Genome-Wide Identification and Expression Profile of TPS Gene Family in Dendrobium officinale and the Role of DoTPS10 in Linalool Biosynthesis

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification and Features of TPS Proteins in D. officinale
2.2. Analysis of Conserved Motifs and Gene Structure
2.3. Phylogenetic Analysis of DoTPS Genes in D. officinale
2.4. Identification of Cis-Acting Elements in the Promoter Region of DoTPS Genes
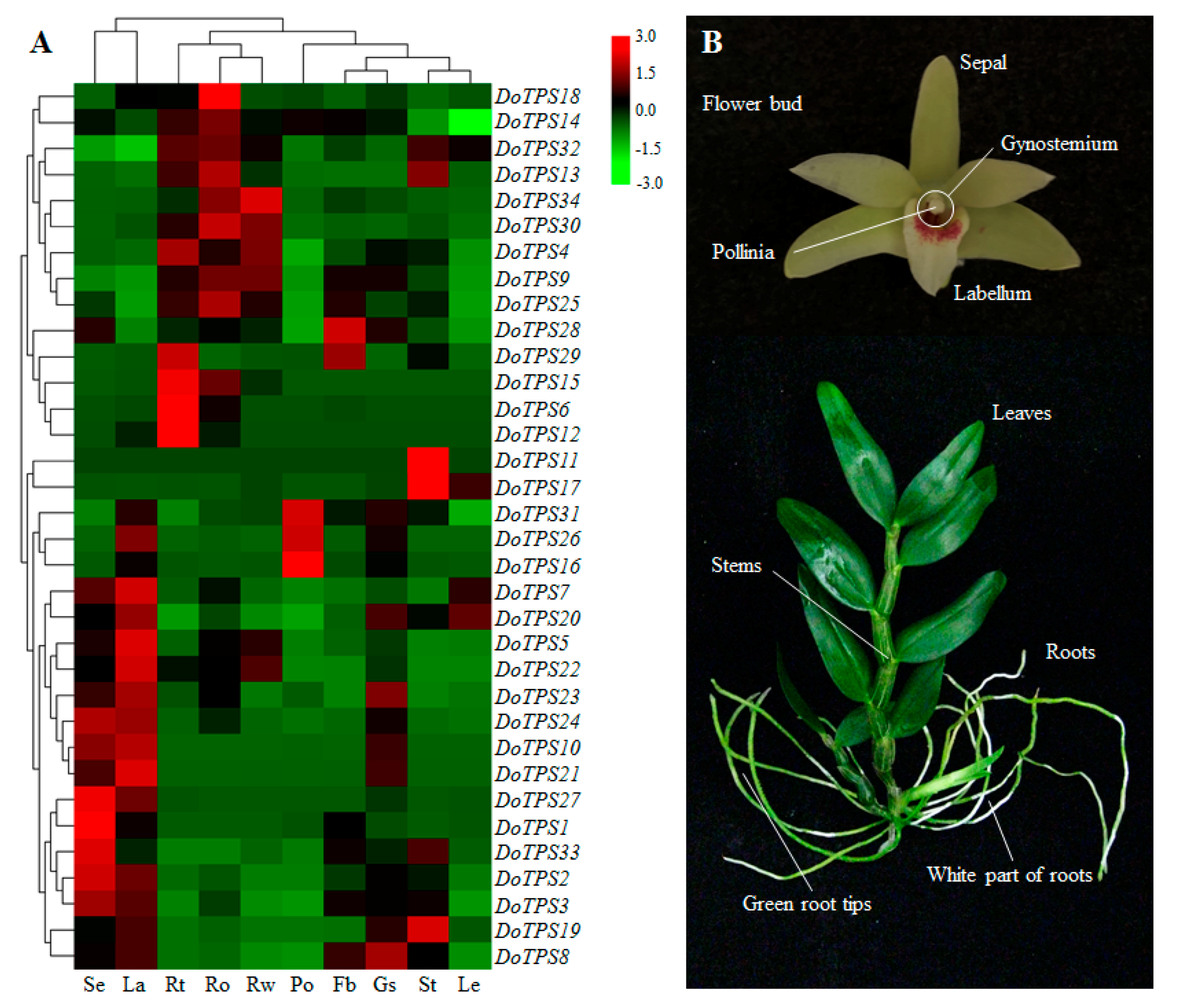
2.5. Tempospatial Expression Patterns of DoTPS Genes in Different D. officinale Organs
2.6. Expression Patterns of DoTPS Genes under Abiotic Stress
2.7. Expression Patterns of DoTPS Genes Subjected to MeJA Treatment
2.8. Transcription Abundance of DoTPS Genes at Budding and Flowering Stages
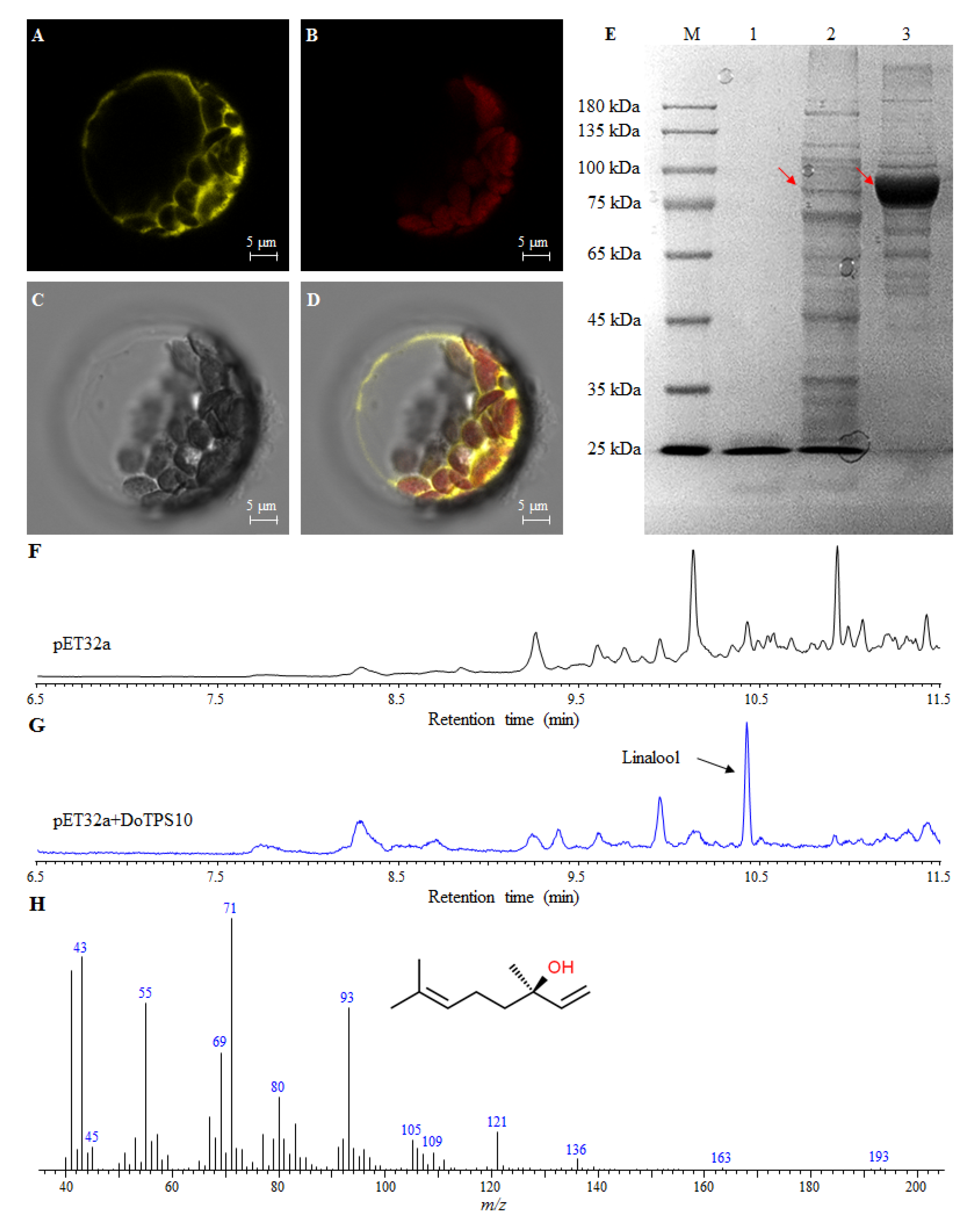
2.9. Subcellular Localization of DoTPS10 in Heterologous Plants
2.10. Functional Characterization of DoTPS10 Involved in the Formation of Linalool
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Identification of TPS Family Members in D. officinale
4.3. Conserved Motifs, Gene Structure, and Phylogenetic Analysis
4.4. Total RNA Isolation, cDNA Reverse Transcription, and RT-qPCR Analysis
4.5. Cis-Acting Elements Analysis of TPS Genes in D. officinale
4.6. Gene Expression Analysis Based on Transcriptome Data
4.7. Gas Chromatography–Mass Spectrometry Analysis of Geraniol and Linalool in Flowers of D. officinale
4.8. Prokaryotic Expression and DoTPS10 Enzyme Assay in Escherichia coli
4.9. Subcellular Localization of DoTPS10 in A. thaliana Mesophyll Protoplasts
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AI | Aliphatic index |
| bp | Base pair |
| CA | Cold acclimation |
| DMAPP | Dimethylallyl diphosphate |
| FPP | Farnesyl diphosphate |
| FPKM | Fragments per kilobase of transcript per million fragments mapped |
| GC–MS | Gas chromatography-mass spectrometry |
| GGPP | Geranylgeranyl diphosphate |
| GPP | Geranyl diphosphate |
| GRAVY | Grand average of hydrophobicity |
| II | Instability index |
| IPP | Isopentenyl diphosphate |
| MeJA | Methyl jasmonate |
| MEP | Methylerythritol phosphate |
| MS | Murashige and Skoog medium |
| Mw | Molecular weight |
| MVA | Mevalonic acid |
| NCBI | National Center for Biotechnology Information |
| NJ | Neighbor-joining |
