
Glyphosate and AMPA Induce Alterations in Expression of Genes Involved in Chromatin Architecture in Human Peripheral Blood Mononuclear Cells (In Vitro)

 , , , ,
, , , ,
Abstract
:
1. Introduction
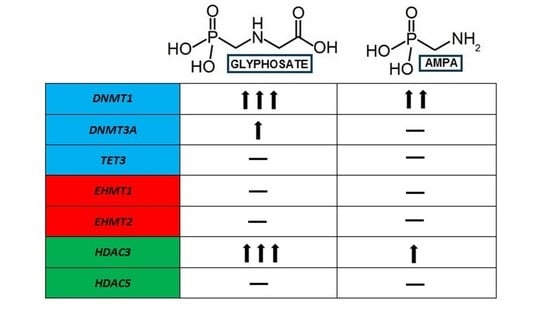
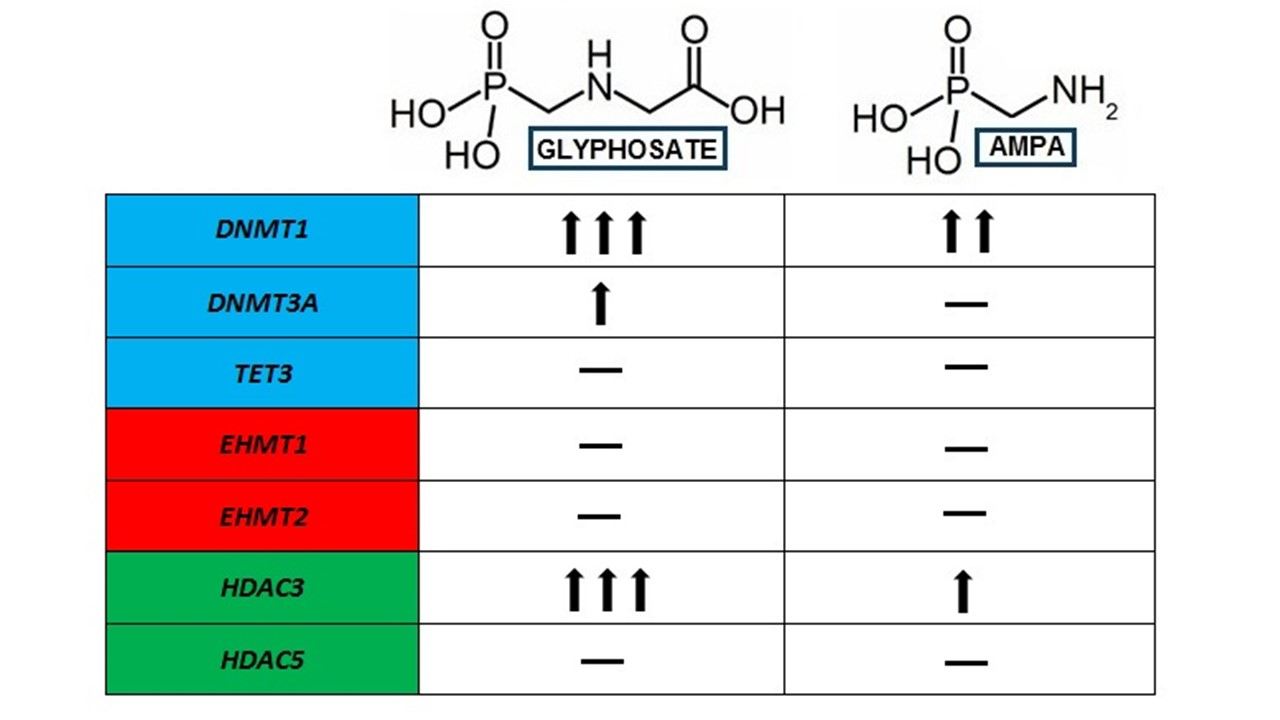
2. Results
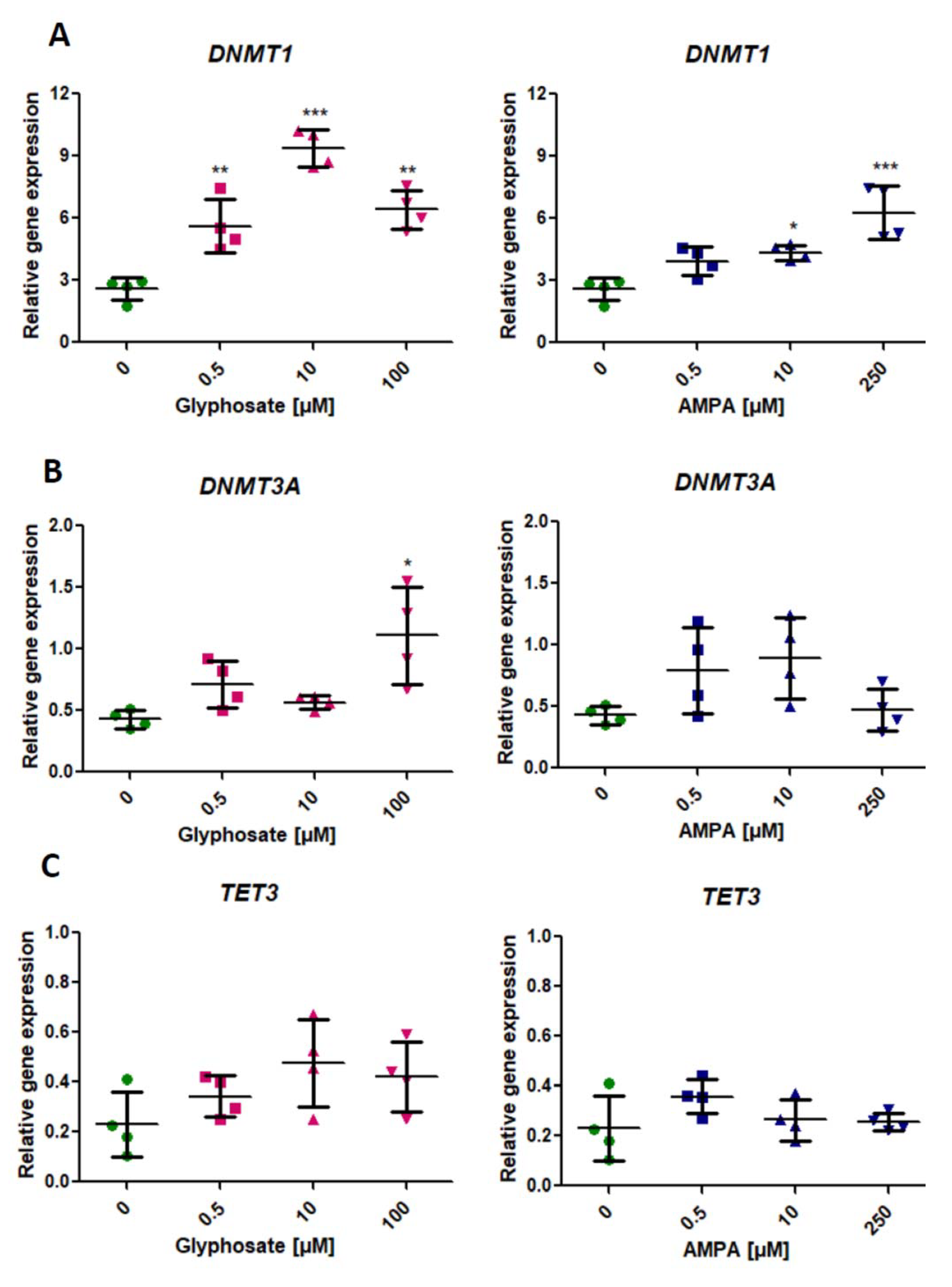
2.1. Analysis of Expression of Genes Involved in the DNA Methylation (DNMT1, DNMT3A) and DNA Demethylation Process (TET3)
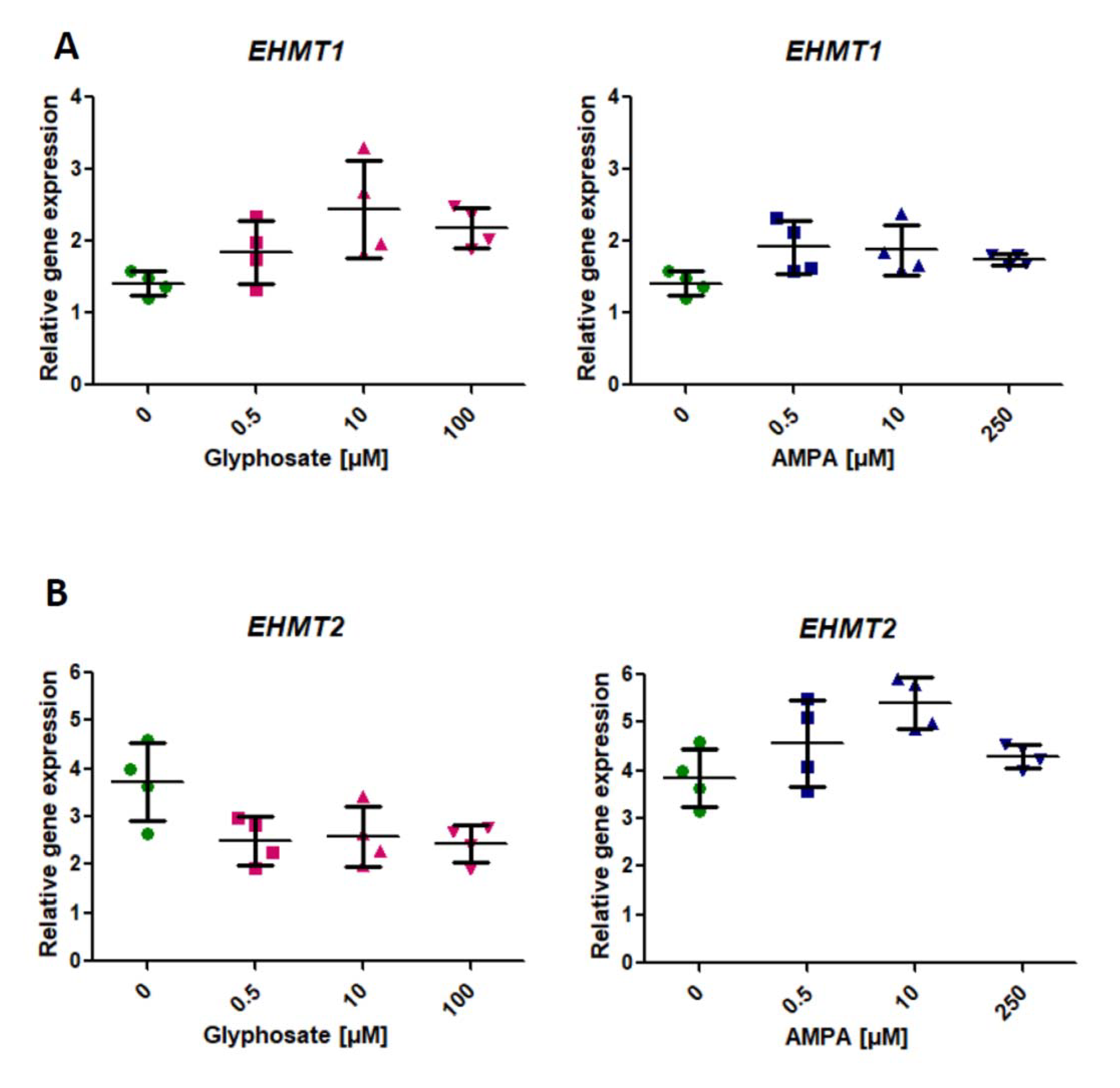
2.2. Analysis of the Expression of Genes Involved in the Modification of Histone Methylation (EHMT1, EHMT)
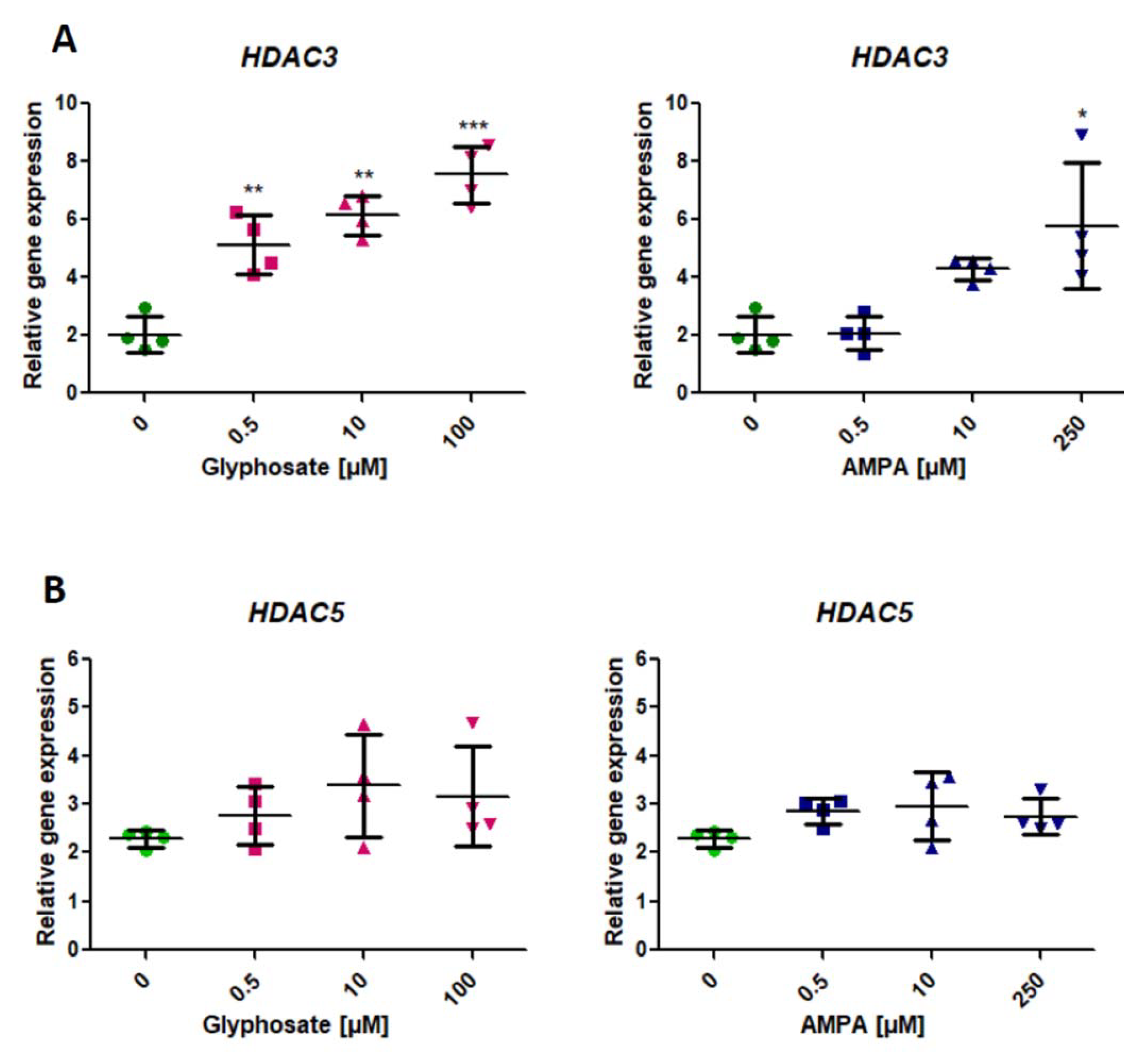
