Competitive Inhibitory Effect of Calcium Polypeptides on Cd Enrichment of Brassia campestris L.


Abstract
:1. Introduction
2. Methods and Materials
2.1. Experimental Materials
2.1.1. Test Plants
2.1.2. Experiment-Soil-Sample Preparation
2.1.3. Calcium Polypeptide Preparation
2.2. Experimental Design
2.3. Sample Processing and Index Measurement
2.3.1. Fresh-Weight Determination
2.3.2. Chlorophyll a Determination
2.3.3. Determination of Calcium, Nitrogen, Phosphorus, Potassium, and Cadmium Contents in Brassia campestris L.
2.3.4. Determination of Available Cadmium in Soil
2.4. Data-Processing Method
3. Results
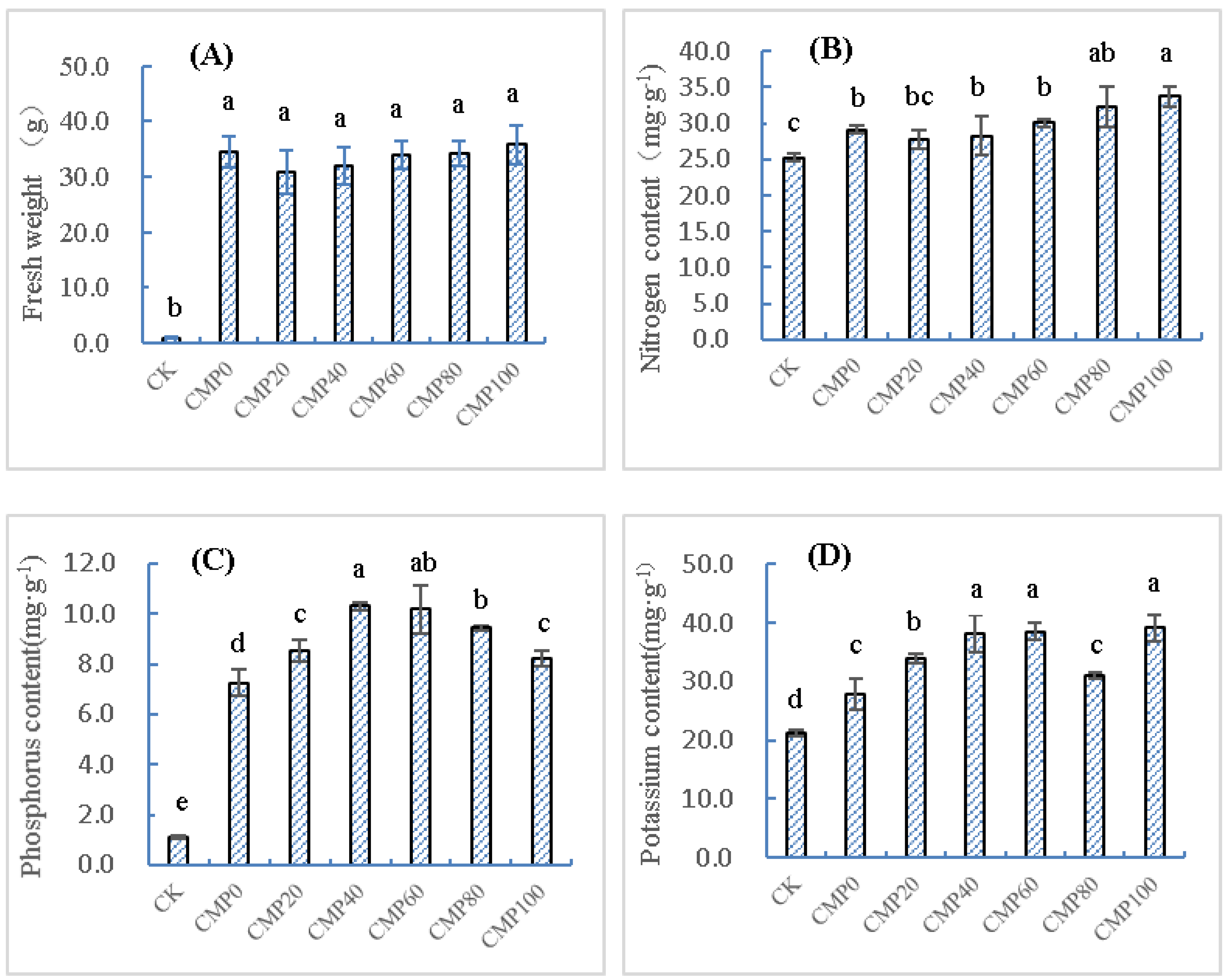
3.1. Effects of Calcium Polypeptide on Growth Characteristics of Brassia campestris L.
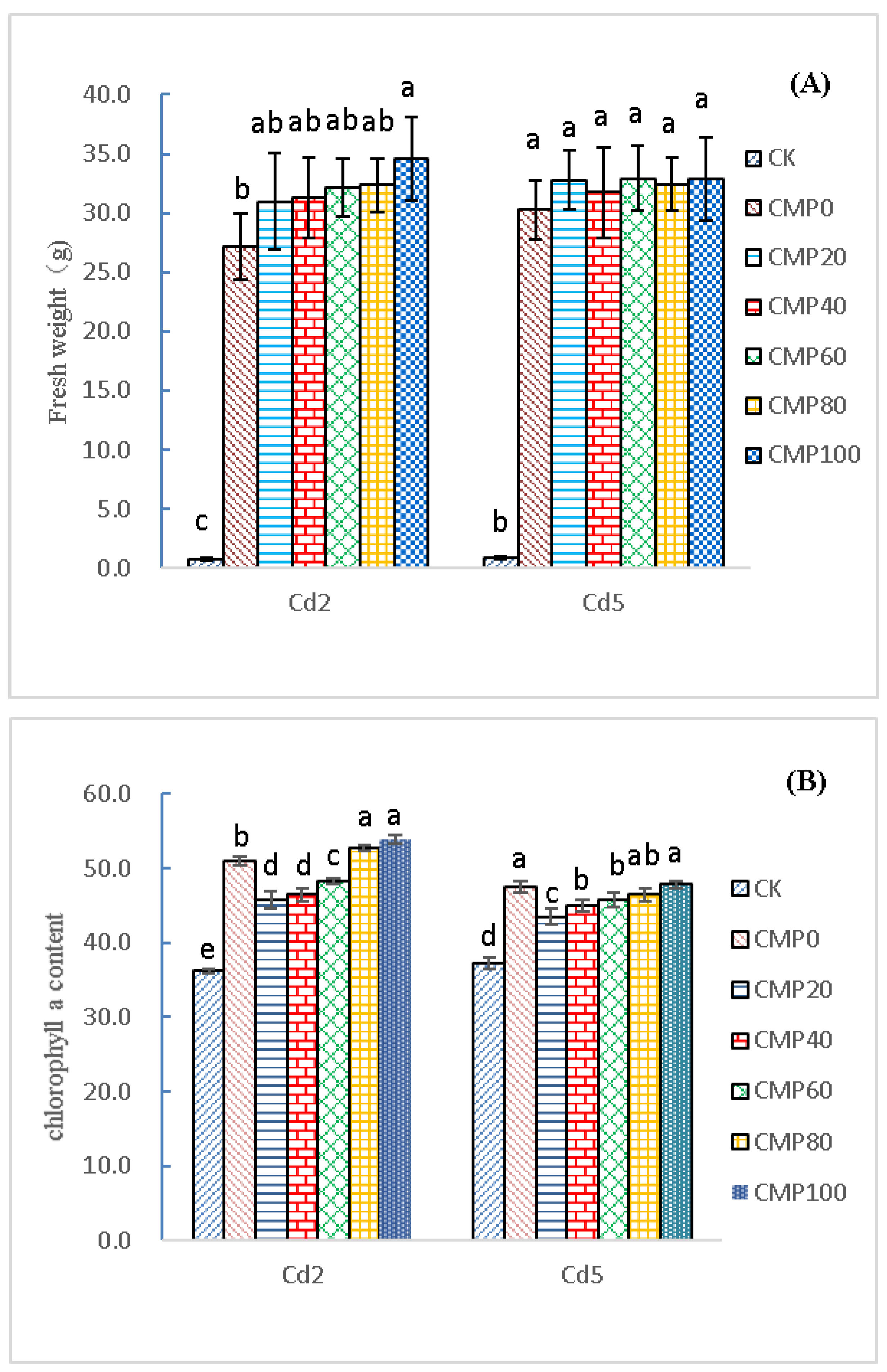
3.2. Control of Brassia campestris L. Growth in Cadmium-Contaminated Soil by Different Calcium Polypeptide Application Amounts
