
Hypoxic Conditions Promote the Angiogenic Potential of Human Induced Pluripotent Stem Cell-Derived Extracellular Vesicles

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
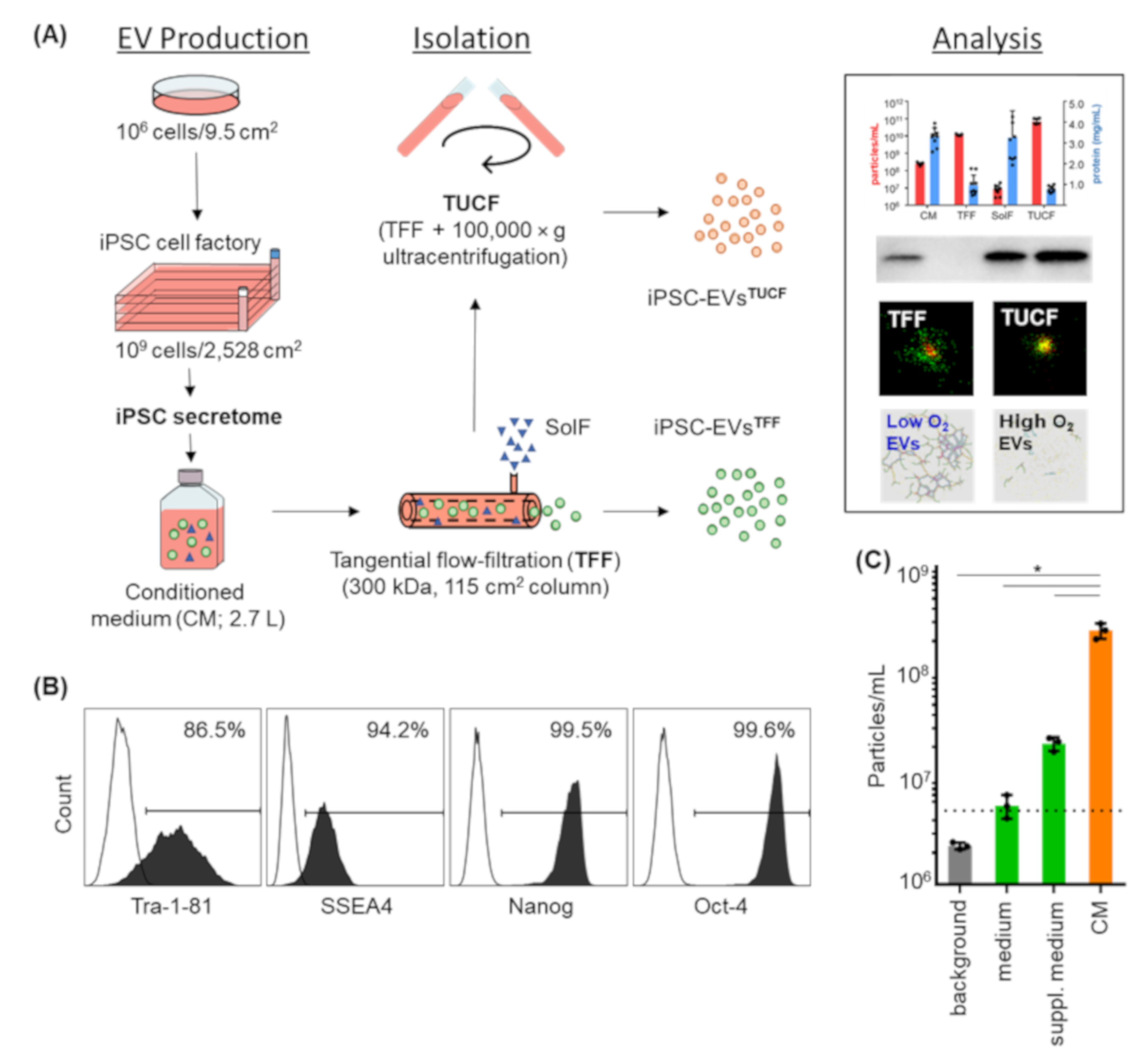
2.1. Pluripotency Markers and Viability of iPSCs Were Maintained after Large-Scale Expansion for EV Production
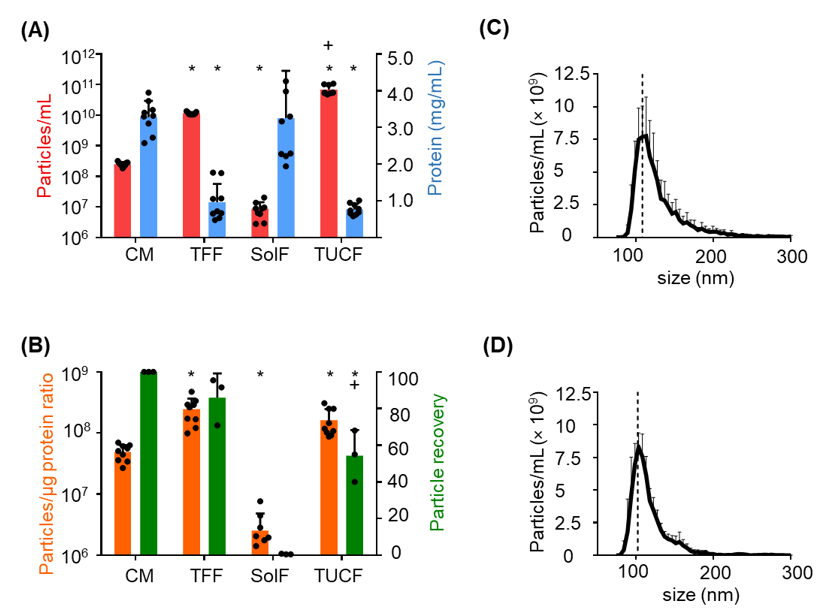
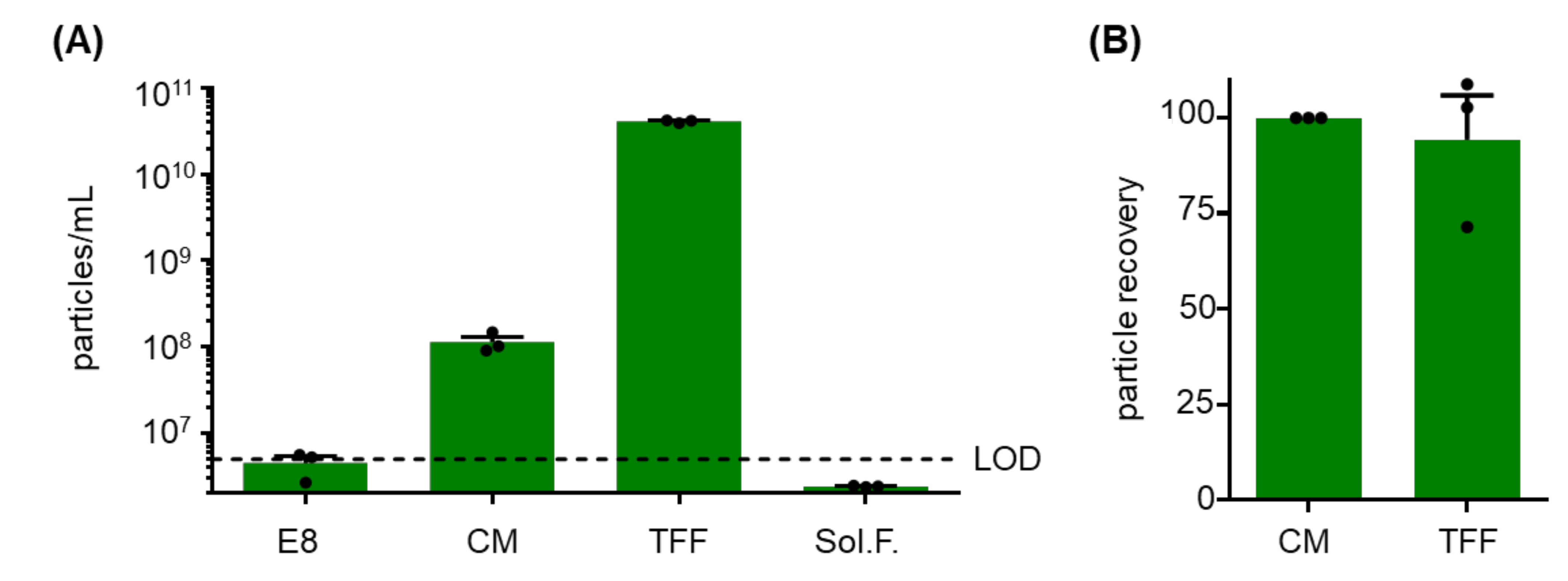
2.2. Monitoring Particle Recovery during Subsequent Purification
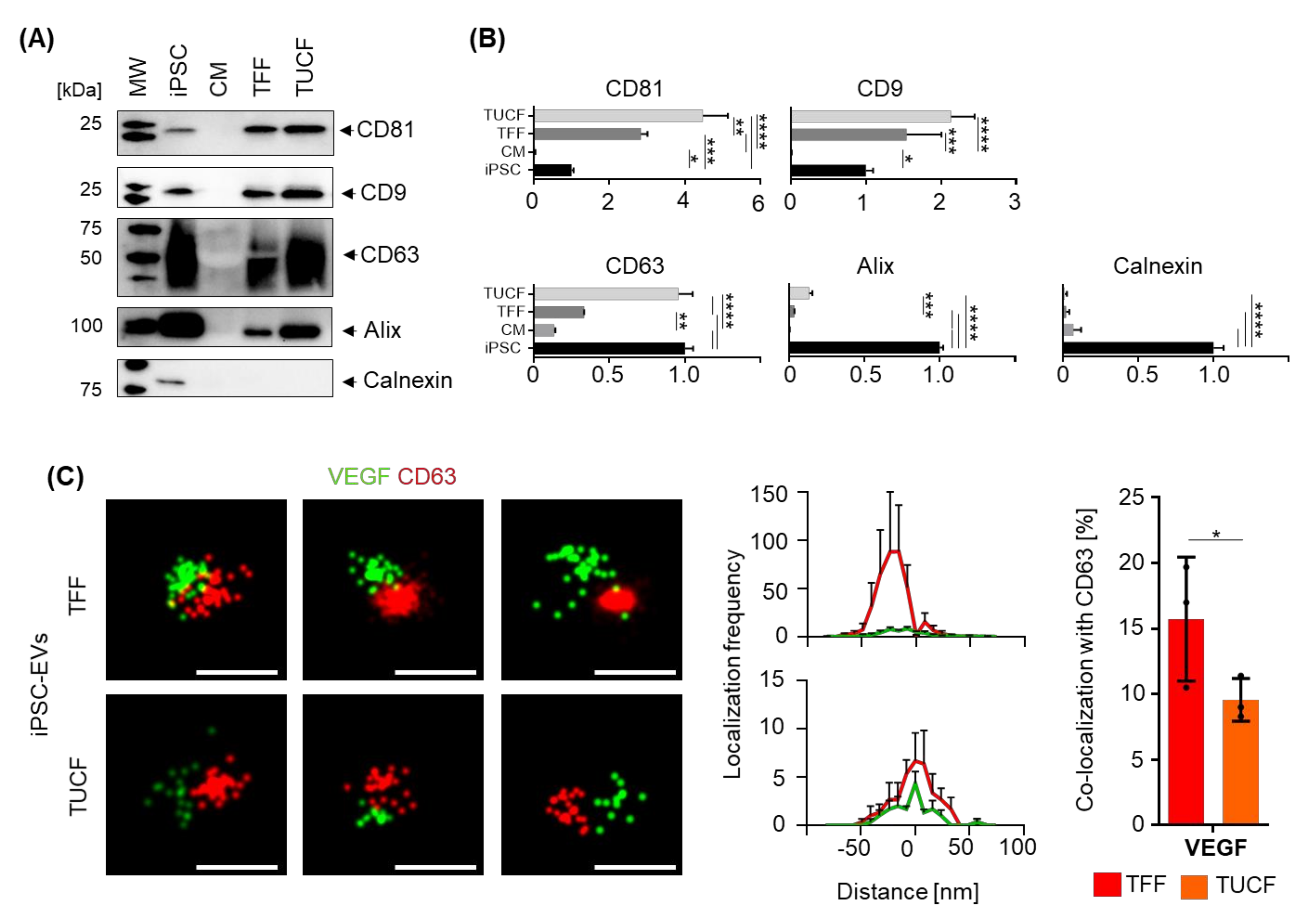
2.3. Marker Identity of iPSC-EVs
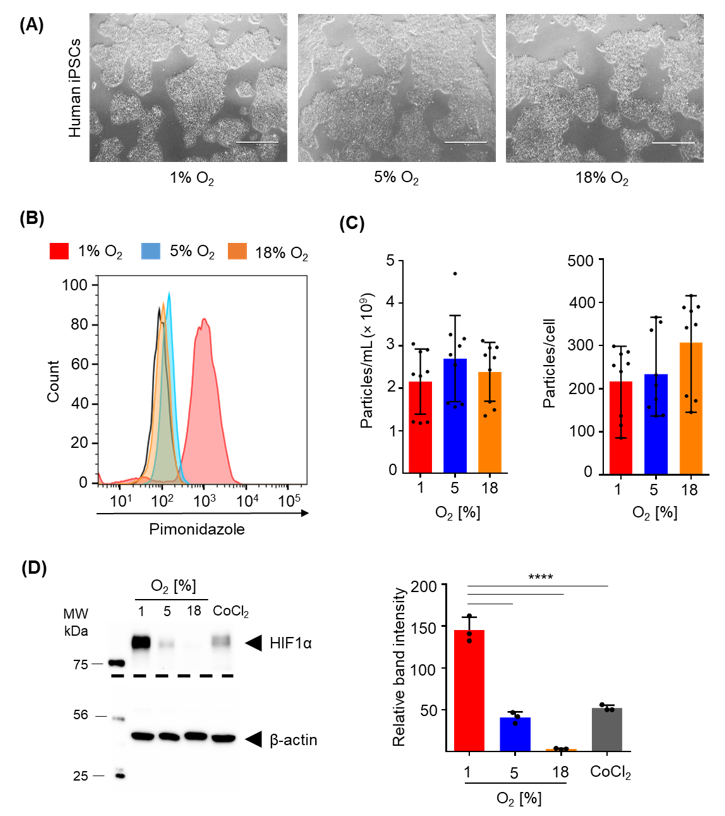
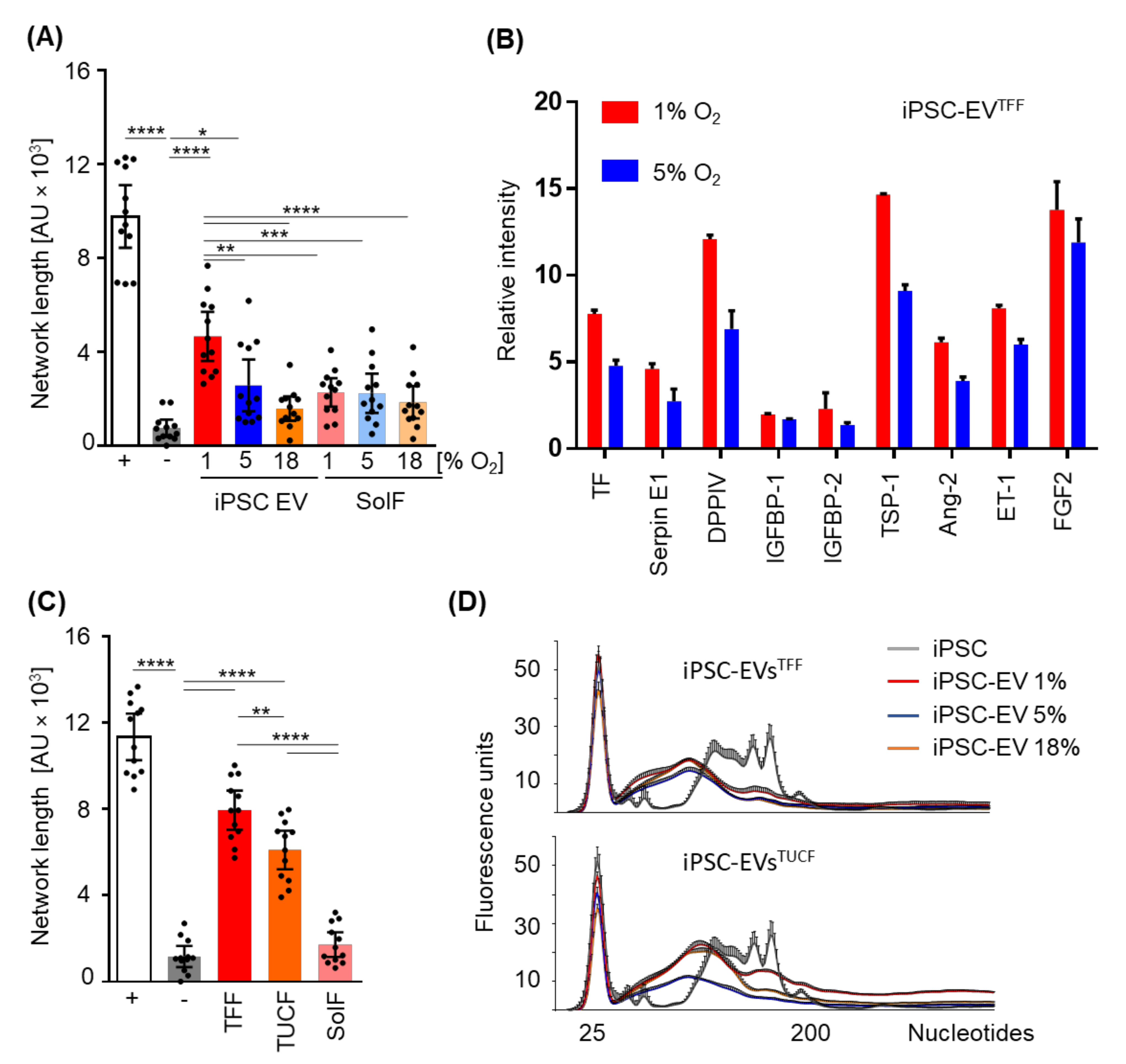
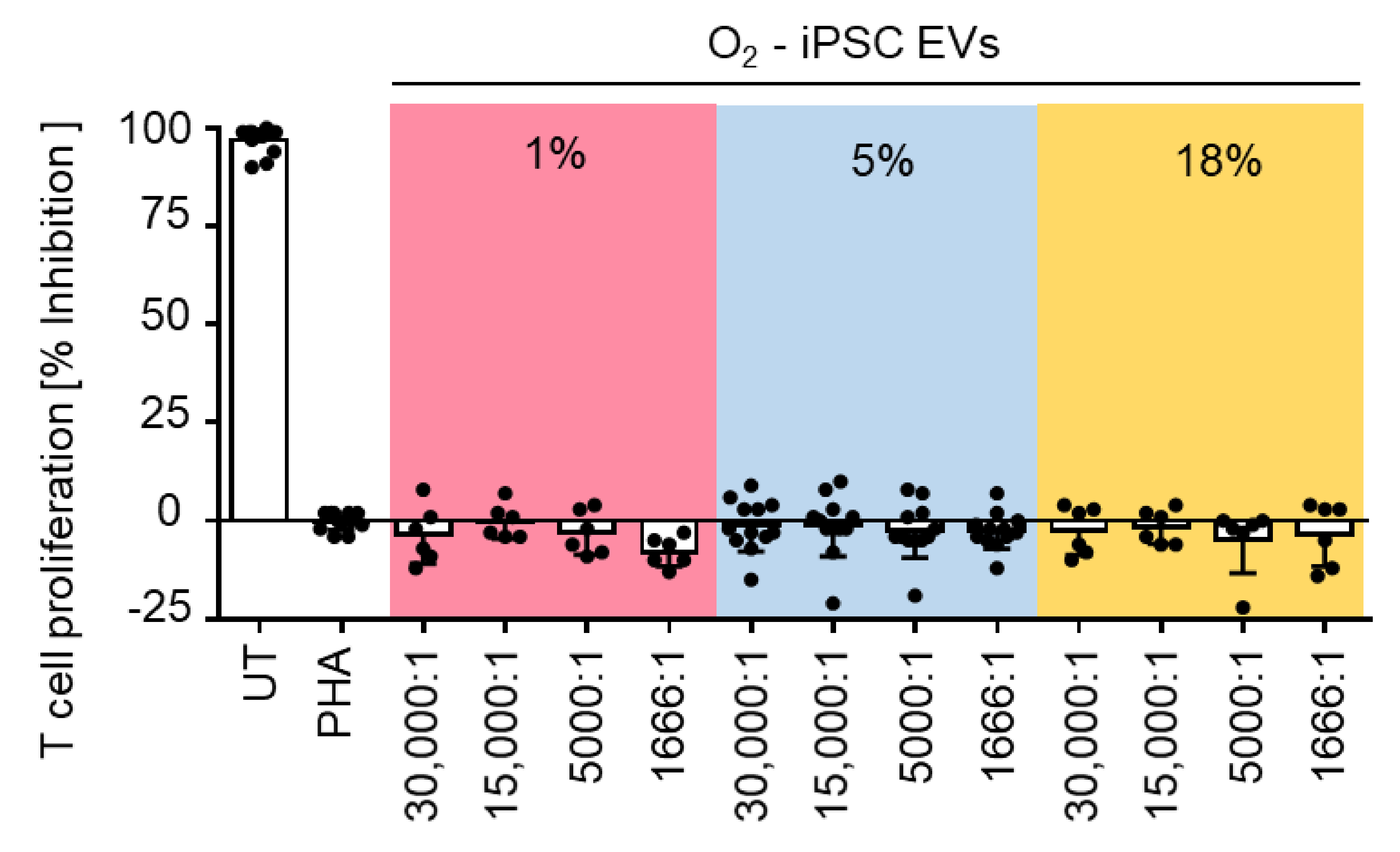
2.4. Efficacy of Hypoxia
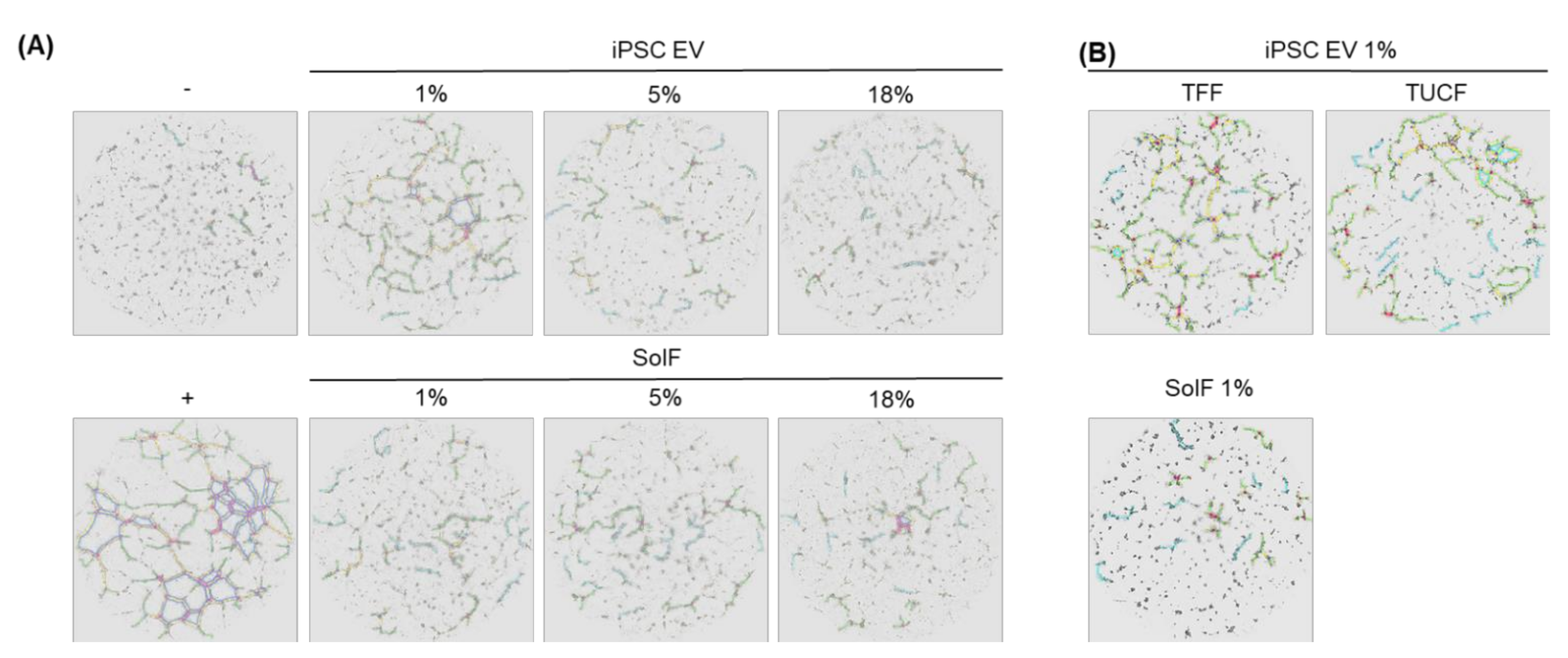
2.5. Angiogenic Potential of Human iPSC-EVs
3. Discussion
4. Materials and Methods
4.1. Human iPSC Culture under Different Oxygen Levels
4.2. 3D Human iPSC Culture
4.3. EV Purification
4.4. Determination of EV Concentration and Size via TRPS
4.5. Analysis of EV Markers and Proteins by Western Blot Analysis
4.6. Flow Cytometry
4.7. Immunomodulation Assay (IMA)
4.8. Super-Resolution Microscopy
4.9. In Vitro Angiogenesis Assay
4.10. Analysis of EV’s RNA Cargo
4.11. Angiogenesis Profiler Arrays
4.12. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EV Source | Intervention | EV Isolation | Outcome | Reference |
|---|---|---|---|---|
| hiPSC | In vitro high glucose challenge | UCF | iPSC-exosomes protected endothelial cells from high glucose-induced senescence | [24] |
| hiPSC | Growth factor depriva-tion during angiogenesis | TFF | EVs from both cell types enhanced growth factor-induced in vitro angiogenesis | [53] |
| Murine iPSC | Mouse myocardial infarction | UCF | Promoted in vitro angiogenesis and improved cardiac recovery post-infarction in vivo | [23] |
| Rhesus macaque iPSC | Macaque skin wound healing | Exo-quick + UCF | In vivo wound healing with increased number of capillaries | [54] |
| hiPSC-MSCs (stromal cells) | Cranial lesions in osteopenic mice | UF + DGC + UF | Promoted in vivo vessel formation in calvaria lesions | [49] |
| hiPSC-MSCs (stromal cells) | Rat stroke model | UCF | Promoted in vitro and in vivo angiogenesis | [55] |
| hiPSC-SMPCs | Cardiotoxin injury | UF + SEC + UF | EVs promoted in vitro angiogenesis | [56] |
| hiPSC-SMCs | Mouse hindlimb ischemia | UCF | iPSC-SMC exosomes promoted endothelial migration and proliferation | [57] |
| hiPSC-CMs | Angio- & scratch assay | Exo-quick | EVs promoted proliferation & angiogenesis | [41] |
| Rat iVPC (vascular progenitor cells) | Mouse hindlimb ischemia | UF + SEC | EVs romoted angiogenesis in vitro and in vivo | [58] |