2.3. Analysis of Expression of Genes Involved in the Modification of Histone Deacetylation (HDAC3, HDAC5) (HDAC3, HDAC5)
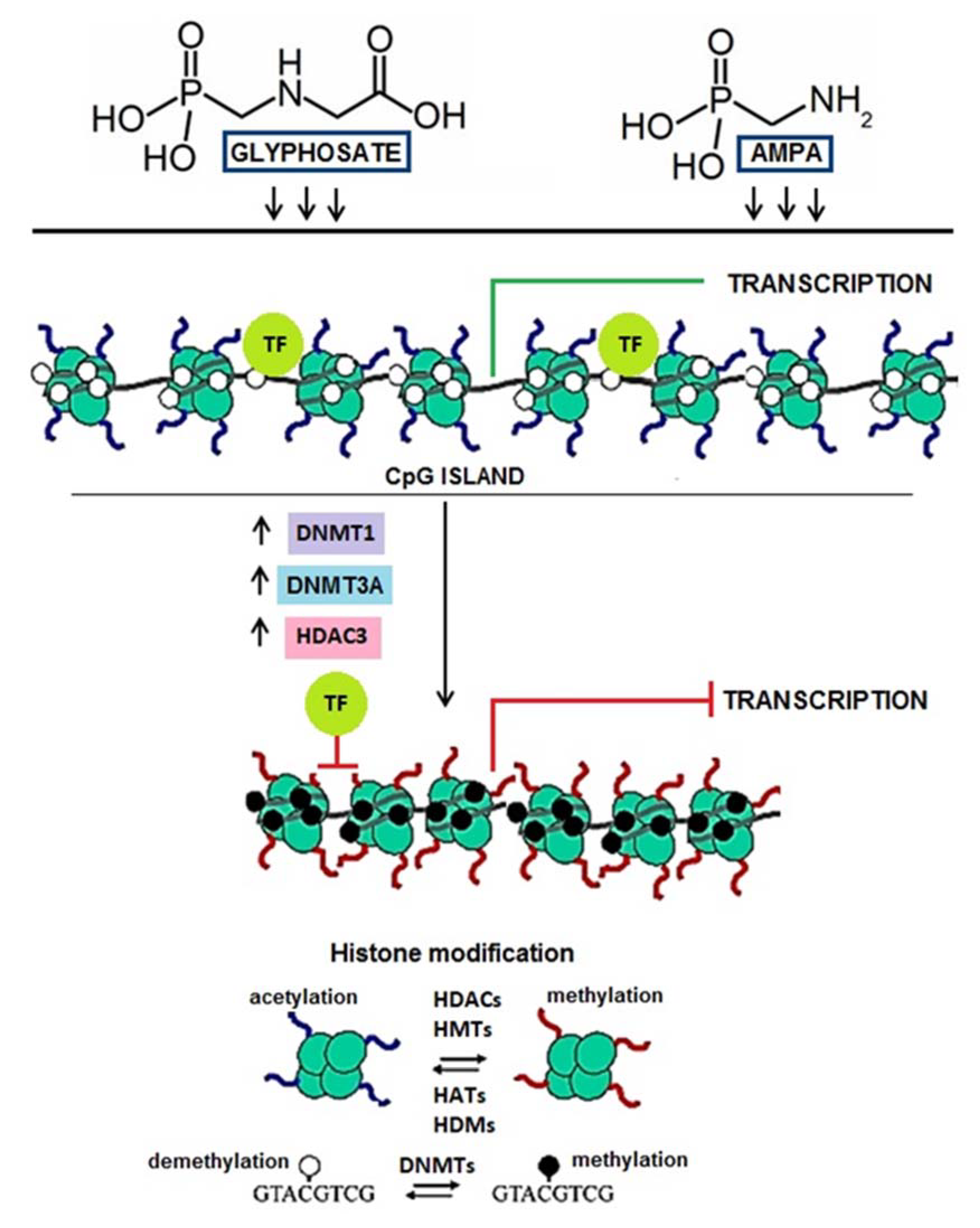
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cells Isolation
4.3. Cells Treatment
4.4. Gene Expression
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMPA | Aminomethylphosphonic acid |
| DNMT1 | DNA methyltransferase |
| DNMT3A | DNA methyltransferase 3 alpha |
| ECHA | European Chemicals Agency |
| EFSA | European Food Safety Authority |
| EHMT1 | Euchromatic histone lysine methyltransferase 1 |
| EHMT2 | Euchromatic histone lysine methyltransferase 2 |
| GMOs | Genetically modified crops |
