
Membrane Profiling by Free Flow Electrophoresis and SWATH-MS to Characterize Subcellular Compartment Proteomes in Mesembryanthemum crystallinum


Abstract
:1. Introduction
2. Results
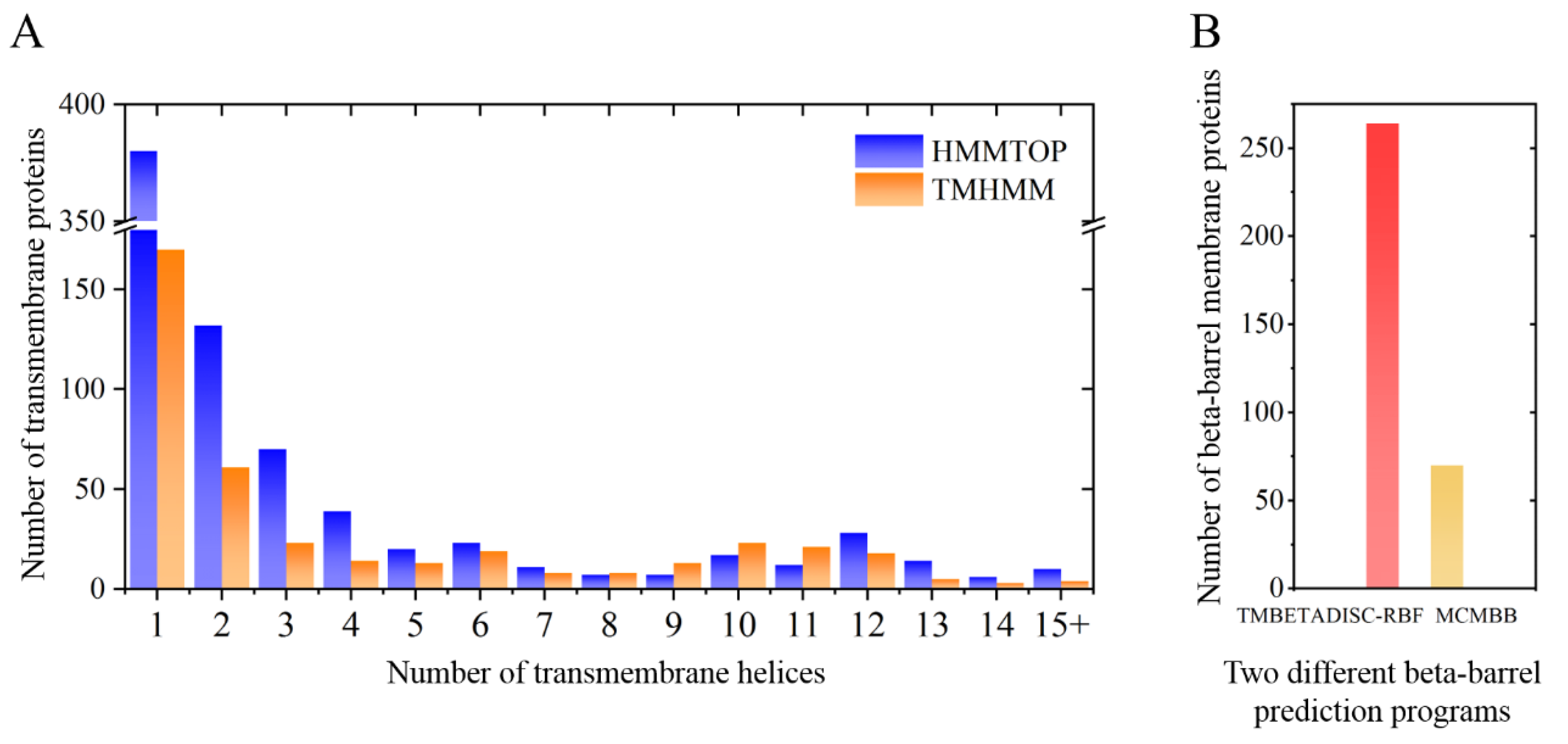
2.1. M. crystallinum Membrane Proteome
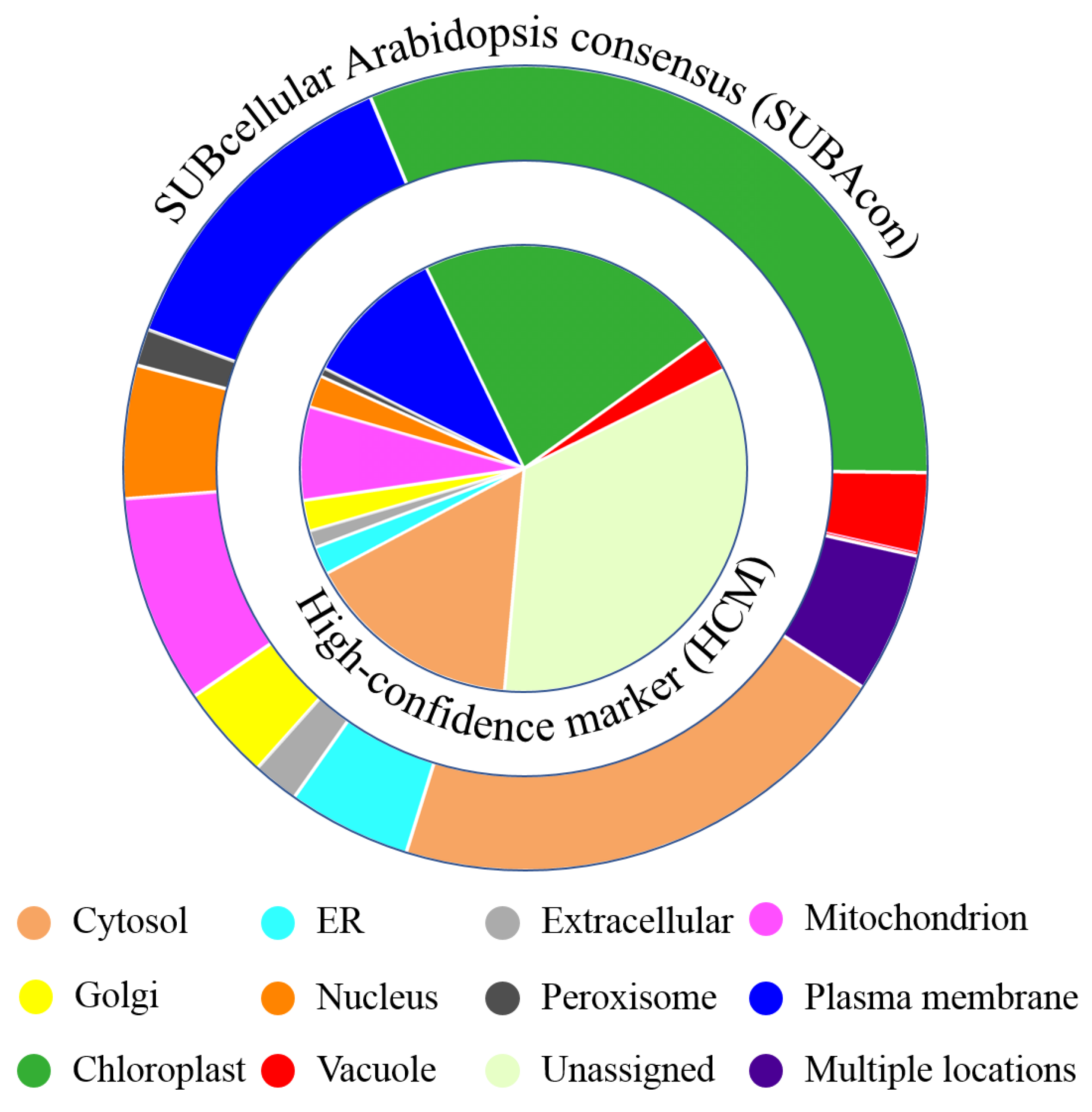
2.2. Functional Annotation and Subcellular Localization
2.3. FFE Profile of Representative Subcellular Membrane Markers
2.4. FFE Profiles of Lipid Metabolism-Related Proteins
3. Discussion
3.1. V-ATPase VHA Subunit Localization
3.2. Proteins with Unexpected FFE Profiles
3.3. A Snapshot of the Components of Electrochemical Reactions
3.4. Subcellular Mapping of Lipid Metabolism
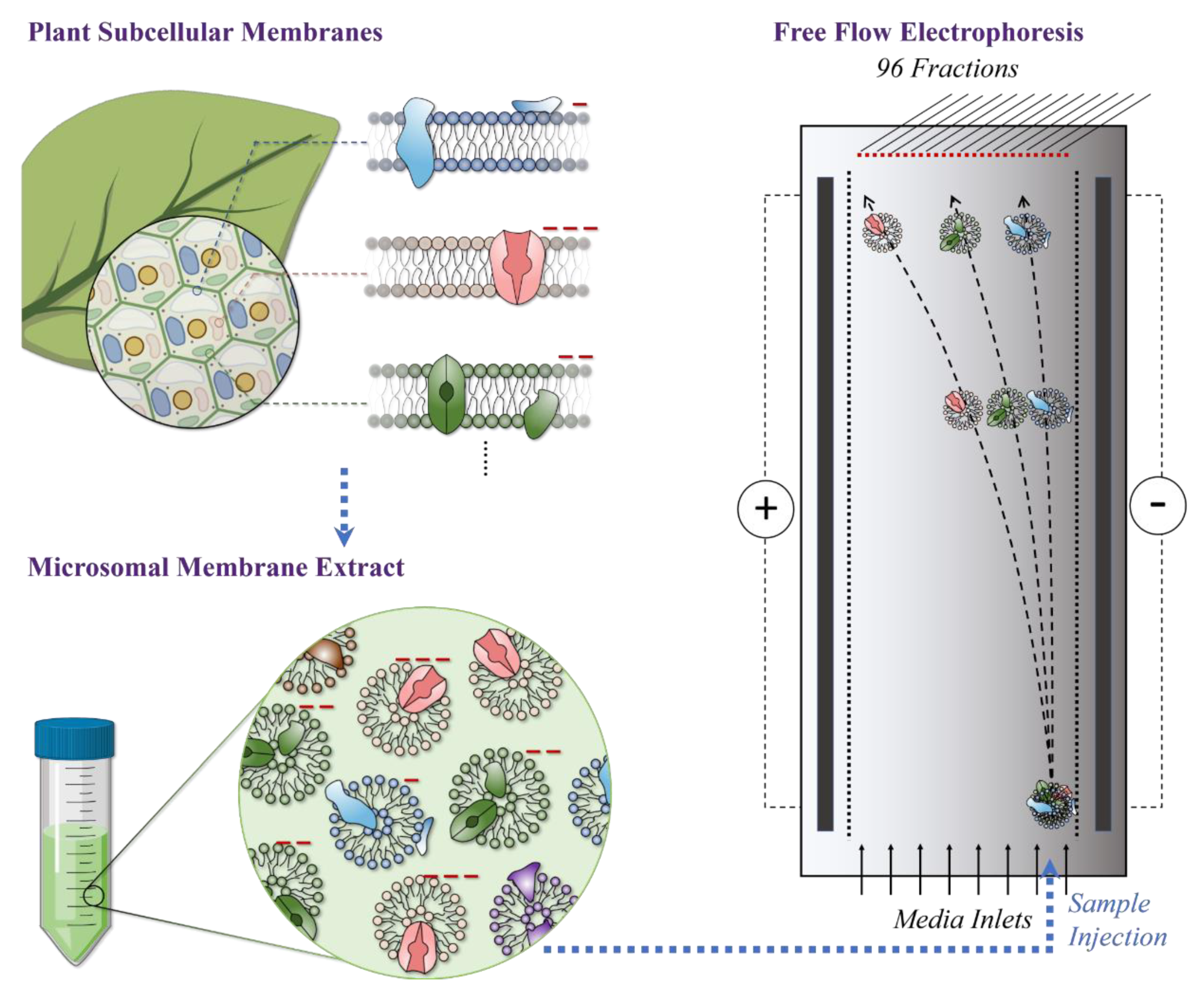
4. Materials and Methods
4.1. M. crystallinum Plant Material
4.2. Microsomal Membrane Extraction
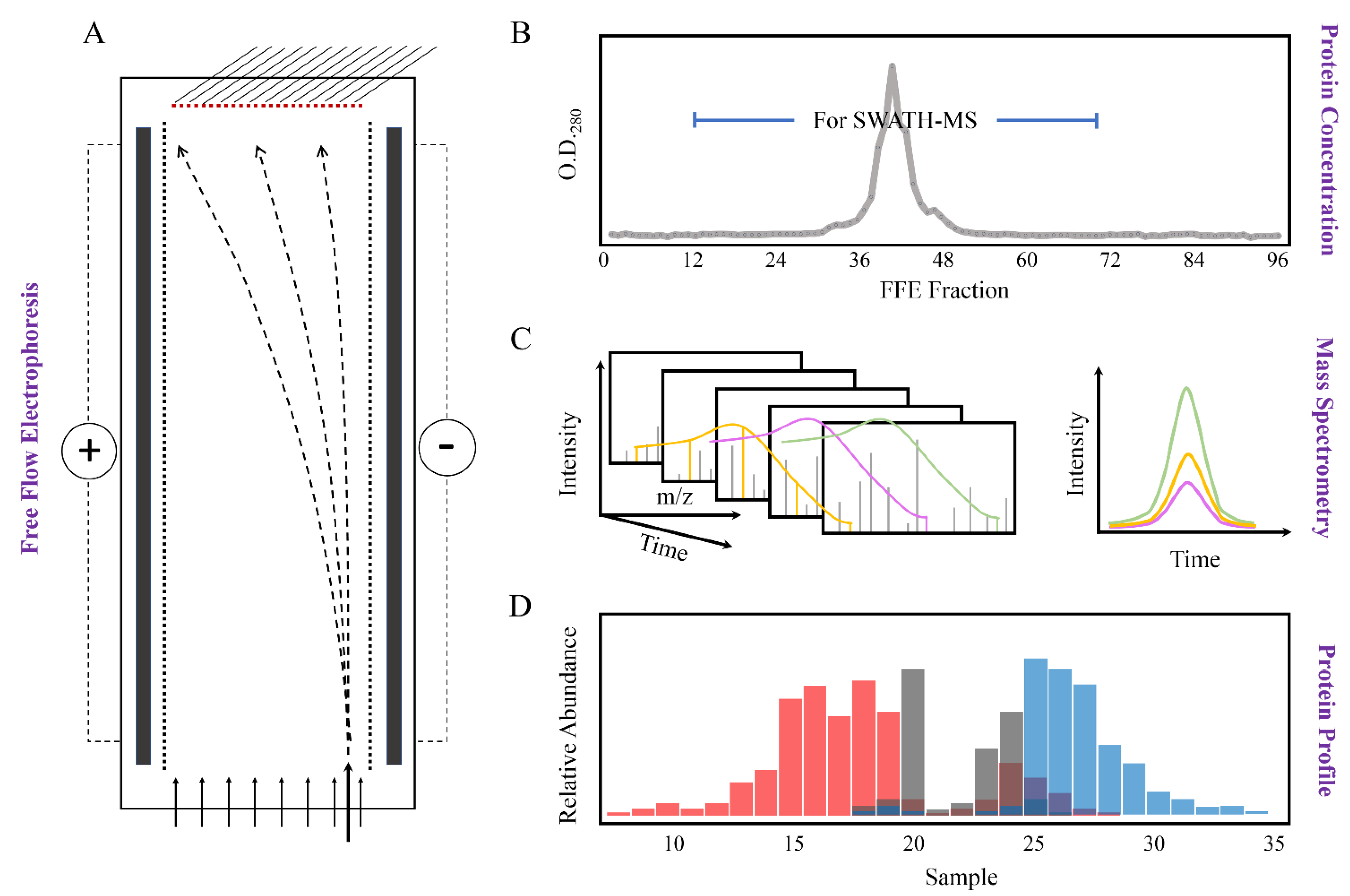
4.3. Free Flow Electrophoresis (FFE)
4.4. TCA Precipitation and Trypsin Digestion of Membrane Proteins
4.5. Information-Dependent Acquisition (IDA)
4.6. Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS)
4.7. Ion Library Generation
4.8. Peptide and Protein Quantification
4.9. Trans Membrane Domain, Functional Annotation and Subcellular Localisation
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maron, L. Divide and conquer: Giving organelle context to genomic data. Plant J. 2019, 100, 436–437. [Google Scholar] [CrossRef] [PubMed]
- Sweetlove, L.J.; Fernie, A.R. The spatial organization of metabolism within the plant cell. Annu. Rev. Plant Biol. 2013, 64, 723–746. [Google Scholar] [CrossRef] [PubMed]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Casares, D.; Escriba, P.V.; Rossello, C.A. Membrane lipid composition: Effect on membrane and organelle structure, function and compartmentalization and therapeutic avenues. Int. J. Mol. Sci. 2019, 20, 2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunkley, T.P.J.; Hester, S.; Shadforth, I.P.; Runions, J.; Weimar, T.; Hanton, S.L.; Griffin, J.L.; Bessant, C.; Brandizzi, F.; Hawes, C.; et al. Mapping the Arabidopsis organelle proteome. Proc. Natl. Acad. Sci. USA 2006, 103, 6518. [Google Scholar] [CrossRef] [Green Version]
- Geladaki, A.; Kočevar Britovšek, N.; Breckels, L.M.; Smith, T.S.; Vennard, O.L.; Mulvey, C.M.; Crook, O.M.; Gatto, L.; Lilley, K.S. Combining LOPIT with differential ultracentrifugation for high-resolution spatial proteomics. Nat. Commun. 2019, 10, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman, M.C.; Brown, P.R. Free Flow Electrophoresis. Anal. Chem. 1994, 66, 86–94. [Google Scholar] [CrossRef]
- Burry, R.W.; Wood, J.G. Contributions of lipids and proteins to the surface charge of membranes. An electron microscopy study with cationized and anionized ferritin. J. Cell Biol. 1979, 82, 726–741. [Google Scholar] [CrossRef] [Green Version]
- Harms, E.; Kern, H.; Schneider, J.A. Human lysosomes can be purified from diploid skin fibroblasts by free-flow electrophoresis. Proc. Natl. Acad. Sci. USA 1980, 77, 6139. [Google Scholar] [CrossRef] [Green Version]
- Barth, F.; Gruetter, M.G.; Kessler, R.; Manz, H.-J. The use of free flow electrophoresis in the purification of recombinant human tissue plasminogen activator expressed in yeast. Electrophoresis 1986, 7, 372–375. [Google Scholar] [CrossRef]
- Marsh, M.; Schmid, S.; Kern, H.; Harms, E.; Male, P.; Mellman, I.; Helenius, A. Rapid analytical and preparative isolation of functional endosomes by free flow electrophoresis. J. Cell Biol. 1987, 104, 875–886. [Google Scholar] [CrossRef] [Green Version]
- Froshauer, S.; Kartenbeck, J.; Helenius, A. Alphavirus RNA replicase is located on the cytoplasmic surface of endosomes and lysosomes. J. Cell Biol. 1988, 107, 2075–2086. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, G.A.; Spiegelberg, A.; Butkowskyj-Walkiw, T.; Bögl, K.W. Assessment of electrophoretic mobility changes of human erythrocytes by free flow electrophoresis after in vitro and in vivo irradiation. Int. J. Radiat. Appl. Instrum. Part B. Nucl. Med. Biol. 1990, 17, 377–380. [Google Scholar] [CrossRef]