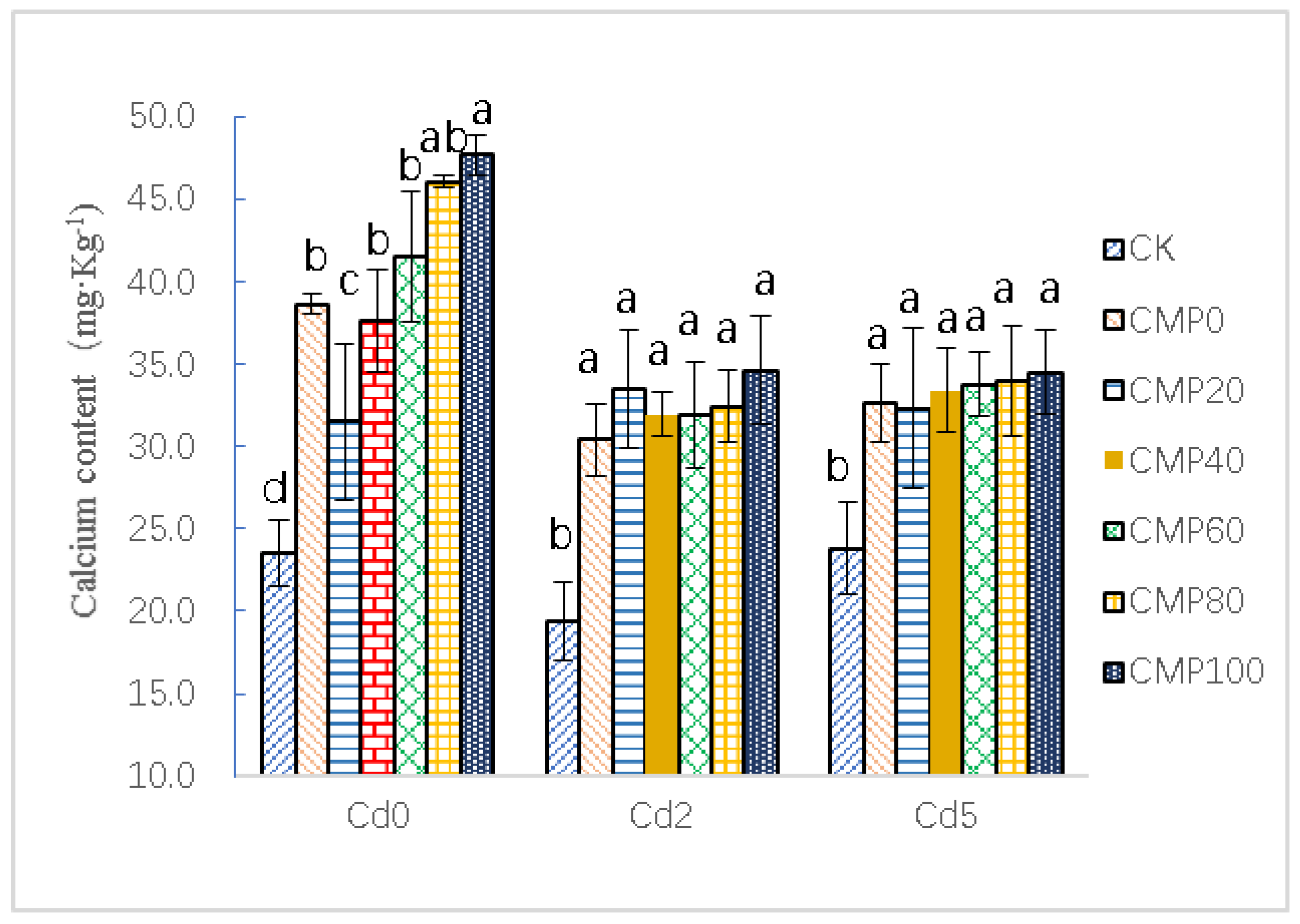
3.3. Effects of Calcium Polypeptide on Ca Content of Brassia campestris L.
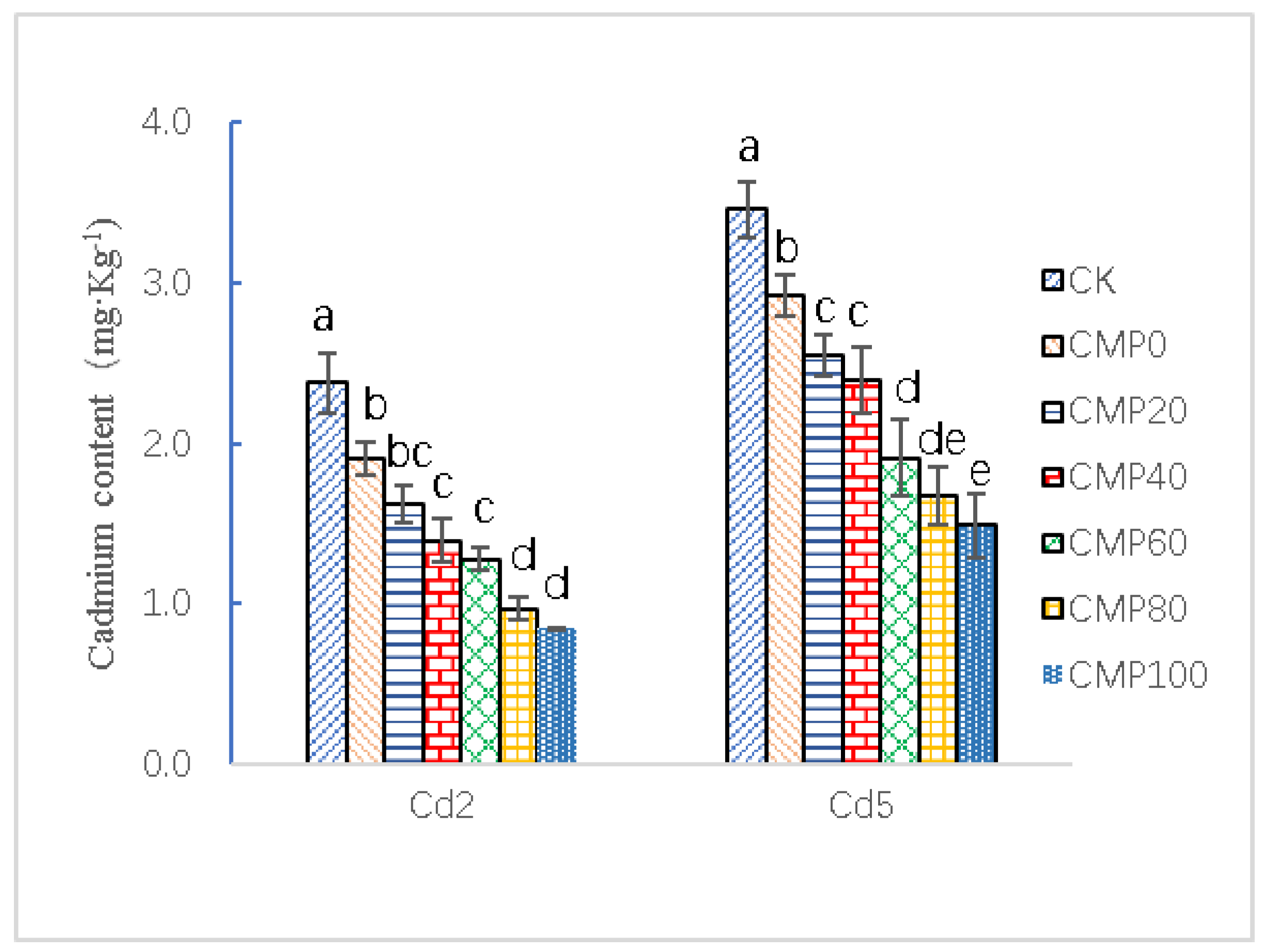
3.4. Effects of Calcium Polypeptide on Cd2+ Content of Brassia campestris L.
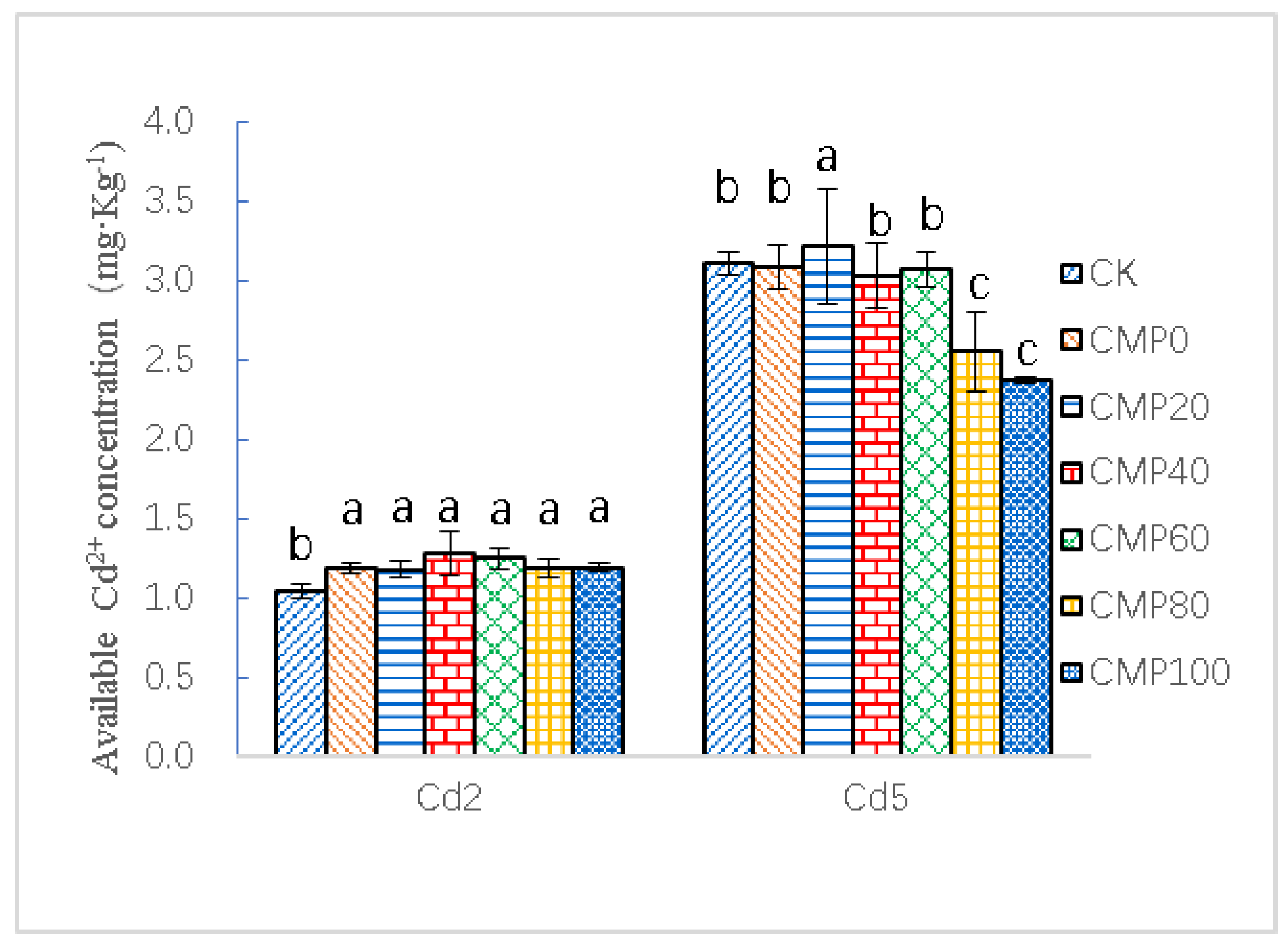
3.5. Effects of Calcium Polypeptide on Available Cadmium Content in Cadmium Contaminated Soil
4. Discussion
4.1. Correlation Analysis
4.2. Effects of Different Calcium Application Rates on Ca2+ Accumulation in Brassia campestris L. under Cadmium Stress
4.3. Control of Calcium Polypeptide on Cadmium Existence in Soil
4.4. Competitive Inhibitory Effect of Calcium polyPeptide on Cadmium Absorption in Brassia campestris L.
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cheng, K.; Tian, H.Z.; Zhao, D.; Lu, L.; Wang, Y.; Chen, J.; Liu, X.G.; Jia, W.X.; Huang, Z. Atmospheric emission inventory of cadmium from anthropogenic sources. Int. J. Env. Sci. Technol. 2014, 11, 605–616. [Google Scholar] [CrossRef]
- Fagerberg, B.; Barregard, L.; Sallsten, G.; Forsgard, N.; Östling, G.; Persson, M.; Borné, Y.; Engström, G.; Hedblad, B. Cadmium exposure and atherosclerotic carotid plaques-results from the Malmö diet and Cancer study. Env. Res. 2015, 136, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Sakan, S.M.; Dević, G.J.; Relić, D.J.; Anđelković, I.B.; Sakan, N.M.; Đorđević, D.S. Environmental Assessment of Heavy Metal Pollution in Freshwater Sediment, Serbia. CLEAN Soil Air Water 2015, 43, 838–845. [Google Scholar] [CrossRef]
- Yang, X.; Duan, J.; Wang, L.; Li, W.; Guan, J.; Beecham, S.; Mulcahy, D. Heavy metal pollution and health risk assessment in the Wei River in China. Env. Monit. Assess. 2015, 187, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Ecology and Environment, PRC. National Soil Pollution Status Survey Bulletin. Available online: http://www.mee.gov.cn/gkml/sthjbgw/qt/201404/t20140417_270670.htm (accessed on 17 April 2014).
- Huang, D.L.; Wang, R.Z.; Liu, Y.G.; Zeng, G.M.; Lai, C.; Xu, P.; Lu, B.A.; Xu, J.J.; Wang, C.; Huang, C. Application of molecularly imprinted polymers in wastewater treatment: A review. Env. Sci. Pollut. Res. 2015, 22, 963–977. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Shu, Y. Cadmium Transporters in the Kidney and Cadmium-Induced Nephrotoxicity. Int. J. Mol. Sci. 2015, 16, 1484–1494. [Google Scholar] [CrossRef]
- Brodziak-Dopierała, B.; Kwapuliński, J.; Sobczyk, K.; Wiechuła, D. Analysis of the Content of Cadmium and Zinc in Parts of the Human Hip Joint. Biol. Trace Elem. Res. 2014, 163, 73–80. [Google Scholar] [CrossRef]
- Degl’Innocenti, E.; Castagna, A.; Ranieri, A.; Guidi, L. Combined effects of cadmium and ozone on photosynthesis of Lycopersicon esculentum. Photosynthetica 2014, 52, 179–185. [Google Scholar] [CrossRef]
