Activity-Based Physical Rehabilitation with Adjuvant Testosterone to Promote Neuromuscular Recovery after Spinal Cord Injury


Abstract
:1. Introduction
2. Neuromuscular Adaptations after SCI
2.1. Ubiquitin-Proteasome Signaling after SCI
2.2. IGF-1 Signaling Pathway after SCI
2.3. PGC-1α and PGC-1β Signaling after SCI
3. Androgenic Regulation of Muscle
3.1. Testosterone Synthesis and Metabolism
3.2. Hypogonadism and Testosterone Replacement after SCI
3.3. Classical Androgen Signaling
3.4. Androgenic Crosstalk with IGF-1 and PI3K/Akt Signaling
3.5. Androgen Crosstalk with the Ubiquitin-Proteasome Pathway
3.6. Androgenic Influence on PGC-1α and PGC-1β Signaling after SCI
4. Androgenic and Estrogenic Regulation of Motoneurons
4.1. Influence of Androgens and Estrogens on Motoneuron Structure after SCI
4.2. Molecular Pathways Underlying Androgen-Mediated Motoneuron Protection
5. Activity-Based Physical Rehabilitation after SCI
5.1. Effects of ABTs on Muscle
5.2. Effects of ABTs on Motoneurons
6. Testosterone Adjuvant to BWSTT
Potential Side-Effects of TRT
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABT | activity-based therapies |
| AR | androgen receptor |
| ARE | androgen response element |
| BDNF | brain-derived neurotrophic factor |
| BWSTT | bodyweight-supported treadmill training |
| CNS | central nervous system |
| CSA | cross-sectional area |
| DHT | dihydrotestosterone |
| E2 | estradiol |
| EPSP | excitatory postsynaptic potential |
| ER | estrogen receptor |
| ERE | estrogen response element |
| fCSA | fiber cross-sectional area |
| FOXO | forkhead box O |
| GPCR | G-protein-coupled receptor |
| GSK3β | glycogen synthase kinase 3β |
| IGF-1 | insulin-like growth factor |
| IGF-1R | insulin-like growth factor receptor |
| IGFBP | insulin-like growth factor binding protein |
| IRS1 | insulin receptor substrate-1 |
| LABC | levator ani/bulbocavernosus |
| MAFbx | muscle atrophy F-box or atrogin-1 |
| MGF | mechano growth factor |
| MHC | myosin heavy chain |
| mTOR | mammalian target of rapamycin |
| MuRF1 | muscle ring finger-1 |
| NT-3 | neurotrophin-3 |
| ORX | orchiectomy |
| PGC-1α | peroxisome proliferator-activated receptor gamma co-activator-1α |
| PGC-1β | peroxisome proliferator-activated receptor gamma co-activator-1β |
| PI3K | phosphatidyl inositol-3 kinase |
| SCI | spinal cord injury |
| SHBG | sex-hormone binding globulin |
| SNB | spinal nucleus of the bulbocavernosus |
| T | testosterone |
| TCF-4 | T-cell factor-4 |
| TE | testosterone enanthate |
| TrkB | tyrosine receptor kinase B |
| TrkC | tyrosine receptor kinase C |
| TRT | testosterone replacement therapy |
References
- Beattie, M.S.; Farooqui, A.A.; Bresnahan, J.C. Review of current evidence for apoptosis after spinal cord injury. J. Neurotrauma 2000, 17, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Hilton, B.J.; Moulson, A.J.; Tetzlaff, W. Neuroprotection and secondary damage following spinal cord injury: Concepts and methods. Neurosci. Lett. 2017, 652, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Kwon, B.K.; Tetzlaff, W.; Grauer, J.N.; Beiner, J.; Vaccaro, A.R. Pathophysiology and pharmacologic treatment of acute spinal cord injury. Spine J. 2004, 4, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Morawietz, C.; Moffat, F. Effects of locomotor training after incomplete spinal cord injury: A systematic review. Arch. Phys. Med. Rehabil. 2013, 94, 2297–2308. [Google Scholar] [CrossRef] [PubMed]
- Battistuzzo, C.R.; Callister, R.J.; Callister, R.; Galea, M.P. A systematic review of exercise training to promote locomotor recovery in animal models of spinal cord injury. J. Neurotrauma 2012, 29, 1600–1613. [Google Scholar] [CrossRef] [PubMed]
- Behrman, A.L.; Ardolino, E.M.; Harkema, S.J. Activity-based therapy: From basic science to clinical application for recovery after spinal cord injury. J. Neurol. Phys. Ther. 2017, 41 (Suppl. S3), S39–S45. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, S.D.; Nash, M.S.; Tefera, E.; Tinsley, E.; Blackman, M.R.; Groah, S. Prevalence and etiology of hypogonadism in young men with chronic spinal cord injury: A cross-sectional analysis from two university-based rehabilitation centers. PM&R 2017, 9, 751–760. [Google Scholar]
- Borst, S.E.; Yarrow, J.F. Injection of testosterone may be safer and more effective than transdermal administration for combating loss of muscle and bone in older men. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E1035–E1042. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.S.; Gray, H.; Schache, A.G.; Hoermann, R.; Lim Joon, D.; Zajac, J.D.; Pandy, M.G.; Grossmann, M. Androgen deprivation causes selective deficits in the biomechanical leg muscle function of men during walking: A prospective case-control study. J. Cachexia Sarcopenia Muscle 2017, 8, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Skinner, J.W.; Otzel, D.M.; Bowser, A.; Nargi, D.; Agarwal, S.; Peterson, M.D.; Zou, B.; Borst, S.E.; Yarrow, J.F. Muscular responses to testosterone replacement vary by administration route: A systematic review and meta-analysis. J. Cachexia Sarcopenia Muscle 2018. [Google Scholar] [CrossRef] [PubMed]
- Snyder, P.J.; Bhasin, S.; Cunningham, G.R.; Matsumoto, A.M.; Stephens-Shields, A.J.; Cauley, J.A.; Gill, T.M.; Barrett-Connor, E.; Swerdloff, R.S.; Wang, C.; et al. Effects of testosterone treatment in older men. N. Engl. J. Med. 2016, 374, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Bauman, W.A.; Cirnigliaro, C.M.; La Fountaine, M.F.; Jensen, A.M.; Wecht, J.M.; Kirshblum, S.C.; Spungen, A.M. A small-scale clinical trial to determine the safety and efficacy of testosterone replacement therapy in hypogonadal men with spinal cord injury. Horm. Metab. Res. 2011, 43, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.D.; Gorgey, A.S.; Wade, R.C.; Khalil, R.E.; Lavis, T.D.; Khan, R.; Adler, R.A. Neuromuscular electrical stimulation and testosterone did not influence heterotopic ossification size after spinal cord injury: A case series. World J. Clin. Cases 2016, 4, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Yarrow, J.F.; Phillips, E.G.; Conover, C.F.; Bassett, T.E.; Chen, C.; Teurlings, T.; Vasconez, A.; Alerte, J.; Prock, H.; Jiron, J.M.; et al. Testosterone plus finasteride prevents bone loss without prostate growth in a rodent spinal cord injury model. J. Neurotrauma 2017, 34, 2972–2981. [Google Scholar] [CrossRef] [PubMed]
- Yarrow, J.F.; Conover, C.F.; Beggs, L.A.; Beck, D.T.; Otzel, D.M.; Balaez, A.; Combs, S.M.; Miller, J.R.; Ye, F.; Aguirre, J.I.; et al. Testosterone dose dependently prevents bone and muscle loss in rodents after spinal cord injury. J. Neurotrauma 2014, 31, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhao, J.; Zhao, W.; Pan, J.; Bauman, W.A.; Cardozo, C.P. Nandrolone normalizes determinants of muscle mass and fiber type after spinal cord injury. J. Neurotrauma 2012, 29, 1663–1675. [Google Scholar] [CrossRef] [PubMed]
- Sengelaub, D.R.; Han, Q.; Liu, N.K.; Maczuga, M.A.; Szalavari, V.; Valencia, S.A.; Xu, X.M. Protective effects of estradiol and dihydrotestosterone following spinal cord injury. J. Neurotrauma 2018, 35, 825–841. [Google Scholar] [CrossRef] [PubMed]
- Byers, J.S.; Huguenard, A.L.; Kuruppu, D.; Liu, N.K.; Xu, X.M.; Sengelaub, D.R. Neuroprotective effects of testosterone on motoneuron and muscle morphology following spinal cord injury. J. Comp. Neurol. 2012, 520, 2683–2696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ung, R.V.; Rouleau, P.; Guertin, P.A. Effects of co-administration of clenbuterol and testosterone propionate on skeletal muscle in paraplegic mice. J. Neurotrauma 2010, 27, 1129–1142. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.M.; Vandenborne, K.; Huang, H.F.; Ottenweller, J.E.; Dudley, G.A. Effects of testosterone replacement therapy on skeletal muscle after spinal cord injury. Spinal Cord 2003, 41, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Sandri, M.; Murgia, M. Activity-dependent signaling pathways controlling muscle diversity and plasticity. Physiology 2007, 22, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.K.; Bakels, R.; Klein, C.S.; Zijdewind, I. Human spinal cord injury: Motor unit properties and behaviour. Acta Physiol. 2014, 210, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.S.; Hager-Ross, C.K.; Thomas, C.K. Fatigue properties of human thenar motor units paralysed by chronic spinal cord injury. J. Physiol. 2006, 573, 161–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraman, A.; Gregory, C.M.; Bowden, M.; Stevens, J.E.; Shah, P.; Behrman, A.L.; Vandenborne, K. Lower extremity skeletal muscle function in persons with incomplete spinal cord injury. Spinal Cord 2006, 44, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Hager-Ross, C.K.; Klein, C.S.; Thomas, C.K. Twitch and tetanic properties of human thenar motor units paralyzed by chronic spinal cord injury. J. Neurophysiol. 2006, 96, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Barbeau, H.; Nadeau, S.; Garneau, C. Physical determinants, emerging concepts, and training approaches in gait of individuals with spinal cord injury. J. Neurotrauma 2006, 23, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Bose, P.; Parmer, R.; Reier, P.J.; Thompson, F.J. Morphological changes of the soleus motoneuron pool in chronic midthoracic contused rats. Exp. Neurol. 2005, 191, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Gazula, V.R.; Roberts, M.; Luzzio, C.; Jawad, A.F.; Kalb, R.G. Effects of limb exercise after spinal cord injury on motor neuron dendrite structure. J. Comp. Neurol. 2004, 476, 130–145. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, N.K.; Zhang, Y.P.; Deng, L.; Lu, Q.B.; Shields, C.B.; Walker, M.J.; Li, J.; Xu, X.M. Treadmill training induced lumbar motoneuron dendritic plasticity and behavior recovery in adult rats after a thoracic contusive spinal cord injury. Exp. Neurol. 2015, 271, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Biering-Sorensen, B.; Kristensen, I.B.; Kjaer, M.; Biering-Sorensen, F. Muscle after spinal cord injury. Muscle Nerve 2009, 40, 499–519. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.D.; Craven, B.C.; Thabane, L.; Laing, A.C.; Frank-Wilson, A.W.; Kontulainen, S.A.; Papaioannou, A.; Adachi, J.D.; Giangregorio, L.M. Lower-extremity muscle atrophy and fat infiltration after chronic spinal cord injury. J. Musculoskelet. Neuronal Interact. 2015, 15, 32–41. [Google Scholar] [PubMed]
- Gorgey, A.S.; Dudley, G.A. Skeletal muscle atrophy and increased intramuscular fat after incomplete spinal cord injury. Spinal Cord 2007, 45, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.J.; Apple, D.F., Jr.; Hillegass, E.A.; Dudley, G.A. Influence of complete spinal cord injury on skeletal muscle cross-sectional area within the first 6 months of injury. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 80, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.K.; Stevens, J.E.; Gregory, C.M.; Pathare, N.C.; Jayaraman, A.; Bickel, S.C.; Bowden, M.; Behrman, A.L.; Walter, G.A.; Dudley, G.A.; et al. Lower-extremity muscle cross-sectional area after incomplete spinal cord injury. Arch. Phys. Med. Rehabil. 2006, 87, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.J.; Apple, D.F., Jr.; Staron, R.S.; Campos, G.E.; Dudley, G.A. Influence of complete spinal cord injury on skeletal muscle within 6 mo of injury. J. Appl. Physiol. 1999, 86, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.M.; Vandenborne, K.; Castro, M.J.; Dudley, G.A. Human and rat skeletal muscle adaptations to spinal cord injury. Can. J. Appl. Physiol. 2003, 28, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Phillips, E.G.; Beggs, L.A.; Ye, F.; Conover, C.F.; Beck, D.T.; Otzel, D.M.; Ghosh, P.; Bassit, A.C.F.; Borst, S.E.; Yarrow, J.F. Effects of pharmacologic sclerostin inhibition or testosterone administration on soleus muscle atrophy in rodents after spinal cord injury. PLoS ONE 2018, 13, e0194440. [Google Scholar] [CrossRef] [PubMed]
- Talmadge, R.J.; Roy, R.R.; Edgerton, V.R. Persistence of hybrid fibers in rat soleus after spinal cord transection. Anat. Rec. 1999, 255, 188–201. [Google Scholar] [CrossRef] [Green Version]
- Talmadge, R.J.; Roy, R.R.; Edgerton, V.R. Prominence of myosin heavy chain hybrid fibers in soleus muscle of spinal cord-transected rats. J. Appl. Physiol. 1995, 78, 1256–1265. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.K.; Zaidner, E.Y.; Calancie, B.; Broton, J.G.; Bigland-Ritchie, B.R. Muscle weakness, paralysis, and atrophy after human cervical spinal cord injury. Exp. Neurol. 1997, 148, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Gaviria, M.; Ohanna, F. Variability of the fatigue response of paralyzed skeletal muscle in relation to the time after spinal cord injury: Mechanical and electrophysiological characteristics. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 80, 145–153. [Google Scholar] [CrossRef] [PubMed]
