
Molecular Mechanisms and Treatment of Sarcopenia in Liver Disease: A Review of Current Knowledge

 , , , , and
, , , , and
Abstract
:1. Introduction
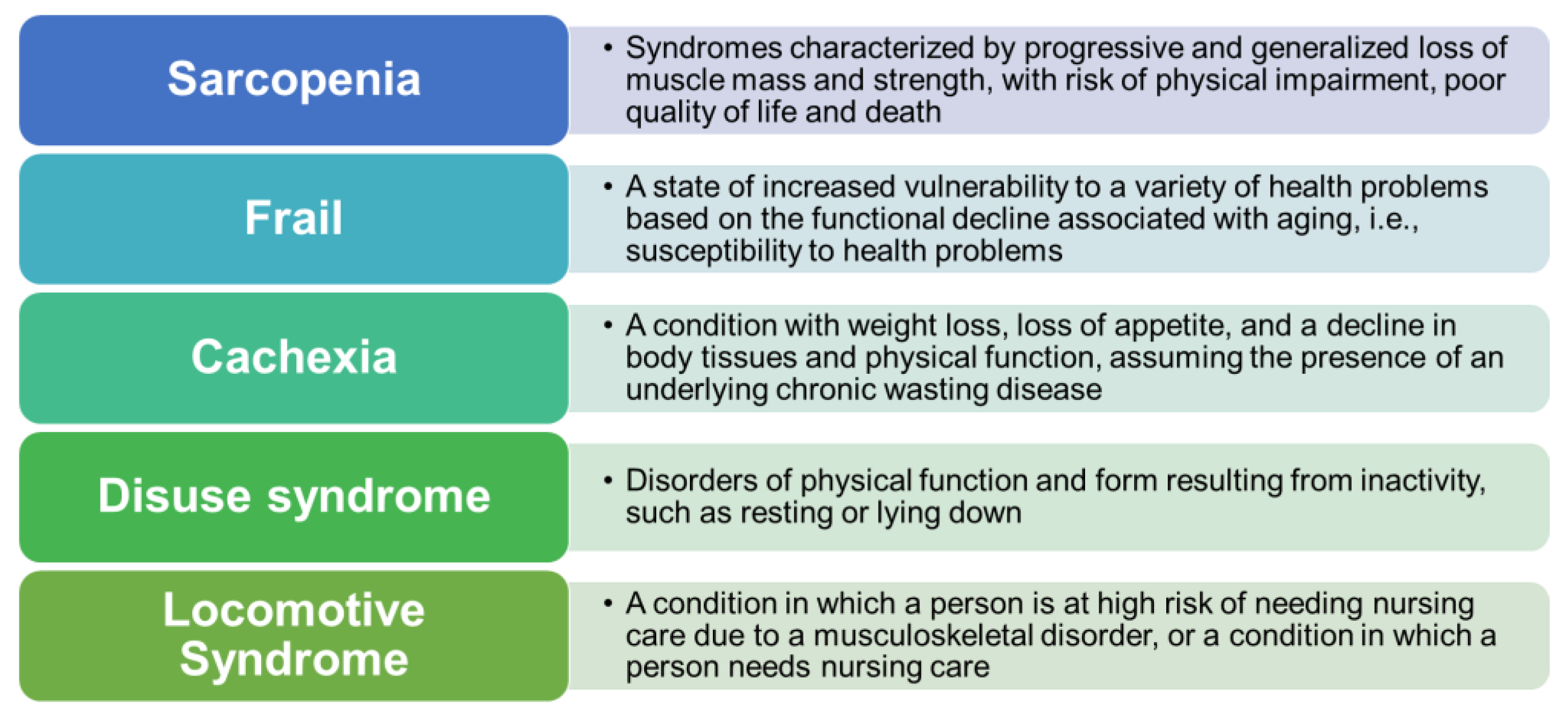
2. Classification of Sarcopenia
3. Mechanism of Sarcopenia in Liver Cirrhosis
3.1. Hyperammonemia
3.2. Lower Levels of BCAAs
3.3. Abnormal Sex Hormones
4. Modalities for the Assessment of Sarcopenia in the Clinical Field
5. Other Effects of Sarcopenia Related to Liver Cirrhosis
6. Nutritional Status in Liver Cirrhosis
7. Treatment of Sarcopenia in Liver Cirrhosis
7.1. Drug Medicine and Nutrients
7.2. Appropriate Exercise for Patients with Liver Disease
7.3. Fluctuations after Liver Transplantation
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Rosenberg, I. Summary comments: Epidemiological and methodological problems in determining the nutritional status of older persons. Am. J. Clin. Nutr. 1989, 50, 1231–1233. [Google Scholar] [CrossRef]
- ICD-11 for Mortality and Morbidity Statistics. Available online: https://icd.who.int/browse11/l-m/en (accessed on 12 March 2019).
