
Nodulated White Lupin Plants Growing in Contaminated Soils Accumulate Unusually High Mercury Concentrations in Their Nodules, Roots and Especially Cluster Roots

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soils
2.2. Plant Material, Inoculation and Growth in Hg-Contaminated Soils
2.3. Cluster Root Promotion Conditions
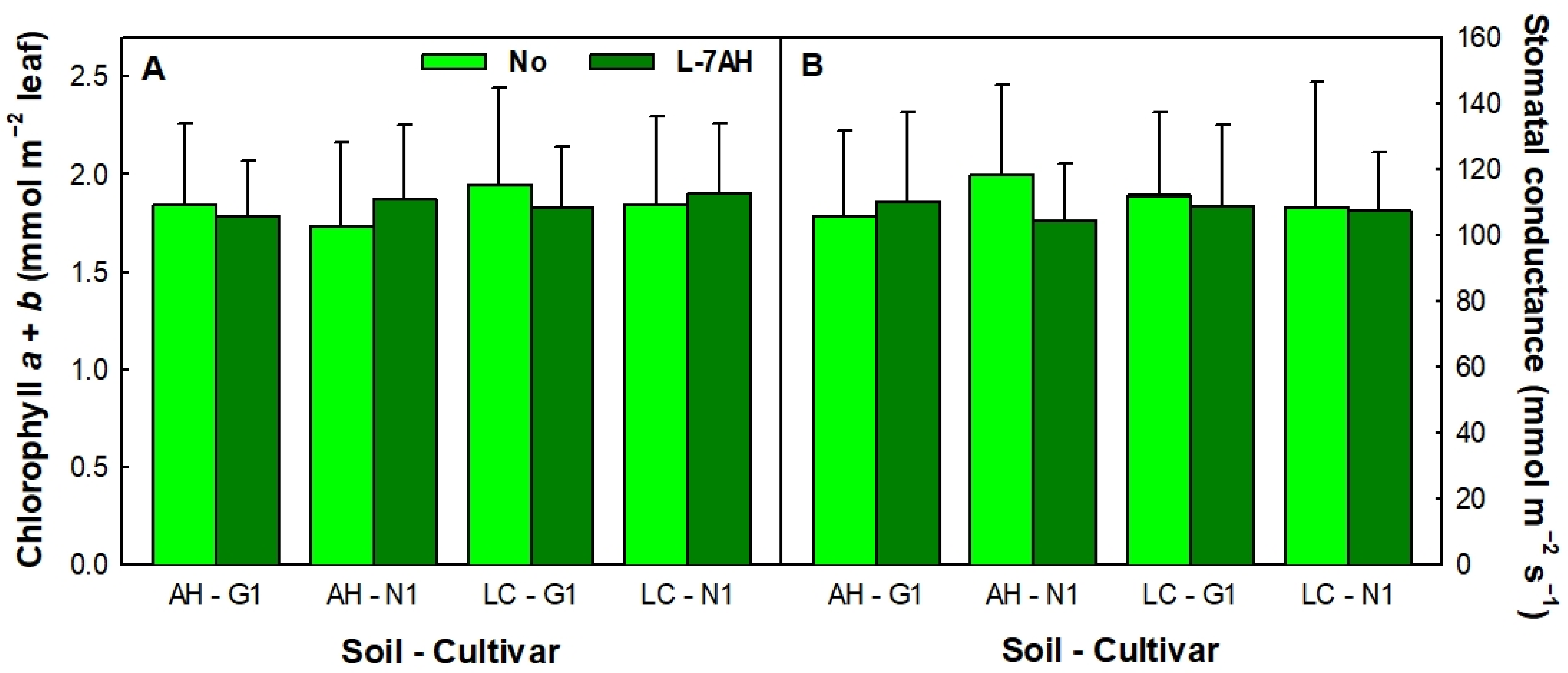
2.4. Chlorophyll Content and Stomatal Conductance
2.5. Sample Collection
2.6. Leaf Area
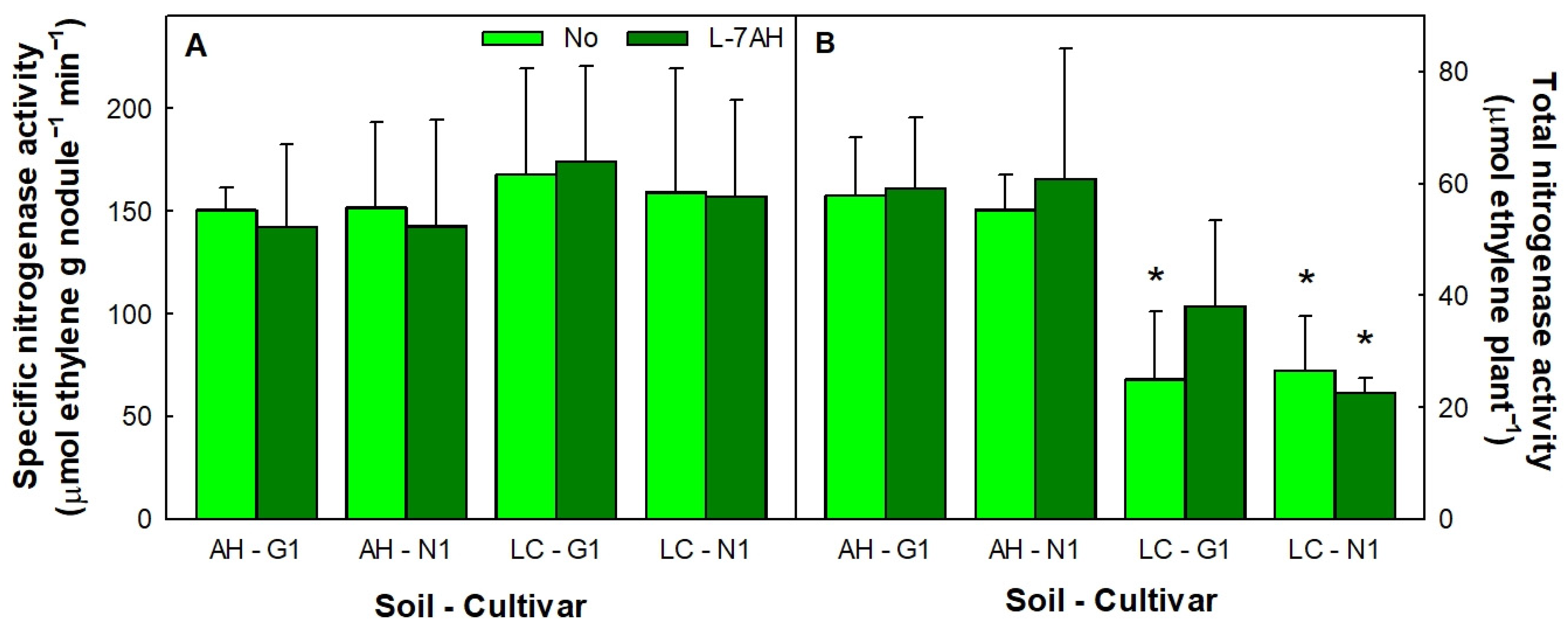
2.7. Nitrogenase Activity
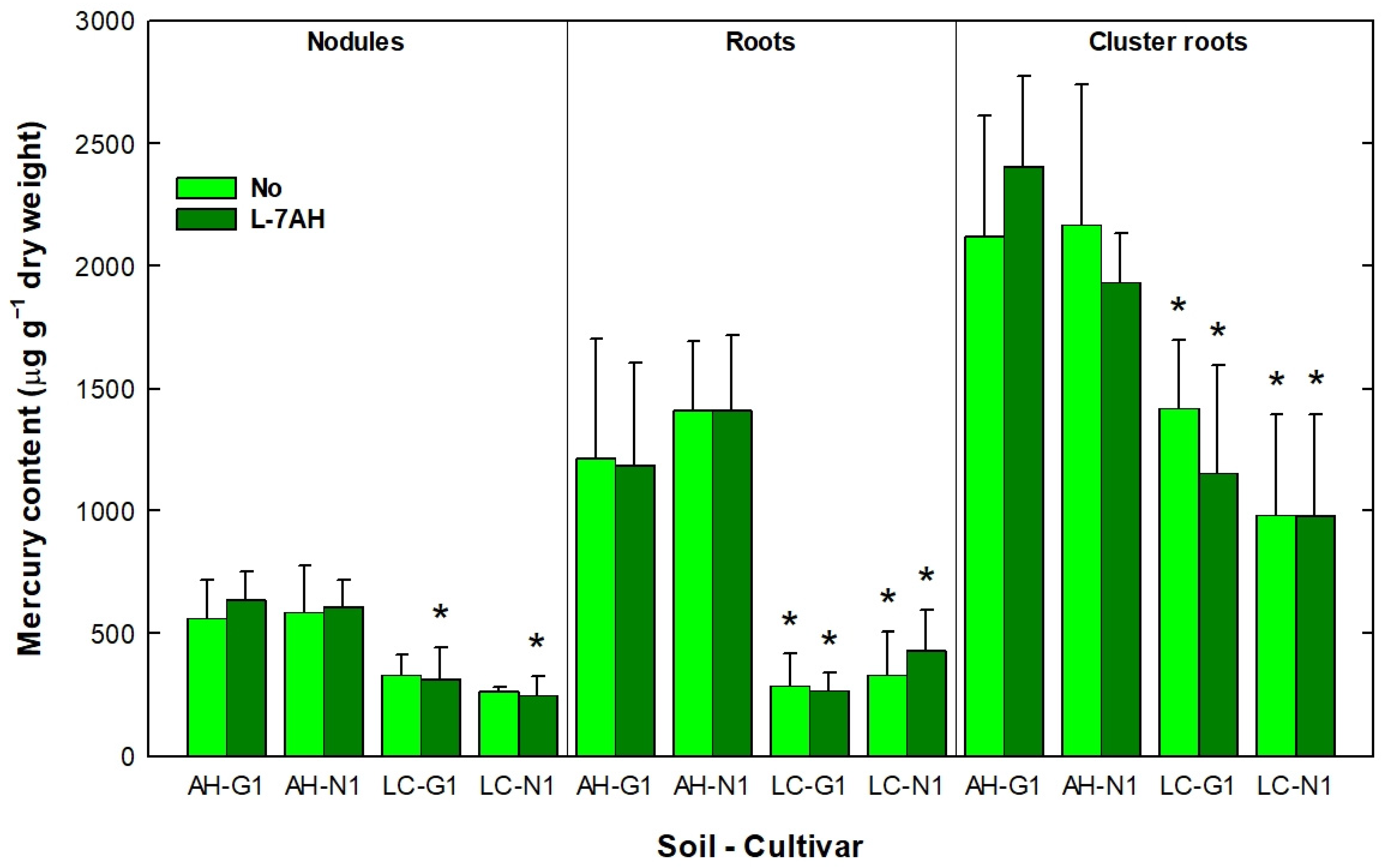
2.8. Mercury Content
2.9. Statistical Analyses
3. Results
3.1. White Lupin Plants Grew Normally in Hg-Contaminated Soils and Were Efficient in Nitrogen Fixation
3.2. Cluster Root Occurrence Was High in Plants Grown in Hg-Contaminated AH and LC Soils
3.3. White Lupin Plants Grown in Hg-Contaminated Soils Accumulated Elevated Amounts of Hg in Nodules, Roots and Cluster Roots
3.4. White Lupin Plants Grown Hydroponically under Cluster Root Promoting Conditions Accumulate High Amounts of Hg in Nodules, Roots and Cluster Roots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brevik, E.C.; Steffan, J.J.; Rodrigo-Comino, J.; Neubert, D.; Burgess, L.C.; Cerdà, A. Connecting the public with soil to improve human health. Environ. J. Soil Sci. 2019, 70, 898–910. [Google Scholar] [CrossRef]
- Amundson, R.; Berhe, A.A.; Hopmans, J.W.; Olson, C.; Sztein, A.E.; Sparks, D.L. Soil and human security in the 21st century. Science 2015, 348, 1261071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.-Q. Heavy metals and pesticides toxicity in agricultural soil and plants: Ecological risks and human health implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef]
- Gao, Y.; Jia, J.; Xi, B.; Tan, W. Divergent response of heavy metal bioavailability in soil rhizosphere to agricultural land use change from paddy fields to various drylands. Environ. Sci. Process. Impacts 2021, 23, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Boening, D.W. Ecological effects, transport, and fate of mercury: A general review. Chemosphere 2000, 40, 1335–1351. [Google Scholar] [CrossRef]
- Molina, J.A.; Oyarzun, R.; Esbrí, J.M.; Higueras, P. Mercury accumulation in soils and plants in the Almadén Mining District, Spain: One of the most contaminated sites on Earth. Environ. Geochem. Health 2006, 28, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Patra, M.; Sharma, A. Mercury toxicity in plants. Bot. Rev. 2000, 66, 379–422. [Google Scholar] [CrossRef]
- Desauziers, V.; Castre, N.; Cloirec, P.L. Sorption of methylmercury by clays and mineral oxides. Environ. Technol. 1997, 18, 1009–1018. [Google Scholar] [CrossRef]
- Millán, R.; Gamarra, R.; Schmid, T.; Sierra, M.J.; Quejido, A.J.; Sánchez, D.M.; Cardona, A.I.; Fernández, M.; Vera, R. Mercury content in vegetation and soils of the Almadén Mining Area (Spain). Sci. Total Environ. 2006, 368, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Kleja, D.B.; Biester, H.; Lagerkvist, A.; Kumpiene, J. Influence of particle size distribution, organic carbon, pH and chlorides on washing of mercury contaminated soil. Chemosphere 2014, 109, 99–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higueras, P.; Oyarzun, R.; Biester, H.; Lillo, J.; Lorenzo, S. A first insight into mercury distribution and speciation in soils from the Almadén Mining District, Spain. J. Geochem. Explor. 2003, 80, 95–104. [Google Scholar] [CrossRef]
