Expression of Genes Encoding Manganese Peroxidase and Laccase of Ganoderma boninense in Response to Nitrogen Sources, Hydrogen Peroxide and Phytohormones



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Fungal Cultures
2.2. Enzyme Assays
2.3. Inoculation of Oil Palms with G. boninense in the Presence of Different Nitrogen Sources
2.4. Primer Design from Partial Transcripts
2.5. Total RNA Extraction and RACE-PCR
2.6. Sequence Analysis of Full-Length cDNA
2.7. Quantitative Reverse Transcription-PCR (qRT-PCR)
3. Results and Discussion
3.1. Sequence Analysis of GbMnPs and GbLacs in G. boninense
3.2. Enzyme Activities of MnP and Lac in G. boninense Treated with Different Nitrogen Sources, Phytohormones and H2O2
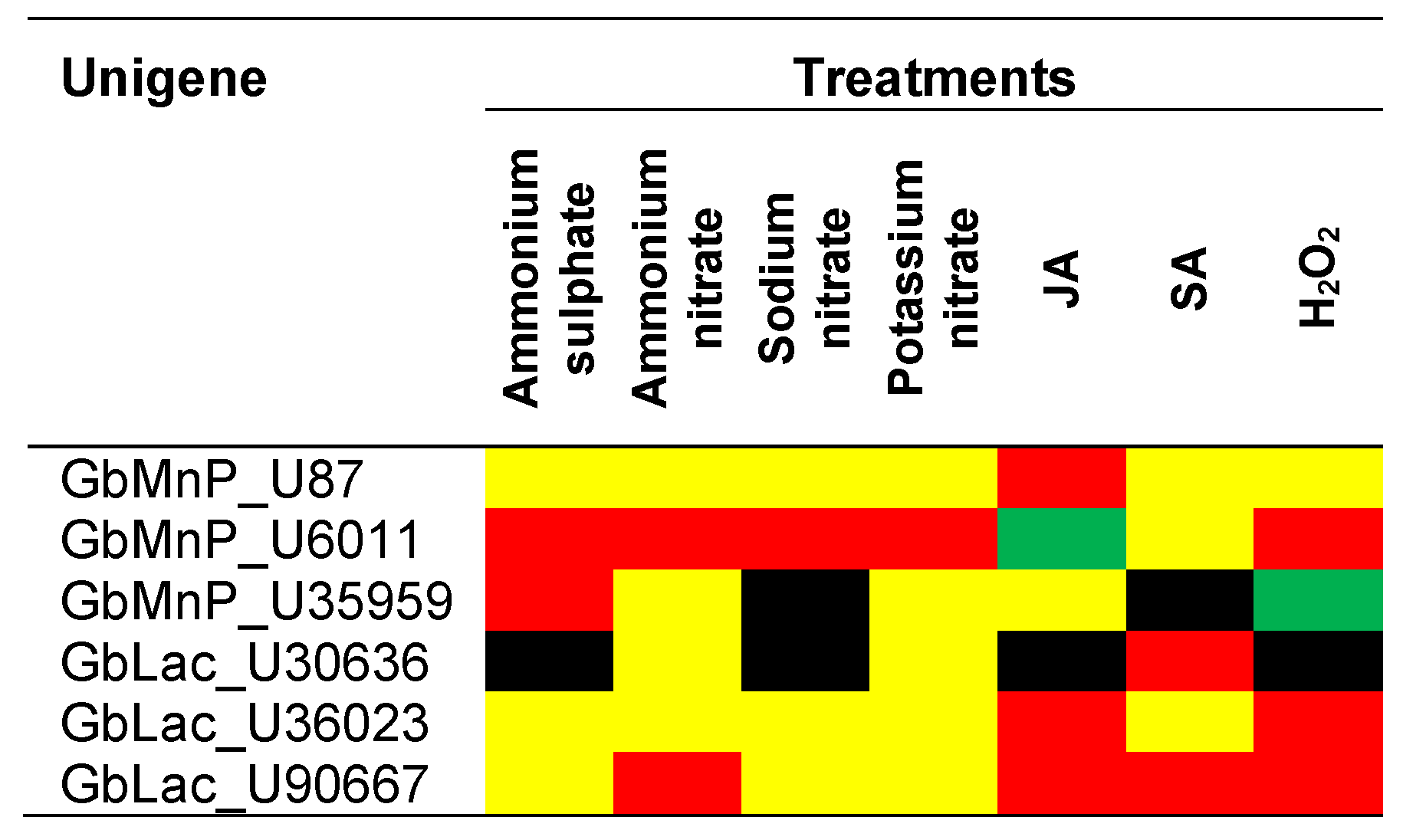
3.3. Transcript Abundance of MnPs and Lacs in G. boninense Treated with Different Nitrogen Sources, Phytohormones and H2O2
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Turner, P.D. Oil Palm Diseases and Disorders; Oxford University Press: Oxford, UK, 1981. [Google Scholar]
- Janusz, G.; Kucharzyk, K.H.; Pawlik, A.; Staszczak, M.; Paszczynski, A.J. Fungal laccase, manganese peroxidase and lignin peroxidase: Gene expression and regulation. Enzyme Microb. Technol. 2013, 52, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Hoyos, C.M.; Morales-Álvarez, E.D.; Poveda-Cuevas, S.A.; Reyes-Guzmán, E.A.; Poutou-Piñales, R.A.; Reyes-Montaño, E.A.; Pedroza-Rodríguez, A.M.; Rodríguez-Vázquez, R.; Cardozo-Bernal, Á.M. Computational analysis and low-scale constitutive expression of laccases synthetic genes GlLCC1 from Ganoderma lucidum and POXA 1B from Pleurotus ostreatus in Pichia pastoris. PLoS ONE 2015, 10, e0116524. [Google Scholar] [CrossRef]
- Martínez, A.T. Molecular biology and structure-function of lignin-degrading heme peroxidases. Enzyme Microb. Technol. 2002, 30, 425–444. [Google Scholar] [CrossRef]
- Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates: An expanded role for laccase in lignin biodegradation. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Eggert, C.; Temp, U.; Eriksson, K.E.L. The ligninolytic system of the white rot fungus Pycnoporus cinnabarinus: Purification and characterization of the laccase. Appl. Environ. Microbiol. 1996, 62, 1151–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giardina, P.; Faraco, V.; Pezzella, C.; Pisutelli, A.; Vanhulle, S.; Sannnia, G. Laccases: A never-ending story. Cell Mol. Life Sci. 2010, 67, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.-M.; Leem, Y.-E.; Choi, H.T. Purification and characterization of laccase isozymes from the white-rot basidiomycete Ganoderma lucidum. Appl. Microbiol. Biotechnol. 2001, 57, 98–102. [Google Scholar] [PubMed]