- Van Der Sluijs, P.; Hull, M.; Zahraoui, A.; Tavitian, A.; Goud, B.; Mellman, I. The small GTP-binding protein rab4 is associated with early endosomes. Proc. Natl. Acad. Sci. USA 1991, 88, 6313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morré, D.J.; Morré, D.M.; Van Alstine, J.M. Separation of endosomes by aqueous two-phase partition and free-flow electrophoresis. J. Chromatogr. B Biomed. Sci. Appl. 1998, 711, 203–215. [Google Scholar] [CrossRef]
- Kronenberger, P.; Schober, D.; Prchla, E.; Ofori-Anyinam, O.; Vrijsen, R.; Rombaut, B.; Blaas, D.; Fuchs, R.; Boeyé, A. Uptake of poliovirus into the endosomal system of HeLa cells. Arch. Virol. 1998, 143, 1417–1424. [Google Scholar] [CrossRef]
- Völkl, A.; Mohr, H.; Fahimi, H.D. Peroxisome subpopulations of the rat liver: Isolation by immune Free Flow Electrophoresis. J. Histochem. Cytochem. 1999, 47, 1111–1117. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Rhodus, N.L.; Griffin, R.J.; Carlis, J.V.; Griffin, T.J. A catalogue of human saliva proteins identified by Free Flow Electrophoresis-based peptide separation and tandem mass spectrometry. Mol. Cell Proteom. 2005, 4, 1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wase, N.V.; Ow, S.Y.; Salim, M.; Nissum, M.; Whalley, M. A Free Flow Electrophoresis separation strategy for segregation of high abundant phycobilisomes from cyanobacterium Nostoc punctiforme PCC 73102. J. Proteom. Bioinform. 2012, 5. [Google Scholar] [CrossRef] [Green Version]
- Murillo, A.; Vera-Estrella, R.; Barkla, B.J.; Méndez, E.; Arias, C.F. Identification of host cell factors associated with astrovirus replication in Caco-2 Cells. J. Virol. 2015, 89, 10359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkla, B.J.; Vera-Estrella, R.; Hernandez-Coronado, M.; Pantoja, O. Quantitative proteomics of the tonoplast reveals a role for glycolytic enzymes in salt tolerance. Plant Cell 2009, 21, 4044–4058. [Google Scholar] [CrossRef] [Green Version]
- Eubel, H.; Meyer, E.H.; Taylor, N.L.; Bussell, J.D.; O’Toole, N.; Heazlewood, J.L.; Castleden, I.; Small, I.D.; Smith, S.M.; Millar, A.H. Novel proteins, putative membrane transporters, and an integrated metabolic network are revealed by quantitative proteomic analysis of Arabidopsis cell culture peroxisomes. Plant Physiol. 2008, 148, 1809–1829. [Google Scholar] [CrossRef] [Green Version]
- Eubel, H.; Lee, C.P.; Kuo, J.; Meyer, E.H.; Taylor, N.L.; Millar, A.H. TECHNICAL ADVANCE: Free-flow electrophoresis for purification of plant mitochondria by surface charge. Plant J. 2007, 52, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Parsons, H.T.; Heazlewood, J.L. Beyond the Western front: Targeted proteomics and organelle abundance profiling. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichacker, L.A.; Weber, G.; Sukop-Köppel, U.; Wildgruber, R. Free Flow Electrophoresis for separation of native membrane protein complexes. In Proteomic Profiling: Methods and Protocols; Posch, A., Ed.; Springer: New York, NY, USA, 2015; pp. 415–425. [Google Scholar] [CrossRef]
- De Michele, R.; McFarlane, H.E.; Parsons, H.T.; Meents, M.J.; Lao, J.; Gonzalez Fernandez-Nino, S.M.; Petzold, C.J.; Frommer, W.B.; Samuels, A.L.; Heazlewood, J.L. Free-Flow Electrophoresis of plasma membrane vesicles enriched by two-phase partitioning enhances the quality of the proteome from Arabidopsis seedlings. J. Proteome Res. 2016, 15, 900–913. [Google Scholar] [CrossRef] [Green Version]
- Parsons, H.T.; Stevens, T.J.; McFarlane, H.E.; Vidal-Melgosa, S.; Griss, J.; Lawrence, N.; Butler, R.; Sousa, M.M.L.; Salemi, M.; Willats, W.G.T.; et al. Separating Golgi proteins from cis to trans reveals underlying properties of cisternal localization. Plant Cell 2019, 31, 2010–2034. [Google Scholar] [CrossRef] [Green Version]
- Guan, Q.; Kong, W.; Zhu, D.; Zhu, W.; Dufresne, C.; Tian, J.; Chen, S. Comparative proteomics of Mesembryanthemum crystallinum guard cells and mesophyll cells in transition from C3 to CAM. J. Proteom. 2021, 231, 104019. [Google Scholar] [CrossRef]