| hiPSC-CM; -EC, -SMCs | Swine myocardial infarction | Exo-quick | EVs promoted reperfusion after infarction | [59] |
| hiPSC-CMPs | Mouse myocardial infarction | UCF | EVs induced CM proliferation & outperformed cells for cardiac recovery post-infarction | [42] |
| hiPSC-ECs | Mouse limb ischemia | UCF | Promoted angiogenesis/blood perfusion | [37] |
| hiPSC-MSCs (stromal cells) | Mouse skin injury | UCF + UF | Promoted wound healing, fibroblast-collagen synthesis and angiogenesis | [60] |
| hiPSC-MSCs (stromal cells) | Steroid-induced rat osteonecrosis | UCF+ UF | Prevented bone loss and promoted angiogenesis | [40] |
| hiPSC-CMs | hIPSC-CM from HyperGEN cohort | UF + PEG + SEC | Delayed angiogenesis with hIPSC-CM-EVs from left ventricular hypertrophy patients | [61] |
| hiPSC-MSCs (stromal cells) | Mouse limb ischemia | UF + DGC + UF | Promoted angiogenesis/blood perfusion | [39] |
| Nomenclature | ||||
|---|---|---|---|---|
| Use of Generic Term EV for iPSC-EVs | ||||
| Collection and Storage | ||||
| Releasing cell information | cell type and origin | Human iPSC reprogramed from bone marrow | ||
| passaging | p28/p35 | |||
| seeding density | cells/cm2 | |||
| culture volume | 500 mL | |||
| culture vessel | 2528 cm2 (CF4) | |||
| oxygen level | 1, 5 and 18% | |||
| Culture conditions | culturing medium | mTeSR basal medium plus supplement (serum-free) | ||
| time of cultivation | 4–8 days | |||
| harvesting medium | mTeSR basal medium plus supplement (serum-free) | |||
| time of cultivation | 24 h | |||
| cell count at harvest | Mean 9.767 × 108 cells | |||
| Storage and recovery | conditioned medium | Storage temperature: −80 °C | ||
| EV preparations | Storage temperature: −80 °C/snap freeze | |||
| Isolation | ||||
| Differential centrifugation | 300× g for 5 min, SX4750, Allegra X-15R, Beckman Coulter 3000× g for 15 min, SX4750, Allegra X-15R, Beckman Coulter | |||
| Ultrafiltration (TFF) | Tangential flow filtration pore size: 300 kDa; concentration factor 150, isovolumetric washing with twice start volume | |||
| Ultracentrifugation (TUCF *) | 100,000× g for 90 min, TH-641 swing rotor Thermo Scientific | |||
| Characterization | ||||
| Parameter | Unit | Method | ||
| Quantification | Size & concentration | particle number | 1.15 × 1010 particles/mL (TFF) 6.95 × 1010 particles/mL (TUCF) | TRPS |
| particle size | 109 nm (TFF) 103 nm (TUCF) | TRPS | ||
| particle/protein ratio | 2.46 × 108 particles/µg (TFF) 1.62 × 108 particles/µg (TFF) | TRPS/Bradford | ||
| Composition | protein content | 578 mg/mL (TFF) 460 mg/mL (TUCF) | Bradford | |
| RNA content | 3 pg/µL (TFF) 1 pg/µL (TUCF) | Bioanalyzer RNA 6000 pico | ||
| Identity | trans membrane proteins | CD9 | WB | |
| trans membrane proteins | CD63 | WB | ||