- Higueras, P.; Oyarzun, R.; Lillo, J.; Sánchez-Hernández, J.C.; Molina, J.A.; Esbrí, J.M.; Lorenzo, S. The Almadén District (Spain): Anatomy of one of the world’s largest Hg-contaminated sites. Sci. Total Environ. 2006, 356, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Zornoza, P.; Millán, R.; Sierra, M.J.; Seco, A.; Esteban, E. Efficiency of white lupin in the removal of mercury from contaminated soils: Soil and hydroponic experiments. J. Environ. Sci. 2010, 22, 421–427. [Google Scholar] [CrossRef]
- de la Peña, T.C.; Pueyo, J.J. Legumes in the reclamation of marginal soils, from cultivar and inoculant selection to transgenic approaches. Agron. Sustain. Dev. 2012, 32, 65–91. [Google Scholar] [CrossRef] [Green Version]
- Frérot, H.; Lefèbvre, C.; Gruber, W.; Collin, C.; Dos Santos, A.; Escarré, J. Specific interactions between local metallicolous plants improve the phytostabilization of mine soils. Plant. Soil 2006, 282, 53–65. [Google Scholar] [CrossRef]
- Fernández-Pascual, M.; Pueyo, J.J.; de Felipe, M.R.; Golvano, M.P.; Lucas, M.M. Singular features of the Bradyrhizobium-Lupinus symbiosis. Dyn. Soil Dyn. Plant 2007, 1, 1–16. [Google Scholar]
- Jansen, P.C.M. Lupinus albus L. Record from PROTA4U; Brink, M., Belay, G., Eds.; PROTA (Plant Resources of Tropical Africa/Ressources végétales de l’Afrique tropicale): Wageningen, The Netherlands, 2006. [Google Scholar]
- Lucas, M.M.; Stoddard, F.L.; Annicchiarico, P.; Frías, J.; Martínez-Villaluenga, C.; Sussmann, D.; Duranti, M.; Seger, A.; Zander, P.M.; Pueyo, J.J. The future of lupin as a protein crop in Europe. Front. Plant Sci. 2015, 6, 705. [Google Scholar] [CrossRef] [PubMed]
- De Ron, A.M.; Sparvoli, F.; Pueyo, J.J.; Bazile, D. Protein crops: Food and feed for the future. Front. Plant Sci. 2017, 8, 6–9. [Google Scholar] [CrossRef] [Green Version]
- Msaddak, A.; Rey, L.; Imperial, J.; Palacios, J.M.; Mars, M.; Pueyo, J.J. Phylogenetic analyses of rhizobia isolated from nodules of Lupinus angustifolius in Northern Tunisia reveal Devosia sp. as a new microsymbiont of lupin species. Agronomy 2021, 11, 1510. [Google Scholar] [CrossRef]
- de la Peña, T.C.; Fedorova, E.; Pueyo, J.J.; Lucas, M.M. The symbiosome: Legume and rhizobia co-evolution toward a nitrogen-fixing organelle? Front. Plant Sci. 2018, 8, 2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Sama, A.; Lucas, M.M.; de Felipe, M.R.; Pueyo, J.J. An unusual infection mechanism and nodule morphogenesis in white lupin (Lupinus albus). New Phytol. 2004, 163, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Purnell, H.M. Studies of the family Proteaceae i. Anatomy and morphology of the roots of some Victorian species. Aust. J. Bot. 1960, 8, 38–50. [Google Scholar] [CrossRef]