- Li, S.; Yang, W.; Yang, T.; Chen, Y.; Ni, W. Effects of Cadmium Stress on Leaf Chlorophyll Fluorescence and Photosynthesis ofElsholtzia argyi-A Cadmium Accumulating Plant. Int. J. Phytoremediation 2014, 17, 85–92. [Google Scholar] [CrossRef]
- Deng, G.; Li, M.; Li, H.; Yin, L.; Li, W. Exposure to cadmium causes declines in growth and photosynthesis in the endangered aquatic fern (Ceratopteris pteridoides). Aquat. Bot. 2014, 112, 23–32. [Google Scholar] [CrossRef]
- Song, W.Y.; Zhang, Z.B.; Shao, H.B.; Guo, X.L.; Cao, H.X.; Zhao, H.B.; Fu, Z.Y.; Hu, X.J. Relationship between calcium decoding elements and plant abiotic-tress resistance. Int. J. Biol. Sci. 2008, 4, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.A.; Popova, L.P. Functions and toxicity of cadmium in plants: Recent advances and future prospects. Turk. J. Bot. 2013, 37, 1–13. [Google Scholar] [CrossRef]
- Li, X.; Zhou, Q.; Sun, X.; Ren, W. Effects of cadmium on uptake and translocation of nutrient elements in different welsh onion (Allium fistulosum L.) cultivars. Food Chem. 2016, 194, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.C.; Monteiro, C.; Moutinho-Pereira, J.; Correia, C.; Gonçalves, B.; Santos, C. Cadmium toxicity affects photosynthesis and plant growth at different levels. Acta Physiol. Plant. 2012, 35, 1281–1289. [Google Scholar] [CrossRef]
- Hasan, S.A.; Fariduddin, Q.; Ali, B.; Hayat, S.; Ahmad, A. Cadmium: Toxicity and tolerance in plants. J. Env. Biol. 2009, 32, 165–174. [Google Scholar]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Env. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Huang, D.L.; Zeng, G.M.; Feng, C.L.; Hu, S.; Jiang, X.F.; Tang, L.; Su, F.F.; Zhang, Y.; Zeng, W.; Liu, H.L. Degradation of lead-contaminated lignocellulosic waste by Phanerochaete chrysosporium and the reduction of lead toxicity. Env. Sci. Technol. 2008, 42, 4946–4951. [Google Scholar] [CrossRef]
- Islam, M.S.; Ahmed, M.K.; Raknuzzaman, M.; Habibullah-Al-Mamun, M.; Islam, M.K. Heavy metal pollution in surface water and sediment: A preliminary assessment of an urban river in a developing country. Ecol. Indic. 2015, 48, 282–291. [Google Scholar] [CrossRef]
- Huang, D.; Xue, W.; Zeng, G.; Wan, J.; Chen, G.; Huang, C.; Xu, P. Immobilization of Cd in river sediments by sodium alginate modified nanoscale zero-valent iron: Impact on enzyme activities and microbial community diversity. Water Res. 2016, 106, 15–25. [Google Scholar] [CrossRef]
- Zhang, S.R.; Guo, A.H.; Han, Y.; Ge, S.H.; Wang, J.; Xu, Z.H.G.; Diao, S. Preparation of Peptide Calcium Chelate with Waste Protein. Leather Chem. 2019, 36, 9–13. (In Chinese) [Google Scholar]