| trans membrane proteins | CD81 | WB | ||
| cytosolic recovered in EVs | Alix-1 | WB | ||
| secretory pathway | Calnexin | WB | ||
| Visualization | TFF: super resolution microscopy (CD63/VEGF) | |||
| TUCF: super resolution microscopy (CD63/VEGF) | ||||
| Functional Studies | ||||
| Angiogenesis | effect of EVs (10,000:1) on endothelial network formation | |||
References
- Critser, P.J.; Yoder, M.C. Endothelial colony-forming cell role in neoangiogenesis and tissue repair. Curr. Opin. Organ Transplant. 2010, 15, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Koike, N.; Fukumura, D.; Gralla, O.; Au, P.; Schechner, J.S.; Jain, R.K. Creation of long-lasting blood vessels. Nature 2004, 428, 138–139. [Google Scholar] [CrossRef] [PubMed]
- Melero-Martin, J.M.; Khan, Z.A.; Picard, A.; Wu, X.; Paruchuri, S.; Bischoff, J. In vivo vasculogenic potential of human blood-derived endothelial progenitor cells. Blood 2007, 109, 4761–4768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinisch, A.; Hofmann, N.A.N.A.; Obenauf, A.C.A.C.; Kashofer, K.; Rohde, E.; Schallmoser, K.; Flicker, K.; Lanzer, G.; Linkesch, W.; Speicher, M.R.M.R.; et al. Humanized large-scale expanded endothelial colony-forming cells function in vitro and in vivo. Blood 2009, 113, 6716–6725. [Google Scholar] [CrossRef] [Green Version]
- Prasain, N.; Lee, M.R.; Vemula, S.; Meador, J.L.; Yoshimoto, M.; Ferkowicz, M.J.; Fett, A.; Gupta, M.; Rapp, B.M.; Saadatzadeh, M.R.; et al. Differentiation of human pluripotent stem cells to cells similar to cord-blood endothelial colony-forming cells. Nat. Biotechnol. 2014, 32, 1151–1157. [Google Scholar] [CrossRef]
- Olmer, R.; Engels, L.; Usman, A.; Menke, S.; Malik, M.N.H.; Pessler, F.; Göhring, G.; Bornhorst, D.; Bolten, S.; Abdelilah-Seyfried, S.; et al. Differentiation of human pluripotent stem cells into functional endothelial cells in scalable suspension culture. Stem Cell Rep. 2018, 10, 1657–1672. [Google Scholar] [CrossRef] [Green Version]
- Peking, P.; Krisch, L.; Wolf, M.; Hoog, A.; Vári, B.; Muigg, K.; Poupardin, R.; Scharler, C.; Russe, E.; Stachelscheid, H.; et al. Self-assembly of progenitor cells under the aegis of platelet factors facilitates human skin organoid formation and vascularized wound healing. bioRxiv 2020. [Google Scholar] [CrossRef]
- Drakhlis, L.; Biswanath, S.; Farr, C.-M.; Lupanow, V.; Teske, J.; Ritzenhoff, K.; Franke, A.; Manstein, F.; Bolesani, E.; Kempf, H.; et al. Human heart-forming organoids recapitulate early heart and foregut development. Nat. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Semenza, G.L. Pharmacologic targeting of hypoxia-inducible factors. Annu. Rev. Pharmacol. Toxicol. 2019, 59, 379–403. [Google Scholar] [CrossRef]