- Lamont, B. Mechanisms for enhancing nutrient uptake in plants, with particular reference to Mediterranean South Africa and Western Australia. Bot. Rev. 1982, 48, 597–689. [Google Scholar] [CrossRef]
- Marschner, H.; Römheld, V.; Horst, W.J.; Martin, P. Root-induced changes in the rhizosphere: Importance for the mineral nutrition of plants. Z. Pflanz. Und Bodenkd 1986, 149, 441–456. [Google Scholar] [CrossRef]
- Neumann, G.; Martinoia, E. Cluster roots—An underground adaptation for survival in extreme environments. Trends Plant. Sci. 2002, 7, 162–167. [Google Scholar] [CrossRef]
- Dinkelaker, B.; Hengeler, C.; Marschner, H. Distribution and function of proteoid roots and other root clusters. Bot. Acta 1995, 108, 183–200. [Google Scholar] [CrossRef]
- Arahou, M.; Diem, H.G. Iron deficiency induces cluster (proteoid) root formation in Casuarina glauca. Plant Soil 1997, 196, 71–79. [Google Scholar] [CrossRef]
- Gerke, J.; Römer, W.; Jungk, A. The excretion of citric and malic acid by proteoid roots of Lupinus albus L.; Effects on soil solution concentrations of phosphate, iron, and aluminum in the proteoid rhizosphere in samples of an oxisol and a luvisol. Z. Pflanz. Und Bodenkd 1994, 157, 289–294. [Google Scholar] [CrossRef]
- Lamont, B. Structure, ecology and physiology of root clusters—A review. Plant Soil 2003, 248, 1–19. [Google Scholar] [CrossRef]
- Gilbert, G.A.; Knight, J.D.; Vance, C.P.; Allan, D.L. Acid phosphatase activity in phosphorus-deficient white lupin roots. Plant Cell Environ. 1999, 22, 801–810. [Google Scholar] [CrossRef]
- Liu, J.; Uhde-Stone, C.; Li, A.; Vance, C.; Allan, D. A phosphate transporter with enhanced expression in proteoid roots of white lupin (Lupinus albus L.). Plant Soil 2001, 237, 257–266. [Google Scholar] [CrossRef]
- Pueyo, J.J.; Quiñones, M.A.; de la Peña, T.C.; Fedorova, E.E.; Lucas, M.M. Nitrogen and phosphorus interplay in lupin root nodules and cluster roots. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Mondal, N.K.; Das, C.; Datta, J.K. Effect of Mercury on Seedling Growth, Nodulation and Ultrastructural Deformation of Vigna Radiata (L) Wilczek. Environ. Monit. Assess. 2015, 187, 241. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, M.A.; Ruiz-Díez, B.; Fajardo, S.; López-Berdonces, M.A.; Higueras, P.L.; Fernández-Pascual, M. Lupinus albus plants acquire mercury tolerance when inoculated with an Hg-resistant Bradyrhizobium strain. Plant Physiol. Biochem. 2013, 73, 168–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Díez, B.; Quiñones, M.A.; Fajardo, S.; Morcillo, C.; Fernández-Pascual, M. Possible reasons for tolerance to mercury of Lupinus albus cv. G1 inoculated with Hg-resistant and sensitive Bradyrhizobium canariense strains. Symbiosis 2015, 67, 91–102. [Google Scholar] [CrossRef]
- de Lorenzo, C.A.; Fernández-Pascual, M.M.; de Felipe, M.R. Protective enzymes against active oxygen species during nitrate-induced senescence of Lupinus albus nodules. J. Plant Physiol. 1994, 144, 633–640. [Google Scholar] [CrossRef]
- Fernández-Pascual, M.; de Lorenzo, C.; de Felipe, M.R.; Rajalakshmi, S.; Gordon, A.J.; Thomas, B.J.; Minchin, F.R. Possible reasons for relative salt stress tolerance in nodules of white lupin cv. Multolupa. J. Exp. Bot. 1996, 47, 1709–1716. [Google Scholar] [CrossRef] [Green Version]