- DiPiro, N.D.; Holthaus, K.D.; Morgan, P.J.; Embry, A.E.; Perry, L.A.; Bowden, M.G.; Gregory, C.M. Lower extremity strength is correlated with walking function after incomplete SCI. Top. Spinal Cord Inj. Rehabil. 2015, 21, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.M.; Bowden, M.G.; Jayaraman, A.; Shah, P.; Behrman, A.; Kautz, S.A.; Vandenborne, K. Resistance training and locomotor recovery after incomplete spinal cord injury: A case series. Spinal Cord 2007, 45, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Yarar-Fisher, C.; Bickel, C.S.; Windham, S.T.; McLain, A.B.; Bamman, M.M. Skeletal muscle signaling associated with impaired glucose tolerance in spinal cord-injured men and the effects of contractile activity. J. Appl. Physiol. 2013, 115, 756–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarar-Fisher, C.; Bickel, C.S.; Kelly, N.A.; Windham, S.T.; McLain, A.B.; Bamman, M.M. Mechanosensitivity may be enhanced in skeletal muscles of spinal cord-injured versus able-bodied men. Muscle Nerve 2014, 50, 599–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panisset, M.G.; Galea, M.P.; El-Ansary, D. Does early exercise attenuate muscle atrophy or bone loss after spinal cord injury? Spinal Cord 2016, 54, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Malavaki, C.J.; Sakkas, G.K.; Mitrou, G.I.; Kalyva, A.; Stefanidis, I.; Myburgh, K.H.; Karatzaferi, C. Skeletal muscle atrophy: Disease-induced mechanisms may mask disuse atrophy. J. Muscle Res. Cell Motil. 2015, 36, 405–421. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K.; et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 2001, 294, 1704–1708. [Google Scholar] [CrossRef] [PubMed]
- Baligand, C.; Chen, Y.W.; Ye, F.; Pandey, S.N.; Lai, S.H.; Liu, M.; Vandenborne, K. Transcriptional pathways associated with skeletal muscle changes after spinal cord injury and treadmill locomotor training. Biomed. Res. Int. 2015, 2015, 387090. [Google Scholar] [CrossRef] [PubMed]
- Haddad, F.; Roy, R.R.; Zhong, H.; Edgerton, V.R.; Baldwin, K.M. Atrophy responses to muscle inactivity. Ii. Molecular markers of protein deficits. J. Appl. Physiol. 2003, 95, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Roy, R.R.; Kim, J.A.; Zhong, H.; Haddad, F.; Baldwin, K.M.; Edgerton, V.R. Gene expression during inactivity-induced muscle atrophy: Effects of brief bouts of a forceful contraction countermeasure. J. Appl. Physiol. 2008, 105, 1246–1254. [Google Scholar] [CrossRef] [PubMed]
- Urso, M.L.; Chen, Y.W.; Scrimgeour, A.G.; Lee, P.C.; Lee, K.F.; Clarkson, P.M. Alterations in mrna expression and protein products following spinal cord injury in humans. J. Physiol. 2007, 579, 877–892. [Google Scholar] [CrossRef] [PubMed]
- Zeman, R.J.; Zhao, J.; Zhang, Y.; Zhao, W.; Wen, X.; Wu, Y.; Pan, J.; Bauman, W.A.; Cardozo, C. Differential skeletal muscle gene expression after upper or lower motor neuron transection. Pflugers Arch. 2009, 458, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Sacheck, J.M.; Hyatt, J.P.; Raffaello, A.; Jagoe, R.T.; Roy, R.R.; Edgerton, V.R.; Lecker, S.H.; Goldberg, A.L. Rapid disuse and denervation atrophy involve transcriptional changes similar to those of muscle wasting during systemic diseases. FASEB J. 2007, 21, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Thakore, N.P.; Samantaray, S.; Park, S.; Nozaki, K.; Smith, J.A.; Cox, A.; Krause, J.; Banik, N.L. Molecular changes in sub-lesional muscle following acute phase of spinal cord injury. Neurochem. Res. 2016, 41, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Baligand, C.; Keener, J.E.; Vohra, R.; Lim, W.; Ruhella, A.; Bose, P.; Daniels, M.; Walter, G.A.; Thompson, F.; et al. Hindlimb muscle morphology and function in a new atrophy model combining spinal cord injury and cast immobilization. J. Neurotrauma 2013, 30, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; Glynn, E.L.; Lujan, H.L.; Dicarlo, S.E.; Rasmussen, B.B. Gene and protein expression associated with protein synthesis and breakdown in paraplegic skeletal muscle. Muscle Nerve 2008, 37, 505–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundell, L.S.; Savikj, M.; Kostovski, E.; Iversen, P.O.; Zierath, J.R.; Krook, A.; Chibalin, A.V.; Widegren, U. Protein translation, proteolysis and autophagy in human skeletal muscle atrophy after spinal cord injury. Acta Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yarar-Fisher, C.; Bickel, C.S.; Kelly, N.A.; Stec, M.J.; Windham, S.T.; McLain, A.B.; Oster, R.A.; Bamman, M.M. Heightened tweak-nf-kappab signaling and inflammation-associated fibrosis in paralyzed muscles of men with chronic spinal cord injury. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E754–761. [Google Scholar] [CrossRef] [PubMed]
- Leger, B.; Senese, R.; Al-Khodairy, A.W.; Deriaz, O.; Gobelet, C.; Giacobino, J.P.; Russell, A.P. Atrogin-1, murf1, and foxo, as well as phosphorylated gsk-3beta and 4e-bp1 are reduced in skeletal muscle of chronic spinal cord-injured patients. Muscle Nerve 2009, 40, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhu, X. The molecular mechanisms of calpains action on skeletal muscle atrophy. Physiol. Res. 2016, 65, 547–560. [Google Scholar] [PubMed]
- Bikle, D.D.; Tahimic, C.; Chang, W.; Wang, Y.; Philippou, A.; Barton, E.R. Role of IGF-I signaling in muscle bone interactions. Bone 2015, 80, 79–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Stevens-Lapsley, J.E.; Jayaraman, A.; Ye, F.; Conover, C.; Walter, G.A.; Bose, P.; Thompson, F.J.; Borst, S.E.; Vandenborne, K. Impact of treadmill locomotor training on skeletal muscle IGF1 and myogenic regulatory factors in spinal cord injured rats. Eur. J. Appl. Physiol. 2010, 109, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Pete, G.; Fuller, C.R.; Oldham, J.M.; Smith, D.R.; D’Ercole, A.J.; Kahn, C.R.; Lund, P.K. Postnatal growth responses to insulin-like growth factor i in insulin receptor substrate-1-deficient mice. Endocrinology 1999, 140, 5478–5487. [Google Scholar] [CrossRef] [PubMed]
- Tsitouras, P.D.; Zhong, Y.G.; Spungen, A.M.; Bauman, W.A. Serum testosterone and growth hormone/insulin-like growth factor-i in adults with spinal cord injury. Horm. Metab. Res. 1995, 27, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Bauman, W.A.; Spungen, A.M.; Flanagan, S.; Zhong, Y.G.; Alexander, L.R.; Tsitouras, P.D. Blunted growth hormone response to intravenous arginine in subjects with a spinal cord injury. Horm. Metab. Res. 1994, 26, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, H.C.; Glynn, E.L.; Lujan, H.L.; Fry, C.S.; DiCarlo, S.E.; Rasmussen, B.B. Chronic paraplegia-induced muscle atrophy downregulates the mtor/s6k1 signaling pathway. J. Appl. Physiol. 2008, 104, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Gorgey, A.S.; Gater, D.R. Insulin growth factors may explain relationship between spasticity and skeletal muscle size in men with spinal cord injury. J. Rehabil. Res. Dev. 2012, 49, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Ji, L.L. Role of PGC-1alpha signaling in skeletal muscle health and disease. Ann. N. Y. Acad. Sci. 2012, 1271, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Gali Ramamoorthy, T.; Laverny, G.; Schlagowski, A.I.; Zoll, J.; Messaddeq, N.; Bornert, J.M.; Panza, S.; Ferry, A.; Geny, B.; Metzger, D. The transcriptional coregulator pgc-1beta controls mitochondrial function and anti-oxidant defence in skeletal muscles. Nat. Commun. 2015, 6, 10210. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Wu, H.; Tarr, P.T.; Zhang, C.Y.; Wu, Z.; Boss, O.; Michael, L.F.; Puigserver, P.; Isotani, E.; Olson, E.N.; et al. Transcriptional co-activator pgc-1 alpha drives the formation of slow-twitch muscle fibres. Nature 2002, 418, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Goodman, C.A.; Hornberger, T.A.; Ji, L.L. PGC-1alpha overexpression by in vivo transfection attenuates mitochondrial deterioration of skeletal muscle caused by immobilization. FASEB J. 2015, 29, 4092–4106. [Google Scholar] [CrossRef] [PubMed]
- Higashino, K.; Matsuura, T.; Suganuma, K.; Yukata, K.; Nishisho, T.; Yasui, N. Early changes in muscle atrophy and muscle fiber type conversion after spinal cord transection and peripheral nerve transection in rats. J. Neuroeng. Rehabil. 2013, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Collier, L.; Qin, W.; Creasey, G.; Bauman, W.A.; Jarvis, J.; Cardozo, C. Electrical stimulation modulates wnt signaling and regulates genes for the motor endplate and calcium binding in muscle of rats with spinal cord transection. BMC Neurosci. 2013, 14, 81. [Google Scholar] [CrossRef] [PubMed]
- Kramer, D.K.; Ahlsen, M.; Norrbom, J.; Jansson, E.; Hjeltnes, N.; Gustafsson, T.; Krook, A. Human skeletal muscle fibre type variations correlate with ppar alpha, ppar delta and PGC-1 alpha mrna. Acta Physiol. 2006, 188, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, K.M.; Haddad, F.; Pandorf, C.E.; Roy, R.R.; Edgerton, V.R. Alterations in muscle mass and contractile phenotype in response to unloading models: Role of transcriptional/pretranslational mechanisms. Front. Physiol. 2013, 4, 284. [Google Scholar] [CrossRef] [PubMed]
- Atherton, P.J.; Greenhaff, P.L.; Phillips, S.M.; Bodine, S.C.; Adams, C.M.; Lang, C.H. Control of skeletal muscle atrophy in response to disuse: Clinical/preclinical contentions and fallacies of evidence. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E594–E604. [Google Scholar] [CrossRef] [PubMed]
- Yarrow, J.F.; McCoy, S.C.; Borst, S.E. Intracrine and myotrophic roles of 5alpha-reductase and androgens: A review. Med. Sci. Sports Exerc. 2012, 44, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Burger, H.G. Androgen production in women. Fertil. Steril. 2002, 77 (Suppl. S4), S3–S5. [Google Scholar] [CrossRef]
- Labrie, F. All sex steroids are made intracellularly in peripheral tissues by the mechanisms of intracrinology after menopause. J. Steroid Biochem. Mol. Biol. 2015, 145, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Conover, C.F.; Yarrow, J.F.; Garrett, T.J.; Ye, F.; Quinlivan, E.P.; Cannady, D.F.; Peterson, M.D.; Borst, S.E. High prevalence of low serum biologically active testosterone in older male veterans. J. Am. Med. Dir. Assoc. 2017, 18, 366.e17–366.e24. [Google Scholar] [CrossRef] [PubMed]
- Laurent, M.R.; Helsen, C.; Antonio, L.; Schollaert, D.; Joniau, S.; Vos, M.J.; Decallonne, B.; Hammond, G.L.; Vanderschueren, D.; Claessens, F. Effects of sex hormone-binding globulin (shbg) on androgen bioactivity in vitro. Mol. Cell. Endocrinol. 2016, 437, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Thaler, M.A.; Seifert-Klauss, V.; Luppa, P.B. The biomarker sex hormone-binding globulin—From established applications to emerging trends in clinical medicine. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Yarrow, J.F.; Wronski, T.J.; Borst, S.E. Testosterone and adult male bone: Actions independent of 5alpha-reductase and aromatase. Exerc. Sport Sci. Rev. 2015, 43, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Lumbroso, S.; Sandillon, F.; Georget, V.; Lobaccaro, J.M.; Brinkmann, A.O.; Privat, A.; Sultan, C. Immunohistochemical localization and immunoblotting of androgen receptor in spinal neurons of male and female rats. Eur. J. Endocrinol. 1996, 134, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, T.; Ogata, A.; Demura, T.; Moriwaka, F.; Tashiro, K.; Koyanagi, T.; Nagashima, K. Identification of androgen receptor in the rat spinal motoneurons. Immunohistochemical and immunoblotting analyses with monoclonal antibody. Neurosci. Lett. 1993, 158, 5–8. [Google Scholar] [CrossRef]
- Cain, M.P.; Kramer, S.A.; Tindall, D.J.; Husmann, D.A. Expression of androgen receptor protein within the lumbar spinal cord during ontologic development and following antiandrogen induced cryptorchidism. J. Urol. 1994, 152, 766–769. [Google Scholar] [CrossRef]
- Bauer, E.R.; Daxenberger, A.; Petri, T.; Sauerwein, H.; Meyer, H.H. Characterisation of the affinity of different anabolics and synthetic hormones to the human androgen receptor, human sex hormone binding globulin and to the bovine progestin receptor. Acta Pathol. Microbiol. Immunol. Scand. 2000, 108, 838–846. [Google Scholar] [CrossRef]