| GBHs | Glyphosate-based herbicides (GBHs) |
| HBM4EU | Human Biomonitoring for Europe initiative |
| HDAC3 | Histone deacetylase 3 |
| HDAC5 | Histone deacetylase 5 |
| IARC | International Agency for Research on Cancer |
| PMBCs | Human peripheral blood mononuclear cells |
| TET1 | Tet methylcytosine dioxygenase 1 |
| TET3 | Tet methylcytosine dioxygenase 3 |
| TF | Transcription factor binding |
| US EPA | United State Environmental Protection Agency |
| WHO | World Human Organization |
References
- Zhang, L.; Rana, J.; Shaffer, R.M.; Taioli, E.; Sheppard, L. Exposure to glyphosate-based herbicides and risk for non-Hodgkin lymphoma: A meta-analysis and supporting evidence. Mutat. Res. 2019, 781, 186–206. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, B.L.; de Oliveira, R.; Abe, F.R.; Brito, L.B.; Moura, D.S.; Valadares, M.C.; Grisolia, C.K.; de Oliveira, D.P.; de Oliveira, G.A.R. Ecotoxicological assessment of glyphosate-based herbicides: Effects on different organisms. Environ. Toxicol. Chem. 2017, 36, 1755–1763. [Google Scholar] [CrossRef] [PubMed]
- Mesnage, R.; Defarge, N.; de Vendômois, J.S.; Séralini, G.E. Potential toxic effects of glyphosate and its commercial formulations below regulatory limits. Food Chem. Toxicol. 2015, 84, 133–153. [Google Scholar] [CrossRef] [Green Version]
- Roy, N.M.; Carneiro, B.; Ochs, J. Glyphosate induces neurotoxicity in zebrafish. Environ. Toxicol. Pharmacol. 2016, 42, 45–54. [Google Scholar] [CrossRef]
- Myers, J.P.; Antoniou, M.N.; Blumberg, B.; Carroll, L.; Colborn, T.; Everett, L.G.; Hansen, M.; Landrigan, P.J.; Lanphear, B.P.; Mesnage, R.; et al. Concerns over use of glyphosate-based herbicides and risks associated with exposures: A consensus statement. Environ. Health 2016, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef] [Green Version]
- HBM4EU Priority Substances. Available online: https://www.hbm4eu.eu/the-substances/ (accessed on 10 February 2021).