- Huang, D.L.; Gong, X.M.; Liu, Y.G.; Zeng, G.M.; Lai, C.; Bashir, H.; Zhou, L.; Wang, D.F.; Xu, P.; Cheng, M.; et al. Effects of calcium at toxic concentrations of cadmium in plants. Planta 2017, 245, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Dayod, M.; Tyerman, S.D.; Leigh, R.A.; Gilliham, M. Calcium storage in plants and the implications for calcium biofortification. Protoplasma 2010, 247, 215–231. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.S.; Huang, Y.X.; Huang, C.Y. Effects of water-soluble organic fertilizer on yield of rice grain and cadmium absorption in rice grain and straw. J. Agro Env. Sci. 2017, 36, 826–831. [Google Scholar] [CrossRef]
- Hayakawa, N.; Tomioka, R.; Takenaka, C. Effects of calcium on cadmium uptake and transport in the tree species Gamblea innovans. Soil Sci. Plant Nutr. 2011, 57, 691–695. [Google Scholar] [CrossRef]
- Marchetti, C. Role of calcium channels in heavy metal toxicity. ISRN Toxicol. 2013, 2013, 184360. [Google Scholar] [CrossRef]
- Li, P.; Zhao, C.; Zhang, Y.; Wang, X.; Wang, X.; Wang, J.; Wang, F.; Bi, Y. Calcium alleviates cadmium-induced inhibition on root growth by maintaining auxin homeostasis in Arabidopsis seedlings. Protoplasma 2015, 253, 185–200. [Google Scholar] [CrossRef]
- Bhargava, A.; Carmona, F.F.; Bhargava, M.; Srivastava, S. Approaches for enhanced phytoextraction of heavy metals. J. Env. Manag. 2012, 105, 103–120. [Google Scholar] [CrossRef]
- Ahmad, P.; Sarwat, M.; Bhat, N.A.; Wani, M.R.; Kazi, A.G.; Tran, L.S.P. Alleviation of Cadmium Toxicity in Brassica juncea L. (Czern. & Coss.) by Calcium Application Involves Various Physiological and Biochemical Strategies. PLoS ONE 2015, 10, e0114571. [Google Scholar] [CrossRef]
- Farzadfar, S.; Zarinkamar, F.; Modarres-Sanavy, S.A.M.; Hojati, M. Exogenously applied calcium alleviates cadmium toxicity in Matricaria chamomilla L. plants. Env. Sci. Pollut. Res. 2012, 20, 1413–1422. [Google Scholar] [CrossRef]
- Singh, A.; Prasad, S.M. Remediation of heavy metal contaminated ecosystem: An overview on technology advancement. Int. J. Env. Sci. Technol. 2014, 12, 353–366. [Google Scholar] [CrossRef]
- Huang, F.; Wen, X.H.; Cai, Y.X.; Cai, K.Z. Silicon-Mediated Enhancement of Heavy Metal Tolerance in Rice at Different Growth Stages. Int. J. Env. Res. Public Health 2018, 15, 2193. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wang, Z.H.; Cai, Y.X.; Chen, S.H.; Tian, J.H.; Cai, K.Z. Heavy metal bioaccumulation and cation release by growing Bacillus cereus RC-1 under culture conditions. Ecotoxicol. Env. Saf. 2018, 157, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Fischer, G.; Winiwarter, W.; Ermolieva, T. Integrated modeling framework for assessment and mitigation of nitrogen pollution from agriculture: Concept and case study for China. Agric. Ecosyst. Env. 2010, 136, 116–124. [Google Scholar] [CrossRef]
- Song, Z.G.; Xu, M.G.; Li, J.M. Effects of calcium on cadmium bioavail-ability in lateritic red soil and related mechanisms. Chin. J. Appl. Ecol. 2009, 20, 1705–1710. (In Chinese) [Google Scholar]
