Unraveling the Chloroplast Genomes of Two Prosopis Species to Identify Its Genomic Information, Comparative Analyses and Phylogenetic Relationship


Abstract
:1. Introduction
2. Results
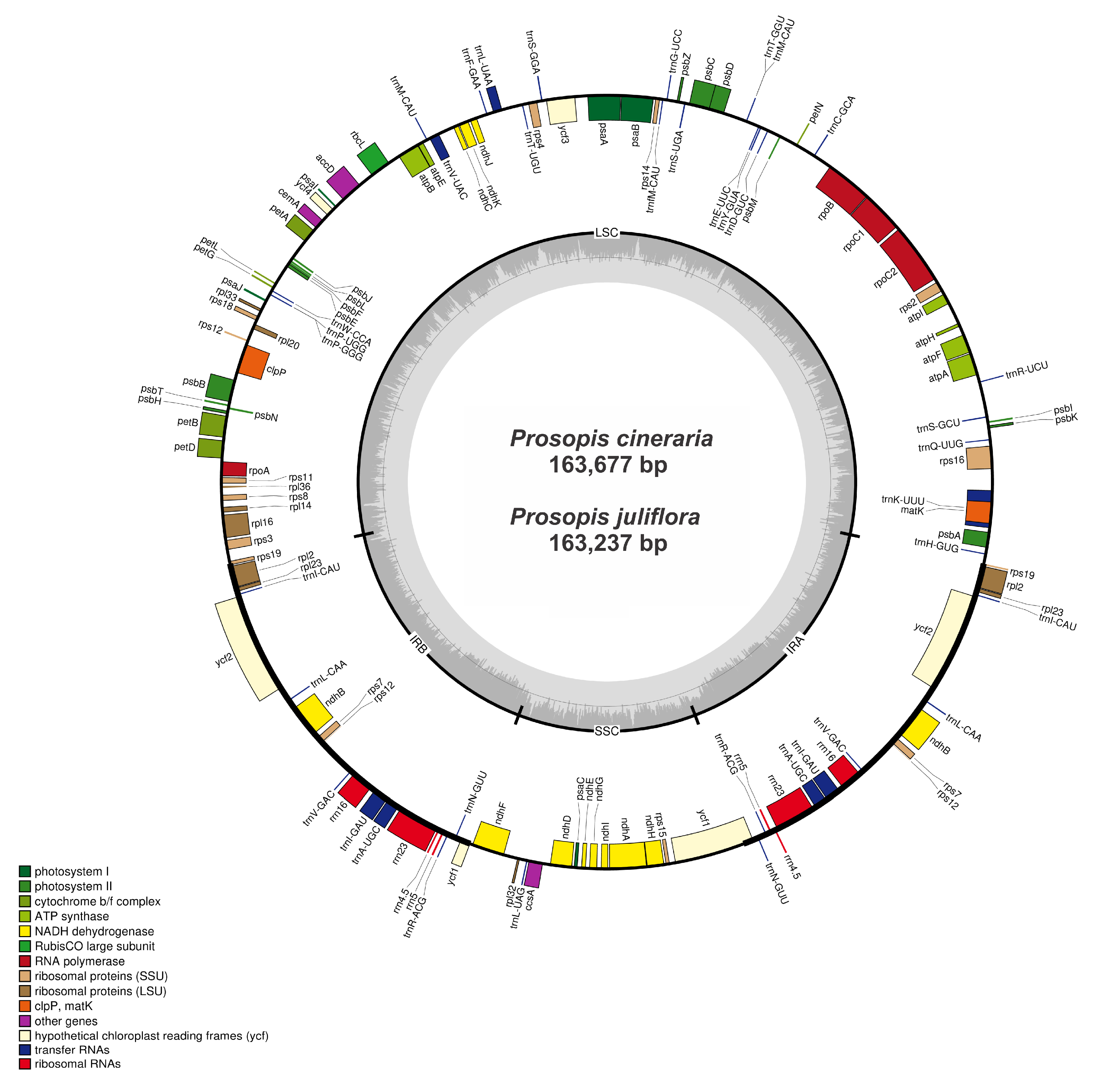
2.1. General Features and Organization of Chloroplast Genomes
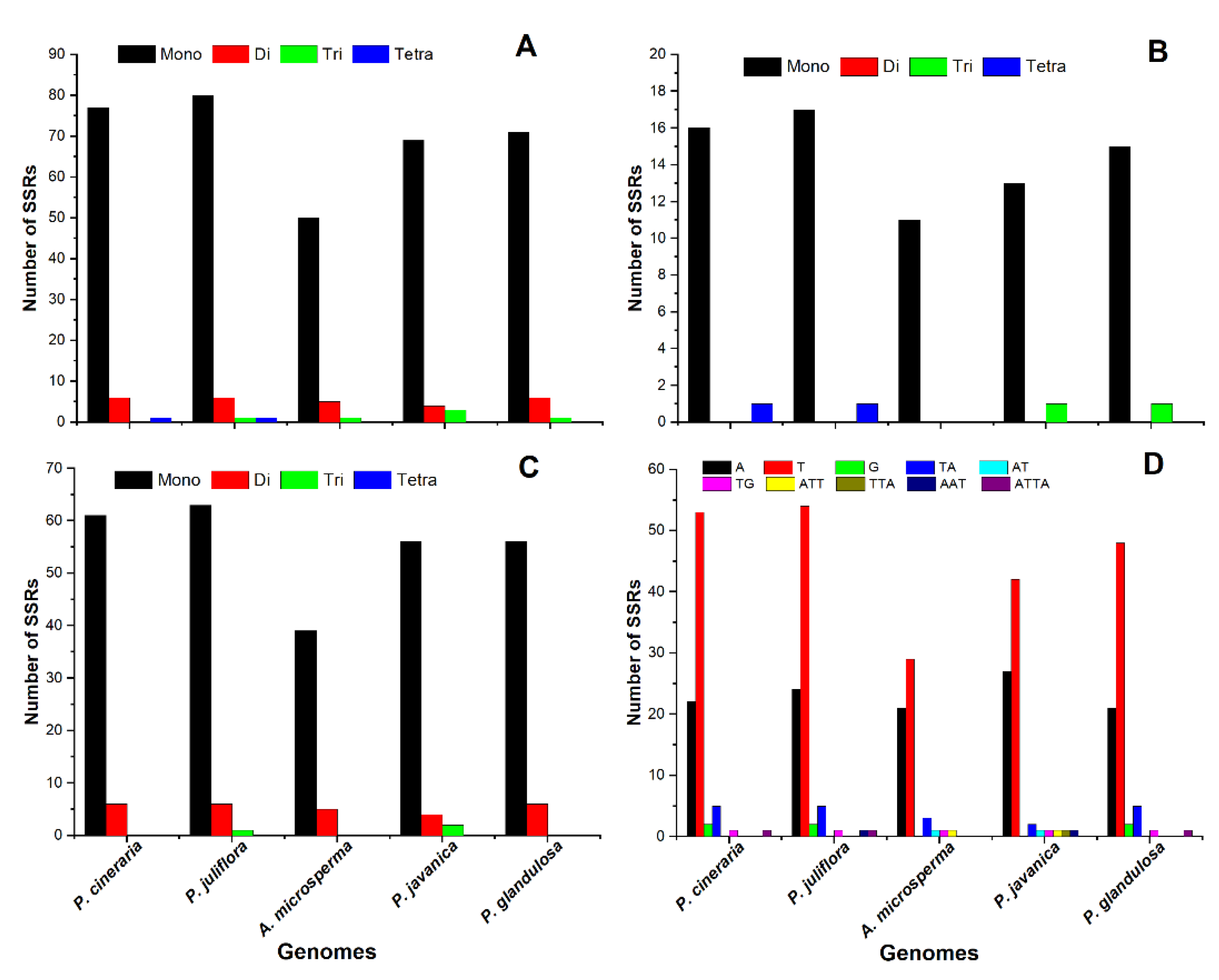
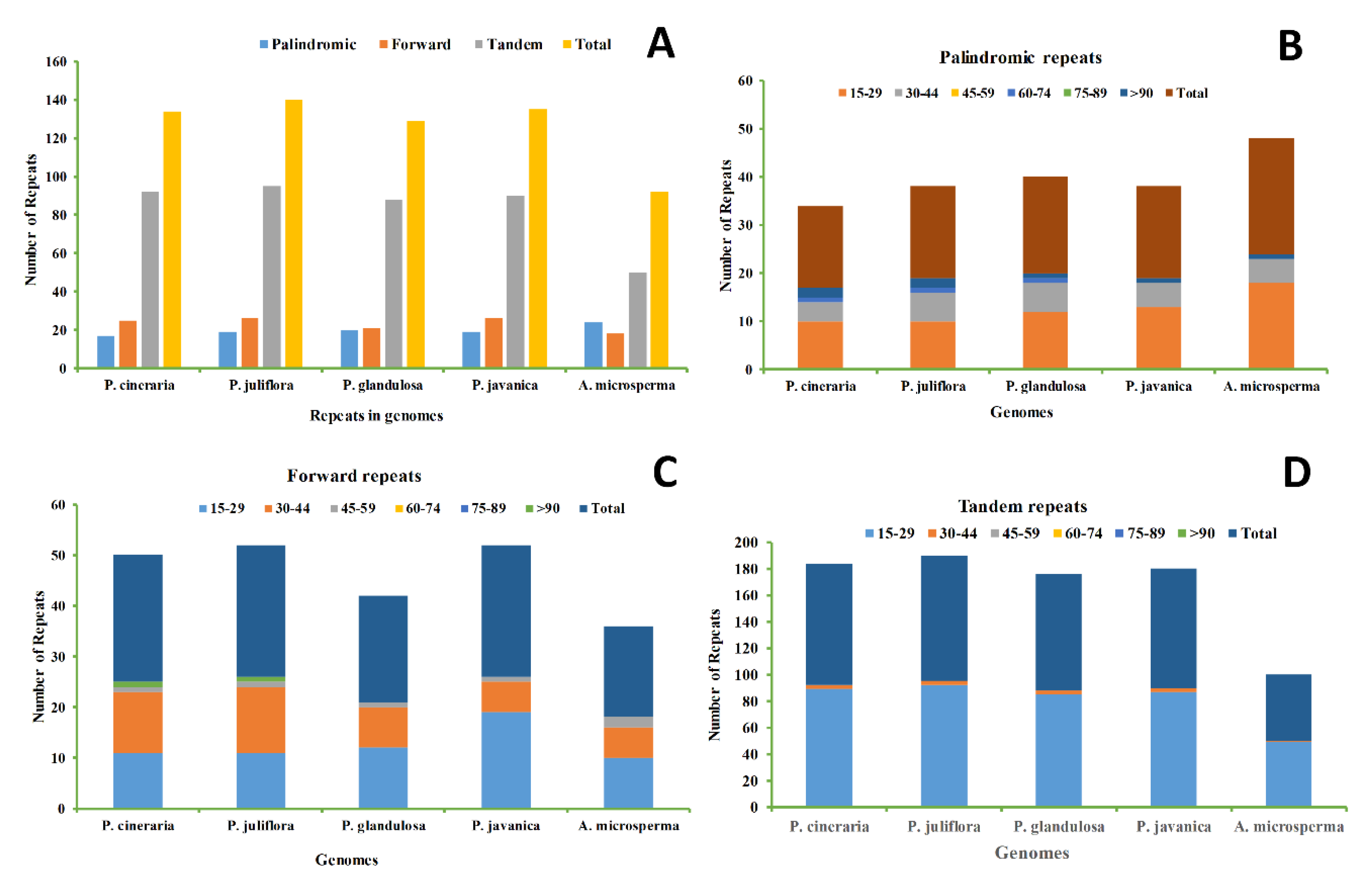
2.2. SSR Analysis and Repeats, an Insight into the Genome
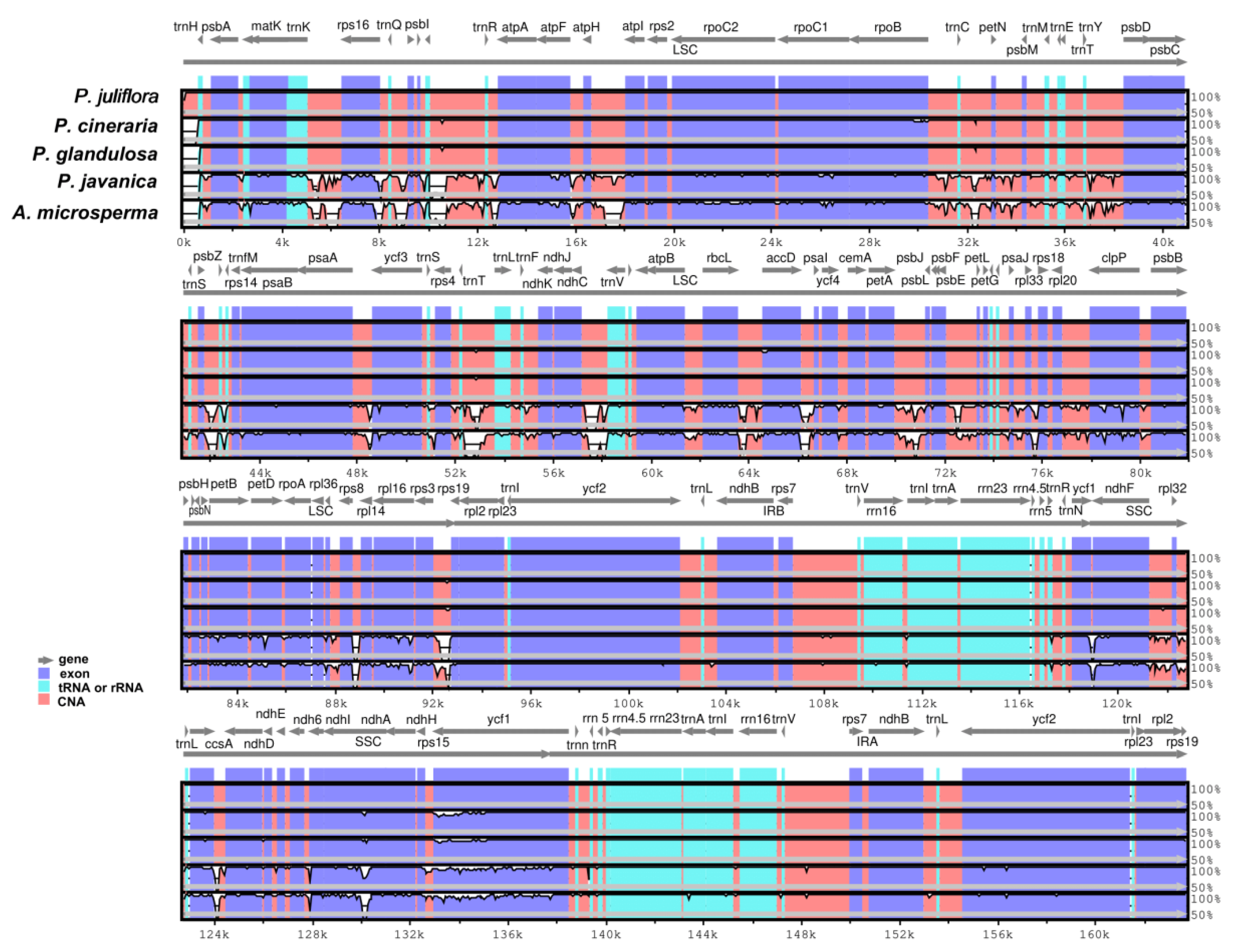
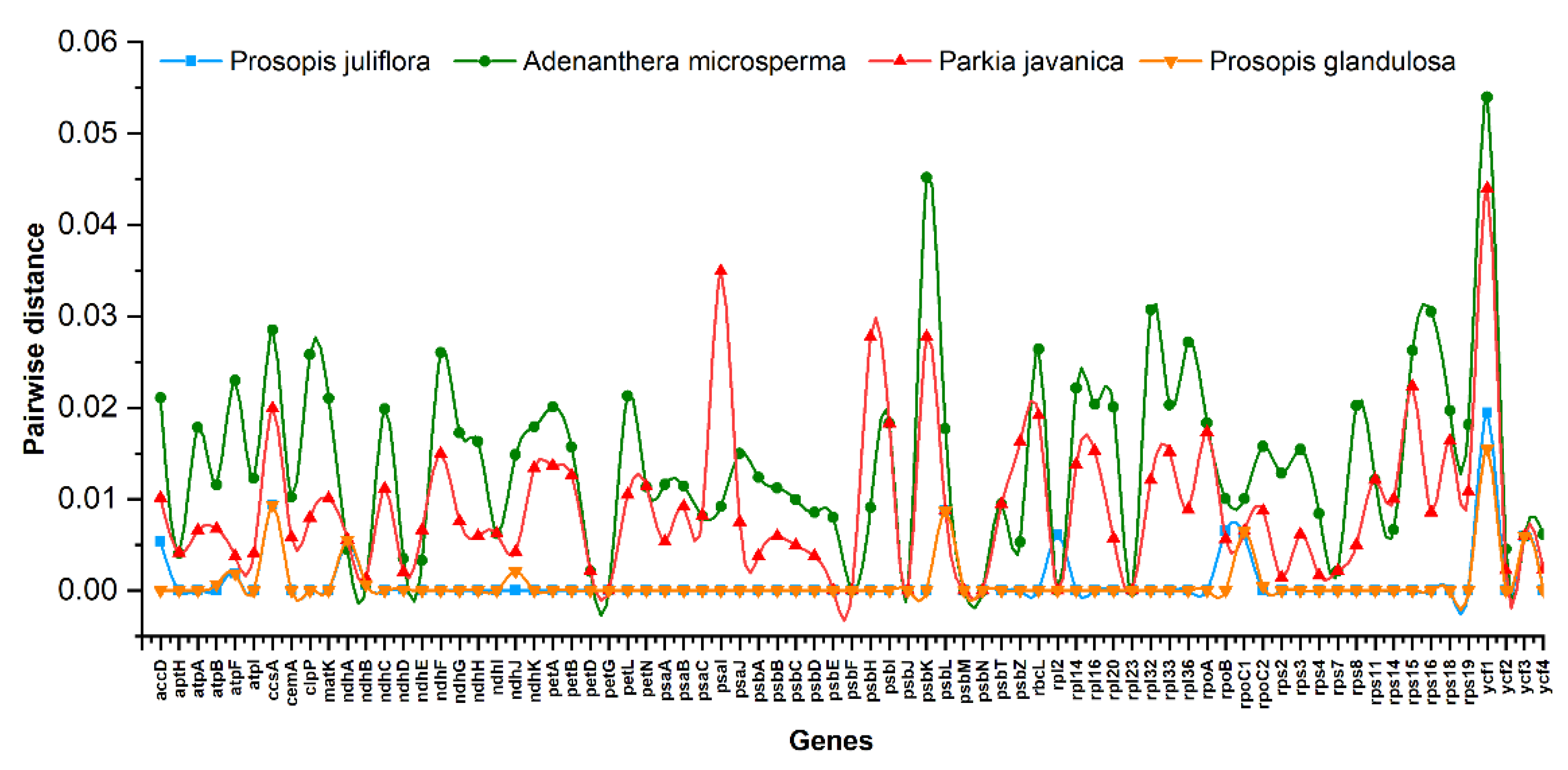
2.3. Comparative Analysis and Sequence Divergence Analyses
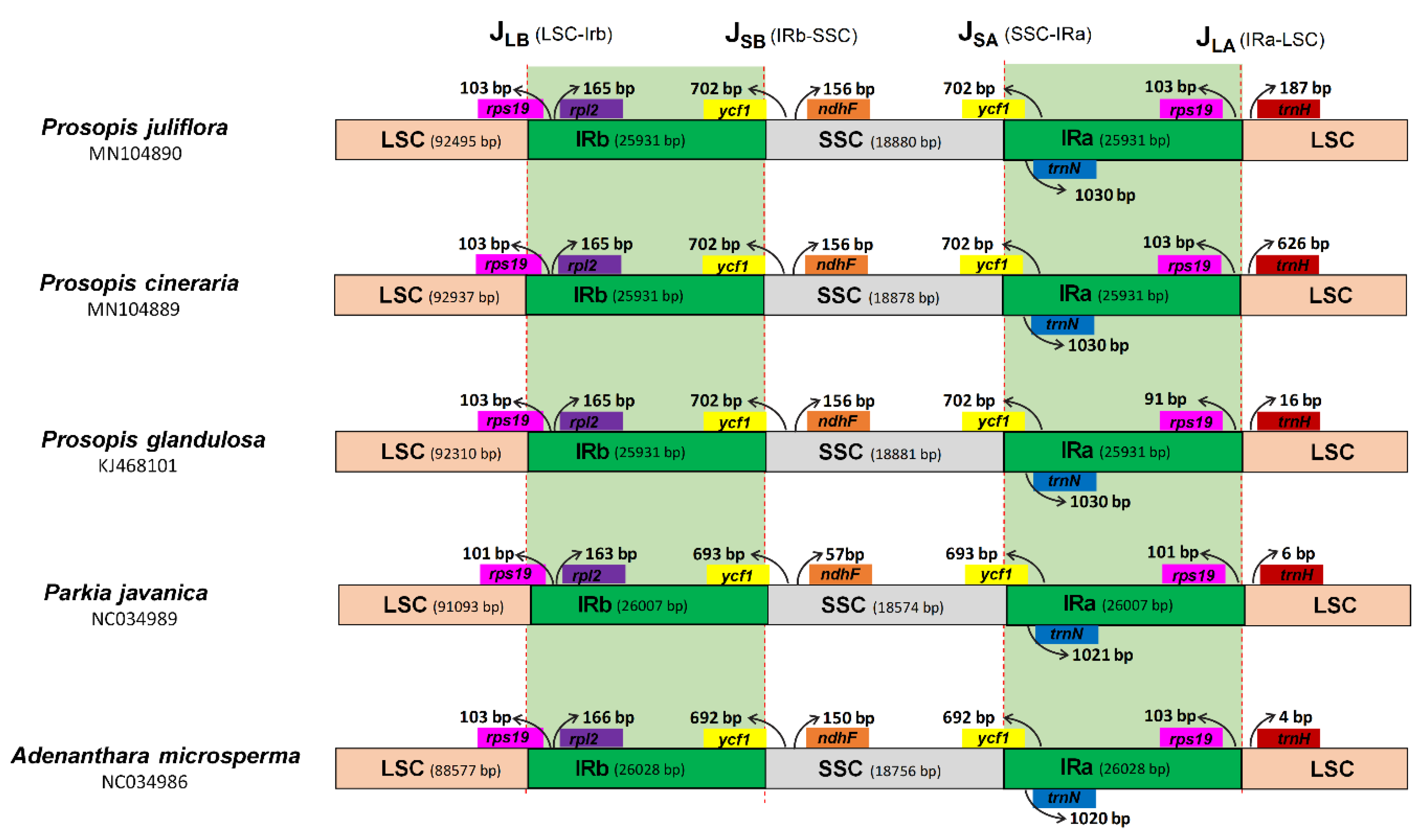
2.4. Boundaries between Inverted Repeat and Single Copy Regions
2.5. Phylogenetic Relationships
3. Discussion
4. Methodology
4.1. Sample Collection
4.2. DNA Extraction and Genome Sequencing
4.3. Chloroplast Reference-Based Genome Assembly
4.4. Prosopis Species Genome Annotation
4.5. Repeat Identification
4.6. Chloroplast Genome Divergence and Phylogenetic Relationship
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BI | Bayesian inference |
| IR | Inverted repeat |
| LSC | Large single copy |
| ML | maximum likelihood |
| MP | Maximum parsimony |
| NGS | Next generation sequencing |
| rRNA | Ribosomal RNA |
| SSC | Short single copy |
| SSRs | Simple sequence repeats |
| tRNA | Transfer RNA |
References
- Burkart, A. A monograph of the genus Prosopis (Leguminosae subfam. Mimosoideae). J. Arnold Arbor. 1976, 57, 450–525. [Google Scholar]
- Shackleton, R.T.; Le Maitre, D.C.; Pasiecznik, N.M.; Richardson, D.M. Prosopis: A global assessment of the biogeography, benefits, impacts and management of one of the world’s worst woody invasive plant taxa. AoB Plants 2014, 6, plu027. [Google Scholar] [CrossRef] [PubMed]
- Prado, D.E.; Gibbs, P.E. Patterns of Species Distributions in the Dry Seasonal Forests of South America. Ann. Mo. Bot. Gard. 1993, 80, 902. [Google Scholar] [CrossRef]
- Bessega, C.; Pometti, C.L.; Ewens, M.; Saidman, B.O.; Vilardi, J.C. Strategies for conservation for disturbed Prosopis alba (Leguminosae, Mimosoidae) forests based on mating system and pollen dispersal parameters. Tree Genet. Genomes 2011, 8, 277–288. [Google Scholar] [CrossRef]
- Bessega, C.; Ferreyra, L.; Julio, N.; Montoya, S.; Saidman, B.; Vilardi, J.C. Mating system parameters in species of genus Prosopis (Leguminosae). Hered. 2000, 132, 19–27. [Google Scholar] [CrossRef]
- Pasiecznik, N.M.; Felker, P.; Harris, P.J.; Harsh, L.; Cruz, G.; Tewari, J.; Cadoret, K.; Maldonado, L.J. The’Prosopis Juliflora’-’Prosopis Pallida’Complex: A Monograph; HDRA: Coventry, UK, 2001; Volume 172. [Google Scholar]
- Rejmánek, M.; Richardson, D.M. Trees and shrubs as invasive alien species - 2013 update of the global database. Divers. Distrib. 2013, 19, 1093–1094. [Google Scholar] [CrossRef]
- Bhandari, M. Flora of the Indian Desert; MPS Repros: Jodhpur, India, 1990. [Google Scholar]
- Felker, P.; Bandurski, R.S. Uses and potential uses of leguminous trees for minimal energy input agriculture. Econ. Bot. 1979, 33, 172–184. [Google Scholar] [CrossRef]
- Felker, P. Mesquite: An all-purpose leguminous arid land tree. New Agric. Crop. 2019, 38, 89–132. [Google Scholar]
- Sawal, R.K.; Ratan, R.; Yadav, S.B.S. Mesquite (Prosopis juliflora) Pods as a Feed Resource for Livestock - A Review -. Asian-Australasian J. Anim. Sci. 2004, 17, 719–725. [Google Scholar] [CrossRef]