- Connolly, A.; Coggins, M.A.; Koch, H.M. Human biomonitoring of glyphosate exposures: State-of-the-art and future research challenges. Toxics 2020, 18, 60. [Google Scholar] [CrossRef]
- Soukup, S.T.; Merz, B.; Bub, A.; Hoffman, I.; Watzl, B.; Steinberg, P.; Kulling, S.E. Glyphosate and AMPA levels in human urine samples and their correlation with food consumption: Results of the cross-sectional KarMeN study in Germany. Arch. Toxicol. 2020, 94, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Aris, A.; Leblanc, S. Maternal and fetal exposure to pesticides associated to genetically modified foods in eastern townships of Quebec, Canada. Reprod. Toxicol. 2011, 31, 528–533. [Google Scholar] [CrossRef]
- Zouaoui, K.; Dulaurent, S.; Gaulier, J.M.; Moesch, C.; Lachâtre, G. Determination of glyphosate and AMPA in blood and urine from humans: About 13 cases of acute intoxication. Forensic Sci. Int. 2013, 226, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Faniband, M.H.; Norén, E.; Littorin, M.; Lindh, C.H. Human experimental exposure to glyphosate and biomonitoring of young Swedish adults. Int. J. Hyg. Environ. Health 2021, 231, 113657. [Google Scholar] [CrossRef] [PubMed]
- Patkin, E.L.; Sofronov, H.A. Population epigenetics, ecotoxicology and human diseases. Rus. J. Genet. App. Res. 2012, 10, 14–28. [Google Scholar] [CrossRef] [Green Version]
- Kubsad, D.; Nilsson, E.E.; King, S.E.; Sadler-Riggleman, I.; Beck, D.; Skinner, M.K. Assessment of Glyphosate Induced Epigenetic Transgenerational Inheritance of Pathologies and Sperm Epimutations: Generational Toxicology. Sci. Rep. 2019, 9, 6372. [Google Scholar] [CrossRef] [Green Version]
- Ben Maamar, M.; Beck, D.; Nilsson, E.E.; Kubsad, D.; Skinner, M.K. Epigenome-wide association study for glyphosate induced transgenerational sperm DNA methylation and histone retention epigenetic biomarkers for disease. Epigenetics 2020, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, D.; Lopes Alderete, B.; de Souza, C.T.; Ferraz Dias, J.; Niekraszewicz, L.; Cappetta, M.; Martínez-López, W.; Da Silva, J. DNA damage and epigenetic alteration in soybean farmers exposed to complex mixture of pesticides. Mutagenesis 2018, 33, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duforestel, M.; Nadaradjane, A.; Bougras-Cartron, G.; Briand, J.; Olivier, C.; Frenel, J.S.; Vallette, F.; Lelièvre, S.; Cartron, P.F. Glyphosate Primes Mammary Cells for Tumorigenesis by Reprogramming the Epigenomeina TET3-Dependent Manner. Front. Genet. 2019, 10, 885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahl, V.F.S.; Dhillon, V.; Fenech, M.; de Souza, M.R.; da Silva, F.N.; Marroni, N.A.P.; Nunes, E.A.; Cerchiaro, G.; Pedron, T.; Batista, B.L.; et al. Occupational Exposure to Pesticides in Tobacco Fields: The Integrated Evaluation of Nutritional Intake and Susceptibility on Genomic and Epigenetic Instability. Oxid. Med. Cell Longev. 2018, 2018, 7017423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiatkowska, M.; Reszka, E.; Woźniak, K.; Jabłońska, E.; Michałowicz, J.; Bukowska, B. DNA damage and methylation induced by glyphosate in human peripheral blood mononuclear cells (in vitro study). Food Chem. Toxicol. 2017, 105, 93–98. [Google Scholar] [CrossRef]
- Woźniak, E.; Reszka, E.; Jabłońska, E.; Balcerczyk, A.; Broncel, M.; Bukowska, B. Glyphosate affects methylation in the promoter regions of selected tumor suppressors as well as expression of major cell cycle and apoptosis drivers in PBMCs (in vitro study). Toxicol. Vitro 2020, 63, 104736. [Google Scholar] [CrossRef]
- Woźniak, E.; Reszka, E.; Jabłońska, E.; Mokra, K.; Balcerczyk, A.; Huras, B.; Zakrzewski, J.; Bukowska, B. The selected epigenetic effects of aminomethylphosphonic acid, a primary metabolite of glyphosate on human peripheral blood mononuclear cells (in vitro). Toxicol. Vitro 2020, 30, 104878. [Google Scholar] [CrossRef]