- Pastor, J.; Hernández, A.J.; Prieto, N.; Fernández-Pascual, M. Accumulating behaviour of Lupinus albus L. growing in a normal and a decalcified calcic luvisol polluted with Zn. J. Plant Physiol. 2003, 160, 1457–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zornoza, P.; Vázquez, S.; Esteban, E.; Fernández-Pascual, M.; Carpena, R. Cadmium-stress in nodulated white lupin: Strategies to avoid toxicity. Plant Physiol. Biochem. 2002, 40, 1003–1009. [Google Scholar] [CrossRef]
- Sánchez-Pardo, B.; Fernández-Pascual, M.; Zornoza, P. Copper microlocalisation, ultrastructural alterations and antioxidant responses in the nodules of white lupin and soybean plants grown under conditions of copper excess. Environ. Exp. Bot. 2012, 84, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, C.; Black, C.R. One step forward, two steps back: The evolution of phytoremediation into commercial technologies. Biosci. Horiz. 2014, 7, 1–15. [Google Scholar] [CrossRef]
- Yang, X.; Feng, Y.; He, Z.; Stoffella, P.J. Molecular mechanisms of heavy metal hyperaccumulation and phytoremediation. J. Trace Elem. Med. Biol. 2005, 18, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, A.; Jeevanantham, S.; Narayanan, V.A.; Kumar, P.S.; Yaashikaa, P.R.; Muthu, C.M.M. Rhizoremediation—A promising tool for the removal of soil contaminants: A review. J. Environ. Chem. Eng. 2020, 8, 103543. [Google Scholar] [CrossRef]
- Dary, M.; Chamber-Pérez, M.A.; Palomares, A.J.; Pajuelo, E. “In situ” phytostabilisation of heavy metal polluted soils using Lupinus luteus inoculated with metal resistant plant-growth promoting rhizobacteria. J. Hazard. Mater. 2010, 177, 323–330. [Google Scholar] [CrossRef]
- Corso, M.; de la Torre, V.S.G. Biomolecular approaches to understanding metal tolerance and hyperaccumulation in plants. Metallomics 2020, 12, 840–859. [Google Scholar] [CrossRef] [PubMed]
- Krämer, U. Metal hyperaccumulation in plants. Annu. Rev. Plant. Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef]
- Reeves, R.D.; van der Ent, A.; Baker, A.J.M. Global distribution and ecology of hyperaccumulator plants. In Agromining: Farming for Metals, Mineral. Resource Reviews; Springer: Cham, Switerland, 2018; pp. 75–92. [Google Scholar]
- Romero-Hernández, J.A.; Amaya-Chávez, A.; Balderas-Hernández, P.; Roa-Morales, G.; González-Rivas, N.; Balderas-Plata, M.Á. Tolerance and hyperaccumulation of a mixture of heavy metals (Cu, Pb, Hg, and Zn) by four aquatic macrophytes. Int. J. Phytoremediation 2017, 19, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant. Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Suman, J.; Uhlik, O.; Viktorova, J.; Macek, T. Phytoextraction of heavy metals: A promising tool for clean-up of polluted environment? Front. Plant Sci. 2018, 9, 1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiodar, E.D.; Văcar, C.L.; Podar, D. Phytoremediation and microorganisms-assisted phytoremediation of mercury-contaminated soils: Challenges and perspectives. Int. J. Environ. Res. Public Health 2021, 18, 2435. [Google Scholar] [CrossRef] [PubMed]