- Teerapatsakul, C.; Abe, N.; Bucke, C.; Chitradon, L. Novel laccases of Ganoderma sp. KU-Alk4, regulated by different glucose concentration in alkaline media. World J. Microbiol. Biotechnol. 2007, 23, 1559–1567. [Google Scholar] [CrossRef]
- Kumar, A.; Kant, K.; Kumar, P.; Ramchiary, N. Laccase isozymes from Ganoderma lucidum MDU-7: Isolation, characterization, catalytic properties and differential role during oxidative stress. J. Mol. Catal. Enzymat. 2015, 113, 68–75. [Google Scholar] [CrossRef]
- Xu, H.; Guo, M.Y.; Gao, Y.H.; Bai, X.H.; Zhou, X.W. Expression and characteristics of manganese peroxidase from Ganoderma lucidum in Pichia pastoris and its application in the degradation of four dyes and phenol. BMC Biotechnol. 2017, 17, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuo, R.; Ma, L.; Fan, F.; Gong, Y.; Wan, X.; Jiang, M.; Zhang, X.; Yang, Y. Decolorization of different dyes by a newly isolated white-rot fungi strain Ganoderma sp. En3 and cloning and functional analysis of its laccase gene. J. Hazard Mater. 2011, 192, 855–873. [Google Scholar] [CrossRef]
- You, L.; Liu, Z.M.; Lin, J.F.; Guo, L.Q.; Huang, X.L.; Yang, H.X. Molecular cloning of a laccase gene from Ganoderma lucidum and heterologous expression in Pichia pastoris. J. Basic Microbiol. 2013, 53, S134–S141. [Google Scholar] [CrossRef]
- Torres-Farradá, G.; Manzano León, A.M.; Rineau, F.; Ledo Alonso, L.L.; Sánchez-López, M.I.; Thijs, S.; Colpaert, J.; Ramos-Leal, M.; Guerra, G.; Vangronsveld, J. Diversity of ligninolytic enzymes and their genes in strains of the genus Ganoderma: Applicable for biodegradation of xenobiotic compounds? Front. Microbiol. 2017, 8, 898. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.L.; Tan, Y.C.; Yeoh, K.A.; Ghazali, A.K.; Yee, W.Y.; Hoh, C.C. De novo transcriptome analyses of host-fungal interactions in oil palm (Elaeis guineensis Jacq.). BMC Genom. 2016, 17, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magill, A.H.; Aber, J.D. Long-term effects of experimental nitrogen additions on foliar litter decay and humus formation in forest ecosystems. Plant Soil 1998, 203, 301–311. [Google Scholar] [CrossRef]
- Kirk, T.K.; Schultz, E.; Connors, W.J.; Lorenz, L.F.; Zeikus, J.G. Influence of culture parameters on lignin metabolism by Phanerochaete chrysosporium. Arch. Microbiol. 1978, 117, 277–285. [Google Scholar] [CrossRef]