- Eto, M. Locomotive syndrome and frailty. Recent advances in sarcopenia research. Clin. Calcium 2012, 22, 75–79. [Google Scholar]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef] [Green Version]
- Breen, L.; Phillips, S.M. Skeletal muscle protein metabolism in the elderly: Interventions to counteract the ‘anabolic resistance’ of aging. Nutr. Metab. 2011, 8, 68. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Moriguch, Y.; Mitani, T.; Aoyama, T.; Arai, H. Age-dependent changes in skeletal muscle mass and visceral fat area in Japanese adults from 40 to 79 years of age. Geriatr. Gerontol. Int. 2014, 14, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Bentov, I.; Kaplan, S.J.; Pham, T.N.; Reed, M.J. Frailty assessment: From clinical to radiological tools. Br. J. Anaesth. 2019, 123, 37–50. [Google Scholar] [CrossRef]
- Nishiguchi, S.; Hino, K.; Moriya, K.; Shiraki, M.; Hiramatsu, A.; Nishikawa, H. Assessment criteria for sarcopenia in liver disease (first edition): Report from the working group for creation of sarcopenia assessment criteria in the Japan Society of Hepatology. Kanzo 2016, 57, 353–368. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.K.; Liu, L.K.; Woo, J.; Assantachai, P.; Auyeung, T.W.; Bahyah, K.S.; Chou, M.Y.; Chen, L.Y.; Hsu, P.S.; Krairit, O.; et al. Sarcopenia in Asia: Consensus report of the Asian Working Group for Sarcopenia. J. Am. Med. Dir. Assoc. 2014, 15, 95–101. [Google Scholar] [CrossRef]
- Nishikawa, H.; Shiraki, M.; Hiramatsu, A.; Moriya, K.; Hino, K.; Nishiguchi, S. Japan Society of Hepatology guidelines for sarcopenia in liver disease (1st edition): Recommendation from the working group for creation of sarcopenia assessment criteria. Hepatol. Res. 2016, 46, 951–963. [Google Scholar] [CrossRef]
- Kobayashi, T.; Kawai, H.; Nakano, O.; Abe, S.; Kamimura, H.; Sakamaki, A.; Kamimura, K.; Tsuchiya, A.; Takamura, M.; Yamagiwa, S.; et al. Rapidly declining skeletal muscle mass predicts poor prognosis of hepatocellular carcinoma treated with transcatheter intra-arterial therapies. BMC Cancer 2018, 18, 756. [Google Scholar] [CrossRef]
- Volpi, E.; Mittendorfer, B.; Rasmussen, B.B.; Wolfe, R.R. The response of muscle protein anabolism to combined hyperaminoacidemia and glucose-induced hyperinsulinemia is impaired in the elderly. J. Clin. Endocrinol. Metab. 2000, 85, 4481–4490. [Google Scholar] [CrossRef] [Green Version]
- Dasarathy, S.; Merli, M. Sarcopenia from mechanism to diagnosis and treatment of liver disease. J. Hepatol. 2016, 65, 1232–1244. [Google Scholar] [CrossRef] [Green Version]
- Kitzmann, M.; Carnac, G.; Vandromme, M.; Primig, M.; Lamb, N.J.; Fernandez, A. The muscle regulatory factors MyoD and myf-5 undergo distinct cell cycle-specific expression in muscle cells. J. Cell Biol. 1998, 142, 1447–1459. [Google Scholar] [CrossRef] [Green Version]
- Egerman, M.A.; Glass, D.J. Signaling pathways controlling skeletal muscle mass. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Jindal, A.; Jagdish, R.K. Sarcopenia: Ammonia metabolism and hepatic encephalopathy. Clin. Mol. Hepatol. 2019, 25, 270–279. [Google Scholar] [CrossRef]
- Chatauret, N.; Butterworth, R.F. Effects of liver failure on inter-organ trafficking of ammonia: Implications for the treatment of HE. J. Gastroenterol. Hepatol. 2004, 19, S219–S223. [Google Scholar] [CrossRef]
- Pistilli, E.E.; Bogdanovich, S.; Goncalves, M.D.; Ahima, R.S.; Lachey, J.; Seehra, J.; Khurana, T. Targeting the activin type IIB receptor to improve muscle mass and function in the mdx mouse model of Duchenne muscular dystrophy. Am. J. Pathol. 2011, 178, 1287–1297. [Google Scholar] [CrossRef]