- Barkla, B.J.; Garibay-Hernandez, A.; Melzer, M.; Rupasinghe, T.W.T.; Roessner, U. Single cell-type analysis of cellular lipid remodelling in response to salinity in the epidermal bladder cells of the model halophyte Mesembryanthemum crystallinum. Plant Cell Environ. 2018, 41, 2390–2403. [Google Scholar] [CrossRef] [PubMed]
- Klink, R.; Haschke, H.-P.; Kramer, D.; Lüttge, U. Membrane Particles, Proteins and ATPase Activity of Tonoplast Vesicles of Mesembryanthemum crystallinum in the C-3 and CAM State*. Bot. Acta 1990, 103, 24–31. [Google Scholar] [CrossRef]
- Barkla, B.J.; Zingarelli, L.; Blumwald, E.; Smith, J.A.C. Tonoplast Na+/H+ antiport activity and tts energization by the bacuolar H+-ATPase in the halophytic plant Mesembryanthemum crystallinum. Plant Physiol. 1995, 109, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, O.T.; Gillet, L.C.; Collins, B.C.; Navarro, P.; Rosenberger, G.; Wolski, W.E.; Lam, H.; Amodei, D.; Mallick, P.; MacLean, B.; et al. Building high-quality assay libraries for targeted analysis of SWATH MS data. Nat. Protoc. 2015, 10, 426–441. [Google Scholar] [CrossRef]
- Tusnády, G.E.; Simon, I. The HMMTOP transmembrane topology prediction server. Bioinformatics 2001, 17, 849–850. [Google Scholar] [CrossRef]
- Sonnhammer, E.L.L.; Heijne, G.v.; Krogh, A. A hidden markov model for predicting transmembrane helices in protein sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1998, 6, 175–182. [Google Scholar] [PubMed]
- Bagos, P.G.; Liakopoulos, T.D.; Spyropoulos, I.C.; Hamodrakas, S.J. A Hidden Markov Model method, capable of predicting and discriminating β-barrel outer membrane proteins. BMC Bioinform. 2004, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Ou, Y.Y.; Gromiha, M.M.; Chen, S.A.; Suwa, M. TMBETADISC-RBF: Discrimination of beta-barrel membrane proteins using RBF networks and PSSM profiles. Comput. Biol. Chem. 2008, 32, 227–231. [Google Scholar] [CrossRef]
- Hooper, C.M.; Castleden, I.R.; Tanz, S.K.; Aryamanesh, N.; Millar, A.H. SUBA4: The interactive data analysis centre for Arabidopsis subcellular protein locations. Nucleic Acids Res. 2017, 45, D1064–D1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, C.M.; Tanz, S.K.; Castleden, I.R.; Vacher, M.A.; Small, I.D.; Millar, A.H. SUBAcon: A consensus algorithm for unifying the subcellular localization data of the Arabidopsis proteome. Bioinformatics 2014, 30, 3356–3364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrie, C.; Murcha, M.W.; Kuehn, K.; Duncan, O.; Barthet, M.; Smith, P.M.; Eubel, H.; Meyer, E.; Day, D.A.; Millar, A.H.; et al. Type II NAD(P)H dehydrogenases are targeted to mitochondria and chloroplasts or peroxisomes in Arabidopsis thaliana. FEBS Lett. 2008, 582, 3073–3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapken, W.; Murphy, A.S. Membrane nanodomains in plants: Capturing form, function, and movement. J. Exp. Bot. 2015, 66, 1573–1586. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Q.S.; Barkla, B.J.; Vera-Estrella, R.; Zhu, J.K.; Schumaker, K.S. Na+/H+ exchange activity in the plasma membrane of Arabidopsis. Plant Physiol. 2003, 132, 1041–1052. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Li, Z.; Wei, J.; Zhao, Z.; Sun, D.; Cui, S. A Na+/Ca2+ exchanger-like protein (AtNCL) involved in salt stress in Arabidopsis. J. Biol. Chem. 2012, 287, 44062–44070. [Google Scholar] [CrossRef] [Green Version]
- Loqué, D.; Yuan, L.; Kojima, S.; Gojon, A.; Wirth, J.; Gazzarrini, S.; Ishiyama, K.; Takahashi, H.; Von Wirén, N. Additive contribution of AMT1;1 and AMT1;3 to high-affinity ammonium uptake across the plasma membrane of nitrogen-deficient Arabidopsis roots. Plant J. 2006, 48, 522–534. [Google Scholar] [CrossRef]
- Kang, J.; Hwang, J.-U.; Lee, M.; Kim, Y.-Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-type ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.; Geisler, M.; Suh, S.J.; Kolukisaoglu, H.Ü.; Azevedo, L.; Plaza, S.; Curtis, M.D.; Richter, A.; Weder, B.; Schulz, B.; et al. Disruption of AtMRP4, a guard cell plasma membrane ABCC-type ABC transporter, leads to deregulation of stomatal opening and increased drought susceptibility. Plant J. 2004, 39, 219–236. [Google Scholar] [CrossRef]