- Choueiri, T.K.; Kaelin, W.G. Targeting the HIF2-VEGF axis in renal cell carcinoma. Nat. Med. 2020, 26, 1519–1530. [Google Scholar] [CrossRef]
- Hofmann, N.A.; Ortner, A.; Jacamo, R.O.; Reinisch, A.; Schallmoser, K.; Rohban, R.; Etchart, N.; Fruehwirth, M.; Beham-Schmid, C.; Andreeff, M.; et al. Oxygen sensing mesenchymal progenitors promote neo-vasculogenesis in a humanized mouse model in vivo. PLoS ONE 2012, 7, e44468. [Google Scholar] [CrossRef]
- Ferland-McCollough, D.; Slater, S.; Richard, J.; Reni, C.; Mangialardi, G. Pericytes, an overlooked player in vascular pathobiology. Pharmacol. Ther. 2017, 171, 30–42. [Google Scholar] [CrossRef]
- Rivera, F.J.; Silva, M.E.; Aigner, L. Editorial: The vascular niche in tissue repair: A therapeutic target for regeneration. Front. Cell Dev. Biol. 2017, 5, 88. [Google Scholar] [CrossRef] [Green Version]
- Rohban, R.; Reinisch, A.; Etchart, N.; Schallmoser, K.; Hofmann, N.A.; Szoke, K.; Brinchmann, J.E.; Rad, E.B.; Rohde, E.; Strunk, D. Identification of an effective early signaling signature during neo-vasculogenesis in vivo by ex vivo proteomic profiling. PLoS ONE 2013, 8, e66909. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367. [Google Scholar] [CrossRef]
- Lee, Y.; El Andaloussi, S.; Wood, M.J.A. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef] [Green Version]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Gudbergsson, J.M.; Jønsson, K.; Simonsen, J.B.; Johnsen, K.B. Systematic review of targeted extracellular vesicles for drug delivery-Considerations on methodological and biological heterogeneity. J. Control. Release 2019, 306, 108–120. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Lener, T.; Gimona, M.; Aigner, L.; Börger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; del Portillo, H.A.; et al. Applying extracellular vesicles based therapeutics in clinical trials-An ISEV position paper. J. Extracell. Vesicles 2015, 4. [Google Scholar] [CrossRef]
- Mancuso, T.; Barone, A.; Salatino, A.; Molinaro, C.; Marino, F.; Scalise, M.; Torella, M.; De Angelis, A.; Urbanek, K.; Torella, D.; et al. Unravelling the biology of adult cardiac stem cell-derived exosomes to foster endogenous cardiac regeneration and repair. Int. J. Mol. Sci. 2020, 21, 3725. [Google Scholar] [CrossRef] [PubMed]