- Almeida, M.; Laurent, M.R.; Dubois, V.; Claessens, F.; O’Brien, C.A.; Bouillon, R.; Vanderschueren, D.; Manolagas, S.C. Estrogens and androgens in skeletal physiology and pathophysiology. Physiol. Rev. 2017, 97, 135–187. [Google Scholar] [CrossRef] [PubMed]
- Gennari, L.; Nuti, R.; Bilezikian, J.P. Aromatase activity and bone homeostasis in men. J. Clin. Endocrinol. Metab. 2004, 89, 5898–5907. [Google Scholar] [CrossRef] [PubMed]
- Durga, A.; Sepahpanah, F.; Regozzi, M.; Hastings, J.; Crane, D.A. Prevalence of testosterone deficiency after spinal cord injury. PM&R 2011, 3, 929–932. [Google Scholar]
- Bauman, W.A.; La Fountaine, M.F.; Spungen, A.M. Age-related prevalence of low testosterone in men with spinal cord injury. J. Spinal Cord Med. 2014, 37, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.J.; Schopp, L.H.; Mazurek, M.O.; Zaniletti, I.; Lammy, A.B.; Martin, T.A.; Thomas, F.P.; Acuff, M.E. Testosterone levels among men with spinal cord injury: Relationship between time since injury and laboratory values. Am. J. Phys. Med. Rehabil. 2008, 87, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Bauman, W.A.; La Fountaine, M.F.; Cirnigliaro, C.M.; Kirshblum, S.C.; Spungen, A.M. Testicular responses to hcg stimulation at varying doses in men with spinal cord injury. Spinal Cord 2017, 55, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Bauman, W.A.; La Fountaine, M.F.; Cirnigliaro, C.M.; Kirshblum, S.C.; Spungen, A.M. Provocative stimulation of the hypothalamic-pituitary-testicular axis in men with spinal cord injury. Spinal Cord 2016, 54, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Bauman, W.A.; La Fountaine, M.F.; Cirnigliaro, C.M.; Kirshblum, S.C.; Spungen, A.M. Administration of increasing doses of gonadotropin-releasing hormone in men with spinal cord injury to investigate dysfunction of the hypothalamic-pituitary-gonadal axis. Spinal Cord 2018, 56, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Kostovski, E.; Iversen, P.O.; Birkeland, K.; Torjesen, P.A.; Hjeltnes, N. Decreased levels of testosterone and gonadotrophins in men with long-standing tetraplegia. Spinal Cord 2008, 46, 559–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkelstein, J.S.; Lee, H.; Burnett-Bowie, S.A.; Pallais, J.C.; Yu, E.W.; Borges, L.F.; Jones, B.F.; Barry, C.V.; Wulczyn, K.E.; Thomas, B.J.; et al. Gonadal steroids and body composition, strength, and sexual function in men. N. Engl. J. Med. 2013, 369, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Schopp, L.H.; Clark, M.; Mazurek, M.O.; Hagglund, K.J.; Acuff, M.E.; Sherman, A.K.; Childers, M.K. Testosterone levels among men with spinal cord injury admitted to inpatient rehabilitation. Am. J. Phys. Med. Rehabil. 2006, 85, 678–684, quiz 685–677. [Google Scholar] [CrossRef] [PubMed]
- Beggs, L.A.; Ye, F.; Ghosh, P.; Beck, D.T.; Conover, C.F.; Balaez, A.; Miller, J.R.; Phillips, E.G.; Zheng, N.; Williams, A.A.; et al. Sclerostin inhibition prevents spinal cord injury-induced cancellous bone loss. J. Bone Miner. Res. 2015, 30, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Collier, L.; Pan, J.; Qin, W.; Bauman, W.A.; Cardozo, C.P. Testosterone reduced methylprednisolone-induced muscle atrophy in spinal cord-injured rats. Spinal Cord 2012, 50, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Borst, S.E.; Yarrow, J.F.; Conover, C.F.; Nseyo, U.; Meuleman, J.R.; Lipinska, J.A.; Braith, R.W.; Beck, D.T.; Martin, J.S.; Morrow, M.; et al. Musculoskeletal and prostate effects of combined testosterone and finasteride administration in older hypogonadal men: A randomized, controlled trial. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E433–E442. [Google Scholar] [CrossRef] [PubMed]
- Cooper, I.S.; Rynearson, E.H.; Mac, C.C.; Power, M.H. The catabolic effect of trauma of the spinal cord and its investigative treatment with testosterone propionate; preliminary report. Proc. Staff Meet. Mayo Clin. 1950, 25, 326–330. [Google Scholar] [PubMed]
- Cooper, I.S.; Rynearson, E.H.; Mac, C.C.; Power, M.H. Testosterone propionate as a nitrogen-sparing agent after spinal cord injury. J. Am. Med. Assoc. 1951, 145, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Bauman, W.A.; La Fountaine, M.F.; Cirnigliaro, C.M.; Kirshblum, S.C.; Spungen, A.M. Lean tissue mass and energy expenditure are retained in hypogonadal men with spinal cord injury after discontinuation of testosterone replacement therapy. J. Spinal Cord Med. 2015, 38, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Bennett, N.C.; Gardiner, R.A.; Hooper, J.D.; Johnson, D.W.; Gobe, G.C. Molecular cell biology of androgen receptor signalling. Int. J. Biochem. Cell Biol. 2010, 42, 813–827. [Google Scholar] [CrossRef] [PubMed]
- MacLean, H.E.; Chiu, W.S.; Notini, A.J.; Axell, A.M.; Davey, R.A.; McManus, J.F.; Ma, C.; Plant, D.R.; Lynch, G.S.; Zajac, J.D. Impaired skeletal muscle development and function in male, but not female, genomic androgen receptor knockout mice. FASEB J. 2008, 22, 2676–2689. [Google Scholar] [CrossRef] [PubMed]
- Revankar, C.M.; Cimino, D.F.; Sklar, L.A.; Arterburn, J.B.; Prossnitz, E.R. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science 2005, 307, 1625–1630. [Google Scholar] [CrossRef] [PubMed]
- Foradori, C.D.; Weiser, M.J.; Handa, R.J. Non-genomic actions of androgens. Front. Neuroendocrinol. 2008, 29, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Bhasin, S.; Braga, M.; Artaza, J.N.; Pervin, S.; Taylor, W.E.; Krishnan, V.; Sinha, S.K.; Rajavashisth, T.B.; Jasuja, R. Regulation of myogenic differentiation by androgens: Cross talk between androgen receptor/beta-catenin and follistatin/transforming growth factor-beta signaling pathways. Endocrinology 2009, 150, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Rudnicki, M.A.; Williams, B.O. Wnt signaling in bone and muscle. Bone 2015, 80, 60–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.H.; Wu, Y.; Yao, S.; Levine, A.C.; Kirschenbaum, A.; Collier, L.; Bauman, W.A.; Cardozo, C.P. Androgens up-regulate transcription of the notch inhibitor numb in c2c12 myoblasts via wnt/beta-catenin signaling to t cell factor elements in the numb promoter. J. Biol. Chem. 2013, 288, 17990–17998. [Google Scholar] [CrossRef] [PubMed]
- Gentile, M.A.; Nantermet, P.V.; Vogel, R.L.; Phillips, R.; Holder, D.; Hodor, P.; Cheng, C.; Dai, H.; Freedman, L.P.; Ray, W.J. Androgen-mediated improvement of body composition and muscle function involves a novel early transcriptional program including igf1, mechano growth factor, and induction of {beta}-catenin. J. Mol. Endocrinol. 2010, 44, 55–73. [Google Scholar] [CrossRef] [PubMed]
- Otto-Duessel, M.; Tew, B.Y.; Vonderfecht, S.; Moore, R.; Jones, J.O. Identification of neuron selective androgen receptor inhibitors. World J. Biol. Chem. 2017, 8, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Pawlowski, J.E.; Ertel, J.R.; Allen, M.P.; Xu, M.; Butler, C.; Wilson, E.M.; Wierman, M.E. Liganded androgen receptor interaction with beta-catenin: Nuclear co-localization and modulation of transcriptional activity in neuronal cells. J. Biol. Chem. 2002, 277, 20702–20710. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.S., Jr.; Fraley, G.S.; Damian, V.; Woodke, L.B.; Zapata, F.; Sopher, B.L.; Plymate, S.R.; La Spada, A.R. Loss of endogenous androgen receptor protein accelerates motor neuron degeneration and accentuates androgen insensitivity in a mouse model of x-linked spinal and bulbar muscular atrophy. Hum. Mol. Genet. 2006, 15, 2225–2238. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Wang, Y.S.; Yuan, Y.J.; Wan, Z.H.; Yao, T.C.; Li, H.H.; Tang, P.F.; Mei, X.F. Neuroprotective effect of rapamycin on spinal cord injury via activation of the wnt/beta-catenin signaling pathway. Neural Regen. Res. 2015, 10, 951–957. [Google Scholar] [PubMed]
- Lu, G.B.; Niu, F.W.; Zhang, Y.C.; Du, L.; Liang, Z.Y.; Gao, Y.; Yan, T.Z.; Nie, Z.K.; Gao, K. Methylprednisolone promotes recovery of neurological function after spinal cord injury: Association with wnt/beta-catenin signaling pathway activation. Neural Regen. Res. 2016, 11, 1816–1823. [Google Scholar] [PubMed]
- Gao, K.; Shen, Z.; Yuan, Y.; Han, D.; Song, C.; Guo, Y.; Mei, X. Simvastatin inhibits neural cell apoptosis and promotes locomotor recovery via activation of wnt/beta-catenin signaling pathway after spinal cord injury. J. Neurochem 2016, 138, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; McCoy, S.C.; Ross, H.H.; Bernardo, J.A.; Beharry, A.W.; Senf, S.M.; Judge, A.R.; Beck, D.T.; Conover, C.F.; Cannady, D.F.; et al. Transcriptional regulation of myotrophic actions by testosterone and trenbolone on androgen-responsive muscle. Steroids 2014, 87, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Bauman, W.A.; Blitzer, R.D.; Cardozo, C. Testosterone-induced hypertrophy of l6 myoblasts is dependent upon erk and mtor. Biochem. Biophys. Res. Commun. 2010, 400, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhao, W.; Zhao, J.; Pan, J.; Wu, Q.; Zhang, Y.; Bauman, W.A.; Cardozo, C.P. Identification of androgen response elements in the insulin-like growth factor i upstream promoter. Endocrinology 2007, 148, 2984–2993. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, A.A.; Sheffield-Moore, M.; Yeckel, C.W.; Gilkison, C.; Jiang, J.; Achacosa, A.; Lieberman, S.A.; Tipton, K.; Wolfe, R.R.; Urban, R.J. Testosterone administration to older men improves muscle function: Molecular and physiological mechanisms. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E601–E607. [Google Scholar] [CrossRef] [PubMed]
- Deane, C.S.; Hughes, D.C.; Sculthorpe, N.; Lewis, M.P.; Stewart, C.E.; Sharples, A.P. Impaired hypertrophy in myoblasts is improved with testosterone administration. J. Steroid Biochem. Mol. Biol. 2013, 138, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.P.; Gao, S.; Puppa, M.J.; Sato, S.; Welle, S.L.; Carson, J.A. Testosterone regulation of akt/mtorc1/foxo3a signaling in skeletal muscle. Mol. Cell. Endocrinol. 2013, 365, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Basualto-Alarcon, C.; Jorquera, G.; Altamirano, F.; Jaimovich, E.; Estrada, M. Testosterone signals through mtor and androgen receptor to induce muscle hypertrophy. Med. Sci. Sports Exerc. 2013, 45, 1712–1720. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Shen, Y.; Pink, H.; Triantafillou, J.; Stimpson, S.A.; Turnbull, P.; Han, B. Phosphorylation of p70s6 kinase is implicated in androgen-induced levator ani muscle anabolism in castrated rats. J. Steroid Biochem. Mol. Biol. 2004, 92, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Serra, C.; Sandor, N.L.; Jang, H.; Lee, D.; Toraldo, G.; Guarneri, T.; Wong, S.; Zhang, A.; Guo, W.; Jasuja, R.; et al. The effects of testosterone deprivation and supplementation on proteasomal and autophagy activity in the skeletal muscle of the male mouse: Differential effects on high-androgen responder and low-androgen responder muscle groups. Endocrinology 2013, 154, 4594–4606. [Google Scholar] [CrossRef] [PubMed]
- Pires-Oliveira, M.; Maragno, A.L.; Parreiras-e-Silva, L.T.; Chiavegatti, T.; Gomes, M.D.; Godinho, R.O. Testosterone represses ubiquitin ligases atrogin-1 and murf-1 expression in an androgen-sensitive rat skeletal muscle in vivo. J. Appl. Physiol. 2010, 108, 266–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrando, A.A.; Sheffield-Moore, M.; Paddon-Jones, D.; Wolfe, R.R.; Urban, R.J. Differential anabolic effects of testosterone and amino acid feeding in older men. J. Clin. Endocrinol. Metab. 2003, 88, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Pan, J.; Wang, X.; Wu, Y.; Bauman, W.A.; Cardozo, C.P. Expression of the muscle atrophy factor muscle atrophy f-box is suppressed by testosterone. Endocrinology 2008, 149, 5449–5460. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ruggiero, C.L.; Bauman, W.A.; Cardozo, C. Ankrd1 is a transcriptional repressor for the androgen receptor that is downregulated by testosterone. Biochem. Biophys. Res. Commun. 2013, 437, 355–360. [Google Scholar] [CrossRef] [PubMed]
- MacKrell, J.G.; Yaden, B.C.; Bullock, H.; Chen, K.; Shetler, P.; Bryant, H.U.; Krishnan, V. Molecular targets of androgen signaling that characterize skeletal muscle recovery and regeneration. Nucl. Recept. Signal. 2015, 13, e005. [Google Scholar] [PubMed]