- Li, J.; Wang, D. Cloning and in vitro expression of the cDNA encoding a putative nucleoside transporter from Arabidopsis thaliana. Plant Sci. 2000, 157, 23–32. [Google Scholar] [CrossRef]
- Bernard, C.; Traub, M.; Kunz, H.H.; Hach, S.; Trentmann, O.; Mohlmann, T. Equilibrative nucleoside transporter 1 (ENT1) is critical for pollen germination and vegetative growth in Arabidopsis. J. Exp. Bot. 2011, 62, 4627–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Zhang, G.; Gonzales, N.; Guo, Y.; Hu, H.; Park, S.; Zhao, J. Ca2+-regulated and diurnal rhythm-regulated Na+/Ca2+ exchanger AtNCL affects flowering time and auxin signalling in Arabidopsis. Plant Cell Environ. 2016, 39, 377–392. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Sancho, J.; Vanneste, S.; Lee, E.; McFarlane, H.E.; Esteban Del Valle, A.; Valpuesta, V.; Friml, J.; Botella, M.A.; Rosado, A. The Arabidopsis synaptotagmin1 is enriched in endoplasmic reticulum-plasma membrane contact sites and confers cellular resistance to mechanical stresses. Plant Physiol. 2015, 168, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, T.; Takata, N.; Uemura, M.; Kawamura, Y. Arabidopsis synaptotagmin SYT1, a type I signal-anchor protein, requires tandem C2 domains for delivery to the plasma membrane. J. Biol. Chem. 2010, 285, 23165–23176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; DeBono, A.; Durrett, T.P.; et al. Acyl-Lipid Metabolism. Arab. Book 2013, e0161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heazlewood, J.L.; Tonti-Filippini, J.; Verboom, R.E.; Millar, A.H. Combining experimental and predicted datasets for determination of the subcellular location of proteins in Arabidopsis. Plant Physiol. 2005, 139, 598–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krebs, M.; Beyhl, D.; Gorlich, E.; Al-Rasheid, K.A.; Marten, I.; Stierhof, Y.D.; Hedrich, R.; Schumacher, K. Arabidopsis V-ATPase activity at the tonoplast is required for efficient nutrient storage but not for sodium accumulation. Proc. Natl. Acad. Sci. USA 2010, 107, 3251–3256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, H.T.; Christiansen, K.; Knierim, B.; Carroll, A.; Ito, J.; Batth, T.S.; Smith-Moritz, A.M.; Morrison, S.; McInerney, P.; Hadi, M.Z.; et al. Isolation and proteomic characterization of the Arabidopsis Golgi defines functional and novel components involved in plant cell wall biosynthesis. Plant Physiol. 2012, 159, 12–26. [Google Scholar] [CrossRef] [Green Version]
- Dettmer, J.; Schubert, D.; Calvo-Weimar, O.; Stierhof, Y.D.; Schmidt, R.; Schumacher, K. Essential role of the V-ATPase in male gametophyte development. Plant J. 2005, 41, 117–124. [Google Scholar] [CrossRef]
- Dettmer, J.; Hong-Hermesdorf, A.; Stierhof, Y.D.; Schumacher, K. Vacuolar H+-ATPase activity is required for endocytic and secretory trafficking in Arabidopsis. Plant Cell 2006, 18, 715–730. [Google Scholar] [CrossRef] [Green Version]
- Strompen, G.; Dettmer, J.; Stierhof, Y.D.; Schumacher, K.; Jurgens, G.; Mayer, U. Arabidopsis vacuolar H+-ATPase subunit E isoform 1 is required for Golgi organization and vacuole function in embryogenesis. Plant J. 2005, 41, 125–132. [Google Scholar] [CrossRef]
- Heard, W.; Sklenář, J.; Tomé, D.F.A.; Robatzek, S.; Jones, A.M.E. Identification of regulatory and cargo proteins of endosomal and secretory pathways in Arabidopsis thaliana by proteomic dissection. Mol. Cell Proteom. 2015, 14, 1796–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Quintero, F.J.; Pardo, J.M.; Zhu, J.-K. The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants. Plant Cell 2002, 14, 465. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.-H.; Lee, S.Y.; Bressan, R.A.; Yun, D.-J.; Bohnert, H.J. Intracellular consequences of SOS1 deficiency during salt stress. J. Exp. Bot. 2010, 61, 1205–1213. [Google Scholar] [CrossRef] [Green Version]
- Undurraga, S.F.; Santos, M.P.; Paez-Valencia, J.; Yang, H.; Hepler, P.K.; Facanha, A.R.; Hirschi, K.D.; Gaxiola, R.A. Arabidopsis sodium dependent and independent phenotypes triggered by H+-PPase up-regulation are SOS1 dependent. Plant Sci. 2012, 183, 96–105. [Google Scholar] [CrossRef]