| ORF | Open reading frame |
| pI | Isoelectric point |
| RT-qPCR | Real-time reverse transcription quantitative polymerase chain reaction |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| TPS | Terpene synthase |
| YFP | Yellow fluorescent protein |
References
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Byers, K.J.; Bradshaw, H.D., Jr.; Riffell, J.A. Three floral volatiles contribute to differential pollinator attraction in monkeyflowers (Mimulus). J. Exp. Biol. 2014, 217, 614–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Gfeller, V.; Erb, M. Root volatiles in plant-plant interactions II: Root volatiles alter root chemistry and plant-herbivore interactions of neighbouring plants. Plant Cell Environ. 2019, 42, 1964–1973. [Google Scholar] [CrossRef] [Green Version]
- Campbell, D.R.; Sosenski, P.; Raguso, R.A. Phenotypic plasticity of floral volatiles in response to increasing drought stress. Ann. Bot. 2019, 123, 601–610. [Google Scholar] [CrossRef]
- Hsiao, Y.Y.; Pan, Z.J.; Hsu, C.C.; Yang, Y.P.; Hsu, Y.C.; Chuang, Y.C.; Shih, H.H.; Chen, W.H.; Tsai, W.C.; Chen, H.H. Research on orchid biology and biotechnology. Plant Cell Physiol. 2011, 52, 1467–1486. [Google Scholar] [CrossRef] [Green Version]
- Ramya, M.; Jang, S.; An, H.R.; Lee, S.Y.; Park, P.M.; Park, P.H. Volatile organic compounds from orchids: From synthesis and function to gene regulation. Int. J. Mol. Sci. 2020, 21, 1160. [Google Scholar] [CrossRef] [Green Version]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar]
- Bohlmann, J.; Meyer-Gauen, G.; Croteau, R. Plant terpenoid synthases: Molecular biology and phylogenetic analysis. Proc. Natl. Acad. Sci. USA 1998, 95, 4126–4133. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.Y.; Jin, J.; Sarojam, R.; Ramachandran, S.A. Comprehensive survey on the terpene synthase gene family provides new insight into its evolutionary patterns. Genome Biol. Evol. 2019, 11, 2078–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubourg, S.; Lecharny, A.; Bohlmann, J. Genomic analysis of the terpenoid synthase (AtTPS) gene family of Arabidopsis thaliana. Mol. Genet. Genom. 2002, 267, 730–745. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.C.; Shamala, L.F.; Yi, X.K.; Yan, Z.; Wei, S. Analysis of terpene synthase family genes in Camellia sinensis with an emphasis on abiotic stress conditions. Sci. Rep. 2020, 10, 933. [Google Scholar] [CrossRef] [PubMed]