- Zhao, J.; Bauman, W.A.; Huang, R.; Caplan, A.J.; Cardozo, C. Oxandrolone blocks glucocorticoid signaling in an androgen receptor-dependent manner. Steroids 2004, 69, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.R.; Shiguemoto, G.E.; Tomaz, L.M.; Ferreira, F.C.; Rodrigues, M.F.; Domingues, M.M.; Souza Master, M.V.; Canevazzi, G.H.; Silva-Magosso, N.S.; Selistre-de-Araujo, H.S.; et al. Resistance training and ovariectomy: Antagonic effects in mitochondrial biogenesis markers in rat skeletal muscle. Int. J. Sports Med. 2016, 37, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.M.; Pereira, R.I.; Erickson, C.B.; Swibas, T.A.; Kang, C.; Van Pelt, R.E. Time since menopause and skeletal muscle estrogen receptors, PGC-1alpha, and ampk. Menopause 2017, 24, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Maher, A.C.; Akhtar, M.; Tarnopolsky, M.A. Men supplemented with 17beta-estradiol have increased beta-oxidation capacity in skeletal muscle. Physiol. Genom. 2010, 42, 342–347. [Google Scholar] [CrossRef] [PubMed]
- McCully, K.K.; Mulcahy, T.K.; Ryan, T.E.; Zhao, Q. Skeletal muscle metabolism in individuals with spinal cord injury. J. Appl. Physiol. 2011, 111, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patte-Mensah, C.; Penning, T.M.; Mensah-Nyagan, A.G. Anatomical and cellular localization of neuroactive 5 alpha/3 alpha-reduced steroid-synthesizing enzymes in the spinal cord. J. Comp. Neurol. 2004, 477, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Poletti, A.; Coscarella, A.; Negri-Cesi, P.; Colciago, A.; Celotti, F.; Martini, L. 5 alpha-reductase isozymes in the central nervous system. Steroids 1998, 63, 246–251. [Google Scholar] [CrossRef]
- Hauser, K.F.; MacLusky, N.J.; Toran-Allerand, C.D. Androgen action in fetal mouse spinal cord cultures: Metabolic and morphologic aspects. Brain Res. 1987, 406, 62–72. [Google Scholar] [CrossRef]
- MacLusky, N.J.; Clark, C.R.; Shanabrough, M.; Naftolin, F. Metabolism and binding of androgens in the spinal cord of the rat. Brain Res. 1987, 422, 83–91. [Google Scholar] [CrossRef]
- Pozzi, P.; Bendotti, C.; Simeoni, S.; Piccioni, F.; Guerini, V.; Marron, T.U.; Martini, L.; Poletti, A. Androgen 5-alpha-reductase type 2 is highly expressed and active in rat spinal cord motor neurones. J. Neuroendocrinol. 2003, 15, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Hauser, K.F.; Toran-Allerand, C.D. Androgen increases the number of cells in fetal mouse spinal cord cultures: Implications for motoneuron survival. Brain Res. 1989, 485, 157–164. [Google Scholar] [CrossRef]
- Fargo, K.N.; Foecking, E.M.; Jones, K.J.; Sengelaub, D.R. Neuroprotective actions of androgens on motoneurons. Front. Neuroendocrinol. 2009, 30, 130–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhovshek, T.; Buckley, K.E.; Sergent, M.A.; Sengelaub, D.R. Testosterone metabolites differentially maintain adult morphology in a sexually dimorphic neuromuscular system. Dev. Neurobiol. 2010, 70, 206–221. [Google Scholar] [CrossRef] [PubMed]
- Huguenard, A.L.; Fernando, S.M.; Monks, D.A.; Sengelaub, D.R. Overexpression of androgen receptors in target musculature confers androgen sensitivity to motoneuron dendrites. Endocrinology 2011, 152, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.X.; Zhao, M.; Liu, Y.L.; Chen, L.S.; Hao, P.L.; Sun, C. Expression of aromatase and estrogen receptors in lumbar motoneurons of mice. Neurosci. Lett. 2017, 653, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Rakotoarivelo, C.; Petite, D.; Lambard, S.; Fabre, C.; Rouleau, C.; Lumbroso, S.; de Weille, J.; Privat, A.; Carreau, S.; Mersel, M. Receptors to steroid hormones and aromatase are expressed by cultured motoneurons but not by glial cells derived from rat embryo spinal cord. Neuroendocrinology 2004, 80, 284–297. [Google Scholar] [CrossRef] [PubMed]
- Nowacek, A.S.; Sengelaub, D.R. Estrogenic support of motoneuron dendritic growth via the neuromuscular periphery in a sexually dimorphic motor system. J. Neurobiol. 2006, 66, 962–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Chew, C.; Munoz, F.; Sengelaub, D.R. Neuroprotective effects of testosterone metabolites and dependency on receptor action on the morphology of somatic motoneurons following the death of neighboring motoneurons. Dev. Neurobiol. 2017, 77, 691–707. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, L.M.; Sengelaub, D.R. Castration-induced upregulation of muscle eralpha supports estrogen sensitivity of motoneuron dendrites in a sexually dimorphic neuromuscular system. Dev. Neurobiol. 2013, 73, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Nakamizo, T.; Urushitani, M.; Inoue, R.; Shinohara, A.; Sawada, H.; Honda, K.; Kihara, T.; Akaike, A.; Shimohama, S. Protection of cultured spinal motor neurons by estradiol. Neuroreport 2000, 11, 3493–3497. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hu, R.; Ge, H.; Duanmu, W.; Li, Y.; Xue, X.; Hu, S.; Feng, H. G-protein-coupled receptor 30-mediated antiapoptotic effect of estrogen on spinal motor neurons following injury and its underlying mechanisms. Mol. Med. Rep. 2015, 12, 1733–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkabes, S.; Nicot, A.B. Sex steroids and neuroprotection in spinal cord injury: A review of preclinical investigations. Exp. Neurol. 2014, 259, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Ottem, E.N.; Bailey, D.J.; Jordan, C.L.; Breedlove, S.M. With a little help from my friends: Androgens tap bdnf signaling pathways to alter neural circuits. Neuroscience 2013, 239, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Verhovshek, T.; Rudolph, L.M.; Sengelaub, D.R. Brain-derived neurotrophic factor and androgen interactions in spinal neuromuscular systems. Neuroscience 2013, 239, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, M.C.; Verhovshek, T.; Sengelaub, D.R. Androgen regulates trkb immunolabeling in spinal motoneurons. J. Neurosci. Res. 2007, 85, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Ottem, E.N.; Beck, L.A.; Jordan, C.L.; Breedlove, S.M. Androgen-dependent regulation of brain-derived neurotrophic factor and tyrosine kinase b in the sexually dimorphic spinal nucleus of the bulbocavernosus. Endocrinology 2007, 148, 3655–3665. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.Y.; Arnold, A.P. Interaction of bdnf and testosterone in the regulation of adult perineal motoneurons. J. Neurobiol. 2000, 44, 308–319. [Google Scholar] [CrossRef]
- Yang, L.Y.; Verhovshek, T.; Sengelaub, D.R. Brain-derived neurotrophic factor and androgen interact in the maintenance of dendritic morphology in a sexually dimorphic rat spinal nucleus. Endocrinology 2004, 145, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Gingras, K.M.; Bengston, L.; Di Marco, A.; Forger, N.G. Blockade of endogenous neurotrophic factors prevents the androgenic rescue of rat spinal motoneurons. J. Neurosci. 2001, 21, 4366–4372. [Google Scholar] [CrossRef] [PubMed]
- Gill, L.C.; Gransee, H.M.; Sieck, G.C.; Mantilla, C.B. Functional recovery after cervical spinal cord injury: Role of neurotrophin and glutamatergic signaling in phrenic motoneurons. Respir. Physiol. Neurobiol. 2016, 226, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona-Rossinyol, A.; Mir, M.; Caraballo-Miralles, V.; Llado, J.; Olmos, G. Neuroprotective effects of estradiol on motoneurons in a model of rat spinal cord embryonic explants. Cell. Mol. Neurobiol. 2013, 33, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.W.; Chen, B.; Huang, K.L.; Dai, Y.S.; Teng, H.L. Inhibition of autophagy by estradiol promotes locomotor recovery after spinal cord injury in rats. Neurosci. Bull. 2016, 32, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Behrman, A.L.; Harkema, S.J. Physical rehabilitation as an agent for recovery after spinal cord injury. Phys. Med. Rehabil. Clin. 2007, 18, 183–202. [Google Scholar] [CrossRef] [PubMed]
- Field-Fote, E.C. Combined use of body weight support, functional electric stimulation, and treadmill training to improve walking ability in individuals with chronic incomplete spinal cord Injury. Arch. Phys. Med. Rehabil. 2001, 82, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Forrest, G.F.; Sisto, S.A.; Barbeau, H.; Kirshblum, S.C.; Wilen, J.; Bond, Q.; Bentson, S.; Asselin, P.; Cirnigliaro, C.M.; Harkema, S. Neuromotor and musculoskeletal responses to locomotor training for an individual with chronic motor complete ais-b spinal cord injury. J. Spinal Cord Med. 2008, 31, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Gorassini, M.A.; Norton, J.A.; Nevett-Duchcherer, J.; Roy, F.D.; Yang, J.F. Changes in locomotor muscle activity after treadmill training in subjects with incomplete spinal cord injury. J. Neurophysiol. 2009, 101, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Harkema, S.J.; Schmidt-Read, M.; Lorenz, D.J.; Edgerton, V.R.; Behrman, A.L. Balance and ambulation improvements in individuals with chronic incomplete spinal cord injury using locomotor training-based rehabilitation. Arch. Phys. Med. Rehabil. 2012, 93, 1508–1517. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Kindig, M.; Mirbagheri, M. Robotic-assisted locomotor training enhances ankle performance in adults with incomplete spinal cord injury. J. Rehabil. Med. 2016, 48, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Nooijen, C.F.; Ter Hoeve, N.; Field-Fote, E.C. Gait quality is improved by locomotor training in individuals with sci regardless of training approach. J. Neuroeng. Rehabil. 2009, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Wernig, A.; Muller, S.; Nanassy, A.; Cagol, E. Laufband therapy based on ‘rules of spinal locomotion’ is effective in spinal cord injured persons. Eur. J. Neurosci. 1995, 7, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Wirz, M.; Bastiaenen, C.; de Bie, R.; Dietz, V. Effectiveness of automated locomotor training in patients with acute incomplete spinal cord injury: A randomized controlled multicenter trial. BMC Neurol. 2011, 11, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirz, M.; Zemon, D.H.; Rupp, R.; Scheel, A.; Colombo, G.; Dietz, V.; Hornby, T.G. Effectiveness of automated locomotor training in patients with chronic incomplete spinal cord injury: A multicenter trial. Arch. Phys. Med. Rehabil. 2005, 86, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Evans, N.; Tefertiller, C.; Backus, D.; Sweatman, M.; Tansey, K.; Morrison, S. Activity-based therapy for recovery of walking in individuals with chronic spinal cord injury: Results from a randomized clinical trial. Arch. Phys. Med. Rehabil. 2014, 95, 2239–2246. [Google Scholar] [CrossRef] [PubMed]
- Knikou, M. Plasticity of corticospinal neural control after locomotor training in human spinal cord injury. Neural Plast. 2012, 2012, 254948. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.M.; Ditor, D.S.; Tarnopolsky, M.A.; Phillips, S.M.; McCartney, N.; Hicks, A.L. The effect of body weight-supported treadmill training on muscle morphology in an individual with chronic, motor-complete spinal cord injury: A case study. J. Spinal Cord Med. 2006, 29, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Alexeeva, N.; Sames, C.; Jacobs, P.L.; Hobday, L.; Distasio, M.M.; Mitchell, S.A.; Calancie, B. Comparison of training methods to improve walking in persons with chronic spinal cord injury: A randomized clinical trial. J. Spinal Cord Med. 2011, 34, 362–379. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.L.; Gorassini, M.A. Increases in corticospinal tract function by treadmill training after incomplete spinal cord injury. J. Neurophysiol. 2005, 94, 2844–2855. [Google Scholar] [CrossRef] [PubMed]
- Coupaud, S.; Jack, L.P.; Hunt, K.J.; Allan, D.B. Muscle and bone adaptations after treadmill training in incomplete spinal cord injury: A case study using peripheral quantitative computed tomography. J. Musculoskelet. Neuronal Interact. 2009, 9, 288–297. [Google Scholar] [PubMed]