- Nühse, T.S.; Stensballe, A.; Jensen, O.N.; Peck, S.C. Phosphoproteomics of the Arabidopsis plasma membrane and a new phosphorylation site database. Plant Cell 2004, 16, 2394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaquinod, M.; Villiers, F.; Kieffer-Jaquinod, S.; Hugouvieux, V.; Bruley, C.; Garin, J.; Bourguignon, J. A Proteomics dissection of Arabidopsis thaliana vacuoles isolated from cell culture. Mol. Cell Proteom. 2007, 6, 394. [Google Scholar] [CrossRef] [Green Version]
- Whiteman, S.A.; Serazetdinova, L.; Jones, A.M.; Sanders, D.; Rathjen, J.; Peck, S.C.; Maathuis, F.J. Identification of novel proteins and phosphorylation sites in a tonoplast enriched membrane fraction of Arabidopsis thaliana. Proteomics 2008, 8, 3536–3547. [Google Scholar] [CrossRef]
- Ping, H.A.; Kraft, L.M.; Chen, W.; Nilles, A.E.; Lackner, L.L. Num1 anchors mitochondria to the plasma membrane via two domains with different lipid binding specificities. J. Cell Biol. 2016, 213, 513–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staehelin, L.A. Chloroplast structure and supramolecular organization of photosynthetic membranes. In Photosynthesis III: Photosynthetic Membranes and Light Harvesting Systems; Staehelin, L.A., Arntzen, C.J., Eds.; Springer: Berlin/Heidelberg, Germany, 1986. [Google Scholar]
- Barber, J. Membrane surface charges and potentials in relation to photosynthesis. Biochim. Biophys. Acta BBA Rev. Bioenergy 1980, 594, 253–308. [Google Scholar] [CrossRef]
- Hanke, G.; Mulo, P. Plant type ferredoxins and ferredoxin-dependent metabolism. Plant Cell Environ. 2013, 36, 1071–1084. [Google Scholar] [CrossRef]
- Kubota-Kawai, H.; Mutoh, R.; Shinmura, K.; Setif, P.; Nowaczyk, M.M.; Rogner, M.; Ikegami, T.; Tanaka, H.; Kurisu, G. X-ray structure of an asymmetrical trimeric ferredoxin-photosystem I complex. Nat. Plants 2018, 4, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Joyard, J.; Ferro, M.; Masselon, C.; Seigneurin-Berny, D.; Salvi, D.; Garin, J.; Rolland, N. Chloroplast proteomics highlights the subcellular compartmentation of lipid metabolism. Prog. Lipid Res. 2010, 49, 128–158. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Kondo, M.; Fukuda, H.; Nishimura, M.; Ohta, H. Galactolipid synthesis in chloroplast inner envelope is essential for proper thylakoid biogenesis, photosynthesis, and embryogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 17216–17221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li-Beisson, Y.; Beisson, F.; Riekhof, W. Metabolism of acyl-lipids in Chlamydomonas reinhardtii. Plant J. 2015, 82, 504–522. [Google Scholar] [CrossRef]
- Kobayashi, K. Role of membrane glycerolipids in photosynthesis, thylakoid biogenesis and chloroplast development. J. Plant Res. 2016, 129, 565–580. [Google Scholar] [CrossRef] [Green Version]
- Hölzl, G.; Dörmann, P. Chloroplast lipids and their biosynthesis. Annu. Rev. Plant Biol. 2019, 70, 51–81. [Google Scholar] [CrossRef]
- Andersson, M.X.; Dörmann, P. Chloroplast Membrane Lipid Biosynthesis and Transport. In The Chloroplast Plant Cell Monographs; Sandelius, A.S., Aronsson, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 13. [Google Scholar]
- Andersson, M.X.; Kjellberg, J.M.; Sandelius, A.S. Chloroplast biogenesis. Regulation of lipid transport to the thylakoid in chloroplasts isolated from expanding and fully expanded leaves of pea. Plant Physiol. 2001, 127, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Ohnishi, M.; Fukao, Y.; Okazaki, Y.; Fujiwara, M.; Song, C.; Nakanishi, Y.; Saito, K.; Shimmen, T.; Suzaki, T.; et al. Studies on vacuolar membrane microdomains isolated from Arabidopsis suspension-cultured cells: Local distribution of vacuolar membrane proteins. Plant Cell Physiol. 2013, 54, 1571–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, S.K.; Walters, B.T.; Clouse, S.D.; Goshe, M.B. An efficient organic solvent based extraction method for the proteomic analysis of Arabidopsis plasma membranes. J. Proteome Res. 2009, 8, 2752–2767. [Google Scholar] [CrossRef] [PubMed]