- Amthor, W.M.; Hoogaars, M.H.W. Interference with myostatin/ActRIIB signaling as a therapeutic strategy for Duchenne muscular dystrophy. Curr. Gene Ther. 2012, 12, 245–259. [Google Scholar] [CrossRef]
- Burks, T.N.; Cohn, R.D. Role of TGF-β signaling in inherited and acquired myopathies. Skeletal Muscle 2011, 1, 19. [Google Scholar] [CrossRef] [Green Version]
- Han, H.Q.; Zhou, X.; Mitch, W.E.; Goldberg, A.L. Myostatin/activin pathway antagonism: Molecular basis and therapeutic potential. Int. J. Biochem. Cell Biol. 2013, 45, 2333–2347. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Benny Klimek, M.E.; Aydogdu, T.; Link, M.J.; Pons, M.; Koniaris, L.G.; Zimmers, T.A. Acute inhibition of myostatin-family proteins preserves skeletal muscle in mouse models of cancer cachexia. Biochem. Biophys. Res. Commun. 2010, 391, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, K.; Nakatani, M.; Uezumi, A.; Murakami, T.; Cui, X. Signal transduction pathways through activin receptors as therapeutic targets for musculoskeletal diseases and cancer. Endocr. J. 2008, 55, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haidet, A.M.; Rizo, L.; Handy, C.; Umapathi, P.; Eagle, A.; Shilling, C.; Boue, D.; Martin, P.T.; Sahenk, Z.; Mendell, J.R.; et al. Long-term enhancement of skeletal muscle mass and strength by single gene administration of myostatin inhibitors. Proc. Natl. Acad. Sci. USA 2008, 105, 4318–4322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmers, T.A.; Davies, M.V.; Koniaris, L.G.; Haynes, P.; Esquela, A.F.; Tomkinson, K.N.; McPherron, A.C.; Wolfman, N.M.; Lee, S.J. Induction of cachexia in mice by systemically administered myostatin. Science 2002, 296, 1486–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amthor, H.; Nicholas, G.; McKinnell, I.; Kemp, C.F.; Sharma, M.; Kambadur, R.; Patel, K. Follistatin complexes myostatin and antagonize myostatin-mediated inhibition of myogenesis. Dev. Biol. 2004, 270, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Swart, G.R.; van den Berg, J.W.; Wattimena, J.L.; Rietveld, T.; van Vuure, J.K.; Frenkel, M. Elevated protein requirements in cirrhosis of the liver were investigated by whole-body protein turnover studies. Clin. Sci. 1988, 75, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, D.; Colombi, M.; Moroni, C.; Hall, M.N. Rapamycin passes the torch, a new generation of mTOR inhibitors. Nat. Rev. Drug Discov. 2011, 10, 868–880. [Google Scholar] [CrossRef]
- Chen, J.; Alberts, I.; Li, X. Dysregulation of the IGF-I/PI3K/AKT/mTOR signaling pathway in autism spectrum disorders. Int. J. Dev. Neurosci. 2014, 35, 35–41. [Google Scholar] [CrossRef]
- Sandri, M. Signaling in muscle atrophy and hypertrophy. Physiology 2008, 23, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Burgos, S.A.; Chandurkar, V.; Tsoukas, M.A.; Chevalier, S.; Morais, J.A.; Lamarche, M.; Marliss, E.B. Insulin resistance of protein anabolism accompanies glucose metabolism in lean, glucose-tolerant offspring of persons with type 2 diabetes. BMJ Open Diabetes Res. Care 2016, 4, e000312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; White, M.F. Targeting Forkhead box O1 from the concept of metabolic diseases: Lessons from mouse models Antioxid. Redox. Signal. 2011, 14, 649–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milan, G.; Romanello, V.; Pescatore, F.; Armani, A.; Paik, J.H.; Frasson, L.; Seydel, A.; Zhao, J.; Abraham, R.; Goldberg, A.L.; et al. Regulation of autophagy and the ubiquitin-proteasome system by the FoxO transcriptional network during muscle atrophy. Nat. Commun. 2015, 6, 6670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, N.; Singh, N.; Joshi, Y.K. Nutrition in the management of hepatic encephalopathy. Trop. Gastroenterol. 2003, 24, 59–62. [Google Scholar]