- Keilwagen, J.; Lehnert, H.; Berner, T.; Budahn, H.; Nothnagel, T.; Ulrich, D.; Dunemann, F. The terpene synthase gene family of carrot (Daucus carota L.): Identification of QTLs and candidate genes associated with terpenoid volatile compounds. Front. Plant Sci. 2017, 8, 1930. [Google Scholar] [CrossRef] [Green Version]
- Külheim, C.; Padovan, A.; Hefer, C.; Krause, S.T.; Köllner, T.G.; Myburg, A.A.; Degenhardt, J.; Foley, W.J. The Eucalyptus terpene synthase gene family. BMC Genom. 2015, 16, 450. [Google Scholar] [CrossRef] [Green Version]
- Nieuwenhuizen, N.J.; Green, S.A.; Chen, X.; Bailleul, E.J.D.; Matich, A.J.; Wang, M.Y.; Atkinson, R.G. Functional genomics reveals that a compact terpene synthase gene family can account for terpene volatile production in apple. Plant Physiol. 2013, 161, 787–804. [Google Scholar] [CrossRef] [Green Version]
- Falara, V.; Akhtar, T.A.; Nguyen, T.T.H.; Spyropoulou, E.A.; Bleeker, P.M.; Schauvinhold, I.; Matsuba, Y.; Bonini, M.E.; Schilmiller, A.L.; Last, R.L.; et al. The tomato terpene synthase gene family. Plant Physiol. 2011, 157, 770–789. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Köllner, T.G.; Yin, Y.; Jiang, Y.; Chen, H.; Xu, Y.; Gershenzon, J.; Pichersky, E.; Chen, F. Nonseed plant Selaginella moellendorffii has both seed plant and microbial types of terpene synthases. Proc. Natl. Acad. Sci. USA 2012, 109, 14711–14715. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.M.; Aubourg, S.; Schouwey, M.B.; Daviet, L.; Schalk, M.; Toub, O.; Lund, S.T.; Bohlmann, J. Functional annotation, genome organization and phylogeny of the grapevine (Vitis vinifera) terpene synthase gene family based on genome assembly, FLcDNA cloning, and enzyme assays. BMC Plant Biol. 2010, 10, 226. [Google Scholar] [CrossRef] [Green Version]
- Chuang, Y.C.; Hung, Y.C.; Tsai, W.C.; Chen, W.H.; Chen, H.H. PbbHLH4 regulates floral monoterpene biosynthesis in Phalaenopsis orchids. J. Exp. Bot. 2018, 69, 4363–4377. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Liu, B.; Li, M.; Gao, X.; Fang, Q.; Liu, C.; Ding, H.; Wang, L.; Gao, X. Identification and characterization of terpene synthase genes accounting for volatile terpene emissions in flowers of Freesia × hybrida. J. Exp. Bot. 2018, 69, 4249–4265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Wang, X.; Liu, H.; Tian, Y.; Lian, J.; Yang, R.; Hao, S.; Wang, X.; Yang, S.; Li, Q.; et al. The genome of Dendrobium officinale illuminates the biology of the important traditional Chinese orchid herb. Mol. Plant 2015, 8, 922–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Xu, Q.; Bian, C.; Tsai, W.C.; Yeh, C.M.; Liu, K.; Yoshida, K.; Zhang, L.; Chang, S.; Chen, F.; et al. The Dendrobium catenatum Lindl genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, W.C.; Dievart, A.; Hsu, C.C.; Hsiao, Y.Y.; Chiou, S.Y.; Huang, H.; Chen, H.H. Post genomics era for orchid research. Bot. Stud. 2017, 58, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.Q.; Liu, K.W.; Li, Z.; Lohaus, R.; Hsiao, Y.Y.; Niu, S.C.; Wang, J.Y.; Lin, Y.C.; Xu, Q.; Chen, L.J.; et al. The Apostasia genome and the evolution of orchids. Nature 2017, 549, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Kaundal, R.; Saini, R.; Zhao, P.X. Combining machine learning and homology-based approaches to accurately predict subcellular localization in Arabidopsis. Plant Physiol. 2010, 154, 36–54. [Google Scholar] [CrossRef] [Green Version]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE. 2010, 5, 11335. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Xiao, X.; Chou, K.C. PLoc-mPlant: Predict subcellular localization of multi-location plant proteins by incorporating the optimal GO information into general PseAAC. Mol. Biosyst. 2017, 13, 1722–1727. [Google Scholar] [CrossRef]
- Wu, Z.; Jiang, W.; Chen, S.; Mantri, N.; Tao, Z.; Jiang, C. Insights from the cold transcriptome and metabolome of Dendrobium officinale: Global reprogramming of metabolic and gene regulation networks during cold acclimation. Front. Plant Sci. 2016, 7, 1653. [Google Scholar] [CrossRef] [Green Version]
- Teixeira da Silva, J.A.; Ng, T.B. The medicinal and pharmaceutical importance of Dendrobium species. Appl. Microbiol. Biotechnol. 2017, 101, 2227–2239. [Google Scholar] [CrossRef]
- Yu, Z.; Yang, Z.; Teixeira da Silva, J.A.; Luo, J.; Duan, J. Influence of low temperature on physiology and bioactivity of postharvest Dendrobium officinale stems. Postharvest Biol. Tech. 2019, 148, 97–106. [Google Scholar] [CrossRef]