- Browse, J.; Warwick, N.; Somerville, C.R.; Slack, C.R. Fluxes through the prokaryotic and eukaryotic pathways of lipid synthesis in the ‘16:3′ plant Arabidopsis thaliana. Biochem. J. 1986, 235, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Zargar, S.M.; Kurata, R.; Inaba, S.; Oikawa, A.; Fukui, R.; Ogata, Y.; Agrawal, G.K.; Rakwal, R.; Fukao, Y. Quantitative proteomics of Arabidopsis shoot microsomal proteins reveals a cross-talk between excess zinc and iron deficiency. Proteomics 2015, 15, 1196–1201. [Google Scholar] [CrossRef] [PubMed]
- Benschop, J.J.; Mohammed, S.; O’Flaherty, M.; Heck, A.J.; Slijper, M.; Menke, F.L. Quantitative phosphoproteomics of early elicitor signaling in Arabidopsis. Mol. Cell Proteom. 2007, 6, 1198–1214. [Google Scholar] [CrossRef] [Green Version]
- Kunst, L.; Samuels, A.L. Biosynthesis and secretion of plant cuticular wax. Prog. Lipid Res. 2003, 42, 51–80. [Google Scholar] [CrossRef]
- Joubès, J.; Raffaele, S.; Bourdenx, B.; Garcia, C.; Laroche-Traineau, J.; Moreau, P.; Domergue, F.; Lessire, R. The VLCFA elongase gene family in Arabidopsis thaliana: Phylogenetic analysis, 3D modelling and expression profiling. Plant Mol. Biol. 2008, 67, 547–566. [Google Scholar] [CrossRef]
- Okazaki, K.; Miyagishima, S.Y.; Wada, H. Phosphatidylinositol 4-phosphate negatively regulates chloroplast division in Arabidopsis. Plant Cell 2015, 27, 663–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.H.; Shilov, I.V.; Seymour, S.L. Nonlinear fitting method for determining local false discovery rates from decoy database searches. J. Proteome Res. 2008, 7, 3661–3667. [Google Scholar] [CrossRef] [PubMed]
- Shilov, I.V.; Seymour, S.L.; Patel, A.A.; Loboda, A.; Tang, W.H.; Keating, S.P.; Hunter, C.L.; Nuwaysir, L.M.; Schaeffer, D.A. The Paragon Algorithm, a next generation search engine that uses sequence temperature balues and feature probabilities to identify peptides from tandem mass spectra. Mol. Cell Proteomics 2007, 6, 1638–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, W.C.; Cushman, J.C. Caryophyllales genomes provide insights into the evolution of Crassulacean acid metabolism (CAM) and halophytism. In Proceedings of the 26th Plant and Animal Genome Conference, San Diego, CA, USA, 12–17 January 2018. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Uniprot Recommended Name (Gene) b | Location c | Digital Western FFE Profiles d |
|---|---|---|
 | ||
| V-type proton ATPase subunit a3 (VHA-a3) | TP |  |
| aquaporin TIP1-1/gamma-TIP (TIP1-1) | TP |  |
| very-long-chain 3-oxoacyl-CoA reductase 1 (KCR1) | ER |  |
| cytochrome b5 isoform B (CYTB5-B) | ER |  |
| polygalacturonate 4-alpha-galacturonosyltransferase (GAUT1) | GA |  |
| galacturonosyltransferase 9 (GAUT9) | GA |  |
| translocase of chloroplast 110 (TIC110) | Chl |  |
| chlorophyll a-b binding protein 1 (LHCB1.3) | Chl |  |
| mitochondrial outer membrane protein porin 2 (VDAC2) | MT |  |
| prohibitin-3 (PHB3) | MT |  |
| ATPase 2, plasma membrane-type (AHA2) | PM |  |
| MipA (mipA) | PM |  |
| Uniprot Recommended Name (Gene) b | Digital Western FFE Profiles c |
|---|---|
 | |
| V-type proton ATPase subunit A (VHA-A) |  |
| V-type proton ATPase subunit B2 (VHA-B2) |  |
| V-type proton ATPase subunit C (VHA-C) |  |
| V-type proton ATPase subunit D (VHA-D) |  |
| V-type proton ATPase subunit E3 (VHA-E3) |  |
| V-type proton ATPase subunit F (VHA-F) |  |
| V-type proton ATPase subunit G1 (VHA-G1) |  |
| V-type proton ATPase subunit H (VHA-H) |  |
| V-type proton ATPase subunit a3 (VHA-a3) |  |
| V-type proton ATPase subunit c2 (VHA-c2) |  |
| V-type proton ATPase subunit d2 (VHA-d2) |  |
| pyrophosphate-energized vacuolar membrane proton pump 1 (AVP1) |  |
| aquaporin TIP2-1/Delta-TIP (TIP2-1) |  |
| aquaporin TIP1-1/gamma-TIP (TIP1-1) |  |
| two-pore calcium channel protein 1 (TPC1) |  |
| chloride channel protein CLC-b (CLC-B) |  |
| chloride channel protein CLC-c (CLC-C) |  |
| Uniprot Recommended Name (Gene) b | Location c | Digital Western FFE Profiles d |
|---|---|---|
 | ||
| calreticulin-1 (CRT1) | ER |  |
| SEC12-like protein 2 (STL2P) | ER |  |
| protein glycosyltransferase subunit 1B (OST1B) | ER |  |
| translocon-associated protein subunit alpha (At2g21160) | ER |  |
| heat shock 70 kDa protein BIP2 (BIP2) | ER |  |