- Menasché, P. Cardiac cell therapy: Current status, challenges and perspectives. Arch. Cardiovasc. Dis. 2020, 113, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Adamiak, M.; Cheng, G.; Bobis-Wozowicz, S.; Zhao, L.; Kedracka-Krok, S.; Samanta, A.; Karnas, E.; Xuan, Y.T.; Skupien-Rabian, B.; Chen, X.; et al. Induced Pluripotent Stem Cell (iPSC)-derived extracellular vesicles are safer and more effective for cardiac repair than iPSCs. Circ. Res. 2018, 122, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Sun, R.; Wang, P.; Zhang, H.; Xiang, M.; Meng, D.; Sun, N.; Chen, A.; Chen, S. Protective effects of human induced pluripotent stem cell-derived exosomes on high glucose-induced injury in human endothelial cells. Exp. Ther. Med. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamò, L.; Fytianos, K.; Caldana, F.; Simillion, C.; Feki, A.; Nita, I.; Heller, M.; Geiser, T.; Gazdhar, A. Interactome analysis of ipsc secretome and its effect on macrophages in vitro. Int. J. Mol. Sci. 2021, 22, 958. [Google Scholar] [CrossRef]
- Place, T.L.; Domann, F.E.; Case, A.J. Limitations of oxygen delivery to cells in culture: An underappreciated problem in basic and translational research. Free Radic. Biol. Med. 2017, 113, 311–322. [Google Scholar] [CrossRef]
- Schito, L.; Rey, S.; Konopleva, M. Integration of hypoxic HIF-α signaling in blood cancers. Oncogene 2017, 36, 5331–5340. [Google Scholar] [CrossRef]
- Schallmoser, K.; Rohde, E.; Reinisch, A.; Bartmann, C.; Thaler, D.; Drexler, C.; Obenauf, A.C.; Lanzer, G.; Linkesch, W.; Strunk, D. Rapid large-scale expansion of functional mesenchymal stem cells from unmanipulated bone marrow without animal serum. Tissue Eng. Part C Methods 2008, 14, 185–196. [Google Scholar] [CrossRef]
- Manstein, F.; Ullmann, K.; Kropp, C.; Halloin, C.; Triebert, W.; Franke, A.; Farr, C.; Sahabian, A.; Haase, A.; Breitkreuz, Y.; et al. High density bioprocessing of human pluripotent stem cells by metabolic control and in silico modeling. Stem Cells Transl. Med. 2021. [Google Scholar] [CrossRef]
- Kropp, C.; Kempf, H.; Halloin, C.; Robles-Diaz, D.; Franke, A.; Scheper, T.; Kinast, K.; Knorpp, T.; Joos, T.O.; Haverich, A.; et al. Impact of feeding strategies on the scalable expansion of human pluripotent stem cells in single-use stirred tank bioreactors. Stem Cells Transl. Med. 2016, 5, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Kordelas, L.; Rebmann, V.; Ludwig, A.-K.; Radtke, S.; Ruesing, J.; Doeppner, T.R.; Epple, M.; Horn, P.A.; Beelen, D.W.; Giebel, B. MSC-derived exosomes: A novel tool to treat therapy-refractory graft-versus-host disease. Leukemia 2014, 28, 970–973. [Google Scholar] [CrossRef]
- Wolf, M.; Vari, B.; Blöchl, C.; Raninger, A.M.; Poupardin, R.; Beez, C.M.; Hoog, A.; Brachtl, G.; Eminger, E.; Binder, H.M.; et al. Extracellular vesicles from therapeutic grade allogeneic human placental stromal cells induce angiogenesis and modulate immunity. bioRxiv 2019, 808808. [Google Scholar] [CrossRef] [Green Version]