- Velthof, G.L.; Oudendag, D.; Witzke, H.P. Integrated assessment of nitrogen emissions from agriculture in EU-27 using Miterra-Europe. J. Env. Qual. 2009, 38, 402–417. [Google Scholar] [CrossRef]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.L.; Han, W.X.; Zhang, W.F.; Christie, P.; Goulding, K.W.T.; Vitousek, P.M.; Zhang, F.S. Significant acidification in major Chinese croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.D.; Wang, X.G.; Yang, L.F.; Li, S.Y. Calcium polypeptide in situ passivating agent for treating heavy metal contaminated soil and its preparation method and application. Patent ZL201410128133.7, 19 August 2015. [Google Scholar]
- Wang, Q.L.; Zou, J.H.; Liu, D.H.; Yue, J.Y. Effects of exogenous calcium (Ca) on tolerance of Allium cepa var.agrogarum L.to cadmium (Cd) stress. Acta Ecol. Sin. 2014, 34, 1165–1177. [Google Scholar] [CrossRef]
- Shi, G.R.; Cai, Q.S.; Liu, Q.Q.; Wu, L. Salicylic acid-mediated alleviation of cadmium toxicity in hemp plants in relation to cadmium uptake, photosynthesis, and antioxidant enzymes. Acta Physiol. Plant. 2009, 31, 969–977. [Google Scholar] [CrossRef]
- Ghasemi, S.; Khoshgoftarmanesh, A.H.; Afyuni, M.; Hadadzadeh, H. The effectiveness of foliar applications of synthesized zinc-amino acid chelates in comparison with zinc sulfate to increase yield and grain nutritional quality of wheat. Eur. J. Agron. 2013, 45, 68–74. [Google Scholar] [CrossRef]
- Wu, H.; Liu, Z.; Zhao, Y.; Zeng, M. Enzymatic preparation and characterization of iron-chelating peptides from anchovy (Engraulis japonicus) muscle protein. Food Res. Int. 2012, 48, 435–441. [Google Scholar] [CrossRef]
- Fytili, D.; Zabaniotou, A. Utilization of sewage sludge in EU application of old and new methods–A review. Renew. Sustain. Energy Rev. 2008, 12, 116–140. [Google Scholar] [CrossRef]
- Xue, W.J. Research on the Effect of Peptide on Soil Microecology and Plant Growth. Master’s Thesis, Hubei University, Wuhan, China, 2014. [Google Scholar]
- Gao, Q. The Effect of Peptide on the Forms of Cd on Soil Contaminated by Cadmium. Master’s Thesis, Hubei University, Wuhan, China, 2016. [Google Scholar]
- Cot, J.; Aramon, C.; Baucells, M. Waste processing in the tannery: Production of gelatin, reconstituted collagen and glue from chrome-tanned leather splits and trimmings subjected to a modified detanning process. J. Soc. Leather Technol. Chem. 2014, 70, 69–76. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Almeida, M.F.; Pinho, S.C. Finished leather waste chromium acid extraction and anaerobic biodegradation of the products. Waste Manag. 2010, 30, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Malek, A.; Hachemi, M.; Didier, V. New approach of depollution of solid chromium leather waste by the use of organic chelates: Economical and environmental impacts. J. Hazard. Mater. 2009, 170, 156–162. [Google Scholar] [CrossRef]