- Giangregorio, L.; McCartney, N. Bone loss and muscle atrophy in spinal cord injury: Epidemiology, fracture prediction, and rehabilitation strategies. J. Spinal Cord Med. 2006, 29, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Giangregorio, L.M.; Hicks, A.L.; Webber, C.E.; Phillips, S.M.; Craven, B.C.; Bugaresti, J.M.; McCartney, N. Body weight supported treadmill training in acute spinal cord injury: Impact on muscle and bone. Spinal Cord 2005, 43, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Stewart, B.G.; Tarnopolsky, M.A.; Hicks, A.L.; McCartney, N.; Mahoney, D.J.; Staron, R.S.; Phillips, S.M. Treadmill training-induced adaptations in muscle phenotype in persons with incomplete spinal cord injury. Muscle Nerve 2004, 30, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Dobkin, B.; Barbeau, H.; Deforge, D.; Ditunno, J.; Elashoff, R.; Apple, D.; Basso, M.; Behrman, A.; Harkema, S.; Saulino, M.; et al. The evolution of walking-related outcomes over the first 12 weeks of rehabilitation for incomplete traumatic spinal cord injury: The multicenter randomized spinal cord injury locomotor trial. NeuroRehabil. Neural Repair 2007, 21, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Field-Fote, E.C.; Roach, K.E. Influence of a locomotor training approach on walking speed and distance in people with chronic spinal cord injury: A randomized clinical trial. Phys. Ther. 2011, 91, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Lucareli, P.R.; Lima, M.O.; Lima, F.P.; de Almeida, J.G.; Brech, G.C.; D’Andrea Greve, J.M. Gait analysis following treadmill training with body weight support versus conventional physical therapy: A prospective randomized controlled single blind study. Spinal Cord 2011, 49, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Postans, N.J.; Hasler, J.P.; Granat, M.H.; Maxwell, D.J. Functional electric stimulation to augment partial weight-bearing supported treadmill training for patients with acute incomplete spinal cord injury: A pilot study. Arch. Phys. Med. Rehabil. 2004, 85, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Mehrholz, J.; Kugler, J.; Pohl, M. Locomotor training for walking after spinal cord injury. Cochrane Database Syst. Rev. 2012, 11, CD006676. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.Y.; Kim, H.J.; Kwon, B.S.; Park, J.W.; Lee, H.J.; Yoo, A. Robot-assisted gait training (lokomat) improves walking function and activity in people with spinal cord injury: A systematic review. J. Neuroeng. Rehabil. 2017, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Wessels, M.; Lucas, C.; Eriks, I.; de Groot, S. Body weight-supported gait training for restoration of walking in people with an incomplete spinal cord injury: A systematic review. J. Rehabil. Med. 2010, 42, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.; Noonan, V.K.; Eng, J.J. A systematic review of functional ambulation outcome measures in spinal cord injury. Spinal Cord 2008, 46, 246–254. [Google Scholar] [CrossRef] [PubMed]
- De Leon, R.D.; Dy, C.J. What did we learn from the animal studies of body weight-supported treadmill training and where do we go from here? J. Neurotrauma 2017, 34, 1744–1750. [Google Scholar] [CrossRef] [PubMed]
- Yarrow, J.F.; Conover, C.F.; Lipinska, J.A.; Santillana, C.A.; Wronski, T.J.; Borst, S.E. Methods to quantify sex steroid hormones in bone: Applications to the study of androgen ablation and administration. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E841–E847. [Google Scholar] [CrossRef] [PubMed]
- Nessler, J.A.; Moustafa-Bayoumi, M.; Soto, D.; Duhon, J.E.; Schmitt, R. Robot applied stance loading increases hindlimb muscle mass and stepping kinetics in a rat model of spinal cord injury. In Proceedings of the 2011 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Boston, MA, USA, 30 August–3 September 2011; Volume 2011, pp. 4145–4148. [Google Scholar]
- Ilha, J.; da Cunha, N.B.; Jaeger, M.; de Souza, D.F.; Nascimento, P.S.; Marcuzzo, S.; Figueiro, M.; Gottfried, C.; Achaval, M. Treadmill step training-induced adaptive muscular plasticity in a chronic paraplegia model. Neurosci. Lett. 2011, 492, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.E.; Liu, M.; Bose, P.; O’Steen, W.A.; Thompson, F.J.; Anderson, D.K.; Vandenborne, K. Changes in soleus muscle function and fiber morphology with one week of locomotor training in spinal cord contusion injured rats. J. Neurotrauma 2006, 23, 1671–1681. [Google Scholar] [CrossRef] [PubMed]
- Bose, P.K.; Hou, J.; Parmer, R.; Reier, P.J.; Thompson, F.J. Altered patterns of reflex excitability, balance, and locomotion following spinal cord injury and locomotor training. Front. Physiol. 2012, 3, 258. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Bose, P.; Walter, G.A.; Thompson, F.J.; Vandenborne, K. A longitudinal study of skeletal muscle following spinal cord injury and locomotor training. Spinal Cord 2008, 46, 488–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Balasubramanian, S.; Murray, M.; Lemay, M.; Houle, J. Role of spared pathways in locomotor recovery after body-weight-supported treadmill training in contused rats. J. Neurotrauma 2011, 28, 2405–2416. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, A.; Shah, P.; Gregory, C.; Bowden, M.; Stevens, J.; Bishop, M.; Walter, G.; Behrman, A.; Vandenborne, K. Locomotor training and muscle function after incomplete spinal cord injury: Case series. J. Spinal Cord Med. 2008, 31, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Duffell, L.D.; Brown, G.L.; Mirbagheri, M.M. Facilitatory effects of anti-spastic medication on robotic locomotor training in people with chronic incomplete spinal cord injury. J. Neuroeng. Rehabil. 2015, 12, 29. [Google Scholar] [CrossRef] [PubMed]
- Varoqui, D.; Niu, X.; Mirbagheri, M.M. Ankle voluntary movement enhancement following robotic-assisted locomotor training in spinal cord injury. J. Neuroeng. Rehabil. 2014, 11, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardozo, C.P.; Graham, Z.A. Muscle-bone interactions: Movement in the field of mechano-humoral coupling of muscle and bone. Ann. N. Y. Acad. Sci. 2017, 1402, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, A.; Liu, M.; Ye, F.; Walter, G.A.; Vandenborne, K. Regenerative responses in slow- and fast-twitch muscles following moderate contusion spinal cord injury and locomotor training. Eur. J. Appl. Physiol. 2013, 113, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Dupont-Versteegden, E.E.; Murphy, R.J.; Houle, J.D.; Gurley, C.M.; Peterson, C.A. Mechanisms leading to restoration of muscle size with exercise and transplantation after spinal cord injury. Am. J. Physiol. Cell Physiol. 2000, 279, C1677–C1684. [Google Scholar] [CrossRef] [PubMed]
- Astorino, T.A.; Harness, E.T.; Witzke, K.A. Chronic activity-based therapy does not improve body composition, insulin-like growth factor-i, adiponectin, or myostatin in persons with spinal cord injury. J. Spinal Cord Med. 2015, 38, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Petrie, M.; Suneja, M.; Shields, R.K. Low-frequency stimulation regulates metabolic gene expression in paralyzed muscle. J. Appl. Physiol. 2015, 118, 723–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorgey, A.S.; Graham, Z.A.; Bauman, W.A.; Cardozo, C.; Gater, D.R. Abundance in proteins expressed after functional electrical stimulation cycling or arm cycling ergometry training in persons with chronic spinal cord injury. J. Spinal Cord Med. 2017, 40, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.M.; Suneja, M.; Dudley-Javoroski, S.; Shields, R.K. Altered mrna expression after long-term soleus electrical stimulation training in humans with paralysis. Muscle Nerve 2011, 43, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Cote, M.P.; Azzam, G.A.; Lemay, M.A.; Zhukareva, V.; Houle, J.D. Activity-dependent increase in neurotrophic factors is associated with an enhanced modulation of spinal reflexes after spinal cord injury. J. Neurotrauma 2011, 28, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Goldshmit, Y.; Lythgo, N.; Galea, M.P.; Turnley, A.M. Treadmill training after spinal cord hemisection in mice promotes axonal sprouting and synapse formation and improves motor recovery. J. Neurotrauma 2008, 25, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Petruska, J.C.; Ichiyama, R.M.; Jindrich, D.L.; Crown, E.D.; Tansey, K.E.; Roy, R.R.; Edgerton, V.R.; Mendell, L.M. Changes in motoneuron properties and synaptic inputs related to step training after spinal cord transection in rats. J. Neurosci. 2007, 27, 4460–4471. [Google Scholar] [CrossRef] [PubMed]
- Knikou, M. Neurophysiological characteristics of human leg muscle action potentials evoked by transcutaneous magnetic stimulation of the spine. Bioelectromagnetics 2013, 34, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Knikou, M.; Mummidisetty, C.K. Locomotor training improves premotoneuronal control after chronic spinal cord injury. J. Neurophysiol. 2014, 111, 2264–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.C.; Mummidisetty, C.K.; Rymer, W.Z.; Knikou, M. Locomotor training alters the behavior of flexor reflexes during walking in human spinal cord injury. J. Neurophysiol. 2014, 112, 2164–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.C.; Knikou, M. A review on locomotor training after spinal cord injury: Reorganization of spinal neuronal circuits and recovery of motor function. Neural Plast. 2016, 2016, 1216258. [Google Scholar] [CrossRef] [PubMed]
- Alcobendas-Maestro, M.; Esclarin-Ruz, A.; Casado-Lopez, R.M.; Munoz-Gonzalez, A.; Perez-Mateos, G.; Gonzalez-Valdizan, E.; Martin, J.L. Lokomat robotic-assisted versus overground training within 3 to 6 months of incomplete spinal cord lesion: Randomized controlled trial. NeuroRehabil. Neural Repair 2012, 26, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Esclarin-Ruz, A.; Alcobendas-Maestro, M.; Casado-Lopez, R.; Perez-Mateos, G.; Florido-Sanchez, M.A.; Gonzalez-Valdizan, E.; Martin, J.L. A comparison of robotic walking therapy and conventional walking therapy in individuals with upper versus lower motor neuron lesions: A randomized controlled trial. Arch. Phys. Med. Rehabil. 2014, 95, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Field-Fote, E.C.; Lindley, S.D.; Sherman, A.L. Locomotor training approaches for individuals with spinal cord injury: A preliminary report of walking-related outcomes. J. Neurol. Phys. Ther. 2005, 29, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Kapadia, N.; Masani, K.; Catharine Craven, B.; Giangregorio, L.M.; Hitzig, S.L.; Richards, K.; Popovic, M.R. A randomized trial of functional electrical stimulation for walking in incomplete spinal cord injury: Effects on walking competency. J. Spinal Cord Med. 2014, 37, 511–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labruyere, R.; van Hedel, H.J. Strength training versus robot-assisted gait training after incomplete spinal cord injury: A randomized pilot study in patients depending on walking assistance. J. Neuroeng. Rehabil. 2014, 11, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, T.; Pauhl, K.; Ferguson, A.; Malik, R.N.; Krassioukov, A.; Eng, J.J. Training with robot-applied resistance in people with motor-incomplete spinal cord injury: Pilot study. J. Rehabil. Res. Dev. 2015, 52, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.A.; Lorenz, D.; Eskay, C.P.; Forrest, G.F.; Basso, D.M. Longitudinal recovery and reduced costs after 120 sessions of locomotor training for motor incomplete spinal cord injury. Arch. Phys. Med. Rehabil. 2018, 99, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Varoqui, D.; Kindig, M.; Mirbagheri, M.M. Prediction of gait recovery in spinal cord injured individuals trained with robotic gait orthosis. J. Neuroeng. Rehabil. 2014, 11, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winchester, P.; McColl, R.; Querry, R.; Foreman, N.; Mosby, J.; Tansey, K.; Williamson, J. Changes in supraspinal activation patterns following robotic locomotor therapy in motor-incomplete spinal cord injury. NeuroRehabil. Neural Repair 2005, 19, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Landry, J.M.; Schmit, B.D.; Hornby, T.G.; Yen, S.C. Robotic resistance treadmill training improves locomotor function in human spinal cord injury: A pilot study. Arch. Phys. Med. Rehabil. 2012, 93, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Galen, S.S.; Clarke, C.J.; McLean, A.N.; Allan, D.B.; Conway, B.A. Isometric hip and knee torque measurements as an outcome measure in robot assisted gait training. NeuroRehabilitation 2014, 34, 287–295. [Google Scholar] [PubMed]
- Maier, I.C.; Ichiyama, R.M.; Courtine, G.; Schnell, L.; Lavrov, I.; Edgerton, V.R.; Schwab, M.E. Differential effects of anti-nogo-a antibody treatment and treadmill training in rats with incomplete spinal cord injury. Brain 2009, 132, 1426–1440. [Google Scholar] [CrossRef] [PubMed]