- de la Torre, V.S.G.; de la Peña, T.C.; Lucas, M.M.; Pueyo, J.J. Rapid screening of Medicago truncatula germplasm for mercury tolerance at the seedling stage. Environ. Exp. Bot. 2013, 91, 90–96. [Google Scholar] [CrossRef] [Green Version]
- de la Torre, V.S.G.; de la Peña, T.C.; Pueyo, J.J.; Lucas, M.M. Cadmium-tolerant and -sensitive cultivars identified by screening of Medicago truncatula germplasm display contrasting responses to cadmium stress. Front. Plant Sci. 2021, 12, 293. [Google Scholar]
- Ruiz-Díez, B.; Quiñones, M.A.; Fajardo, S.; López, M.A.; Higueras, P.; Fernández-Pascual, M. Mercury-resistant rhizobial bacteria isolated from nodules of leguminous plants growing in high Hg-contaminated soils. Appl. Microbiol. Biotechnol. 2012, 96, 543–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Díez, B.; Fajardo, S.; Puertas-Mejía, M.A.; de Felipe, M.R.; Fernández-Pascual, M. Stress tolerance, genetic analysis and symbiotic properties of root-nodulating bacteria isolated from Mediterranean leguminous shrubs in Central Spain. Arch. Microbiol. 2009, 191, 35–46. [Google Scholar] [CrossRef]
- Quiñones, M.A.; Fajardo, S.; Rodríguez-Caviedes, L.; Hortigüela, C.; Contreras, M.; Lucas, M.M.; Pueyo, J.J.; Fernández-Pascual, M. Mercury tolerance of white lupin growing in soils of the Almadén Mining District. In Current and Future Trends in Mercury Research; Chartridge Books Oxford: Oxford, UK, 2018; pp. 31–39. [Google Scholar]
- Millán, R.; Lominchar, M.A.; López-Tejedor, I.; Rodríguez-Alonso, J.; Schmid, T.; Sierra, M.J. Behavior of mercury in the Valdeazogues riverbank soil and transfer to Nerium oleander L. J. Geochem. Explor. 2012, 123, 136–142. [Google Scholar] [CrossRef]
- Brigatti, M.F.; Colonna, S.; Malferrari, D.; Medici, L.; Poppi, L. Mercury adsorption by montmorillonite and vermiculite: A combined XRD, TG-MS, and EXAFS study. Appl. Clay Sci. 2005, 28, 1–8. [Google Scholar] [CrossRef]
- Millán, R.; Esteban, E.; Zornoza, P.; Sierra, M.J. Could an abandoned mercury mine area be cropped? Environ. Res. 2013, 125, 150–159. [Google Scholar]
- Martínez-Coronado, A.; Oyarzun, R.; Esbrí, J.M.; Llanos, W.; Higueras, P. Sampling high to extremely high Hg concentrations at the Cerco de Almadenejos, Almadén Mining District (Spain): The old metallurgical precinct (1794 to 1861AD) and surrounding areas. J. Geochem. Explor. 2011, 109, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Dessureault-Rompré, J.; Nowack, B.; Schulin, R.; Luster, J. Spatial and temporal variation in organic acid anion exudation and nutrient anion uptake in the rhizosphere of Lupinus albus L. Plant Soil 2007, 301, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Dessureault-Rompré, J.; Nowack, B.; Schulin, R.; Tercier-Waeber, M.L.; Luster, J. Metal solubility and speciation in the rhizosphere of Lupinus albus cluster roots. Environ. Sci. Technol. 2008, 42, 7146–7151. [Google Scholar] [CrossRef]
- Yang, Y.; Ratté, D.; Smets, B.F.; Pignatello, J.J.; Grasso, D. Mobilization of soil organic matter by complexing agents and implications for polycyclic aromatic hydrocarbon desorption. Chemosphere 2001, 43, 1013–1021. [Google Scholar] [CrossRef]