- Shiraki, M.; Nishiguchi, S.; Saito, M.; Fukuzawa, Y.; Mizuta, T.; Kaibori, M.; Hanai, T.; Nishimura, K.; Shimizu, M.; Tsurumi, H.; et al. Nutritional status and quality of life in patients with liver cirrhosis as assessed in 2007. Hepatology 2013, 43, 106–112. [Google Scholar] [CrossRef]
- Zhang, S.; Zeng, X.; Ren, M.; Mao, X.; Qiao, S. Novel metabolic and physiological functions of branched chain amino acids: A review. J. Anim. Sci. Biotechnol. 2017, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Kalyani, R.R.; Corriere, M.; Ferrucci, L. Age-related and disease-related muscle loss: The effect of diabetes, obesity, and other diseases. Lancet Diabetes Endocrinol. 2014, 2, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Tajiri, K.; Shimizu, Y. Branched-chain amino acids in liver diseases. World J. Gastroenterol. 2013, 19, 7620–7629. [Google Scholar] [CrossRef]
- Sinclair, M.; Gow, P.J.; Grossmann, M.; Angus, P.W. Review article: Sarcopenia in cirrhosis etiology, implications, and potential therapeutic interventions. Aliment. Pharmacol. Ther. 2016, 43, 765–777. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.; Shi, H. Sex hormones and their receptors regulate liver energy homeostasis. Int. J. Endocrinol. 2015, 2015, 294278. [Google Scholar] [CrossRef]
- Sinclair, M.; Grossmann, M.; Gow, P.J.; Angus, P.W. Testosterone in men with advanced liver disease: Abnormalities and implications. J. Gastroenterol. Hepatol. 2015, 30, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Mowat, N.A.; Edwards, C.R.; Fisher, R.; McNeilly, A.S.; Green, J.R.; Dawson, A.M. Hypothalamic-pituitary-gonadal function in males with cirrhosis of the liver. Gut 1976, 17, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.J.; Wu, J.C.; Lee, S.D.; Tsai, Y.T.; Lo, K.J. Gonadal dysfunction and changes in sex hormones in postnecrotic cirrhotic men: A matched study with alcoholic cirrhotic men. Hepato Gastroenterol. 1991, 38, 531–534. [Google Scholar]
- Sinclair, M. Controversies in Diagnosing Sarcopenia in Cirrhosis-Moving from Research to Clinical Practice. Nutrients 2019, 11, 2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guglielmi, G.; Ponti, F.; Agostini, M.; Amadori, M.; Battista, G.; Bazzocchi, A. The role of DXA in sarcopenia. Aging Clin. Exp. Res. 2016, 28, 1047–1060. [Google Scholar]
- Bera, T.K. Bioelectrical impedance methods for noninvasive health monitoring: A review. J. Med. Eng. 2014, 2014, 381251. [Google Scholar] [CrossRef] [Green Version]
- Anand, A.C. Nutrition and muscle in cirrhosis. J. Clin. Exp. Hepatol. 2017, 7, 340–357. [Google Scholar] [CrossRef]
- Di Renzo, L.; Gratteri, S.; Sarlo, F.; Cabibbo, A.; Colica, C.; De Lorenzo, A. Individually tailored screening of susceptibility to sarcopenia using p53 codon 72 polymorphism, phenotypes, and conventional risk factors. Dis. Markers 2014, 2014. [Google Scholar] [CrossRef]
- Meng, Y.; Wu, H.; Yang, Y.; Du, H.; Xia, Y.; Guo, X.; Liu, X.; Li, C.; Niu, K. Relationship of anabolic and catabolic biomarkers with muscle strength and physical performance in older adults: A population-based cross-sectional study. BMC Musculoskelet. Disord. 2015, 16, 202. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, D.R. Laboratory tests and nutritional assessment. Protein-energy status. Pediatr. Clin. N. Am. 1989, 36, 139–161. [Google Scholar] [CrossRef]
- Ginès, P.; Guevara, M.; Arroyo, V.; Rodés, J. Hepatorenal syndrome. Lancet 2003, 362, 1819–1827. [Google Scholar] [CrossRef]