- Ramírez, L.; De La Vega, A.; Razkin, N.; Luna, V.; Harris, P.J. Analysis of the relationships between species of the genus Prosopis revealed by the use of molecular markers. Agron. 1999, 19, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Landeras, G.; Alfonso, M.; Pasiecznik, N.M.; Harris, P.J.C.; Ramírez, L. Identification of Prosopis juliflora and Prosopis pallida Accessions Using Molecular Markers. Biodivers. Conserv. 2005, 15, 1829–1844. [Google Scholar] [CrossRef]
- Neuhaus, H.; Emes, M. Nonphotosynthetic metabolism in plastids. Annu. Rev. Plant Biol. 2000, 51, 111–140. [Google Scholar] [CrossRef] [PubMed]
- Clegg, M.T.; Gaut, B.S.; Learn, G.H.; Morton, B.R. Rates and patterns of chloroplast DNA evolution. Proc. Natl. Acad. Sci. USA 1994, 91, 6795–6801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, J.D. Plastid Chromosomes: Structure and Evolution. The Molecular Biology of Plastids 1991, 7, 5–53. [Google Scholar] [CrossRef]
- Shaw, J.; Lickey, E.B.; Beck, J.T.; Farmer, S.B.; Liu, W.; Miller, J.; Siripun, K.C.; Winder, C.T.; Schilling, E.E.; Small, R.L. The tortoise and the hare II: Relative utility of 21 noncoding chloroplast DNA sequences for phylogenetic analysis. Am. J. Bot. 2005, 92, 142–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, D.; Kane, N.C.; Ebert, D.P.; Rieseberg, L.H. Genome skimming reveals the origin of the Jerusalem Artichoke tuber crop species: Neither from Jerusalem nor an artichoke. New Phytol. 2013, 201, 1021–1030. [Google Scholar] [CrossRef]
- Shaw, J.; Shafer, H.L.; Leonard, O.R.; Kovach, M.J.; Schorr, M.; Morris, A.B. Chloroplast DNA sequence utility for the lowest phylogenetic and phylogeographic inferences in angiosperms: The tortoise and the hare IV. Am. J. Bot. 2014, 101, 1987–2004. [Google Scholar] [CrossRef] [Green Version]
- Du, F.; Petit, R.; Liu, J.Q. More introgression with less gene flow: Chloroplast vs. mitochondrial DNA in thePicea asperatacomplex in China, and comparison with other Conifers. Mol. Ecol. 2009, 18, 1396–1407. [Google Scholar] [CrossRef]
- Kane, N.C.; King, M.G.; Barker, M.S.; Raduski, A.; Karrenberg, S.; Yatabe, Y.; Knapp, S.J.; Rieseberg, L.H. Comparative genomic and population genetic analyses indicate highly porous genomes and high levels of gene flow between divergent Helianthus species. Evol. Int. J. Org. Evol. 2009, 63, 2061–2075. [Google Scholar] [CrossRef] [Green Version]
- A McGuire, J.; Linkem, C.W.; Koo, M.S.; Hutchison, D.W.; Lappin, A.K.; Orange, D.I.; Lemos-Espinal, J.; Riddle, B.R.; Jaeger, J.R. Mitochondrial Introgression and incomplete lineage sorting through SPACE and time: Phylogenetics of crotaphytid lizards. Evol. 2007, 61, 2879–2897. [Google Scholar] [CrossRef]
- Newton, A.; Allnutt, T.; Gillies, A.; Lowe, A.; Ennos, R. Molecular phylogeography, intraspecific variation and the conservation of tree species. Trends Ecol. Evol. 1999, 14, 140–145. [Google Scholar] [CrossRef]
- Jia, Y.; Yang, J.; He, Y.-L.; He, Y.; Niu, C.; Gong, L.-L.; Li, Z.-H. Characterization of the whole chloroplast genome sequence of Acer davidii Franch (Aceraceae). Conserv. Genet. Resour. 2016, 8, 141–143. [Google Scholar] [CrossRef]