- Warnecke, A.; Harre, J.; Staecker, H.; Prenzler, N.; Strunk, D.; Couillard-Despres, S.; Romanelli, P.; Hollerweger, J.; Lassacher, T.; Auer, D.; et al. Extracellular vesicles from human multipotent stromal cells protect against hearing loss after noise trauma in vivo. Clin. Transl. Med. 2020, 10. [Google Scholar] [CrossRef]
- Mas-Bargues, C.; Sanz-Ros, J.; Román-Domínguez, A.; Inglés, M.; Gimeno-Mallench, L.; El Alami, M.; Viña-Almunia, J.; Gambini, J.; Viña, J.; Borrás, C. Relevance of oxygen concentration in stem cell culture for regenerative medicine. Int. J. Mol. Sci. 2019, 20, 1195. [Google Scholar] [CrossRef] [Green Version]
- Mohyeldin, A.; Garzón-Muvdi, T.; Quiñones-Hinojosa, A. Oxygen in stem cell biology: A critical component of the stem cell niche. Cell Stem Cell 2010, 7, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Tursky, M.L.; Loi, T.H.; Artuz, C.M.; Alateeq, S.; Wolvetang, E.J.; Tao, H.; Ma, D.D.; Molloy, T.J. Direct comparison of four hematopoietic differentiation methods from human induced pluripotent stem cells. Stem Cell Rep. 2020, 15, 735–748. [Google Scholar] [CrossRef]
- Ye, M.; Ni, Q.; Qi, H.; Qian, X.; Chen, J.; Guo, X.; Li, M.; Zhao, Y.; Xue, G.; Deng, H.; et al. Exosomes derived from human induced pluripotent stem cells-endothelia cells promotes postnatal angiogenesis in mice bearing ischemic limbs. Int. J. Biol. Sci. 2019, 15, 158–168. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, L.; Li, Y.; Chen, L.; Wang, X.; Guo, W.; Zhang, X.; Qin, G.; He, S.; Zimmerman, A.; et al. Exosomes/microvesicles from induced pluripotent stem cells deliver cardioprotective miRNAs and prevent cardiomyocyte apoptosis in the ischemic myocardium. Int. J. Cardiol. 2015, 192, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.W.; Li, Q.; Niu, X.; Hu, B.; Liu, J.; Zhou, S.M.; Guo, S.C.; Lang, H.L.; Zhang, C.Q.; Wang, Y.; et al. Exosomes secreted by human-induced pluripotent stem cell-derived mesenchymal stem cells attenuate limb ischemia by promoting angiogenesis in mice. Stem Cell Res. Ther. 2015, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, Q.; Niu, X.; Hu, B.; Chen, S.; Song, W.; Ding, J.; Zhang, C.; Wang, Y. Exosomes secreted from human-induced pluripotent stem cell-derived mesenchymal stem cells prevent osteonecrosis of the femoral head by promoting angiogenesis. Int. J. Biol. Sci. 2017, 13, 232–244. [Google Scholar] [CrossRef]