- Kwiatkowska, M.; Michałowicz, J.; Jarosiewicz, P.; Pingot, D.; Sicińska, P.; Huras, B.; Zakrzewski, J.; Jarosiewicz, M.; Bukowska, B. Evaluation of apoptotic potential of glyphosate metabolites and impurities in human peripheral blood mononuclear cells (in vitro study). Food Chem. Toxicol. 2020, 135, 110888. [Google Scholar] [CrossRef]
- Marsit, C.J. Influence of environmental exposure on human epigenetic regulation. J. Exp. Biol. 2015, 1, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Nair, N.; Shoaib, M.; Sorenson, C.S. Chromatin Dynamics in Genome Stability: Roles in Suppressing Endogenous DNA Dam-age and Facilitating DNA Repair. Int. J. Mol. Sci. 2017, 18, 1486. [Google Scholar] [CrossRef] [Green Version]
- Takata, H.; Hanafusa, T.; Mori, T.; Shimura, M.; Iida, Y.; Ishikawa, K.; Yoshikawa, K.; Yoshikawa, Y.; Maeshima, K. Chromatin Compaction Protects Genomic DNA from Radiation Damage. PLoS ONE 2013, 8, e75622. [Google Scholar] [CrossRef] [Green Version]
- Woźniak, E.; Sicińska, P.; Michałowicz, J.; Woźniak, K.; Reszka, E.; Huras, B.; Zakrzewski, J.; Bukowska, B. The mechanism of DNA damage induced by Roundup 360 PLUS, glyphosate and AMPA in human peripheral blood mononuclear cells—Genotoxic risk assessement. Food Chem. Toxicol. 2018, 120, 510–522. [Google Scholar] [CrossRef]
- Kim, M.; Costello, J. DNA methylation: An epigenetic mark of cellular memory. Exp. Mol. Med. 2017, 49, e322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Liu, S.; Hu, Y.; Fang, L.; Gao, Y.; Xia, H.; Schroeder, S.G.; Rosen, B.D.; Connor, E.E.; Li, C.; et al. Comparative whole genome DNA methylation profiling across cattle tissues reveals global and tissue-specific methylation patterns. BMC Biol. 2020, 18, 85. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, M. DNA hypermethylation in disease: Mechanisms and clinical relevance. Epigenetics 2019, 14, 1141–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coronado-Posada, N.; Olivero-Verbel, J. In silico evaluation of pesticides as potential modulators of human DNA methyltransferases. SAR QSAR Environ. Res. 2019, 30, 865–878. [Google Scholar] [CrossRef]
- Smith, C.M.; Vera, M.K.M.; Bhandari, R.K. Developmental and epigenetic effects of Roundup and glyphosate exposure on Japanese medaka (Oryzias latipes). Aquat. Toxicol. 2019, 210, 215–226. [Google Scholar] [CrossRef]
- Ghosh, K.; Chatterjee, B.; Jayaprasad, A.G.; Kanade, S.R. The persistent organochlorine pesticide endosulfan modulates multiple epigenetic regulators with oncogenic potential in MCF-7 cells. Sci. Total Environ. 2018, 15, 1612–1622. [Google Scholar] [CrossRef]
- Cheng, Y.; He, C.; Wang, M.; Ma, X.; Mo, F.; Yang, S.; Han, J.; Wei, X. Targeting epigenetic regulators for cancer therapy: Mechanisms and advances in clinical trials. Signal Transduct. Target. Ther. 2019, 4, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piyathilake, C.J.; Badiga, S.; Borak, S.G.; Weragoda, J.; Bae, S.; Matthews, R.; Bell, W.C.; Partridge, E.E. A higher degree of expression of DNA methyl transferase 1 in cervical cancer is associated with poor survival outcome. Int. J. Women’s Health 2017, 9, 413–420. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.J.; Huang, F.X.; Sun, Z.T.; Zhang, R.X.; Huang, S.F.; Wang, J. Stat3 inhibits Beclin 1 expression through recruitment of HDAC3 in nonsmall cell lung cancer cells. Tumor Biol. 2014, 35, 7097–7103. [Google Scholar] [CrossRef]
- Chen, G.; Gu, Y.; Han, P.; Li, Z.; Zhao, J.L.; Gao, M.Z. Long noncoding RNA SBF2-AS1 promotes colorectal cancer proliferation and invasion by inhibiting miR-619–5p activity and facilitating HDAC3 expression. J. Cell Physiol. 2019, 234, 18688–18696. [Google Scholar] [CrossRef] [PubMed]