- Li, Z.-H.; Xie, Y.-S.; Zhou, T.; Jia, Y.; He, Y.-L.; Yang, J. The complete chloroplast genome sequence of Acer morrisonense (Aceraceae). Mitochondrial DNA Part A 2015, 28, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-B.; Yang, S.-X.; Li, H.-T.; Yang, J.; Li, D.-Z. Comparative Chloroplast Genomes of Camellia Species. PLoS ONE 2013, 8, e73053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, J.; Ma, P.-F.; Li, H.-T.; Li, D.-Z. Complete Plastid Genome Sequencing of Four Tilia Species (Malvaceae): A Comparative Analysis and Phylogenetic Implications. PLoS ONE 2015, 10, e0142705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asaf, S.; Khan, A.L.; Khan, M.A.; Waqas, M.; Kang, S.-M.; Yun, B.-W.; Lee, I.-J. Chloroplast genomes of Arabidopsis halleri ssp. gemmifera and Arabidopsis lyrata ssp. petraea: Structures and comparative analysis. Sci. Rep. 2017, 7, 7556. [Google Scholar] [CrossRef] [Green Version]
- Nikiforova, M.N.; Wald, A.I.; Roy, S.; Durso, M.B.; Nikiforov, Y.E. Targeted next-generation sequencing panel (ThyroSeq) for detection of mutations in thyroid cancer. J. Clin. Endocrinol. Metab. 2013, 98, E1852–E1860. [Google Scholar] [CrossRef] [Green Version]
- Caballero, J.C.; Alonso, R.; Ibáñez, V.; Terol, J.; Talon, M.; Dopazo, J. A Phylogenetic Analysis of 34 Chloroplast Genomes Elucidates the Relationships between Wild and Domestic Species within the Genus Citrus. Mol. Boil. Evol. 2015, 32, 2015–2035. [Google Scholar] [CrossRef] [Green Version]
- Daniell, H.; Lin, C.; Yu, M.; Chang, W.-J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Boil. 2016, 17, 134. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Hefer, C.A.; Kolosova, N.; Douglas, C.J.; Cronk, Q. Whole plastome sequencing reveals deep plastid divergence and cytonuclear discordance between closely related balsam poplars, Populus balsamifera and P. trichocarpa (Salicaceae). New Phytol. 2014, 204, 693–703. [Google Scholar] [CrossRef]
- Kersten, B.; Rampant, P.F.; Mader, M.; Le Paslier, M.-C.; Bounon, R.; Bérard, A.; Vettori, C.; Schroeder, H.; Leple, J.-C.; Fladung, M. Genome Sequences of Populus tremula Chloroplast and Mitochondrion: Implications for Holistic Poplar Breeding. PLoS ONE 2016, 11, e0147209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.-S.; Li, P.; Qiu, Y.-X. The Complete Chloroplast Genomes of Three Cardiocrinum (Liliaceae) Species: Comparative Genomic and Phylogenetic Analyses. Front. Plant Sci. 2017, 7, 232. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhou, T.; Duan, D.; Yang, J.; Feng, L.; Zhao, G.-F. Comparative Analysis of the Complete Chloroplast Genomes of Five Quercus Species. Front. Plant Sci. 2016, 7, 803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asaf, S.; Khan, A.L.; Khan, A.R.; Waqas, M.; Kang, S.-M.; Khan, M.A.; Lee, S.-M.; Lee, I.-J. Complete Chloroplast Genome of Nicotiana otophora and its Comparison with Related Species. Front. Plant Sci. 2016, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Liao, D.; Jia, C.; Sun, P.; Qi, J.; Li, X. Quality evaluation of Panax quinquefolium from different cultivation regions based on their ginsenoside content and radioprotective effects on irradiated mice. Sci. Rep. 2019, 9, 1079. [Google Scholar] [CrossRef] [Green Version]
- Bessega, C.F.; Pometti, C.L.; Miller, J.T.; Watts, R.; Saidman, B.O.; Vilardi, J.C. New microsatellite loci for Prosopis alba and P. chilensis (Fabaceae)1. Appl. Plant Sci. 2013, 1, 1200324. [Google Scholar] [CrossRef] [PubMed]
- Kuang, D.-Y.; Wu, H.; Wang, Y.-L.; Gao, L.-M.; Zhang, S.-Z.; Lu, L. Complete chloroplast genome sequence of Magnolia kwangsiensis (Magnoliaceae): Implication for DNA barcoding and population genetics. Genome 2011, 54, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Asaf, S.; Waqas, M.; Khan, A.L.; Khan, M.A.; Kang, S.-M.; Imran, Q.M.; Shahzad, R.; Bilal, S.; Yun, B.-W.; Lee, I.-J. The Complete Chloroplast Genome of Wild Rice (Oryza minuta) and Its Comparison to Related Species. Front. Plant Sci. 2017, 8, 345. [Google Scholar] [CrossRef] [Green Version]