- Dougherty, J.A.; Kumar, N.; Noor, M.; Angelos, M.G.; Khan, M.M.; Chen, C.-A.; Khan, M.M. Extracellular vesicles released by human induced-pluripotent stem cell-derived cardiomyocytes promote angiogenesis. Front. Physiol. 2018, 9, 1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Harane, N.; Kervadec, A.; Bellamy, V.; Pidial, L.; Neametalla, H.J.; Perier, M.-C.C.; Lima Correa, B.; Thiébault, L.; Cagnard, N.; Duché, A.; et al. Acellular therapeutic approach for heart failure: In vitro production of extracellular vesicles from human cardiovascular progenitors. Eur. Heart J. 2018, 39, 1835–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltrán-Camacho, L.; Rojas-Torres, M.; Durán-Ruiz, M.C. Current status of angiogenic cell therapy and related strategies applied in critical limb ischemia. Int. J. Mol. Sci. 2021, 22, 2335. [Google Scholar] [CrossRef] [PubMed]
- Reinisch, A.; Strunk, D. Isolation and animal serum free expansion of human umbilical cord derived mesenchymal stromal cells (MSCs) and endothelial colony forming progenitor cells (ECFCs). J. Vis. Exp. 2009. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, N.A.; Reinisch, A.; Strunk, D. Isolation and large scale expansion of adult human endothelial colony forming progenitor cells. J. Vis. Exp. 2009. [Google Scholar] [CrossRef] [Green Version]
- Scharler, C.; Poupardin, R.; Peking, P.; Wolf, M.; Brachtl, G.; Daheron, L.; Schallmoser, K.; Jürchott, K.; Stachelscheid, H.; Volk, H.-D.; et al. Extra-hematopoietic immunomodulatory role of the SCID-susceptibility gene DOCK-2 identified by stepwise maturation of human iPSCs into clonogenic mesodermal stromal progenitors. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ketterl, N.; Brachtl, G.; Schuh, C.; Bieback, K.; Schallmoser, K.; Reinisch, A.; Strunk, D. A robust potency assay highlights significant donor variation of human mesenchymal stem/progenitor cell immune modulatory capacity and extended radio-resistance. Stem Cell Res. Ther. 2015, 6, 236. [Google Scholar] [CrossRef] [Green Version]
- Pachler, K.; Ketterl, N.; Desgeorges, A.; Dunai, Z.A.Z.A.; Laner-Plamberger, S.; Streif, D.; Strunk, D.; Rohde, E.; Gimona, M. An in vitro potency assay for monitoring the immunomodulatory potential of stromal cell-derived extracellular vesicles. Int. J. Mol. Sci. 2017, 18, 1413. [Google Scholar] [CrossRef]
- Qi, X.; Zhang, J.; Yuan, H.; Xu, Z.; Li, Q.; Niu, X.; Hu, B.; Wang, Y.; Li, X. Exosomes secreted by human-induced pluripotent stem cell-derived mesenchymal stem cells repair critical-sized bone defects through enhanced angiogenesis and osteogenesis in osteoporotic rats. Int. J. Biol. Sci. 2016, 12, 836–849. [Google Scholar] [CrossRef] [Green Version]
- Burnouf, T.; Strunk, D.; Koh, M.B.C.; Schallmoser, K. Human platelet lysate: Replacing fetal bovine serum as a gold standard for human cell propagation? Biomaterials 2016, 76. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Carpentier, G. Angiogenesis Analyser for ImageJ. ImageJ News. 2012. Available online: http://image.bio.methods.free.fr (accessed on 26 March 2021).