- He, M.; Tian, L.; Braaten, H.F.V.; Wu, Q.; Luo, J.; Cai, L.M.; Meng, J.H.; Lin, Y. Mercury–organic matter interactions in soils and sediments: Angel or devil? Bull. Environ. Contam. Toxicol. 2019, 102, 621–627. [Google Scholar] [CrossRef]
- Haitzer, M.; Aiken, G.R.; Ryan, J.N. Binding of mercury (II) to aquatic humic substances: Influence of pH and source of humic substances. Environ. Sci. Technol. 2003, 37, 2436–2441. [Google Scholar] [CrossRef]
- Ravichandran, M.; Aiken, G.R.; Reddy, M.M.; Ryan, J.N. Enhanced dissolution of cinnabar (mercuric sulfide) by dissolved organic matter isolated from the Florida Everglades. Environ. Sci. Technol. 1998, 32, 3305–3311. [Google Scholar] [CrossRef]
- Angelini, J.; Taurian, T.; Morgante, C.; Ibáñez, F.; Castro, S.; Fabra, A. Peanut nodulation kinetics in response to low pH. Plant Physiol. Biochem. 2005, 43, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Denton, M.D.; Hill, C.R.; Bellotti, W.D.; Coventry, D.R. Nodulation of Medicago truncatula and Medicago polymorpha in two pastures of contrasting soil pH and rhizobial populations. Appl. Soil Ecol. 2007, 35, 441–448. [Google Scholar] [CrossRef]
- Ferreira, T.C.; Aguilar, J.V.; Souza, L.A.; Justino, G.C.; Aguiar, L.F.; Camargos, L.S. pH effects on nodulation and biological nitrogen fixation in Calopogonium mucunoides. Rev. Bras. Bot. 2016, 39, 1015–1020. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, L.; Rincón, J.; Asencio, I.; Rodríguez-Castellanos, L. Capability of selected crop plants for shoot mercury accumulation from polluted soils: Phytoremediation perspectives. Int. J. Phytoremediation 2007, 9, 1–13. [Google Scholar] [CrossRef]
- Sierra, M.J.; Millán, R.; Esteban, E. Potential use of Solanum melongena in agricultural areas with high mercury background concentrations. Food Chem. Toxicol. 2008, 46, 2143–2149. [Google Scholar] [CrossRef] [PubMed]
- Sierra, M.J.; Millán, R.; Esteban, E. Mercury uptake and distribution in Lavandula stoechas plants grown in soil from Almadén Mining District (Spain). Food Chem. Toxicol. 2009, 47, 2761–2767. [Google Scholar] [CrossRef] [PubMed]
- Sierra, M.J.; Millán, R.; Cardona, A.I.; Schmid, T. Potential cultivation of Hordeum vulgare L. in soils with high mercury background concentrations. Int. J. Phytoremediat. 2011, 13, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Meng, B.; Feng, X.; Qiu, G.; Anderson, C.W.N.; Wang, J.; Zhao, L. Localization and speciation of mercury in brown rice with implications for Pan-Asian public health. Environ. Sci. Technol. 2014, 48, 7974–7981. [Google Scholar] [CrossRef]
- Higueras, P.L.; Amorós, J.; Esbrí, J.M.; de los Reyes, C.P.; López-Berdonces, M.A.; García-Navarro, F.J. Mercury transfer from soil to olive trees. A comparison of three different contaminated sites. Environ. Sci. Pollut. Res. 2016, 23, 6055–6061. [Google Scholar] [CrossRef]