- Wicke, S.; Schneeweiss, G.M.; Depamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Boil. 2011, 76, 273–297. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Kong, H.; Zhou, J.; Fritsch, P.W.; Hao, G.; Gong, W. Complete Chloroplast Genome of Cercis chuniana (Fabaceae) with Structural and Genetic Comparison to Six Species in Caesalpinioideae. Int. J. Mol. Sci. 2018, 19, 1286. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Hu, N.; Wu, H. Analyzing and Characterizing the Chloroplast Genome of Salix wilsonii. BioMed Res. Int. 2019, 2019, 5190425. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, E.N.; Ruhlman, T.A.; Sabir, J.S.M.; Hajrah, N.H.; Alharbi, N.S.; Bailey, C.D.; Jansen, R.K.; Hajarah, N.H.; Al-Malki, A.L. Plastid genome sequences of legumes reveal parallel inversions and multiple losses ofrps16in papilionoids. J. Syst. Evol. 2015, 53, 458–468. [Google Scholar] [CrossRef]
- Chumley, T.W.; Palmer, J.D.; Mower, J.P.; Fourcade, H.M.; Calie, P.J.; Boore, J.L.; Jansen, R.K. The Complete Chloroplast Genome Sequence of Pelargonium × hortorum: Organization and Evolution of the Largest and Most Highly Rearranged Chloroplast Genome of Land Plants. Mol. Boil. Evol. 2006, 23, 2175–2190. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Liu, L.; Zhao, M.; Zhang, C.; Zhang, D.; Li, Y.; Li, S. The complete chloroplast genomes of seventeen Aegilops tauschii: Genome comparative analysis and phylogenetic inference. PeerJ 2020, 8, e8678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, X.; Deng, P.; Feng, K.; Liu, P.; Du, X.; You, F.M.; Weining, S. Comparative analysis of codon usage patterns in chloroplast genomes of the Asteraceae family. Plant Mol. Boil. Rep. 2013, 32, 828–840. [Google Scholar] [CrossRef]
- Vieira, L.D.N.; Faoro, H.; Rogalski, M.; Fraga, H.P.D.F.; Cardoso, R.L.A.; De Souza, E.M.; Pedrosa, F.D.O.; Nodari, R.O.; Guerra, M.P. The Complete Chloroplast Genome Sequence of Podocarpus lambertii: Genome Structure, Evolutionary Aspects, Gene Content and SSR Detection. PLoS ONE 2014, 9, 9. [Google Scholar] [CrossRef]
- Piovani, P.; Leonardi, S.; Piotti, A.; Menozzi, P. Conservation genetics of small relic populations of silver fir (Abies alba Mill.) in the northern Apennines. Plant Biosyst. Int. J. Deal. Asp. Plant Boil. 2010, 144, 683–691. [Google Scholar] [CrossRef]
- Wang, T.; Wang, Z.; Chen, G.; Wang, C.; Su, Y. Invasive Chloroplast Population Genetics of Mikania micrantha in China: No Local Adaptation and Negative Correlation between Diversity and Geographic Distance. Front. Plant Sci. 2016, 7, 2626. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.-S.; Cheon, K.-S.; Hong, S.-Y.; Cho, J.-H.; Im, J.-S.; Mekapogu, M.; Yu, Y.-S.; Park, T.-H. Complete chloroplast genome sequences of Solanum commersonii and its application to chloroplast genotype in somatic hybrids with Solanum tuberosum. Plant Cell Rep. 2016, 35, 2113–2123. [Google Scholar] [CrossRef]
- Williams, A.F.; Miller, J.; Small, I.D.; Nevill, P.G.; Boykin, L.M. Integration of complete chloroplast genome sequences with small amplicon datasets improves phylogenetic resolution in Acacia. Mol. Phylogenetics Evol. 2016, 96, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Milligan, B.G.; Hampton, J.N.; Palmer, J.D. Dispersed repeats and structural reorganization in subclover chloroplast DNA. Mol. Boil. Evol. 1989, 6, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Cavalier-Smith, T. Chloroplast evolution: Secondary symbiogenesis and multiple losses. Curr. Boil. 2002, 12, R62–R64. [Google Scholar] [CrossRef] [Green Version]