- Dang, X.; Shan, Z.; Chen, H. Biodegradable films based on gelatin extracted from chrome leather scrap. Int. J. Biol. Macromol. 2017, 107, 1023–1029. [Google Scholar] [CrossRef]
- Ghasemi, S.; Khoshgoftarmanesh, A.H.; Afyuni, M. Iron (II)-amino acid chelates alleviate salt-stress induced oxidative damages on tomato grown in nutrient solution culture. Sci. Hortic. 2014, 165, 91–98. [Google Scholar] [CrossRef]
- Mohammadi, P.; Khoshgoftarmanesh, A.H. The effectiveness of synthetic zinc(Zn)-amino chelates in supplying Zn and alleviating salt-induced damages on hydroponically grown lettuce. Sci. Hortic. 2014, 172, 117–123. [Google Scholar] [CrossRef]
- Choi, D.W.; Lee, J.H.; Chun, H.H. Isolation of a calciumbinding peptide from bovine serum protein hydrolysates. Food Sci. Biotechnol. 2012, 21, 1663–1667. [Google Scholar] [CrossRef]
- Song, Z.G.; Xu, M.G.; Ding, Y.Z.; Sun, N.; Tang, S.R. Effect of Coexistence Cations (Ca, Zn, K) on Cadmium Bioavailability in Lateritic Red Soils. J. Agro Env. Sci. 2009, 28, 485–489. (In Chinese) [Google Scholar]
- Javed, M.T.; Akram, M.S.; Tanwir, K.; Javed Chaudhary, H.; Ali, Q.; Stoltz, E.; Lindberg, S. Cadmium spiked soil modulates root organic acids exudation and ionic contents of two differentially Cd tolerant maize ( Zea mays L.) cultivars. Ecotoxicol. Env. Saf. 2017, 141, 216–225. [Google Scholar] [CrossRef]
- Nedjimi, B.; Daoud, Y. Cadmium accumulation in Atriplex halimus subsp. schweinfurthii and its influence on growth, proline, root hydraulic conductivity and nutrient uptake. Flora–Morphol. Distrib. Funct. Ecol. Plants 2009, 204, 316–324. [Google Scholar] [CrossRef]
- El-Ashry, Z.M.; Mohamed, F.I. Protective effects of some antioxidant metals against chromosomal damage induced by cadmium in Vicia faba plants. Int. J. Agric. Res. 2012, 7, 376–387. [Google Scholar] [CrossRef] [Green Version]
- Kurtyka, R.; Małkowski, E.; Kita, A.; Karcz, W. Effect of Calcium and Cadmium onGrowth and Accumulation of Cadmium, Calcium, Potassium and Sodium in Maize Seedlings. Pol. J. Env. Stud. 2008, 17, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, P.; Abdel Latef, A.A.; Abd_Allah, E.F.; Hashem, A.; Sarwat, M.; Anjum, N.A.; Gucel, S. Calcium and Potassium Supplementation Enhanced Growth, Osmolyte Secondary Metabolite Production, and Enzymatic Antioxidant Machinery in Cadmium-Exposed Chickpea (Cicer arietinum L.). Front. Plant Sci. 2016, 7, 513. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Sakran, A.M.; Basalah, M.O.; Ali, H.M. Effect of Calcium and Potassium on Antioxidant System of Vicia faba L. Under Cadmium Stress. Int. J. Mol. Sci. 2012, 13, 6604–6619. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.P.; Lan, P.L.; Li, S.Y.; Zhang, Y.C.; Deng, X.W.; Gu, X.F.; Liao, X.R.; Gong, J.B. Effects of different nitrogen fertilizers and fertilization patterns on yield and quality of Brassica chinensis. Ecol. Environ. 2007, 16, 1040–1043. (In Chinese) [Google Scholar]