| ADP-ribosylation factor 1 (ARF1) | ER-GA |  |
| coatomer subunit alpha-1 (At1g62020) | ER-GA |  |
| coatomer subunit beta-1 (At4g31480) | ER-GA |  |
| coatomer subunit beta’-2 (At1g52360) | ER-GA |  |
| coatomer subunit gamma (At4g34450) | ER-GA |  |
| transport protein Sec23/24-like (At3g07100) | ER-GA |  |
| apyrase 2 (APY2) | GA |  |
| polygalacturonate 4-alpha-galacturonosyltransferase (GAUT1) | GA |  |
| galacturonosyltransferase 9 (GAUT9) | GA |  |
| probable methyltransferase PMT8 (At1g04430) | GA |  |
| AP-1 complex subunit gamma-2 (At1g60070) | TGN |  |
| clathrin heavy chain 1 (CHC1) | TGN |  |
| Uniprot Recommended Name (Gene) b | Location c | Digital Western FFE Profiles d |
|---|---|---|
 | ||
| translocase of chloroplast 55 (TIC55) | IME |  |
| translocase of chloroplast 110 (TIC110) | IME |  |
| translocase of chloroplast 34 (TOC34) | OME |  |
| thylakoid lumenal 16.5 kDa protein (At4g02530) | TLK |  |
| ATP synthase gamma chain 1 (ATPC1) | TLK |  |
| ATP synthase subunit delta (ATPD) | TLK |  |
| chlorophyll a-b binding protein 1 (LHCB1.3) | TLK |  |
| chlorophyll a-b binding protein 6 (LHCA1) | TLK |  |
| photosystem I reaction center subunit II-2 (PSAD2) | TLK |  |
| photosystem I reaction center subunit III (PSAF) | TLK |  |
| photosystem I reaction center subunit VI-2 (PSAH2) | TLK |  |
| photosystem I reaction center subunit XI (PSAL) | TLK |  |
| photosystem I reaction center subunit psaK (PSAK) | TLK |  |
| photosystem II stability/assembly factor HCF136 (HCF136) | TLK |  |
| photosystem II 10 kDa polypeptide (PSBR) | TLK |  |
| Uniprot Recommended Name (Gene) b | Location c | Digital Western FFE Profiles d |
|---|---|---|
 | ||
| mitochondrial outer membrane protein porin 1 (VDAC1) | MOM |  |
| mitochondrial outer membrane protein porin 2 (VDAC2) | MOM |  |
| mitochondrial outer membrane protein porin 4 (VDAC4) | MOM |  |
| mitochondrial import receptor subunit TOM9-2 (TOM9-2) | MOM |  |
| prohibitin-2 (PHB2) | MIM |  |
| prohibitin-3 (PHB3) | MIM |  |
| ATP synthase subunit beta-3 (At5g08680) | MIM |  |
| cytochrome b-c1 complex subunit rieske-1 (UCR1-1) | MIM |  |
| cytochrome c oxidase subunit 5b-2 (COX5B-2) | MIM |  |
| mitochondrial-processing peptidase subunit beta (MPPbeta) | MIM |  |
| mitochondrial carnitine/acylcarnitine carrier-like protein (BOU) | MIM |  |
| mitochondrial phosphate carrier protein (MPT3) | MIM |  |
| mitochondrial dicarboxylate/tricarboxylate transporter DTC (DTC) | MIM |  |
| alternative NAD(P)H-ubiquinone oxidoreductase C1 (NDC1) | DL |  |
| Uniprot Recommended Name (Gene) b | Digital Western FFE Profiles c |
|---|---|
 | |
| ATPase 2, plasma membrane-type (AHA2) |  |
| ATPase 4, plasma membrane-type (AHA4) |  |
| ATPase 5, plasma membrane-type (AHA5) |  |
| MipA (mipA) |  |
| MipD (mipD) |  |
| MipH (mipH) |  |
| aquaporin pip2;5 (PIP2-5) |  |
| aquaporin pip2;8 (PIP2-8) |  |
| CSC1-like protein ERD4 (ERD4) |  |
| sucrose transport protein SUC2 (SUC2) |  |
| polyol transporter 5 (PLT5) |  |
| fasciclin-like arabinogalactan protein 2 (FLA2) |  |
| fasciclin-like arabinogalactan protein 6 (FLA6) |  |
| early nodulin-like protein 2 (At4g27520) |  |
| Uniprot Recommended Name (Gene) b | Digital Western FFE Profiles c |
|---|---|
 | |
| sodium/hydrogen exchanger 7 (NHX7) |  |
| sodium/calcium exchanger (NCL) |  |
| ammonium transporter 1 member 1 (AMT1-1) |  |
| sulfite exporter TauE/SafE family protein 4 (TauE) |  |
| ABC transporter G family member 40 (ABCG40) |  |
| ABC transporter C family member 4 (ABCC4) |  |
| equilibrative nucleotide transporter 1 (ENT1) |  |
| Uniprot Recommended Name (Gene) b | Lipid Metabolism Pathway c | Location d | Digital Western FFE Profiles e |
|---|---|---|---|
 | |||
| sterol methyltransferase 1 (SMT1) | sterol synthesis | GA |  |
| glycerol-3-phosphate acyltransferase 4 (GPAT4) | suberin synthesis and transport | GA |  |
| 3-ketoacyl-CoA synthase 10 (FDH) | fatty acid synthesis | GA |  |
| very-long-chain 3-oxoacyl-CoA reductase 1 (KCR1) | fatty acid elongation and wax biosynthesis | ER |  |
| cinnamate-4-hydroxylase (CYP73A5) | fatty acid elongation and wax biosynthesis | ER |  |