- Fukuzawa, Y.; Yoshitomi, A.; Morita, S.; Shiraki, M.; Kato, A.; Haruta, J.; Murakami, M.; Taku, K. Report from the 22nd Tokai Chapter Educational Seminar: Nutrition Care in Internal Medicine. Nihon Naika Gakkai Zasshi 2017, 106, 133–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandai, S.; Furukawa, S.; Kodaka, M.; Hata, Y.; Mori, T.; Nomura, N.; Ando, F.; Mori, Y.; Takahashi, D.; Yoshizaki, Y.; et al. Loop diuretics affect skeletal myoblast differentiation and exercise-induced muscle hypertrophy. Sci. Rep. 2017, 7, 46369. [Google Scholar] [CrossRef] [Green Version]
- Hanai, T.; Shiraki, M.; Miwa, T.; Watanabe, S.; Imai, K.; Suetsugu, A.; Takai, K.; Moriwaki, H.; Shimizu, M. Effect of loop diuretics on skeletal muscle depletion in patients with liver cirrhosis. Hepatol. Res. 2019, 49, 82–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, D.L.; Baumgartner, R.N. Sarcopenia and obesity. Clin. Geriatr. Med. 2011, 27, 401–421.50. [Google Scholar] [CrossRef]
- Park, S.W.; Goodpaster, B.H.; Strotmeyer, E.S.; Kuller, L.H.; Broudeau, R.; Kammerer, C.; De Rekeneire, N.; Harris, T.B.; Schwartz, A.V.; Tylavsky, F.A. Health, aging, and body composition studies. Accelerated loss of skeletal muscle strength in older adults with type 2 diabetes: The health, aging, and body composition study. Diabetes Care 2007, 30, 1507–1512. [Google Scholar] [CrossRef] [Green Version]
- Petroni, M.L.; Caletti, M.T.; Dalle Grave, R.; Bazzocchi, A.; Aparisi Gómez, M.P.; Marchesini, G. Prevention and treatment of sarcopenic obesity in women. Nutrients 2019, 11, 1302. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, D.J.; Erskine, R.M.; Morse, C.I.; Winwood, K.; Onambélé-Pearson, G. The impact of obesity on skeletal muscle strength and structure from adolescence to old age. Biogerontology 2016, 17, 467–483. [Google Scholar] [CrossRef] [Green Version]
- Picca, A.; Fanelli, F.; Calvani, R.; Mulè, G.; Pesce, V.; Sisto, A.; Pantanelli, C.; Bernabei, R.; Landi, F.; Marzetti, E. Gut dysbiosis and muscle aging: Searching for novel targets against sarcopenia. Mediators Inflamm. 2018, 2018, 7026198. [Google Scholar] [CrossRef]
- Picca, A.; Lezza, A.M.S.; Leeuwenburgh, C.; Pesce, V.; Calvani, R.; Bossola, M.; Manes-Gravina, E.; Landi, F.; Bernabei, R.; Marzetti, E. Circulating mitochondrial DNA at the crossroads of mitochondrial dysfunction and inflammation during aging and muscle wasting disorders. Rejuv. Res. 2017, 21, 350–359. [Google Scholar] [CrossRef]
- Haider, M.; Haider, S.Q. Assessment of protein-calorie malnutrition. Clin. Chem. 1984, 30, 1286–1299. [Google Scholar] [CrossRef] [PubMed]
- Mtaweh, H.; Tuira, L.; Floh, A.A.; Parshuram, C.S. Indirect calorimetry: History, technology, and application. Front. Pediatr. 2018, 6, 257. [Google Scholar] [CrossRef] [PubMed]
- Eslamparast, T.; Vandermeer, B.; Raman, M.; Gramlich, L.; Den Heyer, V.; Belland, D.; Ma, M.; Tandon, P. Are predictive energy expenditure equations accurate for cirrhosis? Nutrients 2019, 11, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Text Book of Nutrition Support Team Japan Society of Metabolism and Clinical Nutrition; Nankodo: Tokyo, Japan, 2017.
- Hanai, T.; Shiraki, M.; Nishimura, K.; Imai, K.; Suetsugu, A.; Takai, K.; Shimizu, M.; Naiki, T.; Moriwaki, H. Free fatty acids are markers of energy malnutrition in liver cirrhosis. Hepatol. Res. 2014, 44, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Krähenbühl, L.; Lang, C.; Lüdes, S.; Seiler, C.; Schäfer, M.; Zimmermann, A.; Krähenbühl, S. Reduced hepatic glycogen stores in patients with liver cirrhosis. Liver Int. 2003, 23, 101–109. [Google Scholar] [CrossRef]
- Japanese Society for Parenteral and Enteral Nutrition: Intravenous Enteral Nutrition Guidelines Version 3, Liver Disease; Japanese Society for Parenteral and Enteral Nutrition: Tokyo, Japan, 2013; p. 248.