- Dill, I.; Kraepelin, G. Palo Podrido: Model for extensive delignification of wood by Ganoderma applanatum. Appl. Environ. Microbiol. 1986, 52, 1305–1312. [Google Scholar] [CrossRef] [Green Version]
- Rios, S.; Eyzaguirre, J. Conditions for selective degradation of lignin by the fungus Ganoderma australis. Appl. Microbiol. Biotechnol. 1992, 37, 667–669. [Google Scholar] [CrossRef]
- Mohd Tayeb, D.; Idris, A.S.; Mohd Haniff, H. Reduction of Ganoderma infection in oil palm through balanced fertilization in peat. In Proceedings of the International Palm Oil Congress, Putrajaya, Malaysia, 24–28 August 2003; MPOB: Bangi, Malaysia; pp. 193–218. [Google Scholar]
- Pujianto; Achmad, W.S.; Dafian, P.; Syaiful; Suhardi; Putri, A.W.; Caliman, J.P. Impact of mineral nutrition management on Ganoderma incidence in oil palm planted on peat soil. In Proceedings of the 15th International Peat Congress, Kuching, Malaysia, 17–19 August 2016. [Google Scholar]
- De Coninck, B.; Timmermans, P.; Vos, C.; Cammue, B.P.A.; Kazan, K. What lies beneath: Belowground defense strategies in plants. Trends Plant Sci. 2015, 20, 91–101. [Google Scholar] [CrossRef]
- Surendran, A.; Siddiqui, Y.; Saud, H.M.; Ali, N.S.; Manickam, S. Inhibition and kinetic studies of lignin degrading enzymes of Ganoderma boninense by natural occurring phenolic compounds. J. Appl. Microbiol. 2018, 125, 876–887. [Google Scholar] [CrossRef]
- Ong, C.E.; Goh, Y.K.; Tan, S.Y.; Goh, Y.K.; Goh, K.J. A preliminary study on the effects of salicylic and jasmonic acids on Ganoderma boninense growth, mycelial hydrophobicity, and media pH under in vitro assays. Arch. Phytopath. Plant Prot. 2018, 51, 122–127. [Google Scholar] [CrossRef]
- Rahamah Bivi, M.; Siti Noor Farhana, M.D.; Khairulmazmi, A.; Idris, A.; Ahmed, O.H.; Zamri, R.; Sariah, M. In vitro effects of salicylic acid, calcium and copper ions on growth and sporulation of Ganoderma boninense. Afr. J. Biotech. 2012, 11, 13477–13489. [Google Scholar]
- Rahamah Bivi, M.; Paiko, A.S.; Khairulmazmi, A.; Akhtar, M.S.; Idris, A.S. Control of basal stem rot disease in oil palm by supplementation of calcium, copper, and salicylic acid. Plant Pathol. J. 2016, 32, 396–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermek, H.; Li, K.; Eriksson, K.-E. Studies on inactivation and stabilization of manganese peroxidase from Trametes versicolor. In 8th International Conference on Biotechnology in the Pulp and Paper Industry; Viikari, L., Lantto, R., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2002; pp. 141–149. [Google Scholar]