- Battistuzzo, C.R.; Rank, M.M.; Flynn, J.R.; Morgan, D.L.; Callister, R.; Callister, R.J.; Galea, M.P. Gait recovery following spinal cord injury in mice: Limited effect of treadmill training. J. Spinal Cord Med. 2016, 39, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battistuzzo, C.R.; Rank, M.M.; Flynn, J.R.; Morgan, D.L.; Callister, R.; Callister, R.J.; Galea, M.P. Effects of treadmill training on hindlimb muscles of spinal cord-injured mice. Muscle Nerve 2017, 55, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.K.; Garcia-Alias, G.; Choe, J.; Gad, P.; Gerasimenko, Y.; Tillakaratne, N.; Zhong, H.; Roy, R.R.; Edgerton, V.R. Use of quadrupedal step training to re-engage spinal interneuronal networks and improve locomotor function after spinal cord injury. Brain 2013, 136, 3362–3377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nessler, J.A.; De Leon, R.D.; Sharp, K.; Kwak, E.; Minakata, K.; Reinkensmeyer, D.J. Robotic gait analysis of bipedal treadmill stepping by spinal contused rats: Characterization of intrinsic recovery and comparison with bbb. J. Neurotrauma 2006, 23, 882–896. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.J.; Seo, T.B.; Kwon, K.B.; Yoon, S.J.; Elzi, D.J.; Kim, B.G.; Namgung, U. Axonal outgrowth and erk1/2 activation by training after spinal cord injury in rats. J. Neurotrauma 2009, 26, 2071–2082. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.Y.; Kim, H.; Kwon, M.J.; Hwang, D.H.; Lee, K.; Kim, B.G. Molecular and cellular changes in the lumbar spinal cord following thoracic injury: Regulation by treadmill locomotor training. PLoS ONE 2014, 9, e88215. [Google Scholar] [CrossRef] [PubMed]
- Multon, S.; Franzen, R.; Poirrier, A.L.; Scholtes, F.; Schoenen, J. The effect of treadmill training on motor recovery after a partial spinal cord compression-injury in the adult rat. J. Neurotrauma 2003, 20, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Cao, Y.; Dong, C.; Wang, H.; Wang, Q.; Tong, W.; Li, X.; Shan, C.; Wang, T. Neuromuscular interaction is required for neurotrophins-mediated locomotor recovery following treadmill training in rat spinal cord injury. PeerJ 2016, 4, e2025. [Google Scholar] [CrossRef] [PubMed]
- Foret, A.; Quertainmont, R.; Botman, O.; Bouhy, D.; Amabili, P.; Brook, G.; Schoenen, J.; Franzen, R. Stem cells in the adult rat spinal cord: Plasticity after injury and treadmill training exercise. J. Neurochem. 2010, 112, 762–772. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.J.; Herrity, A.N.; Smith, R.R.; Willhite, A.; Harrison, B.J.; Petruska, J.C.; Harkema, S.J.; Hubscher, C.H. Novel multi-system functional gains via task specific training in spinal cord injured male rats. J. Neurotrauma 2014, 31, 819–833. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Lee, Y.; Park, S.; Lee, S.; Hong, Y.; Kil Lee, S. Synergistic effect of melatonin on exercise-induced neuronal reconstruction and functional recovery in a spinal cord injury animal model. J. Pineal Res. 2010, 48, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, B.; Liu, C.; Zhao, D. Molecular mechanisms underlying the positive role of treadmill training in locomotor recovery after spinal cord injury. Spinal Cord 2017, 55, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Hayashibe, M.; Homma, T.; Fujimoto, K.; Oi, T.; Yagi, N.; Kashihara, M.; Nishikawa, N.; Ishizumi, Y.; Abe, S.; Hashimoto, H.; et al. Locomotor improvement of spinal cord-injured rats through treadmill training by forced plantar placement of hind paws. Spinal Cord 2016, 54, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Heng, C.; de Leon, R.D. Treadmill training enhances the recovery of normal stepping patterns in spinal cord contused rats. Exp. Neurol. 2009, 216, 139–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinozaki, M.; Iwanami, A.; Fujiyoshi, K.; Tashiro, S.; Kitamura, K.; Shibata, S.; Fujita, H.; Nakamura, M.; Okano, H. Combined treatment with chondroitinase abc and treadmill rehabilitation for chronic severe spinal cord injury in adult rats. Neurosci. Res. 2016, 113, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Robert, A.A.; Al Jadid, M.S.; Bin Afif, S.; Al Sowyed, A.A.; Al-Mubarak, S. The effects of different rehabilitation strategies on the functional recovery of spinal cord injured rats: An experimental study. Spine 2010, 35, E1273–E1277. [Google Scholar] [CrossRef] [PubMed]
- Ichiyama, R.; Potuzak, M.; Balak, M.; Kalderon, N.; Edgerton, V.R. Enhanced motor function by training in spinal cord contused rats following radiation therapy. PLoS ONE 2009, 4, e6862. [Google Scholar] [CrossRef] [PubMed]
- Tillakaratne, N.J.; Guu, J.J.; de Leon, R.D.; Bigbee, A.J.; London, N.J.; Zhong, H.; Ziegler, M.D.; Joynes, R.L.; Roy, R.R.; Edgerton, V.R. Functional recovery of stepping in rats after a complete neonatal spinal cord transection is not due to regrowth across the lesion site. Neuroscience 2010, 166, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- See, P.A.; de Leon, R.D. Robotic loading during treadmill training enhances locomotor recovery in rats spinally transected as neonates. J. Neurophysiol. 2013, 110, 760–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timoszyk, W.K.; Nessler, J.A.; Acosta, C.; Roy, R.R.; Edgerton, V.R.; Reinkensmeyer, D.J.; de Leon, R. Hindlimb loading determines stepping quantity and quality following spinal cord transection. Brain Res. 2005, 1050, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ji, S.R.; Wu, C.Y.; Fan, X.H.; Zhou, H.J.; Liu, G.L. Observation of locomotor functional recovery in adult complete spinal rats with bwstt using semiquantitative and qualitative methods. Spinal Cord 2007, 45, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Zdunowski, S.; Edgerton, V.R.; Roy, R.R.; Zhong, H.; Hsiao, I.; Lin, V.W. Improvement of gait patterns in step-trained, complete spinal cord-transected rats treated with a peripheral nerve graft and acidic fibroblast growth factor. Exp. Neurol. 2010, 224, 429–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Leon, R.D.; Acosta, C.N. Effect of robotic-assisted treadmill training and chronic quipazine treatment on hindlimb stepping in spinally transected rats. J. Neurotrauma 2006, 23, 1147–1163. [Google Scholar] [CrossRef] [PubMed]
- Moshonkina, T.R.; Gilerovich, E.G.; Fedorova, E.A.; Avelev, V.D.; Gerasimenko, Y.P.; Otellin, V.A. Morphofunctional basis for recovery of locomotor movements in rats with completely crossed spinal cord. Bull. Exp. Biol. Med. 2004, 138, 198–201. [Google Scholar] [CrossRef] [PubMed]
- Kubasak, M.D.; Jindrich, D.L.; Zhong, H.; Takeoka, A.; McFarland, K.C.; Munoz-Quiles, C.; Roy, R.R.; Edgerton, V.R.; Ramon-Cueto, A.; Phelps, P.E. Oeg implantation and step training enhance hindlimb-stepping ability in adult spinal transected rats. Brain 2008, 131, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Fouad, K.; Metz, G.A.; Merkler, D.; Dietz, V.; Schwab, M.E. Treadmill training in incomplete spinal cord injured rats. Behav. Brain Res. 2000, 115, 107–113. [Google Scholar] [CrossRef]
- Gomez-Pinilla, F.; Ying, Z.; Roy, R.R.; Molteni, R.; Edgerton, V.R. Voluntary exercise induces a bdnf-mediated mechanism that promotes neuroplasticity. J. Neurophysiol. 2002, 88, 2187–2195. [Google Scholar] [CrossRef] [PubMed]
- Skup, M.; Dwornik, A.; Macias, M.; Sulejczak, D.; Wiater, M.; Czarkowska-Bauch, J. Long-term locomotor training up-regulates trkb(fl) receptor-like proteins, brain-derived neurotrophic factor, and neurotrophin 4 with different topographies of expression in oligodendroglia and neurons in the spinal cord. Exp. Neurol. 2002, 176, 289–307. [Google Scholar] [CrossRef] [PubMed]
- Koliatsos, V.E.; Clatterbuck, R.E.; Winslow, J.W.; Cayouette, M.H.; Price, D.L. Evidence that brain-derived neurotrophic factor is a trophic factor for motor neurons in vivo. Neuron 1993, 10, 359–367. [Google Scholar] [CrossRef]
- Blum, R.; Konnerth, A. Neurotrophin-mediated rapid signaling in the central nervous system: Mechanisms and functions. Physiology 2005, 20, 70–78. [Google Scholar] [CrossRef] [PubMed]
- De Leon, R.D.; See, P.A.; Chow, C.H. Differential effects of low versus high amounts of weight supported treadmill training in spinally transected rats. J. Neurotrauma 2011, 28, 1021–1033. [Google Scholar] [CrossRef] [PubMed]
- Dupont-Versteegden, E.E.; Houle, J.D.; Dennis, R.A.; Zhang, J.; Knox, M.; Wagoner, G.; Peterson, C.A. Exercise-induced gene expression in soleus muscle is dependent on time after spinal cord injury in rats. Muscle Nerve 2004, 29, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, F.; Ying, Z.; Opazo, P.; Roy, R.R.; Edgerton, V.R. Differential regulation by exercise of bdnf and nt-3 in rat spinal cord and skeletal muscle. Eur. J. Neurosci. 2001, 13, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, K.J.; Gomez-Pinilla, F.; Crowe, M.J.; Ying, Z.; Basso, D.M. Three exercise paradigms differentially improve sensory recovery after spinal cord contusion in rats. Brain 2004, 127, 1403–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macias, M.; Nowicka, D.; Czupryn, A.; Sulejczak, D.; Skup, M.; Skangiel-Kramska, J.; Czarkowska-Bauch, J. Exercise-induced motor improvement after complete spinal cord transection and its relation to expression of brain-derived neurotrophic factor and presynaptic markers. BMC Neurosci. 2009, 10, 144. [Google Scholar] [CrossRef] [PubMed]
- Ying, Z.; Roy, R.R.; Edgerton, V.R.; Gomez-Pinilla, F. Voluntary exercise increases neurotrophin-3 and its receptor trkc in the spinal cord. Brain Res. 2003, 987, 93–99. [Google Scholar] [CrossRef]
- Ying, Z.; Roy, R.R.; Edgerton, V.R.; Gomez-Pinilla, F. Exercise restores levels of neurotrophins and synaptic plasticity following spinal cord injury. Exp. Neurol. 2005, 193, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Ying, Z.; Roy, R.R.; Zhong, H.; Zdunowski, S.; Edgerton, V.R.; Gomez-Pinilla, F. Bdnf-exercise interactions in the recovery of symmetrical stepping after a cervical hemisection in rats. Neuroscience 2008, 155, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Leech, K.A.; Hornby, T.G. High-intensity locomotor exercise increases brain-derived neurotrophic factor in individuals with incomplete spinal cord injury. J. Neurotrauma 2017, 34, 1240–1248. [Google Scholar] [CrossRef] [PubMed]
- Farinas, I.; Jones, K.R.; Backus, C.; Wang, X.Y.; Reichardt, L.F. Severe sensory and sympathetic deficits in mice lacking neurotrophin-3. Nature 1994, 369, 658–661. [Google Scholar] [CrossRef] [PubMed]
- Frisen, J.; Arvidsson, U.; Lindholm, T.; Fried, K.; Verge, V.M.; Cullheim, S.; Hokfelt, T.; Risling, M. Trkc expression in the injured rat spinal cord. Neuroreport 1993, 5, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Lohof, A.M.; Ip, N.Y.; Poo, M.M. Potentiation of developing neuromuscular synapses by the neurotrophins nt-3 and bdnf. Nature 1993, 363, 350–353. [Google Scholar] [CrossRef] [PubMed]
- Scarisbrick, I.A.; Isackson, P.J.; Windebank, A.J. Differential expression of brain-derived neurotrophic factor, neurotrophin-3, and neurotrophin-4/5 in the adult rat spinal cord: Regulation by the glutamate receptor agonist kainic acid. J. Neurosci. 1999, 19, 7757–7769. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xie, K.; Lu, B. Neurotrophins promote maturation of developing neuromuscular synapses. J. Neurosci. 1995, 15, 4796–4805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, H.; Liu, C.; Yang, M.; Li, H.; Zhang, F.; Zhang, W.; Zhang, J. Neurotrophic factor and trk signaling mechanisms underlying the promotion of motor recovery after acute spinal cord injury in rats. Exp. Ther. Med. 2017, 14, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Bimonte-Nelson, H.A.; Nelson, M.E.; Granholm, A.C. Progesterone counteracts estrogen-induced increases in neurotrophins in the aged female rat brain. Neuroreport 2004, 15, 2659–2663. [Google Scholar] [CrossRef] [PubMed]
- Gorgey, A.S.; Khalil, R.E.; Gill, R.; O’Brien, L.C.; Lavis, T.; Castillo, T.; Cifu, D.X.; Savas, J.; Khan, R.; Cardozo, C.; et al. Effects of testosterone and evoked resistance exercise after spinal cord injury (terex-sci): Study protocol for a randomised controlled trial. BMJ Open 2017, 7, e014125. [Google Scholar] [CrossRef] [PubMed]
- Yarrow, J.F.; Conover, C.F.; Purandare, A.V.; Bhakta, A.M.; Zheng, N.; Conrad, B.; Altman, M.K.; Franz, S.E.; Wronski, T.J.; Borst, S.E. Supraphysiological testosterone enanthate administration prevents bone loss and augments bone strength in gonadectomized male and female rats. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1213–E1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarrow, J.F.; Phillips, E.G.; Kok, H.J.; Conover, C.F.; Basset, T.E.; Vasconez, A.; Alerte, J.; Prock, H.; Flores, M.; Borst, S.E.; et al. Locomotor training with adjuvant testosterone promotes activity-mediated neuromuscular plasticity in spinal cord injured rats. Med. Sci. Sports Exerc. 2017, 49, 1038. [Google Scholar] [CrossRef]
- Wang, X.; Smith, G.I.; Patterson, B.W.; Reeds, D.N.; Kampelman, J.; Magkos, F.; Mittendorfer, B. Testosterone increases the muscle protein synthesis rate but does not affect very-low-density lipoprotein metabolism in obese premenopausal women. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E740–746. [Google Scholar] [CrossRef] [PubMed]
- Spinal Cord Injury (SCI) Facts and Figures at a Glance. Available online: https://www.nscisc.uab.edu/Public/Facts%202016.pdf (accessed on 25 May 2018).