- Park, J.G.; Tak, W.Y.; Park, S.Y.; Kweon, Y.O.; Jang, S.Y.; Lee, Y.R.; Bae, S.H.; Jang, J.Y.; Kim, D.Y.; Lee, J.S.; et al. Effects of branched-chain amino acids on the progression of advanced liver disease: A Korean nationwide, multicenter, retrospective, observational, cohort study. Medicine 2017, 96, e6580. [Google Scholar] [CrossRef]
- Tsien, C.D.; McCullough, A.J.; Dasarathy, S. Late evening snack: Exploiting a period of anabolic opportunity in cirrhosis. J. Gastroenterol. Hepatol. 2012, 27, 430–441. [Google Scholar] [CrossRef]
- Houston, D.K.; Nicklas, B.J.; Ding, J.; Harris, T.B.; Tylavsky, F.A.; Newman, A.B.; Lee, J.S.; Sahyoun, N.R.; Visser, M.; Kritchevsky, S.B.; et al. Health ABC Study. Dietary protein intake is associated with lean mass change in older, community-dwelling adults: The Health, Aging, and Body Composition (Health ABC) Study. Am. J. Clin. Nutr. 2008, 87, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, T.; Izumi, N.; Charlton, M.R.; Sata, M. Branched-chain amino acids as pharmacological nutrients in chronic liver disease. Hepatology 2011, 54, 1063–1070. [Google Scholar] [CrossRef]
- Kobayashi, H.; Kato, H.; Hirabayashi, Y.; Murakami, H.; Suzuki, H. Modulation of muscle protein metabolism by branched-chain amino acids in normal and muscle-atrophying rats. J. Nutr. 2006, 136, 234S–236S. [Google Scholar] [CrossRef] [Green Version]
- Jackman, S.R.; Witard, O.C.; Philp, A.; Wallis, G.A.; Baar, K.; Tipton, K.D. Branched-chain amino acid ingestion stimulates muscle myofibrillar protein synthesis following resistance exercise in humans. Front. Physiol. 2017, 8, 390. [Google Scholar] [CrossRef] [PubMed]
- Bear, D.E.; Langan, A.; Dimidi, E.; Wandrag, L.; Harridge, S.D.R.; Hart, N.; Connolly, B.; Whelan, K. β-Hydroxy-β-methylbutyrate and its impact on skeletal muscle mass and physical function in clinical practice: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2019, 109, 1119–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holeček, M. Beta-hydroxy-beta-methylbutyrate supplementation, and skeletal muscle in healthy and muscle-wasting conditions. J. Cachexia Sarcopenia Muscle 2017, 8, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Black, S.M. Carnitine Homeostasis, Mitochondrial Function, and Cardiovascular Disease. Drug Discov. Today Dis. Mech. 2009, 6, e31–e39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceglia, L. Vitamin D and its role in skeletal muscle. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 628–633. [Google Scholar] [CrossRef] [Green Version]
- Ochi, E.; Tsuchiya, Y. Eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA) in muscle damage and function. Nutrients 2018, 10, 552. [Google Scholar] [CrossRef] [Green Version]
- Gorjao, R.; Dos Santos, C.M.M.; Serdan, T.D.A.; Diniz, V.L.S.; Alba-Loureiro, T.C.; Cury-Boaventura, M.F.; Hatanaka, E.; Levada-Pires, A.C.; Sato, F.T.; Pithon-Curi, T.C.; et al. New insights into the regulation of cancer cachexia by N-3 polyunsaturated fatty acids. Pharmacol. Ther. 2019, 196, 117–134. [Google Scholar] [CrossRef]
- Rodacki, C.L.; Rodacki, A.L.; Pereira, G.; Naliwaiko, K.; Coelho, I.; Pequito, D.; Fernandes, L.C. Fish-oil supplementation enhances the effects of strength training in elderly women. Am. J. Clin. Nutr. 2012, 95, 428–436. [Google Scholar] [CrossRef]
- Nishikawa, H.; Enomoto, H.; Yoh, K.; Iwata, Y.; Sakai, Y.; Kishino, K.; Ikeda, N.; Takashima, T.; Aizawa, N.; Takata, R.; et al. Serum zinc concentration and sarcopenia: A close linkage in chronic liver diseases. J. Clin. Med. 2019, 8, 336. [Google Scholar] [CrossRef] [Green Version]