- Bayat, S.; Derakhshan, S.M.; Derakhshan, N.M.; Khaniani, M.S.; Alivand, M.R. Downregulation of HDAC2 and HDAC3 via oleurope in as a potent prevention and therapeutic agent in MCF-7 breast cancer cells. J. Cell Biochem. 2019, 120, 9172–9180. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.; Hsieh, T.H.; Halicka, H.D.; Darzynkiewicz, Z.; Wu, J.M. Upregulation of PD L1 expression by resveratrol and piceatannol in breast and colorectal cancer cells occurs via HDAC3/p300 mediated NF κB signaling. Int. J. Oncol. 2018, 53, 1469–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Rica, L.; Rodríguez-Ubreva, J.; García, M.; Islam, A.B.M.M.K.; Urquiza, J.M.; Hernando, H.; Christensen, J.; Helin, K.; Gómez-Vaquero, C.; Ballestar, E. PU.1 target genes undergo Tet2-coupled demethylation and DNMT3b-mediated methylation in monocyte-to-osteoclast differentiation. Genome Biol. 2013, 14, R99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seritrakul, P.; Gross, J.M. Tet-mediated DNA hydroxymethylation regulates retinal neurogenesis by modulating cell-extrinsic signaling pathways. PLoS Genet. 2017, 13, e1006987. [Google Scholar] [CrossRef]
- Liu, X.; Lu, H.; Chen, T.; Nallaparaju, K.C.; Yan, X.; Tanaka, S.; Ichiyama, K.; Zhang, X.; Zhang, L.; Wen, X.; et al. Genome-wide analysis identifies Bcl6-controlled regulatory networks during T follicular helper cell differentiation. Cell Rep. 2016, 14, 1735–1747. [Google Scholar] [CrossRef] [Green Version]
- Mesnage, R.; Arno, M.; Costanzo, M.; Malatesta, M.; Séralini, G.E.; Antoniou, M.N. Transcriptome profile analysis reflects rat liver and kidney damage following chronic ultra-low dose Roundup exposure. Environ. Health 2015, 14, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods 2001, 25, 402–4208. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Nucleotide Sequence 5′-3′ | PCR Product [bp] |
|---|---|---|---|
| DNMT1 | forward reverse | GTGGAAGCCGGCAAAGC TCCCACTCGAGCCTTCCATA | 125 |
| DNMT3A | forward reverse | AAGGAGGAGCGCCAAGAG ATCACCGCAGGGTCCTTT | 113 |
| EHMT1 | forward reverse | CCTCGACTCGGAAAAACCCAA AGAGCGCTTATTCTGGTGCT | 148 |
| EHMT2 | forward reverse | CATCGATCGCAACATCACCC GAGCAATCGCCCATCCTTGT | 121 |
| HDAC3 | forward reverse | TGACTCTCTGGGCTGTGATCG CATATTCAACGCATTCCCCATGC | 74 |
| HDAC5 | forward reverse | CTTAGCAAGTGCGAGCGGAT TGTCTAGCTTCTGCCGGTTG | 121 |
| TET3 | forward reverse | CACTAGCTGTACCAACCGCC CAGCCTTTATTTCCACCTCCTTGA | 104 |
| House keeping genes | |||
| GAPDH | forward reverse | AGCCACATCGCTCAGACAC GCCCAATACGACCAAATCC | 66 |
| RPL0 | forward reverse | TCTACAACCCTGAAGTGCTTGAT CAATCTGCAGACAGACACTGG | 96 |
| RPL13 | forward reverse | CAAGCGGATGAACACCAAC TGTGGGGCAGCATACCTC | 95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woźniak, E.; Reszka, E.; Jabłońska, E.; Michałowicz, J.; Huras, B.; Bukowska, B. Glyphosate and AMPA Induce Alterations in Expression of Genes Involved in Chromatin Architecture in Human Peripheral Blood Mononuclear Cells (In Vitro). Int. J. Mol. Sci. 2021, 22, 2966. https://doi.org/10.3390/ijms22062966
Woźniak E, Reszka E, Jabłońska E, Michałowicz J, Huras B, Bukowska B. Glyphosate and AMPA Induce Alterations in Expression of Genes Involved in Chromatin Architecture in Human Peripheral Blood Mononuclear Cells (In Vitro). International Journal of Molecular Sciences. 2021; 22(6):2966. https://doi.org/10.3390/ijms22062966
Chicago/Turabian StyleWoźniak, Ewelina, Edyta Reszka, Ewa Jabłońska, Jaromir Michałowicz, Bogumiła Huras, and Bożena Bukowska. 2021. "Glyphosate and AMPA Induce Alterations in Expression of Genes Involved in Chromatin Architecture in Human Peripheral Blood Mononuclear Cells (In Vitro)" International Journal of Molecular Sciences 22, no. 6: 2966. https://doi.org/10.3390/ijms22062966