, S.K. Improvement in β-glucan extraction from Ganoderma lucidum with high-pressure steaming and enzymatic pre-treatment. Appl. Biol. Chem. 2018, 61, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Cortina-Escribano, M.; Pihlava, J.M.; Miina, J.; Veteli, P.; Linnakoski, R.; Vanhanen, H. Effect of strain, wood substrate and cold treatment on the yield and β-glucan content of Ganoderma lucidum fruiting bodies. Molecules 2020, 25, 4732. [Google Scholar] [CrossRef] [PubMed]
- Kivelä, R.; Sontag-Strohm, T.; Loponen, J.; Tuomainen, P.; Nyström, L. Oxidative and radical mediated cleavage of β-glucan in thermal treatments. Carbohydr. Polym. 2011, 85, 645–652. [Google Scholar] [CrossRef]
- Kivelä, R.; Henniges, U.; Sontag-Strohm, T.; Potthast, A. Oxidation of oat β-glucan in aqueous solutions during processing. Carbohydr. Polym. 2012, 87, 589–597. [Google Scholar] [CrossRef]
- Goudar, G.; Sharma, P.; Janghu, S.; Longvah, T. Effect of processing on barley β-glucan content, its molecular weight and extractability. Int. J. Biol. Macromol. 2020, 162, 1204–1216. [Google Scholar] [CrossRef] [PubMed]
- Mohd Hamim, H.M.; Shuhaimi, M.; Yazid, A.M.; Ali, A.M.; Anas, O.M.; Asilah, A.T.; Wahab, M.N.; Shukor, M.Y.A. Growth of probiotic bacteria in trypticase phytone yeast medium supplemented with crude polysaccharides from Ganoderma lucidum. Malays. J. Microbiol. 2010, 6, 47–56. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Reducing Sugar Content (g 100 g−1) |
|---|---|
| Reishi Control | ND |
| Reishi 70 °C | ND |
| Reishi 120 °C | ND |
| Reishi 150 °C | 0.230 ± 0.009 a |
| Reishi 180 °C | 0.568 ± 0.012 b |
| Heat-Treated Samples (20 min) | E. coli (Log CFU) | L. casei (Log CFU) | ||
|---|---|---|---|---|
| Initial Count | End Count | Initial Count | End Count | |
| Reishi Control | 7.075 ± 0.032 Ad | 7.104 ± 0.027 Ac | 4.719 ± 0.101 Aa | 8.209 ± 0.017 Ba |
| Reishi 70 °C | 7.055 ± 0.027 Acd | 7.190 ± 0.126 Ac | 5.556 ± 0.189 Ac | 8.407 ± 0.025 Bb |
| Reishi 120 °C | 7.004 ± 0.032 Aabc | 6.802 ± 0.191 Ac | 5.594 ± 0.027 Ac | 9.334 ± 0.037 Bd |
| Reishi 150 °C | 6.981 ± 0.025 Bab | 5.358 ± 0.320 Ab | 5.456 ± 0.023 Ac | 8.730 ± 0.045 Bc |
| Reishi 180 °C | 6.963 ± 0.014 Ba | 2.774 ± 0.268 Aa | 5.098 ± 0.069 Ab | 8.085 ± 0.100 Bab |
| Frutafit HD inulin | 7.034 ± 0.011 Abcd | 7.033 ± 0.034 Ac | 5.512 ± 0.088 Ac | 9.252 ± 0.033 Bd |
| TEAC | DPPH | β-glucan | E. coli * | E. coli ** | L. casei * | L. casei ** | RS | TPC | |
|---|---|---|---|---|---|---|---|---|---|
| TEAC | \ | 0.958 | 0.911 | −0.761 | −0.557 | 0.571 | 0.367 | 0.533 | 0.038 |
| DPPH | 0.000 | \ | 0.922 | −0.794 | −0.482 | 0.650 | 0.547 | 0.444 | 0.100 |
| β-glucan | 0.000 | 0.000 | \ | −0.826 | −0.672 | 0.579 | 0.291 | 0.648 | −0.219 |
| E. coli * | 0.001 | 0.000 | 0.000 | \ | 0.748 | −0.327 | −0.136 | −0.703 | 0.284 |
| E. coli ** | 0.031 | 0.069 | 0.006 | 0.001 | \ | 0.151 | 0.393 | −0.991 | 0.772 |
| L. casei * | 0.026 | 0.009 | 0.024 | 0.234 | 0.590 | \ | 0.665 | −0.172 | 0.480 |
| L. casei ** | 0.178 | 0.035 | 0.292 | 0.628 | 0.147 | 0.007 | \ | −0.449 | 0.724 |
| RS | 0.041 | 0.098 | 0.009 | 0.003 | 0.000 | 0.539 | 0.093 | \ | −0.793 |
| TPC | 0.894 | 0.723 | 0.432 | 0.305 | 0.001 | 0.070 | 0.002 | 0.000 | \ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiss, A.; Grünvald, P.; Ladányi, M.; Papp, V.; Papp, I.; Némedi, E.; Mirmazloum, I. Heat Treatment of Reishi Medicinal Mushroom (Ganoderma lingzhi) Basidiocarp Enhanced Its β-glucan Solubility, Antioxidant Capacity and Lactogenic Properties. Foods 2021, 10, 2015. https://doi.org/10.3390/foods10092015
Kiss A, Grünvald P, Ladányi M, Papp V, Papp I, Némedi E, Mirmazloum I. Heat Treatment of Reishi Medicinal Mushroom (Ganoderma lingzhi) Basidiocarp Enhanced Its β-glucan Solubility, Antioxidant Capacity and Lactogenic Properties. Foods. 2021; 10(9):2015. https://doi.org/10.3390/foods10092015
Chicago/Turabian StyleKiss, Attila, Petra Grünvald, Márta Ladányi, Viktor Papp, István Papp, Erzsébet Némedi, and Iman Mirmazloum. 2021. "Heat Treatment of Reishi Medicinal Mushroom (Ganoderma lingzhi) Basidiocarp Enhanced Its β-glucan Solubility, Antioxidant Capacity and Lactogenic Properties" Foods 10, no. 9: 2015. https://doi.org/10.3390/foods10092015