- Kodama, H.; Tanaka, M.; Naito, Y.; Katayama, K.; Moriyama, M. Japan’s practical guidelines for zinc deficiency with a particular focus on taste disorders, inflammatory bowel disease, and liver cirrhosis. Int. J. Mol. Sci. 2020, 22, 2941. [Google Scholar] [CrossRef]
- López-Hortas, L.; Pérez-Larrán, P.; González-Muñoz, M.J.; Falqué, E.; Domínguez, H. Recent developments in the extraction and application of UA. A review. Food Res. Int. 2018, 103, 130–149. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, S.D.; Elmore, C.J.; Bongers, K.S.; Ebert, S.M.; Fox, D.K.; Dyle, M.C.; Bullard, S.A.; Adams, C.M. Ursolic acid increases skeletal muscle and brown fat and decreases diet-induced obesity, glucose intolerance, and fatty liver disease. PLoS ONE 2012, 7, e39332.80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasarathy, S. Consilience in sarcopenia of cirrhosis. J. Cachexia Sarcopenia Muscle 2012, 3, 225–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajaj, J.S.; O’Leary, J.G.; Tandon, P.; Wong, F.; Kamath, P.S.; Biggins, S.W.; Garcia-Tsao, G.; Lai, J.; Fallon, M.B.; Thuluvath, P.J.; et al. Targets to improve quality of care for patients with HE: Data from a multicenter cohort. Aliment Pharmacol. Ther. 2019, 49, 1518–1527. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Davuluri, G.; Silva, R.N.E.; Engelen, M.P.K.J.; Ten Have, G.A.M.; Prayson, R.; Deutz, N.E.; Dasarathy, S. Ammonia lowering reverses the sarcopenia of cirrhosis by restoring skeletal muscle proteostasis. Hepatology 2017, 65, 2045–2058. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.L.; Lai, H.C.; Yeh, S.; Cai, X.; Chang, C. Androgen receptor roles in hepatocellular carcinoma, fatty liver, cirrhosis, and hepatitis. Endocr. Relat. Cancer 2014, 21, R165–R182. [Google Scholar] [CrossRef] [Green Version]
- Buch, S.C.; Kondragunta, V.; Branch, R.A.; Carr, B.I. Gender-based outcomes differences in unresectable hepatocellular carcinoma. Hepatol. Int. 2008, 2, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, M.; Grossmann, M.; Hoermann, R.; Angus, P.W.; Gow, P.J. Testosterone therapy increases muscle mass in men with cirrhosis and low testosterone testosterone: A randomized controlled trial. J. Hepatol. 2016, 65, 906–913. [Google Scholar] [CrossRef]
- Moreno, M.; Chaves, J.F.; Sancho-Bru, P.; Ramalho, F.; Ramalho, L.N.; Mansego, M.L.; Ivorra, C.; Dominguez, M.; Conde, L.; Millán, C.; et al. Ghrelin attenuates hepatocellular injury and liver fibrogenesis in rodents and influences fibrosis progression in humans. Hepatology 2010, 51, 974–985. [Google Scholar] [CrossRef] [Green Version]
- American College of Sports Medicine Geoffrey; Moore, J.L.; Durstine, P. Painter ACSMs Exercise Management for Persons with Chronic Diseases and Disabilities, 4th ed.; Human Kinetics: Champaign, IL, USA, 2016. [Google Scholar]
- Biolo, G.; Tipton, K.D.; Klein, S.; Wolfe, R.R. An abundant supply of amino acids enhances the metabolic effect of exercise on muscle proteins. Am. J. Physiol. 1997, 273, E122–E1229. [Google Scholar] [CrossRef]
- Fiatarone, M.A.; O’Neill, E.F.; Ryan, N.D.; Clements, K.M.; Solares, G.R.; Nelson, M.E.; Roberts, S.B.; Kehayias, J.J.; Lipsitz, L.A.; Evans, W.J. Exercise training and nutritional supplementation for physical frailty in very elderly people. N. Engl. J. Med. 1994, 330, 1769–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratamess, N.A.; Kraemer, W.J.; Volek, J.S.; Maresh, C.M.; Vanheest, J.L.; Sharman, M.J.; Rubin, M.R.; French, D.N.; Vescovi, J.D.; Silvestre, R.; et al. Androgen receptor content following heavy resistance exercise in men J. Steroid Biochem. Mol. Biol. 2005, 93, 35–42. [Google Scholar]