- Bausher, M.G.; Singh, N.D.; Lee, S.-B.; Jansen, R.K.; Daniell, H. The complete chloroplast genome sequence of Citrus sinensis (L.) Osbeck var ’Ridge Pineapple’: Organization and phylogenetic relationships to other angiosperms. BMC Plant Boil. 2006, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Ebert, D.; Peakall, R. Chloroplast simple sequence repeats (cpSSRs): Technical resources and recommendations for expanding cpSSR discovery and applications to a wide array of plant species. Mol. Ecol. Resour. 2009, 9, 673–690. [Google Scholar] [CrossRef]
- Addisalem, A.B.; Esselink, G.D.; Bongers, F.; Smulders, M.J. Genomic sequencing and microsatellite marker development for Boswellia papyrifera, an economically important but threatened tree native to dry tropical forests. AoB PLANTS 2015, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2010, 177, 309–334. [Google Scholar] [CrossRef]
- Sherry, M.; Smith, S.; Patel, A.; Harris, P.; Hand, P.; Trenchard, L.; Henderson, J. RAPD and microsatellite transferability studies in selected species of Prosopis (section Algarobia) with emphasis on Prosopis juliflora and P. pallida. J. Genet. 2011, 90, 251–264. [Google Scholar] [CrossRef]
- Mottura, M.C.; Finkeldey, R.; Verga, A.R.; Gailing, O. Development and characterization of microsatellite markers for Prosopis chilensis and Prosopis flexuosa and cross-species amplification. Mol. Ecol. Notes 2005, 5, 487–489. [Google Scholar] [CrossRef]
- Pomponio, M.F.; Acuña, C.; Petreath, V.; Lauenstein, D.L.; Poltri, S.M.; Torales, S. Characterization of functional SSR markers in Prosopis alba and their transferability across Prosopis species. For. Syst. 2015, 24, 13. [Google Scholar] [CrossRef] [Green Version]
- Yi, X.; Gao, L.; Wang, B.; Su, Y.-J.; Wang, T. The complete chloroplast genome sequence of Cephalotaxus oliveri (Cephalotaxaceae): Evolutionary comparison of cephalotaxus chloroplast DNAs and insights into the loss of inverted repeat copies in gymnosperms. Genome Boil. Evol. 2013, 5, 688–698. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Song, J.; Gao, H.; Zhu, Y.; Xu, J.; Pang, X.; Yao, H.; Sun, C.; Li, X.; Li, C.; et al. The Complete Chloroplast Genome Sequence of the Medicinal Plant Salvia miltiorrhiza. PLoS ONE 2013, 8, e57607. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Hahn, F.M.; Mcmahan, C.; Cornish, K.; Whalen, M.C. Comparative analysis of the complete sequence of the plastid genome of Parthenium argentatum and identification of DNA barcodes to differentiate Parthenium species and lines. BMC Plant Boil. 2009, 9, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.H.; Abe, J.; Sakai, M.; Kanazawa, A.; Shimamoto, Y. Sequence variation of non-coding regions of chloroplast DNA of soybean and related wild species and its implications for the evolution of different chloroplast haplotypes. Theor. Appl. Genet. 2000, 101, 724–732. [Google Scholar] [CrossRef]