| cytochrome b5 isoform B (CYTB5-B) | fatty acid synthesis | ER |  |
| phospholipid: diacylglycerol acyltransferase (PDAT1) | triacylglycerol biosynthesis | ER |  |
| membrane-associated progesterone-binding protein 3 (MSBP2) | steroid binding | ER |  |
| 7-dehydrocholesterol reductase (DWF5) | sterol synthesis | ER |  |
| very-long-chain enoyl-CoA reductase (ECR) | fatty acid elongation and wax biosynthesis | ER |  |
| synaptotagmin-1 (SYT1) | lipid binding | ER,PM |  |
| mitochondrial acyl carrier protein 2 (ACP) | mitochondrial fatty acid and lipoic acid synthesis | MT |  |
| non-specific phospholipase C4 (NPC4) | eukaryotic galactolipid and sulfolipid synthesis | PM |  |
| PLC-like phosphodiesterases superfamily protein (At5g67130) | lipid signaling | PM |  |
| phospholipase D delta (PLDDELTA) | phospholipid signaling | PM |  |
| non-specific lipid transfer protein GPI-anchored 2 (LTPG2) | fatty acid elongation and wax biosynthesis | PM |  |
| phosphatidylinositol-specific phospholipase C4 (PLC4) | phospholipid signaling | PM |  |
| phosphatidylinositol 3- and 4-kinase family protein (PI4KA1) | phospholipid signaling | PM |  |
| AMP-dependent synthetase and ligase family protein (LACS4) | eukaryotic phospholipid synthesis and editing | PM |  |
| delta(14)-sterol reductase (FK) | sterol synthesis | PM |  |
| 3-ketoacyl-CoA synthase 6 (CUT1) | fatty acid elongation and wax biosynthesis | PM |  |
| ABC-2 type transporter family protein (ABCG15) | lipid transport | PM |  |
| ABC-2 type transporter family protein (ABCG22) | lipid transport | PM |  |
| esterase lipase thioesterase family protein (At1g54570) | fatty acid elongation and wax biosynthesis | Chl |  |
| trigalactosyldiacyl-glycerol 2 (TGD2) | eukaryotic galactolipid and sulfolipid synthesis | Chl |  |
| trigalactosyldiacyl-glycerol 4 (TGD4) | eukaryotic galactolipid and sulfolipid synthesis | Chl |  |
| translocon at the outer envelope membrane of chloroplasts 159 (TOC159) | galactolipid, sulfolipid, and phospholipid synthesis | Chl |  |
| alpha beta-hydrolases superfamily protein (DALL3) | oxylipin metabolism | Chl |  |
| single hybrid motif superfamily protein (BCCP2) | fatty acid synthesis | Chl |  |
| long-chain acyl-CoA synthetase 9 (LACS9) | fatty acid synthesis galactolipid, sulfolipid, and phospholipid synthesis | Chl |  |
| fatty acid desaturase 6 (FAD6) | fatty acid desaturation galactolipid, sulfolipid, and phospholipid synthesis | Chl |  |
| fatty acid desaturase 8 (FAD8) | fatty acid desaturation galactolipid, sulfolipid, and phospholipid synthesis | Chl |  |
| phospholipid glycerol acyltransferase family protein (ATS1/GPAT) | galactolipid, sulfolipid, and phospholipid synthesis | Chl |  |
| AMP-dependent synthetase and ligase family protein (AAE16) | fatty acid metabolism | Chl |  |
| acetyl CoA carboxylase carboxyltransferase alpha subunit (CAC3) | fatty acid synthesis | Chl |  |
| Uniprot Recommended Name b | Digital Western FFE Profiles c |
|---|---|
 | |
| ferredoxin-1 (FD1) |  |
| ATP synthase gamma chain 1 (ATPC1) |  |
| ATP synthase subunit delta (ATPD) |  |
| photosystem I reaction center subunit VI-2 (PSAH2) |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Q.; Liu, L.; Yim, W.C.; Cushman, J.C.; Barkla, B.J. Membrane Profiling by Free Flow Electrophoresis and SWATH-MS to Characterize Subcellular Compartment Proteomes in Mesembryanthemum crystallinum. Int. J. Mol. Sci. 2021, 22, 5020. https://doi.org/10.3390/ijms22095020
Guo Q, Liu L, Yim WC, Cushman JC, Barkla BJ. Membrane Profiling by Free Flow Electrophoresis and SWATH-MS to Characterize Subcellular Compartment Proteomes in Mesembryanthemum crystallinum. International Journal of Molecular Sciences. 2021; 22(9):5020. https://doi.org/10.3390/ijms22095020
Chicago/Turabian StyleGuo, Qi, Lei Liu, Won C. Yim, John C. Cushman, and Bronwyn J. Barkla. 2021. "Membrane Profiling by Free Flow Electrophoresis and SWATH-MS to Characterize Subcellular Compartment Proteomes in Mesembryanthemum crystallinum" International Journal of Molecular Sciences 22, no. 9: 5020. https://doi.org/10.3390/ijms22095020