- Schoenfeld, B.J.; Contreras, B.; Krieger, J.; Grgic, J.; Delcastillo, K.; Belliard, R.; Alto, A. Resistance training volume enhances muscle hypertrophy but not strength in trained men. Med. Sci. Sports Exerc. 2018, 51, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, B.J.; Grgic, J.; Ogborn, D.; Krieger, J.W. Strength and hypertrophy adaptations between low- vs. high-load resistance training: A systematic review and meta-analysis. J. Strength Cond. Res. 2017, 31, 3508–3523. [Google Scholar] [CrossRef]
- Kaido, T.; Tamai, Y.; Hamaguchi, Y.; Okumura, S.; Kobayashi, A.; Shirai, H.; Yagi, S.; Kamo, N.; Hammad, A.; Inagaki, N.; et al. Effects of pretransplant sarcopenia and sequential changes in sarcopenic parameters after living donor liver transplantation. Nutrition 2017, 33, 195–198. [Google Scholar] [CrossRef]
- Bergerson, J.T.; Lee, J.G.; Furlan, A.; Sourianarayanane, A.; Fetzer, D.T.; Tevar, A.D.; Landsittel, D.P.; DiMartini, A.F.; Dunn, M.A. Liver transplantation arrests and reverses muscle wasting. Clin. Transplant. 2015, 29, 216–221. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Classification of Sarcopenia | Causes |
|---|---|
| Primary sarcopenia | |
| Age-related | No causes other than aging |
| Secondary sarcopenia | |
| Activity-related | A bedridden state or weightlessness |
| Disease-related | Severe organ (heart, lung, liver, kidney, or brain) dysfunction, inflammatory disease, or malignant tumor |
| Nutrition-related | Energy and protein deficiencies associated with malabsorption, gastrointestinal disease, and/or medication |
| Parameter | Criteria for Diagnosing Sarcopenia |
|---|---|
| Muscle mass at L3 measured with CT | Men: 42 cm2/m2 |
| Women: 38 cm2/m2 | |
| SMI BIA | Men: 7.0 kg/m2 |
| Women: 5.7 kg/m2 | |
| Grip strength | Men: <26 kg |
| Women: <18 kg |
| Factor | The Key Mechanism | References |
|---|---|---|
| Renal Dysfunction-associated Liver Cirrhosis | Restriction of Protein Intake | [52,53] |
| Loop Diuretic Sarcopenia | Regulation for Myoblast Differentiation | [54,55] |
| Sarcopenic Obesity | Qualitative Decline in Muscle Mass for Insulin Resistance | [56,57,58,59] |
| Enterobacteria | Mitochondrial Dysfunction for Disturbance of Intestinal Flora | [60,61] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamimura, H.; Sato, T.; Natsui, K.; Kobayashi, T.; Yoshida, T.; Kamimura, K.; Tsuchiya, A.; Murayama, T.; Yokoyama, J.; Kawai, H.; et al. Molecular Mechanisms and Treatment of Sarcopenia in Liver Disease: A Review of Current Knowledge. Int. J. Mol. Sci. 2021, 22, 1425. https://doi.org/10.3390/ijms22031425
Kamimura H, Sato T, Natsui K, Kobayashi T, Yoshida T, Kamimura K, Tsuchiya A, Murayama T, Yokoyama J, Kawai H, et al. Molecular Mechanisms and Treatment of Sarcopenia in Liver Disease: A Review of Current Knowledge. International Journal of Molecular Sciences. 2021; 22(3):1425. https://doi.org/10.3390/ijms22031425
Chicago/Turabian StyleKamimura, Hiroteru, Takeki Sato, Kazuki Natsui, Takamasa Kobayashi, Tomoaki Yoshida, Kenya Kamimura, Atsunori Tsuchiya, Toshiko Murayama, Junji Yokoyama, Hirokazu Kawai, and et al. 2021. "Molecular Mechanisms and Treatment of Sarcopenia in Liver Disease: A Review of Current Knowledge" International Journal of Molecular Sciences 22, no. 3: 1425. https://doi.org/10.3390/ijms22031425