- Kanazawa, A.; Tozuka, A.; Shimamoto, Y. Sequence variation of chloroplast DNA that involves EcoRI and ClaI restriction site polymorphisms in soybean. Genes Genet. Syst. 1998, 73, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, R.L.; Ryburn, J.A.; Cronn, R.; Seelanan, T.; Wendel, J.F.; Wendel, T.S.F. The tortoise and the hare: Choosing between noncoding plastome and nuclearAdhsequences for phylogeny reconstruction in a recently diverged plant group. Am. J. Bot. 1998, 85, 1301–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, P.-C.; Zhang, Y.-Z.; Geng, H.-M.; Chen, S.-L. The complete chloroplast genome sequence ofGentiana lawrenceivar. farreri(Gentianaceae) and comparative analysis with its congeneric species. PeerJ 2016, 4, e2540. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.-S.; Yun, B.-K.; Yoon, Y.-H.; Hong, S.-Y.; Mekapogu, M.; Kim, K.-H.; Yang, T.-J. Complete Chloroplast Genome Sequence of Tartary Buckwheat (Fagopyrum tataricum) and Comparative Analysis with Common Buckwheat (F. esculentum). PLoS ONE 2015, 10, e0125332. [Google Scholar] [CrossRef]
- Choi, K.S.; Chung, M.G.; Park, S. The Complete Chloroplast Genome Sequences of Three Veroniceae Species (Plantaginaceae): Comparative Analysis and Highly Divergent Regions. Front. Plant Sci. 2016, 7, 183. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.-S.; Park, T.-H. Complete chloroplast genome sequence of Solanum nigrum and development of markers for the discrimination of S. nigrum. Hortic. Environ. Biotechnol. 2016, 57, 69–78. [Google Scholar] [CrossRef]
- Hu, Y.; Woeste, K.E.; Zhao, P. Completion of the Chloroplast Genomes of Five Chinese Juglans and Their Contribution to Chloroplast Phylogeny. Front. Plant Sci. 2017, 7, 803. [Google Scholar] [CrossRef] [Green Version]
- Blazier, J.C.; Jansen, R.K.; Mower, J.P.; Govindu, M.; Zhang, J.; Weng, M.-L.; Ruhlman, T. Variable presence of the inverted repeat and plastome stability inErodium. Ann. Bot. 2016, 117, 1209–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-H.; Wicke, S.; Wang, H.; Jin, J.-J.; Chen, S.-Y.; Zhang, S.-D.; Li, D.-Z.; Yi, T.-S. Plastid Genome Evolution in the Early-Diverging Legume Subfamily Cercidoideae (Fabaceae). Front. Plant Sci. 2018, 9, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, D.R.; Dastidar, S.G.; Cai, Z.; Penaflor, C.; Kuehl, J.; Boore, J.L.; Jansen, R.K. Phylogenetic and evolutionary implications of complete chloroplast genome sequences of four early-diverging angiosperms: Buxus (Buxaceae), Chloranthus (Chloranthaceae), Dioscorea (Dioscoreaceae), and Illicium (Schisandraceae). Mol. Phylogenetics Evol. 2007, 45, 547–563. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Shi, C.; Liu, Y.; Mao, S.-Y.; Gao, L.-Z. Thirteen Camelliachloroplast genome sequences determined by high-throughput sequencing: Genome structure and phylogenetic relationships. Bmc Evol. Biol. 2014, 14, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, S.; Zhou, T.; Han, K.; Yang, Y.; Zhao, J.; Liu, Z.-L. The Complete Chloroplast Genome Sequences of Six Rehmannia Species. Genes 2017, 8, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukas, B.; Novak, J. The complete chloroplast genome of Origanum vulgare L. (Lamiaceae). Gene 2013, 528, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Tan, Y.-H.; Liu, Y.-Y.; Song, Y.; Yang, J.-B.; Corlett, R.T. Chloroplast genome structure in Ilex (Aquifoliaceae). Sci. Rep. 2016, 6, 28559. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.E.; Rousseau-Gueutin, M.; Cordonnier, S.; Lima, O.; Michon-Coudouel, S.; Naquin, D.; De Carvalho, J.F.; Aïnouche, M.; Salmon, A.; Aïnouche, A. The first complete chloroplast genome of the Genistoid legume Lupinus luteus: Evidence for a novel major lineage-specific rearrangement and new insights regarding plastome evolution in the legume family. Ann. Bot. 2014, 113, 1197–1210. [Google Scholar] [CrossRef] [Green Version]