- Chen, J.; Shiyab, S.; Han, F.X.; Monts, D.L.; Waggoner, C.A.; Yang, Z.; Su, Y. Bioaccumulation and physiological effects of mercury in Pteris vittata and Nephrolepis exaltata. Ecotoxicology 2009, 18, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Lombi, E.; Zhao, F.J.; McGrath, S.P.; Young, S.D.; Sacchi, G.A. Physiological evidence for a high-affinity cadmium transporter highly expressed in a Thlaspi caerulescens ecotype. New Phytol. 2001, 149, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regier, N.; Larras, F.; Bravo, A.G.; Ungureanu, V.G.; Amouroux, D.; Cosio, C. Mercury bioaccumulation in the aquatic plant Elodea nuttallii in the field and in microcosm: Accumulation in shoots from the water might involve copper transporters. Chemosphere 2013, 90, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Peel, G.J.; Lei, Z.; Aziz, N.; Dai, X.; He, J.; Watson, B.; Zhao, P.X.; Sumner, L.W.; Dixon, R.A. Transcript and proteomic analysis of developing white lupin (Lupinus albus L.) roots. BMC Plant Biol. 2009, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Boyd, E.S.; Barkay, T. The mercury resistance operon: From an origin in a geothermal environment to an efficient detoxification machine. Front. Microbiol. 2012, 3, 349. [Google Scholar] [CrossRef] [Green Version]
- Uraguchi, S.; Sone, Y.; Yoshikawa, A.; Tanabe, M.; Sato, H.; Otsuka, Y.; Nakamura, R.; Takanezawa, Y.; Kiyono, M. SCARECROW promoter-driven expression of a bacterial mercury transporter MerC in root endodermal cells enhances mercury accumulation in Arabidopsis shoots. Planta 2019, 250, 667–674. [Google Scholar] [CrossRef]
- Arregui, G.; Hipólito, P.; Pallol, B.; Lara-Dampier, V.; García-Rodríguez, D.; Varela, H.P.; Tavakoli Zaniani, P.; Balomenos, D.; Paape, T.; de la Peña, T.C.; et al. Mercury-tolerant Ensifer medicae strains display high mercuric reductase activity and a protective effect on nitrogen fixation in Medicago truncatula nodules under mercury stress. Front. Plant Sci. 2021, 11, 560768. [Google Scholar] [CrossRef]
- Sarret, G.; Saumitou-Laprade, P.; Bert, V.; Proux, O.; Hazemann, J.L.; Traverse, A.; Marcus, M.A.; Manceau, A. Forms of zinc accumulated in the hyperaccumulator Arabidopsis halleri. Plant Physiol. 2002, 130, 1815–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salt, D.E.; Prince, R.C.; Baker, A.J.M.; Raskin, I.; Pickering, I.J. Zinc ligands in the metal hyperaccumulator Thlaspi caerulescens as determined using X-Ray absorption spectroscopy. Environ. Sci. Technol. 1999, 33, 713–717. [Google Scholar] [CrossRef]
- Zhang, H.; Feng, X.; Zhu, J.; Sapkota, A.; Meng, B.; Yao, H.; Qin, H.; Larssen, T. Selenium in soil inhibits mercury uptake and translocation in rice (Oryza sativa L.). Environ. Sci. Technol. 2012, 46, 10040–10046. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.A.T.; Dinh, Q.T.; Zhou, F.; Zhai, H.; Xue, M.; Du, Z.; Bañuelos, G.S.; Liang, D. Mechanisms underlying mercury detoxification in soil–plant systems after selenium application: A review. Environ. Sci. Pollut. Res. 2021, 28, 46852–46876. [Google Scholar] [CrossRef] [PubMed]
- Esteban, E.; Moreno, E.; Peñalosa, J.; Cabrero, J.I.; Millán, R.; Zornoza, P. Short and long-term uptake of Hg in white lupin plants: Kinetics and stress indicators. Environ. Exp. Bot. 2008, 62, 316–322. [Google Scholar] [CrossRef]