- Wan, X.; Zou, L.; Zheng, B.; Tian, Y.; Wang, Y. Transcriptomic profiling for prolonged drought in Dendrobium catenatum. Sci. Data 2018, 5, 180233. [Google Scholar] [CrossRef] [PubMed]
- Kaul, S.; Koo, H.L.; Jenkins, J.; Rizzo, M.; Rooney, T.; Tallon, L.J.; Feldblyum, T.; Nierman, W.; Benito, M.I.; Town, C.D.; et al. The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar]
- Banks, J.A.; Nishiyama, T.; Hasebe, M.; Bowman, J.L.; Gribskov, M.; de Pamphilis, C.; Albert, V.A.; Aono, N.; Aoyama, T.; Ambrose, B.A.; et al. The Selaginella genome identifies genetic changes associated with the evolution of vascular plants. Science 2011, 332, 960–963. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Kurata, N.; Wei, X.; Wang, Z.X.; Wang, A.; Zhao, Q.; Zhao, Y.; Liu, K.; Lu, H.; Li, W.; et al. A map of rice genome variation reveals the origin of cultivated rice. Nature 2012, 490, 497–501. [Google Scholar] [CrossRef] [Green Version]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. French-Italian Public Consortium for Grapevine Genome Characterization. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar]
- Hong, G.J.; Xue, X.Y.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plantarum 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; He, C.; Teixeira da Silva, J.A.; Luo, J.; Yang, Z.; Duan, J. The GDP-mannose transporter gene (DoGMT) from Dendrobium officinale is critical for mannan biosynthesis in plant growth and development. Plant Sci. 2018, 277, 43–54. [Google Scholar] [CrossRef]
- An, H.; Zhu, Q.; Pei, W.; Fan, J.; Liang, Y.; Cui, Y.; Lv, N.; Wang, W. Whole-transcriptome selection and evaluation of internal reference genes for expression analysis in protocorm development of Dendrobium officinale Kimura et Migo. PLoS ONE 2016, 11, 163478. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhang, G.; Teixeira da Silva, J.A.; Yang, Z.; Duan, J. The β-1,3-galactosetransferase gene DoGALT2 is essential for stigmatic mucilage production in Dendrobium officinale. Plant Sci. 2019, 287, 110179. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Liao, Y.; Zeng, L.; Dong, F.; Watanabe, N.; Yang, Z. Transformation of catechins into theaflavins by upregulation of CsPPO3 in preharvest tea (Camellia sinensis) leaves exposed to shading treatment. Food Res. Int. 2020, 129, 108842. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID 1 | ORF 2 (bp) | AA 3 (aa) | pI 4 | Mw 5 (kDa) | AI 6 | II 7 | GRAVY 8 | Localization 9 |