- Kazakoff, S.H.; Imelfort, M.; Edwards, D.; Koehorst, J.J.; Biswas, B.; Batley, J.; Scott, P.T.; Gresshoff, P.M. Capturing the Biofuel Wellhead and Powerhouse: The Chloroplast and Mitochondrial Genomes of the Leguminous Feedstock Tree Pongamia pinnata. PLoS ONE 2012, 7, e51687. [Google Scholar] [CrossRef]
- Wang, R.-J.; Cheng, C.-L.; Chang, C.-C.; Wu, C.-L.; Su, T.-M.; Chaw, S.-M. Dynamics and evolution of the inverted repeat-large single copy junctions in the chloroplast genomes of monocots. BMC Evol. Boil. 2008, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Wambugu, P.W.; Brozynska, M.; Furtado, A.; Waters, D.; Henry, R. Relationships of wild and domesticated rices (Oryza AA genome species) based upon whole chloroplast genome sequences. Sci. Rep. 2015, 5, 13957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, N.; Schmickl, R.; Chiang, T.-Y.; Lučanová, M.; Kolář, F.; Marhold, K.; Koch, M.A. Taming the wild: Resolving the gene pools of non-model Arabidopsis lineages. Bmc Evol. Biol. 2014, 14, 224. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J. Phylogeny of the legume family: An approach to understanding the origins of nodulation. Annu. Rev. Ecol. Syst. 1994, 25, 325–349. [Google Scholar] [CrossRef]
- Chappill, J. Cladistic analysis of the Leguminosae: The development of an explicit phylogenetic hypothesis. In Advances in Legume Systematics; Royal Botanic Gardens: London, UK, 1995; pp. 1–9. [Google Scholar]
- Doyle, J.J.; Doyle, J.L.; Ballenger, J.A.; Dickson, E.E.; Kajita, T.; Ohashi, H. A Phylogeny of the Chloroplast Gene RBC L in the Leguminosae: Taxonomic correlations and Insights Into the Evolution of Nodulation. Am. J. Bot. 1997, 84, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowski, M.F.; Lavin, M.; Sanderson, M.J.; Belmonte, J.; Vilà, M. A phylogeny of legumes (Leguminosae) based on analysis of the plastid mat K gene resolves many well-supported subclades within the family. Am. J. Bot. 2004, 91, 1846–1862. [Google Scholar] [CrossRef] [PubMed]
- Bruneau, A.; Mercure, M.; Lewis, G.P.; Herendeen, P. Phylogenetic patterns and diversification in the caesalpinioid legumesThis paper is one of a selection of papers published in the Special Issue on Systematics Research. Bot. 2008, 86, 697–718. [Google Scholar] [CrossRef]
- Lewis, G.; Schrire, B.; Mackinder, B.; Lock, M. Legumes of the World; Royal Botanic Gardens, Kew: London, UK, 2005. [Google Scholar]
- Shi, C.; Hu, N.; Huang, H.; Gao, J.; Zhao, Y.-J.; Gao, L.-Z. An Improved Chloroplast DNA Extraction Procedure for Whole Plastid Genome Sequencing. PLoS ONE 2012, 7, e31468. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Hahn, C.; Bachmann, L.; Chevreux, B. Reconstructing mitochondrial genomes directly from genomic next-generation sequencing reads--A baiting and iterative mapping approach. Nucleic Acids Res. 2013, 41, e129. [Google Scholar] [CrossRef] [Green Version]
- Wyman, S.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schattner, P.; Brooks, A.; Lowe, T. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res 2005, 33, W686–W689. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, S. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. Paup*: Phylogenetic analysis using parsimony (and other methods) 4.0. B5. 2001. Available online: http://citeseerx.ist.psu.edu/viewdoc/summary?doi=10.1.1.458.6867 (accessed on 22 January 2020).