- Calof, O.M.; Singh, A.B.; Lee, M.L.; Kenny, A.M.; Urban, R.J.; Tenover, J.L.; Bhasin, S. Adverse events associated with testosterone replacement in middle-aged and older men: A meta-analysis of randomized, placebo-controlled trials. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 1451–1457. [Google Scholar] [CrossRef] [PubMed]
- Basaria, S.; Coviello, A.D.; Travison, T.G.; Storer, T.W.; Farwell, W.R.; Jette, A.M.; Eder, R.; Tennstedt, S.; Ulloor, J.; Zhang, A.; et al. Adverse events associated with testosterone administration. N. Engl. J. Med. 2010, 363, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Finkle, W.D.; Greenland, S.; Ridgeway, G.K.; Adams, J.L.; Frasco, M.A.; Cook, M.B.; Fraumeni, J.F., Jr.; Hoover, R.N. Increased risk of non-fatal myocardial infarction following testosterone therapy prescription in men. PLoS ONE 2014, 9, e85805. [Google Scholar] [CrossRef] [PubMed]
- Vigen, R.; O’Donnell, C.I.; Baron, A.E.; Grunwald, G.K.; Maddox, T.M.; Bradley, S.M.; Barqawi, A.; Woning, G.; Wierman, M.E.; Plomondon, M.E.; et al. Association of testosterone therapy with mortality, myocardial infarction, and stroke in men with low testosterone levels. JAMA 2013, 310, 1829–1836. [Google Scholar] [CrossRef] [PubMed]
- Morgentaler, A.; Lunenfeld, B. Testosterone and cardiovascular risk: World’s experts take unprecedented action to correct misinformation. Aging Male 2014, 17, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Traish, A.M.; Guay, A.T.; Morgentaler, A. Death by testosterone? We think not! J. Sex. Med. 2014, 11, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Borst, S.E.; Shuster, J.J.; Zou, B.; Ye, F.; Jia, H.; Wokhlu, A.; Yarrow, J.F. Cardiovascular risks and elevation of serum dht vary by route of testosterone administration: A systematic review and meta-analysis. BMC Med. 2014, 12, 211. [Google Scholar] [CrossRef] [PubMed]
- Onasanya, O.; Iyer, G.; Lucas, E.; Lin, D.; Singh, S.; Alexander, G.C. Association between exogenous testosterone and cardiovascular events: An overview of systematic reviews. Lancet Diabetes Endocrinol. 2016, 4, 943–956. [Google Scholar] [CrossRef]
- Sharma, R.; Oni, O.A.; Gupta, K.; Chen, G.; Sharma, M.; Dawn, B.; Parashara, D.; Savin, V.J.; Ambrose, J.A.; Barua, R.S. Normalization of testosterone level is associated with reduced incidence of myocardial infarction and mortality in men. Eur. Heart J. 2015, 36, 2706–2715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Fountaine, M.F.; Wecht, J.M.; Cirnigliaro, C.M.; Kirshblum, S.C.; Spungen, A.M.; Bauman, W.A. Testosterone replacement therapy improves qtavi in hypogonadal men with spinal cord injury. Neuroendocrinology 2013, 97, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Hamid, R.; Averbeck, M.A.; Chiang, H.; Garcia, A.; Al Mousa, R.T.; Oh, S.J.; Patel, A.; Plata, M.; Del Popolo, G. Epidemiology and pathophysiology of neurogenic bladder after spinal cord injury. World J. Urol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Page, S.T.; Amory, J.K.; Bowman, F.D.; Anawalt, B.D.; Matsumoto, A.M.; Bremner, W.J.; Tenover, J.L. Exogenous testosterone (T) alone or with finasteride increases physical performance, grip strength, and lean body mass in older men with low serum T. J. Clin. Endocrinol. Metab. 2005, 90, 1502–1510. [Google Scholar] [CrossRef] [PubMed]
- Amory, J.K.; Watts, N.B.; Easley, K.A.; Sutton, P.R.; Anawalt, B.D.; Matsumoto, A.M.; Bremner, W.J.; Tenover, J.L. Exogenous testosterone or testosterone with finasteride increases bone mineral density in older men with low serum testosterone. J. Clin. Endocrinol. Metab. 2004, 89, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Clinicaltrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02248701 (accessed on 25 May 2018).
- Kaplan, A.L.; Hu, J.C. Use of testosterone replacement therapy in the united states and its effect on subsequent prostate cancer outcomes. Urology 2013, 82, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Sullivan, C.T.; McCoy, S.C.; Yarrow, J.F.; Morrow, M.; Borst, S.E. Review of health risks of low testosterone and testosterone administration. World J. Clin. Cases 2015, 3, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Cole, A.P.; Hanske, J.; Jiang, W.; Kwon, N.K.; Lipsitz, S.R.; Kathrins, M.; Learn, P.A.; Sun, M.; Haider, A.H.; Basaria, S.; et al. Impact of testosterone replacement therapy on thromboembolism, heart disease and obstructive sleep apnoea in men. BJU Int. 2018, 121, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Chiodo, A.E.; Sitrin, R.G.; Bauman, K.A. Sleep disordered breathing in spinal cord injury: A systematic review. J. Spinal Cord Med. 2016, 39, 374–382. [Google Scholar] [CrossRef] [PubMed]


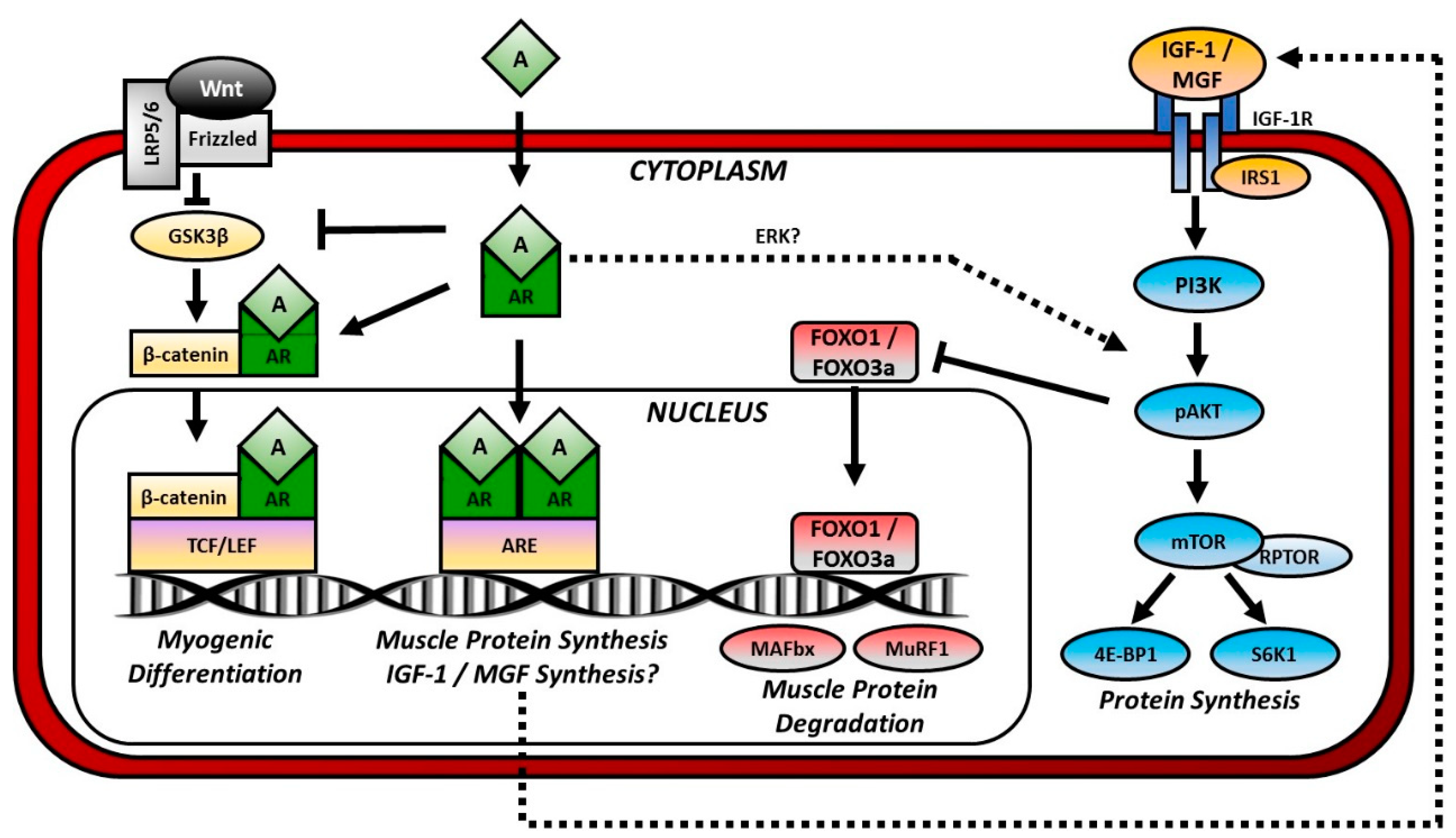{kind=link}
| Study | Intervention | Duration | Population | N | Outcomes |
|---|---|---|---|---|---|
| Studies Assessing Gait | |||||
| Dobkin et al., 2007 [185] | Manual-assisted BWSTT vs. OG training | 12 wk | 38 ASIA B, 107 ASIA C&D | 145 | BWSTT increased walking speed from ~0.40 m/s at 2 weeks post-entry to 0.85 m/s for ASIA C&D. OG training increased walking speed from ~0.50 at 2 weeks post-entry to 0.84 m/s for ASIA C&D. Poor walking outcomes were observed for ASIA B participants. |
| Field-Fote et al., 2001 [167] | BWSTT with ES | 3 days/wk, 12 wk | ASIA C | 19 | Walking speed increased from 0.12 to 0.21 m/s. a |
| Alcobendas-Maestro et al., 2012 [218] | Robotic-assisted BWSTT vs. OG training | 3–6 mo | ASIA C&D | 48 M/32F | Robotic-assisted BWSTT and OG training walking speed remained the same from baseline to post-training. Robotic-assisted BWSTT increased 6 min walk distance from 91 to 169 m. a |
| Alexeeva et al., 2011 [179] | BWS training on a fixed track vs. BWSTT vs. Comprehensive PT | 3 days/wk 13 wk | ASIA C&D | 30 M/5 F | BWS on a fixed track increased walking speed from 0.33 to 0.44 m/s. a BWSTT increased walking speed from 0.30 to 0.46 m/s. a PT increased walking speed from 0.41 to 0.51 m/s. a |
| Duffell et al., 2015 [202] | Robotic-assisted BWSTT | 3 days/wk, 4 wk | ASIA C&D | 19 M/7 F | Walking speed increased from ~0.55 to ~0.58 m/s. a |
| Esclarin-Ruz et al., 2014 [219] | Robotic-assisted BWSTT + OG training (LKOGT) vs. Conventional OG training (OGT) | 5 days/wk, 8 wk | ASIA C&D | 59 M/24 F | LKOGT increased walking speed from 0.48 to 0.54 m/s and 6 min walk distance from 122 to 187 m b in participants with upper motor neuron injury. LKOGT increased walking speed from 0.24 to 0.46 m/s and 6 min walk distance from 82 to 157 m b in participants with lower motor neuron injury. OGT increased walking speed from 0.36 to 0.39 m/s and 6 min walk distance from 93 to 119 m in participants with upper motor neuron injury. OGT increased walking speed from 0.28 to 0.45 m/s and 6 min walk distance from 94 to 145 m in participants with lower motor neuron injury. |
| Field-Fote et al., 2005 [220] | Manual-assisted BWSTT (TM) vs. BWSTT + ES (TS) vs. BWS OG training + ES (OGS) vs. Robotic-assisted BWSTT (LR) | 5 days/wk 12 wk | ASIA C&D | 22 M/5 F | TM increased walking speed from ~0.07 to ~0.10. TS increased walking speed from ~0.12 to ~0.16 m/s. a OG increased walking speed from ~0.14 to ~0.19 m/s. a LR increased walking speed from ~0.09 to ~0.11. |
| Field-Fote & Roach 2011 [186] | Manual-assisted BWSTT (TM) vs. BWSTT + ES (TS) vs. BWS OG training +ES (OGS) vs. Robotic-assisted BWSTT (LR) | 5 days/wk, 12 wk | ASIA C&D | 52 M/12 F | TM increased walk speed from 0.17 to 0.22 m/s a and 2 min distance from 22.1 to 23.0 m. TS increased walk speed from 0.18 to 0.23 m/s a and 2 min distance from 20.6 to 24.4 m. a OGS increased walk speed from 0.19 to 0.28 m/s a and 2 min distance from 24.0 to 38.3 m. a,b LR increased walk speed from 0.17 to 0.18 m/s and 2 min distance from 16.8 to 17.9 m. |
| Gorassini et al., 2009 [169] | Manual-assisted BWSTT | 5 days/wk, 14 wk | ASIA C&D | 14 M/3 F | In 9 responders, walking speed increased from 0.31 to 0.55 m/s. a In 8 non-responders, there was no change in walking speed. |
| Harkema et al., 2012 [170] | Manual-assisted BWSTT | 3 day/wk, 4 to 92 wks | ASIA C&D | 148M/48 F | Walking speed increased from 0.31 to 0.51 m/s a and 6 min walk distance increased from 91 to 154 m. a |
| Kapadia et al., 2014 [221] | Robotic-assisted BWSTT + ES | 3 days/wk, 16 wk | ASIA C&D | 13 M/3 F | Walking speed increased from 0.23 to 0.28 m/s and 6 min walk distance increased from 187.9 to 217.1 m. a |
| Knikou 2013 [214] | Manual-assisted BWSTT | 5 days/wk, 1.5–3.5 mo | ASIA C&D | 9 M/3 F | 6 min walk distance increased from 36.25 to 39.05 m for ASIA C and from 252 to 279.5 m for ASIA D participants. |
| Krishnan et al., 2016 [171] | Robotic-assisted BWSTT | 3 day/wk, 4 wk | ASIA C&D | 8 M/8 F | Median walking speed increased from 0.58 to 0.66 m/s. a 6 min walking distance did not change. |
| Labruyere et al., 2014 [222] | Robotic-assisted BWSTT | 4 days/wk, 4 wk | ASIA C&D | 5 M/4 F | Walking speed increased from 0.62 to 0.66 m/s. |