|---|---|---|---|---|---|---|---|---|---|
| DoTPS1 | Dca014928 | 960 | 319 | 6.31 | 36.82 | 90.78 | 48.81 | −0.342 | Chloroplast a,b,c |
| DoTPS2 | Dca000724 | 1902 | 633 | 6.47 | 74.02 | 88.44 | 43.11 | −0.260 | Chloroplast a,b,c |
| DoTPS3 | Dca000725 | 1827 | 608 | 5.46 | 70.52 | 89.67 | 41.24 | −0.231 | Chloroplast a,b,c |
| DoTPS4 | Dca022838 | 2571 | 856 | 7.18 | 100.05 | 80.35 | 47.78 | −0.429 | Chloroplast a,b,c |
| DoTPS5 | Dca003141 | 1692 | 563 | 5.11 | 65.84 | 98.86 | 42.13 | −0.228 | Chloroplast a,b/Cytoplasm c |
| DoTPS6 | Dca019411 | 1521 | 506 | 5.13 | 59.48 | 93.68 | 43.71 | −0.133 | Chloroplast a,b/Cytoplasm c |
| DoTPS7 | Dca003139 | 1692 | 563 | 5.67 | 65.72 | 92.49 | 39.06 | −0.266 | Chloroplast a,b/Cytoplasm c |
| DoTPS8 | Dca028160 | 579 | 192 | 6.83 | 22.63 | 100.62 | 45.31 | −0.121 | Chloroplast a/Unknown b/Cytoplasm c |
| DoTPS9 | Dca019412 | 1665 | 554 | 5.03 | 64.93 | 92.94 | 39.76 | −0.189 | Chloroplast a,b/Cytoplasm c |
| DoTPS10 | Dca007746 | 1797 | 598 | 5.73 | 69.73 | 93.61 | 47.41 | −0.242 | Chloroplast a,b/Cytoplasm c |
| DoTPS11 | Dca022749 | 696 | 231 | 5.13 | 27.40 | 104.20 | 49.98 | −0.045 | Chloroplast a,b/Cytoplasm c |
| DoTPS12 | Dca024936 | 378 | 125 | 5.64 | 14.98 | 98.32 | 40.18 | −0.326 | Chloroplast a/Cytoplasm b,c |
| DoTPS13 | Dca026570 | 1659 | 552 | 5.59 | 64.94 | 96.97 | 38.16 | −0.266 | Chloroplast a,b,c |
| DoTPS14 | Dca005188 | 2550 | 849 | 6.71 | 98.69 | 86.21 | 46.66 | −0.352 | Chloroplast a,b,c |
| DoTPS15 | Dca025698 | 1659 | 552 | 5.31 | 64.82 | 89.60 | 44.73 | −0.284 | Chloroplast a,b,c |
| DoTPS16 | Dca016979 | 1650 | 549 | 5.62 | 64.23 | 91.62 | 36.94 | −0.374 | Chloroplast a,b/Cytoplasm c |
| DoTPS17 | Dca008309 | 1653 | 550 | 5.42 | 64.89 | 97.13 | 47.01 | −0.233 | Chloroplast a,b,c |
| DoTPS18 | Dca011215 | 1674 | 557 | 5.36 | 64.80 | 95.10 | 44.24 | −0.302 | Chloroplast a,b,c |
| DoTPS19 | Dca010855 | 1446 | 481 | 4.94 | 55.95 | 110.46 | 34.63 | 0.013 | Chloroplast a,b,c |
| DoTPS20 | Dca026890 | 1749 | 582 | 5.20 | 68.07 | 92.84 | 33.82 | −0.295 | Chloroplast a,b,c |
| DoTPS21 | Dca007747 | 1797 | 598 | 5.62 | 69.60 | 94.92 | 48.15 | −0.224 | Chloroplast a,b/Cytoplasm c |