- Chan-Cupul, W.; Arámbula-Zúñiga, C.S.; Fan, Z.; Heredia, G. Oxidative enzymes activity and hydrogen peroxide production in white-rot fungi and soil-borne micromycetes co-cultures. Ann. Microbiol. 2019, 69, 171–181. [Google Scholar]
- Li, D.; Alic, M.; Brown, J.A.; Gold, M.H. Regulation of manganese peroxidase gene transcription by hydrogen peroxide, chemical stress, and molecular oxygen. Appl. Environ. Microbiol. 1995, 61, 341–345. [Google Scholar] [CrossRef] [Green Version]
- Branden, R.; Malmstrom, B.; Vanngard, T. The interaction of fungal laccase with hydrogen peroxide and the removal of fluoride from the inhibited enzyme. Eur. J. Biochem. 1971, 18, 238–241. [Google Scholar] [CrossRef]
- Asther, M.; Lesag, L.; Drapron, R.; Corrieu, G.; Odier, E. Phospholipid and fatty acid enrichment of Phanerochaete chrysosporium INA-12 relation to ligninase production. Appl. Microbiol. Biotechnol. 1988, 27, 393–398. [Google Scholar] [CrossRef]
- Wolfenden, B.S.; Wilson, R.L. Radical cations as reference chromogens in studies of one-electron transfer reactions: Pulse radio analysis studies of 2,2’-azinobis-(3-ethlbenzthiazoline-6-sulfonate). J. Chem. Soc. Perkin Trans. 1982, 2, 805–812. [Google Scholar] [CrossRef]
- Zaiton, S.; Sariah, M.; Zainal, A.M.A. Effect of endophytic bacteria on growth and suppression of Ganoderma infection in oil palm. Int. J. Agric. Biol. 2008, 10, 127–132. [Google Scholar]
- Song, W.; Zhou, L.; Yang, C.; Cao, X.; Zhang, L.; Liu, X. Tomato Fusarium wilt and its chemical control strategies in a hydroponic system. Crop Prot. 2004, 23, 243–247. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, N.; Du, L. Isolation of RNA of high quality and yield from Ginkgo biloba leaves. Biotechnol. Lett. 2005, 27, 629e633. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Lim, F.H.; Nor Fakhrana, I.; Abdul Rasid, O.; Idris, A.S.; Ahmad Parveez, G.K.; Ho, C.L.; Shaharuddin, N.A. Isolation and selection of reference genes for Ganoderma boninense gene expression study using quantitative real-time PCR (qPCR). J. Oil Palm Res. 2014, 26, 170–181. [Google Scholar]
- Tan, Y.C.; Yeoh, K.A.; Wong, M.Y.; Ho, C.L. Expression profiles of putative defence-related proteins in oil palm (Elaeis guineensis) colonized by Ganoderma boninense. J. Plant Physiol. 2013, 170, 1455–1460. [Google Scholar] [CrossRef]
- Ren, A.; Qin, L.; Shi, L.; Dong, X.; Mu, D.S.; Li, Y.X.; Zhao, M.W. Methyl jasmonate induces ganoderic acid biosynthesis in the basidiomycetous fungus Ganoderma lucidum. Bioresour. Technol. 2010, 101, 6785–6790. [Google Scholar] [CrossRef]