- Branscome, H.; Paul, S.; Khatkar, P.; Kim, Y.; Barclay, R.A.; Pinto, D.O.; Yin, D.; Zhou, W.; Liotta, L.A.; El-Hage, N.; et al. Stem cell extracellular vesicles and their potential to contribute to the repair of damaged CNS cells. J. Neuroimmune Pharmacol. 2020, 15, 520–537. [Google Scholar] [CrossRef]
- Lu, M.; Peng, L.; Ming, X.; Wang, X.X.X.; Cui, A.; Li, Y.; Wang, X.X.X.; Meng, D.; Sun, N.; Xiang, M.; et al. Enhanced wound healing promotion by immune response-free monkey autologous iPSCs and exosomes vs. their allogeneic counterparts. EBioMedicine 2019, 42, 443–457. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Ling, X.; Hu, G.; Zhu, Q.; Zhang, J.; Li, Q.; Zhao, B.; Wang, Y.; Deng, Z. Small extracellular vesicles secreted by human iPSC-derived MSC enhance angiogenesis through inhibiting STAT3-dependent autophagy in ischemic stroke. Stem Cell Res. Ther. 2020, 11, 313. [Google Scholar] [CrossRef]
- Xuan, W.; Khan, M.; Ashraf, M. Pluripotent stem cell-induced skeletal muscle progenitor cells with givinostat promote myoangiogenesis and restore dystrophin in injured Duchenne dystrophic muscle. Stem Cell Res. Ther. 2021, 12, 131. [Google Scholar] [CrossRef]
- Park, J.J.; Kwon, Y.W.; Kim, J.W.; Park, G.T.; Yoon, J.W.; Kim, Y.S.; Kim, D.S.; Kwon, S.M.; Bae, S.S.; Ko, K.; et al. Coadministration of endothelial and smooth muscle cells derived from human induced pluripotent stem cells as a therapy for critical limb ischemia. Stem Cells Transl. Med. 2021, 10, 414–426. [Google Scholar] [CrossRef]
- Johnson, T.K.; Zhao, L.; Zhu, D.; Wang, Y.; Xiao, Y.; Oguljahan, B.; Zhao, X.; Kirlin, W.G.; Yin, L.; Chilian, W.M.; et al. Exosomes derived from induced vascular progenitor cells promote angiogenesis in vitro and in an in vivo rat hindlimb ischemia model. Am. J. Physiol. Circ. Physiol. 2019, 317, H765–H776. [Google Scholar] [CrossRef]
- Gao, L.; Wang, L.; Wei, Y.; Krishnamurthy, P.; Walcott, G.P.; Menasché, P.; Zhang, J. Exosomes secreted by hiPSC-derived cardiac cells improve recovery from myocardial infarction in swine. Sci. Transl. Med. 2020, 12. [Google Scholar] [CrossRef]
- Zhang, J.; Guan, J.; Niu, X.; Hu, G.; Guo, S.; Li, Q.; Xie, Z.; Zhang, C.; Wang, Y. Exosomes released from human induced pluripotent stem cells-derived MSCs facilitate cutaneous wound healing by promoting collagen synthesis and angiogenesis. J. Transl. Med. 2015, 13, 49. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.; Aggarwal, P.; Matter, A.; Olson, B.; Gu, C.C.; Hunt, S.C.; Lewis, C.E.; Arnett, D.K.; Lorier, R.; Broeckel, U. Donor-specific phenotypic variation in hiPSC cardiomyocyte-derived exosomes impacts endothelial cell function. Am. J. Physiol. Circ. Physiol. 2021, 320, H954–H968. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade, A.C.; Wolf, M.; Binder, H.-M.; Gomes, F.G.; Manstein, F.; Ebner-Peking, P.; Poupardin, R.; Zweigerdt, R.; Schallmoser, K.; Strunk, D. Hypoxic Conditions Promote the Angiogenic Potential of Human Induced Pluripotent Stem Cell-Derived Extracellular Vesicles. Int. J. Mol. Sci. 2021, 22, 3890. https://doi.org/10.3390/ijms22083890
Andrade AC, Wolf M, Binder H-M, Gomes FG, Manstein F, Ebner-Peking P, Poupardin R, Zweigerdt R, Schallmoser K, Strunk D. Hypoxic Conditions Promote the Angiogenic Potential of Human Induced Pluripotent Stem Cell-Derived Extracellular Vesicles. International Journal of Molecular Sciences. 2021; 22(8):3890. https://doi.org/10.3390/ijms22083890
Chicago/Turabian StyleAndrade, André Cronemberger, Martin Wolf, Heide-Marie Binder, Fausto Gueths Gomes, Felix Manstein, Patricia Ebner-Peking, Rodolphe Poupardin, Robert Zweigerdt, Katharina Schallmoser, and Dirk Strunk. 2021. "Hypoxic Conditions Promote the Angiogenic Potential of Human Induced Pluripotent Stem Cell-Derived Extracellular Vesicles" International Journal of Molecular Sciences 22, no. 8: 3890. https://doi.org/10.3390/ijms22083890