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Briefings Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Boil. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Gascuel, O. BIONJ: An improved version of the NJ algorithm based on a simple model of sequence data. Mol. Boil. Evol. 1997, 14, 685–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. Cineraria | P. Juliflora | P. Glandulosa | A. Microsperma | P. Javanica | |
|---|---|---|---|---|---|
| Size (bp) | 163,677 | 163,237 | 163,040 | 159,389 | 161,681 |
| Overall GC contents | 35.9 | 35.9 | 35.9 | 36.5 | 35.9 |
| LSC size (bp) | 92,937 | 92,495 | 92,310 | 88,577 | 91,093 |
| SSC size (bp) | 18,878 | 18,880 | 19,132 | 18,756 | 18,574 |
| IR size (bp) | 25,931 | 25,931 | 25,931 | 26,028 | 26,007 |
| Protein coding regions (bp) | 78,883 | 78,421 | 78,039 | 78,030 | 78,075 |
| tRNA size (bp) | 2868 | 2927 | 2810 | 2793 | 2794 |
| rRNA size (bp) | 9052 | 9052 | 9052 | 9052 | 9052 |
| Number of genes | 131 | 132 | 128 | 128 | 130 |
| Number of protein coding genes | 85 | 85 | 83 | 83 | 83 |
| Number of rRNAs | 8 | 8 | 8 | 8 | 8 |
| Number of tRNA s | 38 | 39 | 37 | 37 | 37 |
| Genes with introns | 21 | 21 | 23 | 22 | 23 |
| Gene | Location | Exon I (bp) | Intron 1 (bp) | Exon II (bp) | Intron II (bp) | Exon III (bp) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. j | P. c | P. g | P. j | P. c | P. g | P. j | P. c | P. g | P. j | P. c | P. g | P. j | P. c | P. g | ||
| atpF | LSC | 145 | 145 | 145 | 744 | 726 | 727 | 389 | 407 | 407 | ||||||
| petB | LSC | 6 | 6 | 6 | 815 | 815 | 815 | 642 | 642 | 642 | ||||||
| petD | LSC | 8 | 8 | 8 | 720 | 720 | 720 | 475 | 475 | 475 | ||||||
| rpl2 a | IR | 391 | 393 | 393 | 665 | 662 | 662 | 434 | 435 | 435 | ||||||
| rpl16 | LSC | 9 | 9 | 9 | 1173 | 1173 | 1173 | 399 | 399 | 399 | ||||||
| rps16 | LSC | 40 | 40 | 40 | 884 | 883 | 883 | 245 | 245 | 245 | ||||||
| rpoC1 | LSC | 432 | 432 | 432 | 842 | 802 | 802 | 1578 | 1617 | 1617 | ||||||
| rps12 * | LSC-IR | 114 | 114 | 114 | 232 | 232 | 232 | 540 | 540 | 540 | 26 | 26 | ||||
| clpP | LSC | 69 | 69 | 69 | 789 | 790 | 789 | 291 | 291 | 291 | 642 | 642 | 642 | 228 | 228 | 228 |
| ndhA | SSC | 552 | 553 | 552 | 1449 | 1452 | 1457 | 540 | 539 | 540 | ||||||
| ndhB a | IR | 777 | 777 | 777 | 685 | 684 | 685 | 756 | 756 | 756 | ||||||
| ycf3 | LSC | 126 | 126 | 126 | 729 | 729 | 728 | 228 | 228 | 228 | 738 | 738 | 738 | 153 | 153 | 153 |
| trnA-UGC a | IR | 38 | 38 | 38 | 802 | 794 | 802 | 35 | 35 | 35 | ||||||
| trnI –GAUa | IR | 42 | 42 | 42 | 948 | 948 | 948 | 35 | 35 | 35 | ||||||
| trnL-UAA | LSC | 37 | 37 | 37 | 536 | 536 | 536 | 50 | 50 | 50 | ||||||
| trnK-UUU | LSC | 29 | 29 | 29 | 2492 | 2491 | 2491 | 37 | 37 | 37 | ||||||
| trnG-GCC | LSC | 23 | 702 | 49 | ||||||||||||
| trnV-UAC | LSC | 37 | 37 | 37 | 619 | 619 | 619 | 39 | 39 | 39 | ||||||
| Category | Group of Genes | Name of Genes |
|---|---|---|
| Self-replication | Large subunit of ribosomal proteins | rpl2, 14, 16, 20, 23, 32, 33, 36 |
| Small subunit of ribosomal proteins | rps2, 3, 4, 7, 8, 11, 12, 14, 15, 16, 18, 19 | |
| DNA dependent RNA polymerase | rpoA, B, C1, C2 | |
| rRNA genes | rrn4.5, rrn5, rrn16, rrn23 | |
| tRNA genes | trnA-UGC, trnC-GCA, trnD-GUC, trnE-UUC trnF-GAA, trnfM-CAU, trnG-UCC, trnH-GUG, trnI-CAU, trnI-GAU, trnK-UUU, trnL-CAA, trnL-UAA, trnL-UAG, trnM-CAU, trnN-GUU, trnP-GGG, trnP-UGG, trnQ-UUG, trnR-ACG, trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC, trnV-UAC, trnW-CCA, trnY-GUA | |
| Photosynthesis | Photosystem I | psaA, B, C, I, |
| Photosystem II | psbA, B, C, D, E, F, H, I, J, K, L, M, N, T, Z | |
| Cytochrome b6/f complex | petA, B, D, G, L, N | |
| ATP synthase | atpA, B, E, F, H, I | |
| Rubisco | rbcL | |
| Maturase | matK | |
| Protease | clpP | |
| Envelop membrane protein | cemA | |
| Subunit Acetyl- CoA-Carboxylate | accD | |
| c-type cytochrome synthesis gene | ccsA | |
| Unknown | Conserved Open reading frames | ycf1, 2, 3, 4 |
| T/U | C | A | G | Length (bp) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. j | P. c | P. g | P. j | P. c | P. g | P. j | P. c | P. g | P. j | P. c | P. g | P. j | P. c | P. g | |
| Genome | 32.5 | 32.5 | 32.5 | 18.3 | 18.3 | 18.3 | 31.6 | 31.6 | 31.7 | 17.6 | 17.6 | 17.6 | 163,237 | 163,677 | 163,040 |
| LSC | 34.2 | 34.2 | 34.2 | 17.0 | 17.0 | 17.0 | 32.6 | 32.6 | 32.6 | 16.1 | 16.2 | 16.1 | 92,495 | 92,937 | 92,310 |
| SSC | 34.9 | 34.9 | 34.6 | 15.9 | 16.0 | 15.7 | 34.7 | 34.6 | 35.2 | 14.5 | 14.5 | 14.4 | 18,880 | 18,878 | 19,132 |
| IR | 28.8 | 28.8 | 28.5 | 20.6 | 20.6 | 22.1 | 28.5 | 28.5 | 28.8 | 22.1 | 22.1 | 20.6 | 25,931 | 25,931 | 25,931 |
| tRNA | 25.1 | 24.9 | 24.8 | 23.4 | 23.5 | 23.8 | 22.1 | 22.0 | 21.9 | 29.4 | 29.6 | 29.5 | 2927 | 2868 | 2810 |
| rRNA | 18.9 | 18.9 | 18.8 | 23.7 | 23.7 | 23.6 | 25.7 | 25.7 | 25.7 | 31.7 | 31.7 | 31.9 | 9052 | 9052 | 9052 |
| Protein coding genes | 31.7 | 31.7 | 31.7 | 17.4 | 17.4 | 17.4 | 30.9 | 30.8 | 30.8 | 20.0 | 20.1 | 20.1 | 78,421 | 78,883 | 78,039 |
| 1st position | 32.6 | 33.0 | 32.4 | 17.9 | 18.14 | 18.2 | 31.4 | 31.5 | 31.7 | 17.9 | 17.5 | 17.5 | 54,412 | 54,559 | 54,347 |
| 2nd position | 32.4 | 32.33 | 32.7 | 18.5 | 18.4 | 17.7 | 31.5 | 31.3 | 32.1 | 17.4 | 17.8 | 17.5 | 54,412 | 54,559 | 54,347 |
| 3rd position | 32.3 | 32.44 | 32.4 | 20.9 | 18.1 | 18.8 | 32.0 | 31.9 | 31 | 17.4 | 17.3 | 17.7 | 54,412 | 54,559 | 54,347 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asaf, S.; Khan, A.L.; Khan, A.; Al-Harrasi, A. Unraveling the Chloroplast Genomes of Two Prosopis Species to Identify Its Genomic Information, Comparative Analyses and Phylogenetic Relationship. Int. J. Mol. Sci. 2020, 21, 3280. https://doi.org/10.3390/ijms21093280
Asaf S, Khan AL, Khan A, Al-Harrasi A. Unraveling the Chloroplast Genomes of Two Prosopis Species to Identify Its Genomic Information, Comparative Analyses and Phylogenetic Relationship. International Journal of Molecular Sciences. 2020; 21(9):3280. https://doi.org/10.3390/ijms21093280
Chicago/Turabian StyleAsaf, Sajjad, Abdul Latif Khan, Arif Khan, and Ahmed Al-Harrasi. 2020. "Unraveling the Chloroplast Genomes of Two Prosopis Species to Identify Its Genomic Information, Comparative Analyses and Phylogenetic Relationship" International Journal of Molecular Sciences 21, no. 9: 3280. https://doi.org/10.3390/ijms21093280