- Ren, A.; Li, M.-J.; Shi, L.; Mu, D.-S.; Jiang, A.-L.; Han, Q.; Zhao, M.-W. Profiling and quantifying differential gene transcription provide insights into ganoderic acid biosynthesis in Ganoderma lucidum in response to methyl jasmonate. PLoS ONE 2013, 8, e65027. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, H.F.S.; El-Arab, M.M.; Omar, S.A. Differential effect of jasmonic acid on the defense of faba bean sgainst Fusarium Wilt: Modulation of other phytohormones and simple phenols. Int. J. Agri. Biol. 2002, 4, 447–453. [Google Scholar]
- Sengottayan, S.-N.; Kandaswamy, K.; Choi, M.Y.; Paik, C.-H. Effects of jasmonic acid-induced resistance in rice on the plant brownhopper, Nilaparvata lugens Stål (Homoptera: Delphacidae). Pestic. Biochem. Physiol. 2009, 95, 77–84. [Google Scholar]
- Levine, A.; Tenhaken, R.; Dixon, R.A.; Lamb, C.J. H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell 1994, 79, 583–593. [Google Scholar] [CrossRef]
- Cheeseman, J.M. Hydrogen peroxide concentrations in leaves under natural conditions. J. Exp. Bot. 2006, 57, 2435–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.L.; Liu, Y.L.; Cao, W.X.; Huai, M.F.; Xu, B.G.; Huang, B.G. Effects of salicylic acid on heat tolerance associated with antioxidant metabolism in Kentucky bluegrass. Crop Sci. 2005, 45, 988–995. [Google Scholar] [CrossRef]
- Aver‘yanov, A.A.; Lapikova, V.P.; Pasechnik, T.D.; Kuznetsov, V.V.; Baker, C.J. Suppression of early stages of fungus development by hydrogen peroxide at low concentrations. Plant Pathol. J. 2007, 6, 242–247. [Google Scholar]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef] [PubMed]
- Saxena, I.; Srikanth, S.; Chen, Z. Cross talk between H2O2 and interacting signal molecules under plant stress response. Front. Plant Sci. 2016, 7, 570. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, P.-Y.; Namasivayam, P.; Sundram, S.; Ho, C.-L. Expression of Genes Encoding Manganese Peroxidase and Laccase of Ganoderma boninense in Response to Nitrogen Sources, Hydrogen Peroxide and Phytohormones. Genes 2020, 11, 1263. https://doi.org/10.3390/genes11111263
Ho P-Y, Namasivayam P, Sundram S, Ho C-L. Expression of Genes Encoding Manganese Peroxidase and Laccase of Ganoderma boninense in Response to Nitrogen Sources, Hydrogen Peroxide and Phytohormones. Genes. 2020; 11(11):1263. https://doi.org/10.3390/genes11111263
Chicago/Turabian StyleHo, Pei-Yin, Parameswari Namasivayam, Shamala Sundram, and Chai-Ling Ho. 2020. "Expression of Genes Encoding Manganese Peroxidase and Laccase of Ganoderma boninense in Response to Nitrogen Sources, Hydrogen Peroxide and Phytohormones" Genes 11, no. 11: 1263. https://doi.org/10.3390/genes11111263