| DoTPS22 | Dca003142 | 1692 | 563 | 5.24 | 65.61 | 95.26 | 43.73 | −0.245 | Chloroplast a,b/Cytoplasm c |
| DoTPS23 | Dca011214 | 1674 | 557 | 5.22 | 65.03 | 90.36 | 37.30 | −0.331 | Chloroplast a,b/Cytoplasm c |
| DoTPS24 | Dca000728 | 1386 | 461 | 6.38 | 53.78 | 93.08 | 38.16 | −0.180 | Chloroplast a,b/Cytoplasm c |
| DoTPS25 | Dca013782 | 1794 | 597 | 5.31 | 69.67 | 94.61 | 44.76 | −0.274 | Chloroplast a,b/Cytoplasm c |
| DoTPS26 | Dca026369 | 1650 | 549 | 5.42 | 64.61 | 90.73 | 42.39 | −0.438 | Chloroplast a,b/Cytoplasm c |
| DoTPS27 | Dca000723 | 1938 | 645 | 5.89 | 74.89 | 91.74 | 48.68 | −0.240 | Chloroplast a,b/Cytoplasm c |
| DoTPS28 | Dca003295 | 1863 | 620 | 5.91 | 72.57 | 92.35 | 47.89 | −0.262 | Chloroplast a,b/Cytoplasm c |
| DoTPS29 | Dca018407 | 1653 | 550 | 5.57 | 64.68 | 95.89 | 50.10 | −0.251 | Chloroplast a,b/Cytoplasm c |
| DoTPS30 | Dca013784 | 1377 | 458 | 5.07 | 53.35 | 93.52 | 38.64 | −0.254 | Chloroplast a,b |
| DoTPS31 | Dca016966 | 1089 | 362 | 7.07 | 41.65 | 99.70 | 51.56 | −0.193 | Chloroplast a,b,c |
| DoTPS32 | Dca018946 | 2433 | 810 | 5.75 | 91.13 | 88.99 | 46.05 | −0.169 | Chloroplast a,b,c |
| DoTPS33 | Dca017971 | 1536 | 511 | 5.57 | 58.70 | 97.18 | 42.66 | −0.109 | Chloroplast a,b/Cytoplasm c |
| DoTPS34 | Dca020940 | 1797 | 598 | 5.19 | 69.59 | 91.52 | 45.72 | −0.280 | Chloroplast a,b,c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Zhao, C.; Zhang, G.; Teixeira da Silva, J.A.; Duan, J. Genome-Wide Identification and Expression Profile of TPS Gene Family in Dendrobium officinale and the Role of DoTPS10 in Linalool Biosynthesis. Int. J. Mol. Sci. 2020, 21, 5419. https://doi.org/10.3390/ijms21155419
Yu Z, Zhao C, Zhang G, Teixeira da Silva JA, Duan J. Genome-Wide Identification and Expression Profile of TPS Gene Family in Dendrobium officinale and the Role of DoTPS10 in Linalool Biosynthesis. International Journal of Molecular Sciences. 2020; 21(15):5419. https://doi.org/10.3390/ijms21155419
Chicago/Turabian StyleYu, Zhenming, Conghui Zhao, Guihua Zhang, Jaime A. Teixeira da Silva, and Jun Duan. 2020. "Genome-Wide Identification and Expression Profile of TPS Gene Family in Dendrobium officinale and the Role of DoTPS10 in Linalool Biosynthesis" International Journal of Molecular Sciences 21, no. 15: 5419. https://doi.org/10.3390/ijms21155419