- Guan, M.Y.; Zhang, H.H.; Pan, W.; Jin, C.W.; Lin, X.Y. Sulfide alleviates cadmium toxicity in Arabidopsis plants by altering the chemical form and the subcellular distribution of cadmium. Sci. Total Environ. 2018, 627, 663–670. [Google Scholar] [CrossRef]
- Rodriguez-Hernandez, M.C.; Bonifas, I.; Alfaro-De la Torre, M.C.; Flores-Flores, J.L.; Bañuelos-Hernández, B.; Patiño-Rodríguez, O. Increased accumulation of cadmium and lead under Ca and Fe deficiency in Typha latifolia: A study of two pore channel (TPC1) gene responses. Environ. Exp. Bot. 2015, 115, 38–48. [Google Scholar] [CrossRef]
- Mleczek, M.; Kozlowska, M.; Kaczmarek, Z.; Chadzinikolau, T.; Golinski, P. Influence of Ca/Mg Ratio on Phytoextraction Properties ofSalix ViminalisI. The Effectiveness of Cd, Cu, Pb, and Zn Bioaccumulation and Plant Growth. Int. J. Phytoremediation 2012, 14, 75–88. [Google Scholar] [CrossRef]
- Suzuki, N. Alleviation by calcium of cadmium-induced root growth inhibition in Arabidopsis seedlings. Plant Biotechnol. 2005, 22, 19–25. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Matter (g·kg−1) | Total Nitrogen (g·kg−1) | Total Phosphorus (g·kg−1) | Total Potassium (g·kg−1) | Alkali-Hydrolyzed Nitrogen (mg·kg−1) | Total Cadmium (mg·kg−1) | pH |
|---|---|---|---|---|---|---|
| 13.50 | 1.437 | 0.735 | 8.53 | 98.03 | 0.002 | 7.1 |
| Indicators | Fresh Weight | Chlorophyll a Content | Calcium Content | Cadmium Content | Available Cd2+ Concentration |
|---|---|---|---|---|---|
| Fresh weight | 1 | 0.772 ** | 0.875 ** | −0.755 ** | 0.539 * |
| Chlorophyll a content | 0.734 ** | 1 | 0.756 ** | −0.803 ** | 0.495 * |
| Calcium content | 0.828 ** | 0.866 ** | 1 | −0.734 ** | 0.594 ** |
| Cadmium content | −0.611 ** | −0.697 ** | −0.673 ** | 1 | −0.530 * |
| Available Cd2+ concentration | −0.129 | −0.324 | −0.174 | 0.442* | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Shu, F.; Yang, S.; Li, Y.; Wang, S. Competitive Inhibitory Effect of Calcium Polypeptides on Cd Enrichment of Brassia campestris L. Int. J. Environ. Res. Public Health 2019, 16, 4472. https://doi.org/10.3390/ijerph16224472
Chen H, Shu F, Yang S, Li Y, Wang S. Competitive Inhibitory Effect of Calcium Polypeptides on Cd Enrichment of Brassia campestris L. International Journal of Environmental Research and Public Health. 2019; 16(22):4472. https://doi.org/10.3390/ijerph16224472
Chicago/Turabian StyleChen, Hongbing, Fangfang Shu, Sheng Yang, Yadong Li, and Shilin Wang. 2019. "Competitive Inhibitory Effect of Calcium Polypeptides on Cd Enrichment of Brassia campestris L." International Journal of Environmental Research and Public Health 16, no. 22: 4472. https://doi.org/10.3390/ijerph16224472