- Sobrino-Plata, J.; Herrero, J.; Carrasco-Gil, S.; Pérez-Sanz, A.; Lobo, C.; Escobar, C.; Millán, R.; Hernández, L.E. Specific stress responses to cadmium, arsenic and mercury appear in the metallophyte Silene vulgaris when grown hydroponically. RSC Adv. 2013, 3, 4736–4744. [Google Scholar] [CrossRef]
- Carrasco-Gil, S.; Álvarez-Fernández, A.; Sobrino-Plata, J.; Millán, R.; Carpena-Ruiz, R.O.; Leduc, D.L.; Andrews, J.C.; Abadía, J.; Hernández, L.E. Complexation of Hg with phytochelatins is important for plant Hg tolerance. Plant Cell Environ. 2011, 34, 778–791. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Gil, S.; Siebner, H.; LeDuc, D.; Webb, M.; Millán, R.; Andrews, J.C.; Hernández, L.E. Mercury localization and speciation in plants grown hydroponically or in a natural environment. Environ. Sci. Technol. 2013, 47, 3082–3090. [Google Scholar] [CrossRef]
- Duranti, M.; Scarafoni, A.; Di Cataldo, A.; Sessa, F. Interaction of metal ions with lupin seed conglutin γ. Phytochemistry 2001, 56, 529–533. [Google Scholar] [CrossRef]
- Bolan, N.S.; Park, J.H.; Robinson, B.; Naidu, R.; Huh, K.Y. Phytostabilization. A green approach to contaminant containment. Adv. Agron. 2011, 112, 145–204. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil | pH | Organic Matter (%) | C (%) | N (%) | Bioavailable Fe (μg g−1) | P (μg g−1) | Total Hg (μg g−1) | Bioavailable Hg (μg g−1) |
|---|---|---|---|---|---|---|---|---|
| AH | 6.87 | 8.59 | 3.33 | 0.26 | 106.3 | 11.2 | 21,845 | 2.69 |
| LC | 5.09 | 9.85 | 3.82 | 0.35 | 158.9 | 3.6 | 2622 | 0.82 |
| Tissue | Almadenejos | Las Cuevas | ||
|---|---|---|---|---|
| G1 | N1 | G1 | N1 | |
| Nodules | 443.5 ± 98.1 | 441.6 ± 107.8 | 787.2 ± 215.7 * | 624.1 ± 117.6 * |
| Roots | 889.2 ± 313 | 1046 ± 202.1 | 675.7 ± 241.4 | 927.0 ± 406.5 |
| Cluster roots | 1676.8 ± 318.4 | 1519 ± 308.3 | 3139.3 ± 900.9 * | 2390.9 ± 932.9 * |
| Tissue | 0.05 mM P | 0.5 mM P |
|---|---|---|
| Nodules | 131.9 ± 33 | 142 ± 48.5 |
| Roots | 137.4 ± 45.7 | 146.8 ± 29.7 |
| Cluster roots | 309.4 ± 66.1 | 345 ± 40.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiñones, M.A.; Fajardo, S.; Fernández-Pascual, M.; Lucas, M.M.; Pueyo, J.J. Nodulated White Lupin Plants Growing in Contaminated Soils Accumulate Unusually High Mercury Concentrations in Their Nodules, Roots and Especially Cluster Roots. Horticulturae 2021, 7, 302. https://doi.org/10.3390/horticulturae7090302
Quiñones MA, Fajardo S, Fernández-Pascual M, Lucas MM, Pueyo JJ. Nodulated White Lupin Plants Growing in Contaminated Soils Accumulate Unusually High Mercury Concentrations in Their Nodules, Roots and Especially Cluster Roots. Horticulturae. 2021; 7(9):302. https://doi.org/10.3390/horticulturae7090302
Chicago/Turabian StyleQuiñones, Miguel A., Susana Fajardo, Mercedes Fernández-Pascual, M. Mercedes Lucas, and José J. Pueyo. 2021. "Nodulated White Lupin Plants Growing in Contaminated Soils Accumulate Unusually High Mercury Concentrations in Their Nodules, Roots and Especially Cluster Roots" Horticulturae 7, no. 9: 302. https://doi.org/10.3390/horticulturae7090302