| Lam et al., 2015 [223] | Robotic-assisted BWSTT with resistance (LR) vs. Robotic-assisted BWSTT only (LO) | 3 days/wk, 12 wk | ASIA C&D | 9 M/6 F | LR increased walking speed from 0.29 to 0.40 m/s. LO increased walking speed from 0.33 to 0.44 m/s. |
| Lucarelli et al., 2011 [187] | BWSTT vs. Conventional gait training | 2 days/wk, 12 wk | ASIA C&D | 20 M/10 F | BWSTT increased walking speed from 0.85 to 1.25 m/s a as well as increased cadence, distance, step length and swing phase. Conventional training did not improve gait quality or speed. |
| Morrison et al., 2018 [224] | Manual-assisted BWSTT | 120 sessions | ASIA C&D | 49 M/20 F | Median walking speed increased by 0.25 m/s. a Median 6 min walk distance increased by 66 m. a |
| Niu et al., 2014 [225] | Robotic-assisted BWSTT | 3 days/wk, 4 wk | ASIA C&D | 27 M/13 F | Walking speed increased in the low-walking capacity group from 0.12 to 0.15 m/s and in the high-walking capacity group from 0.84 to 0.97 m/s. 6 min walk distance did not change. |
| Nooijen et al., 2009 [172] | Manual-assisted BWSTT vs. Manual-assisted BWSTT with ES vs. OG training with ES vs. Robotic-assisted BWSTT | 4 days/wk, 12 wk | ASIA C&D | 40 M/11 F | All therapies led to small improvements in gait quality (increased cadence and step length) with no differences among groups. |
| Potsans et al., 2004 [188] | BWSTT with ES Cross-over design: treatment-control (AB); control-treatment (BA) | 5 days/wk, 4 wk | ASIA C&D | 12 M/2 F | In AB group, walking speed increased 0.23 m/s and 6 min walk distance increased 72.2 m. In BA group, walking speed increased 0.17 m/s and 6 min walk distance increased 63.8 m. |
| Thomas et al., 2005 [180] | Manual-assisted BWSTT | 4 days/wk, 16 wk | ASIA C&D | 8 M/2 F | Walking speed increased from 0.15 to 0.53 m/s a and 6 min walk distance increased from 34.2 to 167.6 m. |
| Varoqui et al., 2014 [203] | Robotic-assisted BWSTT | 3 days/wk, 4 wk | ASIA C&D | 14 M/ 1 F | Walking speed increased from 0.56 to 0.64 m/s a and 6 min walk distance did not change (207 to 209 m). |
| Winchester et al., 2005 [226] | Robotic-assisted BWSTT | 3 days/wk 12 wk | ASIA C&D | 4 M | Walking speed increased for 3 participants from 0.0 to 0.11, 0.0 to 0.81, 0.24 to 0.62 m/s and one remained unable to ambulate. |
| Wirz et al., 2005 [175] | Robotic-assisted BWSTT | 4 days/wk, 8 wk | ASIA C&D | 18 M/2 F | Walking speed increased from ~0.37 to ~.48 m/s a and 6 min walk distance 120 to 160 m. a |
| Wu et al., 2012 [227] | 4 wk Robotic-assistance BWSTT + 4 wk Robotic-resistance BWSTT | 3 days/wk, 8 wk | ASIA D | 8 M/2 F | Walking speed increased from 0.67 to 0.76 m/s a and 6 min walk distance increased from 223 to 247 m. Step length and cadence increased. a |
| Studies Assessing Muscle | |||||
| Duffell et al., 2015 [202] | Robotic-assisted BWSTT | 3 days/wk, 4 wk | ASIA C&D | 19 M/7 F | Maximal isometric ankle dorsiflexion torque increased from 12.3 to 13.2 Nm, a but plantar flexion torque did not change (28.1 vs. 28.4 Nm). |
| Galen et al., 2014 [228] | Robotic-assisted BWSTT | 5 days/wk, 6 wk | ASIA C&D | 14 M/4 F | Percent peak torque increased 68% for hip flexion, 54% for hip extension, 93% for knee flexion and 71% for knee extension. |
| Gorassini et al., 2009 [169] | Manual-assisted BWSTT | 5 days/wk, 14 wk | ASIA C&D | 14 M/3 F | In 9 responders, peak electromyography activity increased from 67 to 135 μV in the tibialis anterior muscle and 36 to 50 μV in the hamstrings. In 8 non-responders, there was no change. |
| Jayaraman et al., 2008 [201] | Manual-assisted BWSTT | 5 days/wk, 9 wk | ASIA C&D | 4 M/1 F | Isometric knee extension strength increased 21%. Isometric plantar flexion strength increased 44%. Knee extension and plantar flexion voluntary muscle activation improved. Maximal CSA of the plantar flexors increased and 15%. |
| Krishnan et al., 2016 [171] | Robotic-assisted BWSTT | 3 day/wk, 4 wk | ASIA C&D | 8 M/8 F | BWSTT increased isometric ankle dorsiflexion by 20% and ankle plantar flexion by 22%. |
| Thomas et al., 2005 [180] | Manual-assisted BWSTT | 4 days/wk, 16 wk | ASIA C&D | 8 M/2 F | Peak electromyography activity averaged from four lower limb muscles increased during treadmill walking from 82.4 to 137.1 μV. |
| Varoqui et al., 2014 [203] | Robotic-assisted BWSTT | 3 days/wk, 4 wk | ASIA C&D | 14 M/1 F | Ankle dorsiflexion torque increased from 26.8 to 29.1 Nma and ankle plantar flexion torque increased from 10.9 to 13.5 Nm. a |
| Study | Sex/Age | Injury Level | Start of Training | Training Duration | BBB wk 1 | BBB End | Gait Outcome | Muscle/Electrophysiology |
|---|---|---|---|---|---|---|---|---|
| Studies Assessing BWSTT after Spinal Hemisection | ||||||||
| Maier et al., 2009 [229] | F/A | T8 | 1 wk | 8 wk | ~12 | N/R | Stepping ↑ | N/R |
| Battistuzzo et al., 2016 [230] | M/A | T10 | 1 wk | 9 wk | N/R | N/R | Kinematics ↑ | N/R |
| Battistuzzo et al., 2017 [231] | M/A | T10 | 1 wk | 9 wk | N/R | N/R | N/R | G fCSA ↑ |
| Shah et al., 2013 [232] | N/R/N/R | T10 | 5 days | 2.5 wk | N/R | N/R | Stepping ↑ | N/R |
| Goldshmit et al., 2008 [212] | N/R/A | T12 | 1 wk | 4 wk | ~3 | 11T/6C | Kinematics ↑ | G-S fCSA ↑ |
| Studies Assessing BWSTT after Mild Contusion SCI | ||||||||
| Nessler et al., 2006 [233] | F/Y | T9 | 1 wk | 12 wk | 13 | 14T/No C | N/R | N/R |
| Oh et al., 2009 [234] | M/Y | T9–10 | 1 wk | 4wk | ~4 | 13T/8C | N/R | N/R |
| Studies Assessing BWSTT after Moderate Contusion SCI | ||||||||
| Stevens et al., 2006 [197] | F/Y | T8 | 1 wk | 1 wk | ~5 | 10T/6C | N/R | Sol F ↑/fCSA ↑ |
| Liu et al., 2008 [199] | F/Y | T8 | 8 days | 12 wk | 3–7 | 15T/11C | N/R | G-S CSA ↑ |
| Nessler et al., 2006 [233] | F/Y | T9 | 1 wk | 12 wk | 9 | 11T/No C | N/R | N/R |
| Shin et al., 2014 [235] | F/Y | T9 | 1 wk | 8 wk | ~6 | ~13T/~10C | N/R | N/R |
| Wang et al., 2015 [29] | F/Y | T9 | 1 wk | 16 wk | ~7 | ~13T/~11C | N/R | N/R |
| Singh et al., 2011 [200] | F/Y | T9–10 | 1 wk | 8 wk | ~5 | ~8T/~9C | N/R | mass/bw ↑ |
| Bose et al., 2012 [198] | F/A | T8 | 1 wk | 12 wk | ~3 | ~15T/~11C | Stepping ↑ | N/R |
| Multon et al., 2003 [236] | F/A | T9 | 2–4 days | 12 wk | ~2 | 10T/8C | N/R | N/R |
| Wu et al., 2016 [237] | F/A | T10 | 1 wk | 4 wk | ~5 | ~13T/~9C | N/R | N/R |
| Foret et al., 2010 [238] | F/A | T10 | 1 day | 4 wk | ~2 | ~9T/~7C | N/R | N/R |
| Ward et al., 2014 [239] | M/Y | T8 | 2 wk | 12 wk | ~7 | ~12T/~9C | N/R | Sol EMG ↑ |
| Park et al., 2010 [240] | M/Y | T10 | 3 days | 25 days | ~2 | 11.5 | N/R | N/R |
| Liu et al., 2017 [241] | N/R/N/R | T9 | 1 wk | 2 wk | N/R | N/R | Stepping ↑ | N/R |
| Studies Assessing BWSTT after Severe Contusion SCI | ||||||||
| Hayashibe et al., 2016 [242] | F/Y | T8–9 | 1 wk | 4 wk | <1 | 16T/10C | N/R | N/R |
| Heng et al., 2009 [243] | F/Y | T9 | 42 days | 8 wk | N/R | N/R | Stepping ↑ | N/R |
| Nessler et al., 2006 [233] | F/Y | T9 | 1 wk | 12 wk | 4.5 | ~9T/No C | N/R | N/R |
| Shinozaki et al., 2016 [244] | F/Y | T10 | 6 wk | 8 wk | ~1 | 4T/3C | N/R | N/R |
| Robert et al., 2010 [245] | F/A | T7–8 | 2 wk | 2 wk | ~3 | 4T/3.5C | N/R | N/R |
| Ichiyama et al., 2009 [246] | F/A | T10 | 30 days | 8 wk | N/R | N/R | Gait not improved | N/R |
| Studies Assessing BWSTT after Spinal Transection | ||||||||
| Petruska et al., 2007 [213] | F/N | T6–8 | 16 days | 6 wk | N/R | N/R | Stepping ↑ | G EPSP ↑ |
| Tillakaratne et al., 2010 [247] | F/N | T7–8 | 26 days | 8 wk | N/R | N/R | Stepping ↑ | N/R |
| See et al., 2013 [248] | F/N | T8–9 | 3 wk | 4 wk | N/R | N/R | Kinematics ↑ | N/R |
| Timoszyk et al., 2005 [249] | F/N | N/R | 64 days | 40 days | N/R | N/R | Gait not improved | N/R |
| Zhang et al., 2007 [250] | F/Y | T8 | 5 days | 40 days | <1 | ~6.5T/~2C | N/R | N/R |
| Lee et al., 2010 [251] | F/Y | T8 | 3 wk | 42 wk | N/R | N/R | Gait not improved | N/R |
| De Leon et al., 2006 [252] | F/Y | T9 | 3 wk | 16 wk | N/R | N/R | Kinematics ↑ | N/R |
| Moshonkina et al., 2004 [253] | F/Y | T9 | 1 day | 9 wk | N/R | N/R | Kinematics ↑ | N/R |
| Kubasak et al., 2008 [254] | F/N/R | T9 | 4 wk | 20 wk | N/R | N/R | Gait not improved | N/R |
| Fouad et al., 2000 [255] | B/A | T8 | 3 days | 5 wk | ~9 | 14T/14C | Kinematics ↑ | N/R |
| Ihla et al., 2011 [196] | M/A | T8–9 | 6 days | 9 wk | N/R | N/R | N/R | Sol fCSA ↑ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otzel, D.M.; Lee, J.; Ye, F.; Borst, S.E.; Yarrow, J.F. Activity-Based Physical Rehabilitation with Adjuvant Testosterone to Promote Neuromuscular Recovery after Spinal Cord Injury. Int. J. Mol. Sci. 2018, 19, 1701. https://doi.org/10.3390/ijms19061701
Otzel DM, Lee J, Ye F, Borst SE, Yarrow JF. Activity-Based Physical Rehabilitation with Adjuvant Testosterone to Promote Neuromuscular Recovery after Spinal Cord Injury. International Journal of Molecular Sciences. 2018; 19(6):1701. https://doi.org/10.3390/ijms19061701
Chicago/Turabian StyleOtzel, Dana M., Jimmy Lee, Fan Ye, Stephen E. Borst, and Joshua F. Yarrow. 2018. "Activity-Based Physical Rehabilitation with Adjuvant Testosterone to Promote Neuromuscular Recovery after Spinal Cord Injury" International Journal of Molecular Sciences 19, no. 6: 1701. https://doi.org/10.3390/ijms19061701